Novel Approaches to Cancer Therapy
Writer sand Curator: Larry H. Bernstein, MD, FCAP
11.1 Novel Approaches to Cancer Therapy
11.1.1 Electrically-driven modulation of surface-grafted RGD peptides for .. cell adhesion
11.1.2 The metabolic state of cancer stem cells—a target for cancer therapy
11.1.3 Regulation of tissue morphogenesis by endothelial cell-derived signals
11.1.4 Novel approach to bis(indolyl)methanes. De novo synthesis of 1-hydroxyimino-methyl derivatives with anti-cancer properties
11.1.5 Synthesis and Biological Evaluation of New 1,3-Thiazolidine-4-one Derivatives of 2-(4-Isobutylphenyl)propionic Acid molecules
11.1.6 Targeting pyruvate kinase M2 contributes to radiosensitivity of NSCLC cells
11.1.7 The tyrosine kinase inhibitor nilotinib has antineoplastic activity in prostate cancer cells but up-regulates the ERK survival signal—Implications for targeted therapies
11.1.8 PAF and EZH2 Induce Wnt.β-Catenin Signaling Hyperactivation
11.1.9 PAF Makes It EZ(H2) for β-Catenin Transactivation
11.1.10 PI3K.AKT.mTOR pathway as a therapeutic target in ovarian cancer
11.1.11 Endogenous, hyperactive Rac3 controls proliferation of breast cancer cells by a p21-activated kinase-dependent pathway
11.1.12 Curcumin-could-reduce-the-monomer-of-ttr-with-tyr114cys-mutation via autophagy in cell model of familial amyloid polyneuropathy.
11.1.1 Electrically-driven modulation of surface-grafted RGD peptides for .. cell adhesion
Lashkor M1, Rawson FJ, Stephenson-Brown A, Preece JA, Mendes PM.
Chem Commun (Camb). 2014 Dec 21; 50(98):15589-92
http://dx.doi.org/10.1039%2Fc4cc06649a
Reported herein is a switchable surface that relies on electrically-induced conformational changes within surface-grafted arginine–glycine–aspartate (RGD) oligopeptides as the means of modulating cell adhesion
Stimuli-responsive surfaces that are capable of modulating their biological properties in response to an external stimuli, including temperature,1,2 light,3 magnetic field4 and electrical potential,5–9 are of growing interest for a variety of biological and medical applications.10,11 Switchable surfaces that can be controlled on-demand are playing an increasingly important part in the development of highly sensitive biosensors,12–15novel drug delivery systems16–18 and functional microfluidic, bioanalysis, and bioseparation systems.19–22Additionally, dynamic, synthetic surfaces that can control the presentation of regulatory signals to a cell are expected to have a significant impact in the field of tissue engineering and regenerative medicine, and to provide unprecedented opportunities in fundamental studies of cell biology.23,24 The availability of sophisticated and functional switchable surfaces is expected to emulate more complex in vivo like extracellular environments, and provide a powerful means to probe and control the dynamic interactions between the cell and its external environments.
The majority of studies on stimuli-responsive surfaces reported to date either rely25–29 on controlling non-specific interactions (i.e., hydrophobic/hydrophilic and electrostatic) of the biomolecules with the active surface, or have focused30–32 on demonstrating modulation of specific biomolecular interactions using relatively simple biological systems (e.g. biotin–streptavidin) and conditions (i.e. water or buffer solutions). For example, Zareie et al. 30 fabricated a mixed self-assembled monolayer (SAM) on gold comprising oligo(ethylene glycol) (OEG) thiol molecules and shorter disulfides carrying biotin end-groups that regulated the interaction between biotin and streptavidin in water. The OEG thiols were able to switch in response to a change in temperature below and above their lower critical solution temperature (LCST = 37 °C). At 23 °C the structure of the OEG molecules was fully extended hindering the shorter biotin disulfide components. On the contrary, at 45 °C the OEG backbone collapsed, thus allowing the specific interaction between the biotin molecule on the surface and the protein streptavidin in solution. In our previous work,7–9 electrically controlled switching has been applied to regulate the conformational changes of modified positively charged oligolysine peptides tethered to a gold surface, such that biotin moieties incorporated into the oligolysines could be reversibly exposed or concealed on demand, as a function of surface potential. Switchable SAMs used to control biomolecular interactions via an electrical stimulus are particularly appealing because of their fast response times, ease of creating multiple individually addressable switchable regions on the same surface, as well as low-drive voltage and electric fields, which are compatible with biological systems.33 Our previous reported electrically switchable surface was able to control directly the biomolecular interactions between biotin and neutravidin in phosphate buffer saline (PBS) solution.
However, switchable surfaces have been scarcely used, thus far, to control biomolecular interactions on more complex systems such as those involving modulation of cell responsiveness.34–37 Jonkheijm and co-workers35 have reported a cucurbit[8]uril-based SAM system to electrochemically control the release of cells. Charged end groups on SAM surfaces have been exploited to electrically control the early stages of bacterial cell adhesion37 and form patterned surfaces with two independent dynamic functions for inducing cell migration.36 In spite of these efforts, given cellular complexity and diversity, such studies are very limited in number, as are the opportunities to further understand and control the complex interplay of events and interactions occurring within living cells.
Herein, we report on a stimuli-responsive surface that relies on electrically-induced conformational changes within surface-grafted arginine–glycine–aspartate (RGD) oligopeptides as the means of modulating cell adhesion. RGD, which is present in most of the adhesive ECM proteins (e.g. fibronectin, vitronectin, laminin and collagen), is specific for integrin-mediated cell adhesion.38 The RGD modified electrode is used here to dynamically regulate the adhesion of immune macrophage cells. The stimuli-responsive surface is fabricated on a gold surface and comprises a mixed SAM consisting of two components (Fig. 1): (i) an oligopeptide containing a terminal cysteine for attachment to the gold surface, three lysine residues as the main switching unit, and a glycine–arginine–glycine–aspartate–serine (GRGDS) as the recognition motif for cell adhesion –C3K-GRGDS, and (ii) an ethylene glycol-terminated thiol (C11TEG) to space out the oligopeptides. Since the charged backbone of the oligopeptide can be potentially harnessed7–9 to induce its folding on the surface upon an application of an electrical potential, we reasoned that such conformational changes can be employed to selectively expose under open circuit (OC) conditions (bio-active state) or conceal under negative potential (bio-inactive state) the RGD to the cell and dynamically regulate cell adhesion.

rdg-oligopeptide-sam-utilised-for-controlling-specific-cellular-interactions-c4cc06649a
RDG oligopeptide SAM utilised for controlling specific cellular interactions
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4230383/bin/c4cc06649a-f1.jpg
Fig. 1 Schematic of the dynamic RDG oligopeptide SAM utilised for controlling specific cellular interactions. The electrically switchable SAM exposes the RGD peptide and supports cell adhesion under open circuit (OC) conditions (no applied potential), while …
Mixed SAMs of C3K-GRGDS : C11TEG were formed from a solution ratio of 1 : 40 and characterised by X-ray photoelectron spectroscopy (XPS) (Fig. S2, ESI†). XPS analysis confirmed the formation of the C3K-GRGDS:C11TEG mixed monolayer and displayed signals from S, N, C and O. The chemical state of the sulphur atom was probed using the XPS spectra of the S 2p emission (Fig. S2, ESI†). The S 2p spectrum (Fig. S2a, ESI†) consists of two doublet peaks, with one doublet peak at 162.0 eV (S 2p3/2) and 163.2 eV (S 2p1/2), indicating that the sulphur is chemisorbed on the gold surface.39 A second small doublet peak can be observed at 163.8 eV and 165.0 eV, which can be attributed to the S–H bond, indicating a small presence of unbound sulphur. No sulphur peaks above 166 eV were observed, indicating that no oxidised sulphur is present at the surface. The N 1s spectrum (Fig. S2b, ESI†) can be de-convoluted into two peaks, which support the presence of the peptide on the surface. The first peak centred at 400.5 eV is attributed to amino (NH2) and amide (CONH) moieties. The second peak centred at 402.8 eV is ascribed to protonated amino groups.40 Note that no nitrogen peak was observed for pure C11TEG SAMs. The C 1s spectrum (Fig. S2c, ESI†) can be de-convoluted into three peaks, which are attributed to five different binding environments. The peak at 285.0 eV is attributed to C–C bonds,41 while the peak at 286.7 eV corresponds to C 1s of the three binding environments of C–S, C–N and C–O.41 The third and smaller peak (288.6 eV) is assigned to the C 1s photoelectron of the carbonyl moiety, C O.41 The O 1s spectrum (Fig. S2d, ESI†) is de-convoluted into two different peaks, corresponding to two different binding environments, arising from the C–O (533.3 eV) and C O (532.0 eV) bonds.41 From integrating the area of the S 2 p and N 1s peaks and taking into consideration that the C3K-GRGDS oligopeptide consists of 15 N atoms and 1 S atom and C11TEG has no N and 1 S atom only, it was possible to infer that the ratio of C3K-GRGDS:C11TEG on the surface is 1 : 10 ± 2. The presence of C11TEG was utilised not only to ensure sufficient spatial freedom for molecular reorientation of the surface bound oligopeptide, but also to stop non-specific binding to the surface.
The C3K-GRGDS:C11TEG mixed SAMs were shown to support adhesion of immune macrophage cells as determined by cell counting42,43 (Fig. 2). When RAW 264.7 mouse macrophages were cultured on theC3K-GRGDS:C11TEG mixed SAM in supplemented Dulbecco’s Modified Eagle Medium (DMEM), the number of cells adhered to the surface increased with incubation time, reaching 1792 ± 157 cells per mm2after 24 hours. This is in contrast with the weak cell adhesion observed in two control surfaces, pureC11TEG SAMs and clean gold, in which the number of cells that adhere was 60% and 50% lower, respectively, after 24 hours (Fig. 2).

microscopic-images-and-density-of-adhered-cells-on-c3k-grgds-c11teg-mixed-sam-pure-c11teg-sam-and-bare-gold-surfaces
Microscopic images and density of adhered cells on C3K-GRGDS:C11TEG mixed SAM, pure C11TEG SAM and bare gold surfaces
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4230383/bin/c4cc06649a-f2.jpg
Fig. 2 Microscopic images and density of adhered cells on C3K-GRGDS:C11TEG mixed SAM, pure C11TEG SAM and bare gold surfaces that were normalized against the density of cells adherent onto the C3K-GRGDS:C11TEG mixed SAM. The surfaces were cultured in RAW 264.7 mouse macrophage cells under OC conditions for 24 hours.
In order to demonstrate that the C3K-GRGDS:C11TEG mixed SAMs can support or resist cell adhesion on demand, the macrophage cells were cultured on the C3K-GRGDS:C11TEG mixed SAM in DMEM medium under OC conditions and applied negative potential (–0.4 V) for a period of 1 h. Note that DMEM contains a mixture of inorganic salts, amino acids, glucose and vitamins. On application of the applied potential of –0.4 V the number of adherent cells was 70% less compared to the C3K-GRGDS:C11TEGmixed SAMs under OC conditions, Fig. 3. Similar switching efficiencies have been observed in another oligopeptide system using different DMEM solutions.44 These findings suggest that the negative potential induces the conformational changes in the C3K moiety of C3K-GRGDS in the SAM which in turn leads to the RGD moiety being concealed and hence reducing the binding of the cells.

density-of-adhered-cells-on-c3k-grgds-c11teg-c11teg-c6eg-grgds-c11teg-mixed-sams-c4cc06649a-f3
Density of adhered cells on C3K-GRGDS:C11TEG, C11TEG, C6EG-GRGDS:C11TEG mixed SAMs
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4230383/bin/c4cc06649a-f3.jpg
Fig. 3 Density of adhered cells on C3K-GRGDS:C11TEG, C11TEG, C6EG-GRGDS:C11TEG mixed SAMs that were normalized against the density of cells adherent onto the C3K-GRGDS:C11TEG mixed SAM. The surfaces were cultured in RAW 264.7 for 1 h under OC conditions or while applying –0.4 V.
Previous studies have shown that small conformational and orientational changes in proteins and peptides modulate the availability and potency of the active sites for cell surface receptors.45–47 Thus, in a similar manner, small changes in the conformation/orientation of the RGD peptide on the surface induced by application of an electrical potential are able to affect the binding activity of the peptide. Recently, we have conducted detailed theoretical8 and experimental9 studies aimed at understanding the switching mechanism of oligopeptide-based switchable surfaces, that similarly as in the case of the C3K-GRGDS:C11TEG mixed SAMs, use lysine residues to act as the switching unit. These previous studies unraveled that the surface-appended oligolysines undergo conformational changes between fully extended, partially extended and collapsed conformer structures in response to an applied positive potential, open circuit conditions and negative electrical potential, respectively. Thus, these previous findings allow us to propose that when a negative potential is applied to the GRGDS:C11TEG mixed SAM surface, the oligopeptide chain adopts a collapsed conformation on the surface and the RGD binding motif is partially embedded on the C11TEGmatrix, thus showing no bioactivity (“OFF” state).
In order to verify that the changes in adhesion upon application of a negative surface potential occur due to changes in the conformational orientation of the RGD instead of cell repulsion or cell damage due to the presence of an electrical potential, control mixed SAMs were also prepared using C11TEG and a peptide where the 3 lysine residues as the switching unit were replaced by 6 non-switchable ethylene glycol units –C6EG-GRGDS (Fig. S1, ESI†). Fig. 3 demonstrates that cells adhered in similar numbers to the C11TEGand C6EG-GRGDS:C11TEG mixed SAMs under OC conditions and an applied negative potential. These results provide strong evidence that control over cell adhesion using the C3K-GRGDS:C11TEG mixed SAM is due to a conformational behaviour of the lysine-containing oligopeptide that can either expose or conceal the RGD moiety.
Cell viability was checked following application of –0.4 V for 1 h by performing a trypan blue assay. Cells that were dead were stained blue due to a break down in membrane integrity. Incubation of the cells under a negative potential had negligible effect on cell viability, which was greater than 98%. Cyclic voltammetric studies (outlined in detail in the Fig. S3, ESI†) were also performed to demonstrate that no significant faradaic process occur over the potential range studied, and thus ions are not participating in redox reactions and consequently redox chemistry is not being significantly affected by application of the potential used. In agreement with other studies,35,36,48 we conclude that the electrical modulation of the surface neither affected cell viability nor induced any redox process in the medium that could have had an effect on cells.
We then addressed the question of whether the C3K-GRGDS:C11TEG surfaces could be switched between different cell adhesive states (cell-resistant and cell-adhesive states). To begin with, we investigated the switching from a cell-adhesive state to a cell-resistant state, and the possibility to detach the cells from the substrate upon the application of a negative potential. Cells were incubated in the C3K-GRGDS:C11TEGmixed SAMs for 1 h under OC conditions, thereby exposing the RGD moiety and allowing for cell attachment. This step was followed by the application of a potential of –0.4 V for 1 h in order to detach the cells from the surface, by concealing the RGD moieties. Cell counts showed no significant differences between the pre and post application of the –0.4 V, suggesting that the electrostatic force generated by the applied negative electrical potential might not be sufficient to disrupt the RGD–integrin interaction. These results were to a certain extent expected since adherent cells are able to withstand strong detachment forces due to the adhesion being mediated by multiple RGD–integrin bonds in parallel.49
In contrast, a reversal of the switching sequence demonstrated that our surfaces can be dynamically switched from a non-adhesive to cell-adhesive state. Cells were incubated in the C3K-GRGDS:C11TEG mixed SAMs for 1 h while holding the potential at –0.4 V for 1 h making the RGD peptide inaccessible for recognition by the corresponding integrin. As above, the number of adherent cells when a negative potential of –0.4 V was applied was 70% of the number that adhered to the C3K-GRGDS:C11TEG mixed SAMs under OC conditions, Fig. 4. The potential was then shifted to open circuit conditions for 1 h on those exposed to a potential of –0.4 V, which resulted in a significant increase in the number of cells as a result of the exposure of the RGD moiety to the cells (Fig. 4). These values were similar to those obtained for the samples that were only incubated for 1 hour under OC conditions (Fig. 4), indicating that the surfaces were highly effective at switching from a non-adhesive to cell-adhesive state.

microscopic-images-and-density-of-adhered-cells-on-c3k-grgds-c11teg-mixed-sams-c4cc06649a-f4
Microscopic images and density of adhered cells on C3K-GRGDS:C11TEG mixed SAMs
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4230383/bin/c4cc06649a-f4.jpg
Fig. 4 Microscopic images and density of adhered cells on C3K-GRGDS:C11TEG mixed SAMs that were incubated with cells for 1 h while applying –0.4 V and subsequently in OC conditions for 1 h. The density was normalized against the density of cells adherent onto C3K-GRGDS:C11TEG mixed SAMs that were incubated with cells in OC conditions for 1 h.
In summary, an electrically switchable surface has been devised and fabricated that is capable of efficiently exposing and concealing the RGD cell adhesion motif and dynamically regulate the adhesion of immune macrophage cells. This study will no doubt be useful in developing more realistic dynamic extracellular matrix models and is certainly applicable in a wide variety of biological and medical applications. For instance, macrophage cell adhesion to surfaces plays a key role in mediating immune response to foreign materials.50 Thus, development of such dynamic in vitro model systems that can control macrophage cell adhesion on demand are likely to provide new opportunities to understand adhesion signaling in macrophages51 and develop effective approaches for prolonging the life-span of implantable medical devices and other biomaterials.52
11.1.2 The metabolic state of cancer stem cells—a target for cancer therapy
Vlashi E1, Pajonk F2.
Free Radic Biol Med. 2015 Feb; 79:264-8
http://dx.doi.org:/10.1016/j.freeradbiomed.2014.10.732
Highlights
- Bulk tumor cell populations rely on aerobic glycolysis.
- Cancer stem cells are in a specific metabolic state.
- Cancer stem cells in breast cancer, glioblastoma, and leukemia rely on oxidative phosphorylation of glucose.
In the 1920s Otto Warburg first described high glucose uptake, aerobic glycolysis, and high lactate production in tumors. Since then high glucose uptake has been utilized in the development of PET imaging for cancer. However, despite a deepened understanding of the molecular underpinnings of glucose metabolism in cancer, this fundamental difference between normal and malignant tissue has yet to be employed in targeted cancer therapy in the clinic. In this review, we highlight attempts in the recent literature to target cancer cell metabolism and elaborate on the challenges and controversies of these strategies in general and in the context of tumor cell heterogeneity in cancer.
11.1.3 Regulation of tissue morphogenesis by endothelial cell-derived signals
Saravana K. Ramasamy, Anjali P. Kusumbe, Ralf H. Adams
Trends Cell Biol Mar 2015; 25(3):148–157
http://dx.doi.org/10.1016/j.tcb.2014.11.007
Highlights
- Endothelial cells lining blood vessels induce organ formation and other morphogenetic processes in the embryo.
- Blood vessels are also an important source of paracrine (angiocrine) signals acting on other cell types in organ regeneration.
- Vascular niches and endothelial cell-derived signals generate microenvironments for stem and progenitor cells.
Endothelial cells (ECs) form an extensive network of blood vessels that has numerous essential functions in the vertebrate body. In addition to their well-established role as a versatile transport network, blood vessels can induce organ formation or direct growth and differentiation processes by providing signals in a paracrine (angiocrine) fashion. Tissue repair also requires the local restoration of vasculature. ECs are emerging as important signaling centers that coordinate regeneration and help to prevent deregulated, disease-promoting processes. Vascular cells are also part of stem cell niches and have key roles in hematopoiesis, bone formation, and neurogenesis. Here, we review these newly identified roles of ECs in the regulation of organ morphogenesis, maintenance, and regeneration.
http://ars.els-cdn.com/content/image/1-s2.0-S0962892414002104-gr1.sml
Figure 1. Role of endothelial cells (ECs) during organogenesis
http://ars.els-cdn.com/content/image/1-s2.0-S0962892414002104-gr2.sml
Figure 2. Endothelial cells (ECs) in lung regeneration
http://ars.els-cdn.com/content/image/1-s2.0-S0962892414002104-gr3.sml
Figure 3. Liver endothelium in regeneration and fibrosis.

Vascular cells have key roles in morphogenesis and regeneration
http://ars.els-cdn.com/content/image/1-s2.0-S0962892414002104-gr4.sml
Figure 4. Functional roles of the bone vasculature
http://ars.els-cdn.com/content/image/1-s2.0-S0962892414002104-gr5.sml
Figure 5. Vascular niche for neurogenesis.
Concluding remarks
The examples provided in this review highlight the important roles of ECs in tissue development, patterning, homeostasis, and regeneration. The endothelium often takes a central position in these processes and there are many reasons why ECs are ideally positioned as the source of important instructive, angiocrine signals. The vascular transport network extends into every organ system and needs to be embedded in those tissues in a certain spacing or pattern, which places ECs in central and, therefore, strategic positions for the regulation of morphogenesis and organ homeostasis.
Given that ECs and other cell types frequently form functional units, such as kidney glomeruli, liver lobules, or lung alveoli, the assembly, differentiation, and function of the different cellular components needs to be tightly coordinated. In addition, because circulating blood cells extensively rely on the vascular conduit system and frequently interact with the endothelium, it is perhaps not surprising that ECs contribute to niche microenvironments. During tissue repair, proliferative cell expansion processes are sometimes temporally separated from cell differentiation and tissue patterning events. The latter has to involve the restoration of a fully functional vascular network so that ECs appear ideally suited as the source of molecular signals that can trigger or suppress processes in the surrounding tissue.
11.1.4 Novel approach to bis(indolyl)methanes. De novo synthesis of 1-hydroxyimino-methyl derivatives with anti-cancer properties
Grasso C, et al.
Eur J Medicinal Chem 01/2015; 93:9-15.
http://dx.doi.org:/10.1016/j.ejmech.2015.01.050
A versatile and broad range approach to previously unknown bis(indolyl)methane oximes based on two consecutive hetero Diels-Alder cycloaddition reactions of electrophilic conjugated nitrosoalkenes with indoles is disclosed. The cytotoxic properties and selectivity of some adducts against several human cancer cell lines pointing to a promising role in the development of anti-tumoural drugs, in particular for leukemia and lymphoma.
Novel approach to bis(indolyl)methanes: De novo synthesis of 1-hydroxyiminomethyl derivatives with anti-cancer properties. Available from:
https://www.researchgate.net/publication/271525370
_Novel_approach_to_bis-28indolyl-29methanes_De_novo_synthesis_of_1-hydroxyiminomethyl_ derivatives_with_anti-cancer_properties [accessed Apr 11, 2015].
The one-pot synthetic strategy to bis(indolyl)methanes is outlined in Scheme 3. The starting a,a 0-dihalogenooximes 3 were efficiently prepared from the respective ketones by known procedures [58,61]. These compounds, in the presence of base, were converted, in situ, into the corresponding transient and reactive nitrosoalkenes 4, which were intercepted bya first molecule of the appropriate indole 5 originating the intermediate indole oximes 6. The initially formed tetrahydroxazines undergo ring-opening to the corresponding oximes, under the driving force of the energy gain on rearomatisation. Subsequent dehydro-halogenation of 6 produces nitrosoalkenes 7 which reacted with a second molecule of indole, producing the target bis(indolyl)methanes 8. The results obtained are summarised in Table 1.
The reaction yields may be considered generally good, taking into account that the synthetic process involves a sequence of reactions. On the other hand, no other products could be obtained, which indicates that the reactions were regioselective. The results have shown also that both alkyl and aryl oximes can be used in the synthesis of bis(indolyl)methanes. Starting from aryl oximes 3aef the expected (E) oximes 9 were obtained as single or major products (Entries 1e11) whereas alkyl oxime 3g reacted with indole to give the (Z)-oxime 10g as the major product (Entries 12e13). The stereochemistry assignment of oximes 9 and 10 was confirmed by analysis of the NOESY spectra of 9d, 9g, 10d and 10g. In the spectra of 10d and 10g, connectivity was observed between the hydroxyl proton and the phenyl protons and the methyl protons, respectively, whereas in the case of 9d and 9g no connectivity was observed. Moreover, oximes 9 and 10 are also characterized by 1H NMR spectra with different features. The chemical shift of the methylenic proton appears at higher value for (E)-oximes 9 (9b: δ 6.81 ppm; 9d: δ = 6.82 ppm; 9g: δ = 6.39 ppm) than for the corresponding (Z) oximes 10 (10b: δ = 5.74 ppm; 10d: δ = 5.77 ppm; 10g: δ = 5.41 ppm).
The synthesis of two isomeric oximes from the reaction of arylnitrosoethylenes with pyrrole and dipyrromethanes has been previously observed [62]. The process was rationalized considering the conjugate addition of the heterocycle to the nitrosoalkene, at the s-cis or s-trans conformation, followed by rearomatization of the pyrrole unit leading to (E)- and (Z)-oxime, respectively. Thus, the synthesis of the BIM oximes via 1,4-conjugate addition of indole to the nitrosoelkene cannot be ruled out.
The use of water as solvent in Diels- Alder reactions has been shown to be advantageous, not only in environmental terms but also inducing critical improvements in reaction times, yields and selectivity [51,63]. We observed that carrying out the synthesis of bis(indolyl)methanes in water using dichloromethane as co-solvent is a valuable alternative to the use of dichloromethane as the only solvent. Generally the yields were better or comparable to those obtained in dichloromethane and reaction time significantly shorter (the reaction time was reduced from 36 h to 3 h). Clearly the efficiency of the reaction, using H2O/CH2Cl2 system, amongst the nitrosoalkenes bearing halogenated aryl substituents increases in the order F > Cl > Br > H the order of electron withdrawing ability and consequently the order of the expected effectiveness for an inverse electron demand Diels-Alder reaction (entries 2, 5, 7 and 9). However, the isolated yields from the reaction carried out in CH2Cl2 do not reflect the expected reactivity, which can be explained considering differences in the efficiency of the purification process.
The cytotoxicity of compounds 9a, 9e and 9d was evaluated in different tumorl cell lines, namely HepG2 (hepatocellular carcinoma), MDA-MB-468 (human breast carcinoma), RAW 264.7 (murine leukemic monocyte macrophages), THP1 (human acute monocytic leukaemia), U937 (human leukaemic monocytic lymphoma) and EL4 cells (murine T-lymphoma). The compounds’ selectivity towards tumoural cells was assessed determining their cytotoxicity with respect to two non-tumoural derived cell lines S17 (murine bone marrow) and N9 cells (murine microglial). Results of the half maximal concentrations (IC50) are shown in Table 2 together with the toxicity of etoposide, a known antitumoural drug. Compound 9e was considerably less cytotoxic on tumor cell lines than the other two compounds, with IC50 values ranging from 35.7 (HepG2) to 124 mM (THP1) and was not selective. Compounds 9a and 9d, however, were considerably cytotoxic to all cells tested, with IC50 values ranging from 1.62 (THP1) to 23.9 mM (RAW) and from 10.7 (MDA) to 34.1 mM (U937), respectively. Compound 9a was particularly active against non-adherent cell lines with IC50 values ranging from 1.62 in THP1 to 1.65 mM in EL4.
Some conclusions regarding structure activity relationships can be redrawn based on the biological evaluation of these bis(indolyl)methanes. There is a dramatic difference in anticancer activitybetweenN-unsubstituted bis(indolyl)methanes 9a and the Nmethyl substituted derivative 9e, the latter characterized by high IC50 values. On the other hand, the significantly lower IC50 values observed for 9a for non-adherent cell lines in comparisonwith the ones obtained for 9d demonstrates that the presence of the bromo substituent leads to higher cytotoxic activity.
The observed high cytotoxicity of compound 9a against THP1, EL4 and U937 cell lines led us to extend the study to BIMs 9c, 9g and 10g (Table 3). Compound 9c, bearing a 4-fluorophenyl substituent, showed moderate anti-cancer activity which reinforces the observation that the 4-bromophenyl group is crucial to ensure low IC50 values. On the other hand, alkyl oximes 9g and 10g were even less cytotoxic against THP1, EL4 and U937 cell lines. None of these compounds were selective towards the tumor cell lines (selectivity index calculated for non-tumour cell line S17). In addition to having displayed higher toxicity towards the nontumor cell lines than all the studied compounds, compound 9a demonstrated the highest selectivity indexes: 9.86-14.2. Further studies using 9a as scaffold in the development of anti-tumoural drugs for leukaemia and lymphoma is worth pursuing since it presents lower IC50 and higher selectivity than etoposide.
Conclusions
The reliable preparation of a variety of unknown BIMs bearing different oxime substituents at the methylene bridge was presented. This strategy, supported on the robust and proved methodology of Diels-Alder cyclo addition reactions of electrophilic nitrosoalkenes with electron rich indoles, may pave the way for the synthesis of a vast library of new compounds.
Table 1 Preparation of bis(indolyl)methane oxime
Scheme 1. Selected biological active bis(indolyl)methanes.
Scheme 2. Common methods for BIMs’ preparation [27e44].
Scheme 3. Synthetic strategy towards BIM oximes.
Synthesis of a new bis(indolyl)methane that inhibits growth and induces apoptosis in human prostate cancer cells
Marrelli M., et al.
Natural product research 08/2013; 27(21).
http://dx.doi.org:/10.1080/14786419.2013.824440
The synthesis and the antiproliferative activity against the human breast MCF-7, SkBr3 and the prostate LNCaP cancer cell lines of a series of bis(indolyl)methane derivatives are reported. The synthesis of new compounds was first accomplished by the reaction of different indoles with trimethoxyacetophenone in the presence of catalytic amounts of hydrochloric acid. A second procedure involving the use of oxalic acid dihydrate [(CO2H)2·2H2O] and N-cetyl-N,N,N-trimethylammonium bromide in water was carried out and led to better yields. Compound 5b significantly reduced LNCaP prostate cancer cell viability in a dose-dependent manner, with an IC50 of 0.64 ± 0.09 μM. To determine whether the growth inhibition was associated with the induction of apoptosis, treated cells were stained using DAPI. LNCaP cells treated with 1 μM of 5b showed the morphological changes characteristic of apoptosis after 24 h of incubation.
11.1.5 Synthesis and Biological Evaluation of New 1,3-Thiazolidine-4-one Derivatives of 2-(4-Isobutylphenyl)propionic Acid molecules
Vasincu IM1, Apotrosoaei M2, Panzariu AT3, Buron F4, Routier S5, Profire L6
Molecules. 2014 Sep 18; 19(9):15005-25
http://dx.doi.org/10.3390/molecules190915005
New thiazolidine-4-one derivatives of 2-(4-isobutylphenyl)propionic acid (ibuprofen) have been synthesized as potential anti-inflammatory drugs. The structure of the new compounds was proved using spectral methods (FR-IR, 1H-NMR, 13C-NMR, MS). The in vitro antioxidant potential of the synthesized compounds was evaluated according to the total antioxidant activity, the DPPH and ABTS radical scavenging assays. Reactive oxygen species (ROS) and free radicals are considered to be involved in many pathological events like diabetes mellitus, neurodegenerative diseases, cancer, infections and more recently, in inflammation. It is known that overproduction of free radicals may initiate and amplify the inflammatory process via upregulation of genes involved in the production of proinflammatory cytokines and adhesion molecules. The chemical modulation of acyl hydrazones of ibuprofen 3a–l through cyclization to the corresponding thiazolidine-4-ones 4a–n led to increased antioxidant potential, as all thiazolidine-4-ones were more active than their parent acyl hydrazones and also ibuprofen. The most active compounds are the thiazolidine-4-ones 4e, m, which showed the highest DPPH radical scavenging ability, their activity being comparable with vitamin E.
In order to improve the anti-inflammatory effect and safety profile of representative NSAIDs, one research strategy is derivatization of the carboxylic acid group with various heterocyclic systems (oxazole, izoxazole, pyrazole, oxadiazole, thiazole, thiadiazole, triazole, etc.) [9,10]. In the past two decades there has been considerable interest in the role of reactive oxygen species (ROS) in inflammation [11]. ROS mediate the oxidative degradation of cellular components and alteration of protease/antiprotease balance with damage to the corresponding tissue. In the early stages of the inflammatory process, ROS exert their actions through activation of nuclear factors, such as NFkB or AP-1, that induce the synthesis of cytokines. In later stages, endothelial cells are activated due to the synergy between free radicals and cytokines, promoting the synthesis of inflammatory mediators and adhesion of molecules. In the last step free radicals react with different cellular components (trypsin, collagen, LDL, DNA, lipids) inducing the death of cells [12,13].
The thiazolidine-4-one moiety is a heterocycle that has received more attention in the last years due its important biological properties [14]. Many effects have been found, including anti-inflammatory and analgesic [15], antitubercular [16], antimicrobial and antifungal [17], antiviral, especially as anti-HIV agents [18], anticancer, antioxidants [19], anticonvulsants [20] and antidiabetic activity [21]. In the present study, some new derivatives of ibuprofen that contain thiazolidine-4-one scaffolds were synthesized in order to obtain compounds with double effect—antioxidant and anti-inflammatory properties. The structures of the compounds were assigned based on their spectral data (FT-IR, 1H-NMR, 13C-NMR, MS) and the compounds were screened for their in vitro antioxidant potential.
The 1,3-thiazolidine-4-one derivatives 4a–m were synthesized in several steps using the method summarized in Scheme 1 and Table 1. First 2-(4-isobutylphenyl)propionic acid (ibuprofen, 1) was reacted with thionyl chloride, followed by treatment with dry ethanol to get 2-(4-isobutylphenyl)propionic acid ethyl ester, which was turned in 2-(4-isobutylphenyl)propionic acid hydrazide (2) by reaction with 66% hydrazine hydrate [22]. The condensation of compound 2 with various aromatic aldehydes allowed the preparation of the corresponding hydrazone derivatives 3a–l in satisfactory yields. Finally, the hydrazone derivatives of ibuprofen upon reaction with mercaptoacetic acid led to the thiazolidine-4-one derivatives 4a–l in moderate to good yields. By reduction of compound 4g in presence of tin chloride and few drops of acetic acid in ethanol, the thiazolidine-4-one 4m was obtained in 90% yield. Acetylation of 4m with acetyl chloride gave thiazolidine-4-one 4n in moderate yield.
In the acyl hydrazone series most of the the tested compounds showed a radical scavenging ability comparable with ibuprofen (Table 4). The most active compounds were 3e and 3f which are about three times and two times more active than their parent compound, respectively. The scavenging ability of the acyl hydrazones was improved by cyclization to the corresponding thiazolidine-4-one derivatives, these compounds all being more active than ibuprofen, except for compound 4j which contains a CF3 group in the metaposition of phenyl ring (Table 5). The most active compounds were 4e and 4m which contain NO2 and NH2 groups in ortho and paraposition of the phenyl ring, respectively. For these compounds the radical scavenging ability (%) was 94.42 ± 0.43 and 94.88 ± 0.57, which means that the compounds are about 23 times more active than ibuprofen (4.15 ± 0.22). The activity of these compounds is comparable with that of vitamin E used as positive control. Important radical scavenging ability was also shown by compound 4b(81.31 ± 0.55), which contains a Cl group in the para position of the phenyl ring, the compound being 20 times more active than ibuprofen.
The acyl hydrazone derivatives showed an antioxidant activity comparable with ibuprofen. The most active compound in this series was 3h, with radical scavenging activity of 13.31 ± 0.81, which means that this compound is three times more active than ibuprofen (4.42 ± 0.18). In the thiazolidine-4-one series the most active compounds were 4b, 4e and 4k, which contain Cl(4), NO2(2) and CN(4), respectively, as substituents on the phenyl ring. These compounds, which showed a scavenging ability of around 50%, are 12 times more active than ibuprofen. In comparison with the corresponding acyl hydrazones 3b, 3e and 3k the thiazolidine-4-ones were 10 times (4b), seven times (4e) and 13 times (3k) more active. The improved antiradical activity of acyl hydrazones by cyclization to form thiazolidine-4-ones was also observed for compounds 3d, 3f and 3g. The most favorable influence was observed for acyl hydrazone 4g, which contains a NO2 in the para position of the phenyl ring. The corresponding thiazolidine-4-one (4g, 37.14 ± 1.10) is 22 times more active than 3g (1.67 ± 0.35). These data strongly support the favorable influence of the thiazolidine-4-one ring on the antioxidant potential of these compounds. The tested compounds were less active than vitamin E.
In this study new heterocyclic compounds that combine the thiazolidine-4-one structure with the arylpropionic acid one have been synthesized. The structure of the new compounds was proved using spectral methods (IR, 1H-NMR, 13C-NMR, MS). The compounds were evaluated for their antioxidant effects using in vitro assays: total antioxidant activity, DPPH and ABTS radical scavenging ability. All thiazolidin-4-one derivatives 4a–n showed improved antioxidant effects in comparison with the corresponding acyl hydrazones 3a–l and ibuprofen, the parent compound. The encouraging preliminary results illustrate the antioxidant potential of the synthesized compounds and motivate our next research focused on their anti-inflammatory effects in chronic and acute inflammation models.
11.1.6 Targeting pyruvate kinase M2 contributes to radiosensitivity of NSCLC cells
Meng MB1, Wang HH2, Guo WH3, Wu ZQ2, Zeng XL2, Zaorsky NG4, et al.
Cancer Lett. 2015 Jan 28; 356(2 Pt B):985-93
http://dx.doi.org:/10.1016/j.canlet.2014.11.016
Aerobic glycolysis, a metabolic hallmark of cancer, is associated with radioresistance in non-small cell lung cancer (NSCLC). Pyruvate kinase M2 isoform (PKM2), a key regulator of glycolysis, is expressed exclusively in cancers. However, the impact of PKM2 silencing on the radiosensitivity of NSCLC has not been explored. Here, we show a plasmid of shRNA-PKM2 for expressing a short hairpin RNA targeting PKM2 (pshRNA-PKM2) and demonstrate that treatment with pshRNA-PKM2 effectively inhibits PKM2 expression in NSCLC cell lines and xenografts. Silencing of PKM2 expression enhanced ionizing radiation (IR)-induced apoptosis and autophagy in vitro and in vivo, accompanied by inhibiting AKT and PDK1 phosphorylation, but enhanced ERK and GSK3β phosphorylation. These results demonstrated that knockdown of PKM2 expression enhances the radiosensitivity of NSCLC cell lines and xenografts as well as may aid in the design of new therapies for the treatment of NSCLC.
Knockdown of PKM2 expression increases the sensitivity of NSCLC cells to radiotherapy in vitro
To examine PKM2 expressions levels in the normal lung epithelial cell and the NSCLC cell lines, we evaluated the expression levels of PKM2 in normal lung bronchial epithelial cell BEAS-2B and five NSCLC cell lines including A549, H460, H1299, H292, and H520 by Western blotting assays, and our results demonstrated that PKM2 expression was elevated in almost five NSCLC cell lines examined compared to autologous normal lung bronchial epithelial cell, although the expression levels fluctuated slightly depending on the different cell lines (Fig.1A). To test the role of PKM2 in the sensitivity of NSCLC to radiotherapy, we generated plasmids of pshRNA-PKM2 and control pshRNA-Con by inserting the DNA fragment for a pshRNA specifically targeting the PKM2 or control into the pGenesil2 vector. After demonstrating the authenticity, A549 and H460 cells were transfected with the plasmid for 48h and the levels of PKM2 expression were tested by Western blot assays. Obviously, transfection with control plasmid did not significantly modulate PKM2 expression; while transfection with pshRNA-PKM2 reduced the levels of PKM2 expression (Fig.1B and Appendix: Supplementary Fig.S1A). Quantitative analysis revealed that transfection with pshRNA-PKM2 significantly reduced PKM2 expressions as compared with that in the mock-treated and control pshRNA-Con plasmid-transfected cells, respectively (p<0.05, Fig.1C). Mock-treated and pshRNA-PKM2-trasnfected A549 and H460 cells were subjected to IR (4Gy), and 12 and 24h after IR, these cells, together with un-irradiated mock-treated, pshRNA-Con-transfected, and pshRNA-PKM2-trasnfected cells, were tested for cell viability by trypan blue staining. Knockdown of PKM2 reduced the percentage of A549 viable cells by 12.6–20% and IR treatment decreased the frequency of viable cells by 17.1–28.2%. However, the percentages of viable cells in the PKM2-silencing and irradiated cells were reduced by 27.7–48.7%, which were significantly lower than that in other groups (Fig.1D, p<0.05). Furthermore, it was consistent with the above results of A549 cells that knockdown of PKM2 significantly reduced the percentage of H460 viable cells (Appendix: Supplementary Fig.S1B). In addition, to further validate PKM2 silencing on their radiosensitivity,unirradiated control, mock-treated, and pshRNA-PKM2 transfected A549 cells were subjected to IR (0, 2, 4, 6, and 8Gy), and two weeks after IR, these cells were tested for the capacity for colony formation. The results showed that the numbers of colonies formed by pshRNA-PKM2 cells were significantly decreased compared with that of mock-treated and control cells; however, there was no significant change in mock-treated cells compared with control cells. These results suggested that pshRNA-PKM2 cells were more sensitive to IR than mock-treated and control cells (Fig.1E and F). Given that IR usually causes DNA double-strand breaks [28], we characterized the frequency of γ-H2AX nuclear foci positive cells by immunofluorescent assays. While IR treatment dramatically increased the frequency of γ-H2AX+ cells, the same dose of IR further significantly increased the percentages of γ-H2AX+ cells when combined with PKM2 silencing at 12 and 24h after IR, and there was a significant difference in γ-H2AX+ cells between these two groups at 12 and 24 h after IR (Fig. 1G and H, p < 0.05).
Fig. 1. The PKM2 expression levels in the normal lung epithelial cell and the NSCLC cell lines and knockdown of PKM2 expression enhance the radiosensitivity of A549 cells in vitro. The expression levels of PKM2 in normal lung bronchial epithelial cell BEAS-2B and five NSCLC cell lines including A549, H460, H1299, H292, and H520 were determined by Western blotting assay (A). A549 cells were transfected with pshRNA-PKM2 or pshRNA-Con plasmid for 48h, and the levels of PKM2 expression were determined by Western blot assays using a PKM2-specific antibody and β-actin as an internal control (B and C). Data are representative images or expressed as mean±SD of the relative levels of PKM2 to control β-actin in individual groups of cells from three separate experiments. # p
Knockdown of PKM2 enhances IR-induced apoptosis in NSCLC cells
Next, we tested the impact of PKM2-silencing on IR-induced cell death types. One day after IR, the apoptotic cells in the irradiatedmock-treated,pshRNA-PKM2-trasnfected cells, and one group of cells that had been pre-treated with 30μM Z-VAD for 1h prior to IR, together with mock-treated, unirradiated pshRNA-Contransfected, and pshRNA-PKM2-trasnfected groups of cells were characterized by TUNEL assays and/or FACS analysis (Fig.2A and C). In comparison with that in mock-treated and control plasmid transfected cells, the frequency of apoptotic cells in the PKM2 silencing or IR-treated cells increased moderately, while the percentages of apoptotic cells in the cells receiving combined treatment with IR and PKM2-silencing were significantly greater. However, the frequency of apoptotic cells in the Z-VAD-pretreated cells was partially reduced. Apparently, knockdown of PKM2 and IR induced apoptosis in NSCLC cells in vitro (Fig. 2B and D, and Appendix: Supplementary Fig.S1C).
Fig. 2. Knockdown of PKM2 expression enhances IR-induced apoptosis in A549 cells. A549 cells were transfected with, or without, pshRNA-Con or pshRNA-PKM2 for 48h and treated with, or without, Z-VAD for 1h. Subsequently, the cells were subjected to IR, and 24h later, the frequency of apoptotic cells was determined by TUNEL assays and FACS. (A and C) TUNEL and FACS analyses of apoptotic cells. (B and D) Quantitative analysis of the percentage of apoptotic cells. Data are representative images or expressed as mean%±SD of individual groups of cells from three independent experiments. * p
Knockdown of PKM2 enhances IR-induced autophagy in NSCLC cells
The cell autophagy is characterized by the formation of numerous autophagic vacuoles, autophagosome, in the cytoplasm and elevated levels of the microtubule-associated protein 1 light chain 3 (LC3)-II [29]. To test the impact of PKM2 silencing on IR-induced autophagy, the presence of autophagosome in mock-treated, pshRNACon-transfected, pshRNA-PKM2-transfected, IR-treated alone, IR + pshRNA-PKM2-transfected, and 1 mM 3-MA-pretreated IR + pshRNA-PKM2-transfected cells was characterized by electronic microphotography (EM). Intriguingly and importantly, numerous autophagosomes were detected in the IR + pshRNAPKM2-transfected cells, and only a few were detected in the sensitivity of the NSCLC cells to radiotherapy in vitro. It was noted that pshRNA-Con had almost no effect on A549 cells, therefore, some subsequently experiments did not set this group.
Fig. 3. Knockdown of PKM2 and IR induce A549 cell autophagy. A549 cells were transfected with, or without, pshRNA-Con or pshRNA-PKM2 for 48h and treated with, or without, 3-MA for 1h. Subsequently, the cells were subjected to IR, and 2h later, the presence of autophagic vacuoles and autolysosomes in A549 cells was determined by EM and the relative levels of LC3-I, LC3-II, AKT, ERK1/2, and control β-actin expression and AKT, ERK1/2, GSK3β, PDK1 phosphorylation were determined by Western blot assays using specific antibodies. Data are representative images and expressed as mean values of the relative levels of target protein to control in individual groups of cells from three separate experiments. The relative levels of target protein to control in mock-treated cells were designated as 1. (A) EM analysis of autophagic vacuoles and autophagosomes. Black arrows point to autophagic vacuoles and autophagosomes in the cytoplasma of A549 cells. (B) Western blot analysis of LC3-I and LC3-II expression. The values indicate the ratios of the relative levels of LC3-II to LC3-I in individual groups. (C) Western blotting analysis of individual signal events. The values indicate the relative levels of target protein to control β-actin in individual groups of cell
Fig. 4. The impact of 3-MA or/and V-ZAD on cell viability, colony formation, apoptosis and autophagy in A549 cells. A549 cells were transfected with, or without, pshRNACon or pshRNA-PKM2 for 48h and pre-treated with, or without, 3-MA or V-ZAD for 1h, respectively. Subsequently, the cells were subjected to IR. Twenty-four hours later and two weeks, the viability, apoptosis, and colony formation were determined. Two hours after treatment, autophagy and the relative levels of LC3-I and LC3-II expression in different groups of cells were determined. Data are representative images and expressed as mean%±SD of individual groups of cells from three separate experiments. (A) The percentages of viable cells. (B) The capacity of cell colony formation. (C) Quantitative analysis of apoptotic cells. (D) Western blot analysis of LC3-I and LC3-II expression. The values indicate the ratios of LC3-II to LC3-I in individual groups of cells. * p
Fig. 5. Treatment with pshRNA-PKM2 enhances the IR-inhibited growth of implanted tumors in mice. The nude mice were inoculated with A549 cells and when the tumor grew at 50mm3 in one dimension, the mice were randomized and treated with vehicle (PS), plasmid of pshRNA-Con or pshRNA-PKM2 alone or IR (4Gy×7f) alone or in combination with pshRNA-PKM2 and IR, respectively. The body weights and tumor growths of individual mice were monitored longitudinally. At the end of the in vivo experiment, the tumor tissues were dissected out and the frequency of apoptotic cells, the presence of autophagosomes and the expression of PKM2 were determined by TUNEL, EM and immunohistochemistry, respectively. Data are representative images or expressed as mean±SD of individual groups of mice (n=6 per group). (A) The body weights of mice. (B and C) The tumor growth curve of implanted tumors and the log-transformed tumor growth curve of implanted tumors in mice. (D) Quantitative analysis of the frequency of apoptotic cells.(E) EM analysis of autophagy. (F)The expression of PKM2.(G) Quantitative analysis of PKM2 expression.The cells with brown cytoplasma were considered as positive anti-PKM2 staining and the percentage of PKM2-positive cells was obtained by dividing the numbers of the PKM2-positive cells by the total number of cancer cells in the same field.
11.1.7 The tyrosine kinase inhibitor nilotinib has antineoplastic activity in prostate cancer cells but up-regulates the ERK survival signal—Implications for targeted therapies
Schneider M1, Korzeniewski N2, Merkle K2, Schüler J, et al.
Urol Oncol. 2015 Feb; 33(2):72.e1-7
http://dx.doi.org:/10.1016/j.urolonc.2014.06.001
Background: Novel therapeutic options beyond hormone ablation and chemotherapy are urgently needed for patients with advanced prostate cancer. Tyrosine kinase inhibitors (TKIs) are an attractive option as advanced prostate cancers show a highly altered phosphotyrosine proteome. However, despite favorable initial clinical results, the combination of the TKI dasatinib with docetaxel did not result in improved patient survival for reasons that are not known in detail. Methods: The National Cancer Institute-Approved Oncology Drug Set II was used in a phenotypic drug screen to identify novel compounds with antineoplastic activity in prostate cancer cells. Validation experiments were carried out in vitro and in vivo. Results: We identified the TKI nilotinib as a novel compound with antineoplastic activity in hormone-refractory prostate cancer cells. However, further analyses revealed that treatment with nilotinib was associated with a significant up-regulation of the phospho-extracellular-signal-regulated kinases (ERK) survival signal. ERK blockade alone led to a significant antitumoral effect and enhanced the cytotoxicity of nilotinib when used in combination. Conclusions: Our findings underscore that TKIs, such as nilotinib, have antitumoral activity in prostate cancer cells but that survival signals, such as ERK up-regulation, may mitigate their effectiveness. ERK blockade alone or in combination with TKIs may represent a promising therapeutic strategy in advanced prostate cancer.
Identification of nilotinib as a novel antineoplastic compound in prostate cancer cells
Using the NCI-Approved Oncology Drug Panel II for a phenotypic drug screen of normal prostate epithelial cells and prostate cancer cell lines (Fig. 1) [7], we identified the TKI nilotinib as a positive hit in hormone-refractory DU-145 prostate cancer cells.
Fig. 1. Discovery of nilotinib as a novel antineoplastic agent in prostate cancer cells using a phenotypic drug screen. Overview of the drug screen procedure (see text for details).
Results were confirmed using annexin V staining, which showed a significant induction of apoptosis beginning at 24 hours (Fig. 2A). The IC50 of nilotinib against DU-145 cells was determined at 10 μM using an MTT cell viability assay (Fig. 2B). Immunoblot experiments confirmed an induction of apoptosis using PARP cleavage in DU-145 cells and in hormonerefractory PC-3 prostate cancer cells at this drug concentration (Fig. 2C). An onset of apoptosis at 24 hours was likewise confirmed using PARP cleavage at a nilotinib concentration of 10 μM(Fig. 2D). PWR-1E prostate epithelial cells and hormone-sensitive prostate LNCaP prostate cancer cells were not found to undergo enhanced apoptosis when treated with nilotinib (not shown).
Fig. 2. Antitumoral effects of nilotinib in prostate cancer cells: (A) flow cytometric analysis of DU-145 prostate cancer cells for annexin V to detect apoptotic cells after treatment with 10 μM of nilotinib for the indicated intervals; (B) cell viability (MTT) assay to determine the IC50 of nilotinib in DU-145 cells (24-h treatment); (C and D) immunoblot analysis of DU-145 and PC-3 prostate cancer cells for PARP cleavage (arrow) at nilotinib concentrations and time intervals as indicated. GAPDH is shown for protein loading; and (E) colony growth assay of DU-145 cells after drug treatment and washout as shown. Cells grown in 60-mm dishes were stained with crystal violet to visualize viable cells at the time points indicated. (Color version of figure is available online.
To further confirm the effect of nilotinib on prostate cancer cell growth, we performed a colony growth assay in which DU-145 cells were treated with nilotinib for 72 hours followed by a washout of the drug and continued culture for additional 9 days (Fig. 2E). We found that nilotinib induced significant cytotoxicity after 72 hours and that a minor regrowth of cancer cells did not occur until 6 to 9 days after the washout, which is comparable to other TKIs [8]. Next, we sought to identify the targets of nilotinib in DU-145 prostate cancer cells. Overall, 5 well-established targets, including ABL1, KIT, PDGFRA, DDR1, and NQO2, were analyzed for their role in the drug response. We found that protein expression of 3 of these targets (ABL1, KIT, and PDGFRA) was not detectable in DU-145 cells and that small interfering RNA–mediated knockdown of the remaining 2 targets, DDR1 and NQO2, did not result in apoptosis (not shown). Collectively, these results show a significant antitumoral activity of nilotinib in prostate cancer cells. However, this effect was associated with a relatively high IC50 and was independent of known nilotinib targets.
Nilotinib up-regulates the ERK survival signal in prostate cancer cells
To further investigate why relatively high concentrations of nilotinib were required to induce cytotoxicity, we analyzed 40,6-diamidino-2-phenylindole–stained DU-145 cells treated with 10 μM of nilotinib for 24 hours using fluorescence microscopy (Fig. 3A).
Fig. 3. Nilotinib up-regulates the ERK survival signal in prostate cancer cells. (A) Fluorescence microscopic analysis of DAPI-stained DU-145 cells. (B and C) Immunoblot analyses of DU-145 cells (B) or DU-145 cells in comparison with LNCaP and PC-3 cells (C) treated with nilotinib for the expression of phospho-ERK1/2 T202/Y204 and total ERK. Immunoblot for GAPDH is shown as a loading control. (D) Immunohistochemical staining of xenografted DU-145 cells after 21 days of treatment with 75 mg/kg/d of nilotinib for phospho-ERK1/2 T202/Y204 expression. It can be noted that tumors explanted from vehicle-treated mice showed mostly positivity at the tumor periphery, whereas tumors explanted from nilotinib-treated mice showed a more evenly distributed phospho-ERK immunostaining (left panels). Quantification of phospho-ERK–positive DU-145 xenografts explanted after 21 days of treatment. Mean and standard errors of positive cells per high-power field (HPF; [1]40) from at least 3 tumors are given (right panel). (E) Immunoblot analysis of DU-145 cells treated with U0126 alone or in combination with nilotinib shows abrogation of phospho-ERK1/2 T202/Y204 expression by U0126. (F) Quantification of viable cells compared with that of controls using the MTT assay after treatment with U0126 (10 μM) or nilotinib (10 μM) or both and after either pretreatment (24 h) or simultaneous treatment (72 h). DAPI ¼ 40,6-diamidino-2-phenylindole. (Color version of figure is available online.)
We found that, despite the presence of apoptotic cells, there was also a population of actively dividing tumor cells in the presence of nilotinib as well as a population of viable but multinucleated cells (Fig. 3A). We interpreted these results as evidence that a subset of tumor cells has the ability to resist TKI treatment. To reconcile these results, we analyzed the activation of ERK1/2, which is known to function as a prosurvival signal in TKI-treated tumor cells [9,10]. We detected a robust overexpression of phospho-ERK1/2 T202/Y204 in nilotinib-treated DU-145 cells (Fig. 3B). An up-regulation of phospho-ERK1/2 T202/Y204 was also detectable in nilotinib-treated LNCaP cells, albeit at a lower level, and was not found in PC-3 cells (Fig. 3C). To further corroborate the evidence of phospho-ERK upregulation in vivo, we analyzed explanted DU-145 xenografts from a representative experiment in which nilotinib was used at a 75-mg/kg/d concentration. This initial dosage was based on published animal experiments [11] but yielded no or incomplete tumor control in our experiment (data not shown).
In vivo antitumoral activity of nilotinib and ERK blockade
Our results raised 2 important questions First, can a higher dose of nilotinib induce improved tumor control, and second, is a combination of nilotinib with the MEK inhibitor U0126 to block ERK activity superior to nilotinib alone?
Fig. 4. In vivo antitumoral activity of nilotinib and ERK blockade in prostate cancer cells: (A) tumor growth curves of DU-145 xenografts in NMRI-nude mice and (B) analysis of tumor volumes on day 21. Asterisks indicate statistical significance (**P r 0.01 and ***P r 0.001). (Color version of figure is available online.)
11.1.8 PAF and EZH2 Induce Wnt.β-Catenin Signaling Hyperactivation
Jung HY1, Jun S, Lee M, Kim HC, Wang X, Ji H, McCrea PD, Park JI
Mol Cell. 2013 Oct 24; 52(2):193-205
http://dx.doi.org/10.1016%2Fj.molcel.2013.08.028
Fine-control of Wnt signaling is essential for various cellular and developmental decision making processes. However, deregulation of Wnt signaling leads to pathological consequences including cancer. Here, we identify a novel function of PAF, a component of translesion DNA synthesis, in modulating Wnt signaling. PAF is specifically overexpressed in colon cancer cells and intestinal stem cells, and required for colon cancer cell proliferation. In Xenopus laevis, ventrovegetal expression of PAF hyperactivates Wnt signaling, developing secondary axis with β-catenin target gene upregulation. Upon Wnt signaling activation, PAF is dissociated from PCNA, and directly binds to β-catenin. Then, PAF recruits EZH2 to β-catenin transcriptional complex, and specifically enhances Wnt target gene transactivation, independently of EZH2’s methyltransferase activity. In mouse, conditional expression of PAF induces intestinal neoplasia via Wnt signaling hyperactivation. Our studies reveal an unexpected role of PAF in regulating Wnt signaling, and propose a novel regulatory mechanism of Wnt signaling during tumorigenesis. Fine-control of Wnt signaling is essential for various cellular and developmental decision making processes. However, deregulation of Wnt signaling leads to pathological consequences including cancer. Here, we identify a novel function of PAF, a component of translesion DNA synthesis, in modulating Wnt signaling. PAF is specifically overexpressed in colon cancer cells and intestinal stem cells, and required for colon cancer cell proliferation. In Xenopus laevis, ventrovegetal expression of PAF hyperactivates Wnt signaling, developing secondary axis with β-catenin target gene upregulation. Upon Wnt signaling activation, PAF is dissociated from PCNA, and directly binds to β-catenin. Then, PAF recruits EZH2 to β-catenin transcriptional complex, and specifically enhances Wnt target gene transactivation, independently of EZH2’s methyltransferase activity. In mouse, conditional expression of PAF induces intestinal neoplasia via Wnt signaling hyperactivation. Our studies reveal an unexpected role of PAF in regulating Wnt signaling, and propose a novel regulatory mechanism of Wnt signaling during tumorigenesis.
Keywords: Wnt, β-catenin, PAF, KIAA0101, EZH2
Strict regulation of stem cell proliferation and differentiation is required for mammalian tissue homeostasis, and its repair in the setting of tissue damage. These processes are precisely orchestrated by various developmental signaling pathways, with dysregulation contributing to disease and genetic disorders, including cancer (Beachy et al., 2004). Cancer is initiated by the inactivation of tumor suppressor genes and activation of oncogenes. For instance, colon cancer cells harbor genetic mutations in Wnt/β-catenin pathway constituents such as adenomatous polyposis coli (APC), Axin, and β-catenin (Polakis, 2007). In mouse models, inactivation of APC or activation of β-catenin results in the development of intestinal hyperplasia and adenocarcinoma (Moser et al., 1990), indicating that hyperactivation of Wnt signaling promotes intestinal tumorigenesis.
In canonical Wnt signaling, Wnt ligand induces stabilization of β-catenin protein via inhibition of the protein destruction complex (glycogen synthase kinase 3, APC, casein kinase I, and Axin). Then, activated β-catenin is translocated into the nucleus and binds to its nuclear interacting partners, TCF/LEF. Finally, β-catenin-TCF/LEF transactivates the expression of its target genes (Clevers and Nusse, 2012).
Although various Wnt/β-catenin modulators have been identified (Wnt homepage; wnt.stanford.edu), the pathological relevance of these modulators to tumorigenesis remains elusive. Also, many reports have suggested that mutation-driven Wnt signaling activation can be enhanced further (Goentoro and Kirschner, 2009; He et al., 2005; Suzuki et al., 2004; Vermeulen et al., 2010), which implies the presence of an additional layer of Wnt-signaling regulation in cancer beyond genetic mutations in APC or β-catenin. Here, we unraveled a novel function of the DNA repair gene, PAF (PCNA-associated factor) /KIAA0101). PAF was shown to be involved in translesion DNA synthesis (TLS), an error-prone DNA repair process that permits DNA replication machinery to replicate DNA lesions with specialized translesion DNA polymerase (Emanuele et al., 2011; Povlsen et al., 2012; Sale et al., 2012). Our comprehensive approaches uncover that cancer-specifically expressed PAF hyperactivates Wnt/β-catenin signaling and induces intestinal tumorigenesis.
Mitogenic role of PAF via Wnt signaling
To identify colon cancer-specific Wnt signaling regulators, we analyzed multiple sets of human colon cancer tissue samples using the publicly available database (www.oncomine.org), and selected genes that are highly expressed in colon cancer cells (fold change > 2; P < 0.0001; top 10% ranked). Among several genes, we investigated the biological role of PAF, based on its significant overexpression in human colon adenocarcinoma with correlated expression of Axin2, a well-established specific target gene of β-catenin (Jho et al., 2002; Lustig et al., 2002)(Figure 1A). To validate our in silico analysis, we performed immunostaining of colon cancer tissue microarray, and confirmed that PAF was highly expressed in colon cancer cells, whereas its expression was barely detectable in normal intestine (Figure 1B). Consistently, PAF was strongly expressed and mainly localized in the nucleus of colon cancer cell lines (Figure 1C). Additionally, we found that PAF was not expressed in non-transformed cells such as NIH3T3, mouse embryonic fibroblasts, and mammary epithelial cells (data not shown). Next, to assess the relevance of PAF upregulation in colon cancer cell proliferation, we depleted endogenous PAF using short hairpin RNAs (shRNAs) in these cell lines. Intriguingly, PAF knockdown (sh-PAF) inhibited colon cancer cell proliferation (Figures 1D and 1E). Given that PAF was shown to interact with PCNA via PIP box (Yu et al., 2001), we also examined whether PAF-PCNA interaction is required for mitogenic effects of PAF. In reconstitution experiments, sh-PAF-induced cell growth inhibition was rescued by ectopic expression of both shRNA non-targetable wild-type PAF (nt-PAF) and PIP mutant PAF (mutPIP-PAF) (Figure 1F), indicating that the PAF-PCNA interaction is not necessary for PAF-mediated colon cancer cell proliferation. Interestingly, PAF knockdown downregulated cell proliferation–related genes (Cyclin D1 and c-Myc) (Figure 1G). Given that Cyclin D1 and c-Myc are β-catenin direct target genes (He et al., 1998; Tetsu and McCormick, 1999), PAF likely participates in regulating Wnt/β-catenin signaling. Interestingly, PAF depletion-induced downregulation of Cyclin D1 andc-Myc was only observed in SW620 colon cancer cells, but not in Panc-1 and MDA-MB-231 cells (Figure 1G), indicating the specific effects of PAF on Cyclin D1 and c-Myc expression in colon cancer cells. We also assessed the effects of PAF knockdown on Axin2. Indeed, PAF knockdown suppressed Axin2transcription in colon cancer cells (Figure 1H). Moreover, as nt-PAF did, β-catenin ectopic expression reverted sh-PAF–induced cell growth arrest (Figure 1I), implying that PAF might be functionally associated with Wnt/β-catenin. We also examined whether other mitogenic signaling pathways mediate PAF’s mitogenic role. Of note, except HT29, other colon cancer cell lines (SW620, HCT116, HCC2998, and HCT15) harbor oncogenic mutations in K-Ras gene. Nonetheless, PAF depletion induced the suppression of cell growth on all five colon cancer cells (Figure 1D), indicating that PAF’s mitogenic function is independent of Ras/MAPK signaling activation. Additionally, overexpression of wild-type Akt or constitutively active form of Akt (myristoylated form of Akt [Myr-Akt]) did not rescue sh-PAF-induced inhibition of cell proliferation (Figure 1I). Moreover, β-catenin activation did not revert cell proliferation suppression resulted from MAPK or PI3K inhibition (Figure 1J), indicating that β-catenin-mediated mitogenic function is independent of MAPK and PI3K signaling pathways. These results suggest that PAF contributes to colon cancer cell proliferation, possibly via Wnt/β-catenin signaling.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040269/bin/nihms573362f1.gif
Figure 1 Mitogenic role of PAF in colon cancer cells
PAF positively modulates Wnt signaling
Given that many cancers develop as a result of deregulation of developmental signalings (Beachy et al., 2004), analyzing PAF expression during development may provide insights into the mechanisms of PAF-mediated signaling regulation. Whole mount immunostaining of mouse embryos, showed that PAF was specifically enriched in the apical ectodermal ridge (AER) of the limb bud, midbrain, hindbrain, and somites (Figure 2A and data not shown). During limb development, AER induction is specifically coordinated by active Wnt signaling (Figure 2B)(Kengaku et al., 1998). Using, Axin2-LacZ, a β-catenin reporter (Lustig et al., 2002), mouse embryos, we confirmed the specific activation of Wnt signaling in AER (Figure 2C). Intriguingly, Wnt signaling activity as exhibited in the AER, overlapped with the pattern of PAF expression (Figures 2A and 2C). Given that (1) Wnt signaling is deregulated in most colon cancer, (2) PAF is highly overexpressed in colon cancer cells, (3) PAF is required for colon cancer cell proliferation (Figure 1D), and (4) PAF is enriched in AER where Wnt signaling is active (Figure 2A), we hypothesized that PAF modulates the Wnt signaling pathway. To test this, we first examined the impact of PAF on β-catenin transcriptional activity using TOPFLASH reporter assays. In HeLa cells, PAF knockdown decreased β-catenin reporter activation by 6-bromoindirubin-3′-oxime, a GSK3 inhibitor (Figure 2D). Similarly, Wnt3A-induced transcriptional activation of Axin2 was also inhibited by PAF depletion (Figure 2E). These data suggest that PAF might be required for Wnt target gene expression.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040269/bin/nihms573362f2.gif
Figure 2 Activation of Wnt signaling by PAF
To gain better insight of PAF’s role in Wnt signaling regulation, we utilized Xenopus laevis embryos for axis duplication assays (Funayama et al., 1995), as previously performed (Park et al., 2009). Because of Wnt signaling’s pivotal role in vertebrate anterior-posterior axis development, the effects of Xenopus PAF (xPAF) on Wnt signaling can be monitored and quantified on the basis of secondary axis formation following injection of in vitro transcribed mRNAs. xβ-catenin mRNA, titrated to a subphenotypic level when expressed in isolation, was co-injected with xPAF mRNA into ventrovegetal blastomeres. Unlike the controls (β-catenin and β-galactosidase mRNA), the experimental group (β-catenin and xPAF mRNA) displayed axis-duplications (Figures 2F-H). Of note, the ventrovegetal injection of xPAF mRNA alone failed to induce secondary axes (data not shown), showing that PAF hyperactivates Wnt/β-catenin signaling only in the presence of active β-catenin. Consistent with the results of axis duplication assays, qRT-PCR assays showed that xPAF expression upregulated expression of Siamois and Xnr3, β-catenin targets in frogs (Figure 2I). Furthermore, we examined the specificity of PAF on Wnt/β-catenin signaling activity, using various luciferase assays. Ectopic expression of PAF hyperactivates Wnt3A or LiCl, a GSK3 inhibitor, -induced activation of β-catenin target gene reporter activity (MegaTOPFLASH, Siamois, c-Myc, and Cyclin D1). Of note, BMP/Smad pathway also plays an essential role in the developing limb AER (Ahn et al., 2001). However, PAF knockdown or overexpression did not affect BMP/Smad or FoxO signalings, respectively, (Figure 2J) indicating the specificity of PAF in regulating Wnt signaling. These results suggest that PAF positively modulates Wnt/β-catenin signaling in vitro and in vivo.
PAF-EZH2-β-catenin transcriptional complex formation
Next, we investigated the molecular mechanism underlying PAF hyperactivation of Wnt signaling. Given that stabilization of β-catenin protein is a key process in transducing Wnt signaling, we asked whether PAF affects β-catenin protein level. However, we found that the level of β-catenin protein was not altered by PAF knockdown or overexpression (Figures 2E and and3A),3A), leading us to test whether PAF controls the β-catenin/TCF transcriptional complex activity. Owing to the nuclear specific localization of PAF in colon cancer cells (Figure 1C), we tested whether PAF interacts with β-catenin transcriptional complex. Using a glutathione S-transferase (GST) pull-down assay, we found that PAF bound to β-catenin and TCF proteins (Figure 3B). Also, endogenous PAF interacted with β-catenin and TCF3 in SW620 cells that display constitutive hyperactivation of Wnt signaling by APC mutation (Figure 3C). Moreover, binding domain mapping assays showed that the Armadillo repeat domain of β-catenin was essential for its interaction with PAF (Figure 3D). Although PAF is a cell cycle-regulated anaphase-promoting complex substrate (Emanuele et al., 2011), PAF-β-catenin interaction was not affected (Figure S1). These data suggest that PAF directly binds to β-catenin transcriptional complex and this interaction is independent of cell cycle. Next, due to interaction of PAF with β-catenin and TCF, we tested whether PAF is also associated with Wnt/β-catenin target genes. First, we analyzed the subnuclear localization of PAF by chromatin fractionation. We found that PAF was only detected in the chromatin fraction of HCT116 cells (Figure 3E). Additionally, chromatin immunoprecipitation (ChIP) assays showed that both ectopically expressed and endogenous PAF occupied the TCF-binding element (TBE)-containing proximal promoter of the β-catenin targets (c-Myc and Cyclin D1) in HCT116 cells (Figures 3F and 3G). These data show that PAF is specifically associated with the promoters of Wnt/β-catenin target genes.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040269/bin/nihms573362f3.gif
Figure 3 PAF-EZH2-β-catenin transcriptional complex at target gene promoters
In intestine, Wnt/β-catenin signaling constitutively activates intestinal stem cells (ISCs) to give rise to progenitor cells, which replenishes intestinal epithelium (Figure 3H). Given the involvement of PAF on Wnt/β-catenin signaling regulation (Figure 2), we analyzed the spatial expression of PAF in intestinal epithelium. Immunostaining showed that PAF was specifically expressed in B lymphoma Mo-MLV insertion region 1 homolog (Bmi1) positive intestinal stem cells (ISCs)(Figures 3I and 3J). Bmi1 and its associated components in Polycomb-repressive complex 1 (PRC1) and 2 (PRC2) are shown to epigenetically regulate gene expression (Sparmann and van Lohuizen, 2006). Due to (1) specific association of PAF with TBEs of β-catenin target promoters (Figures 3F and 3G) and (2) co-localization with Bmi1 positive ISCs (Figure 3J), we asked whether PAF is associated with components of PRC1 and PRC2, using co-immunoprecipitation (co-IP) assays. Intriguingly, PAF interacted with both Bmi1 and enhancer of zeste homolog 2 (EZH2) in SW620 cells (Figure 3K), which led us to test whether either Bmi1 or EZH2 is functionally associated with PAF-mediated Wnt signaling hyperactivation. To do this, we assessed the effects of Bmi1 and EZH2 on β-catenin transcriptional activity, using β-catenin reporter assays. We observed that ectopic expression of EZH2 upregulated β-catenin transcriptional activity, but Bmi1 overexpression did not (data not shown), implying that EZH2 might be associated with Wnt signaling activation. Binding domain mapping analysis showed that EZH2 bound to PAF via the middle region of EZH2 including the CXC cysteine-rich domain (Figure 3L). In conjunction with the Bmi1-containing PRC1, EZH2-containing PRC2 catalyzes histone H3 lysine 27 trimethylation (H3K27me3) via histone methyltransferase domain. Despite the crucial role of EZH2 in H3K27me3-meidated gene regulation, we found that other core components of PRC2, EED, and Suz12 were not associated with PAF (Figure 3K). Moreover, although EZH2 overexpression in cancer induces PRC4 formation in association with the NAD+-dependent histone deacetylase Sirt1 (Kuzmichev et al., 2005), the PAF-EZH2 complex did not contain Sirt1 (Figure 3K). These data indicate that PAF-EZH2 complex is distinct from the conventional PRCs in cancer cells. Also, we questioned whether PCNA is required for PAF’s interaction with either PAF or β-catenin. Interestingly, β-catenin-PAF and EZH2-PAF complexes existed only in PCNA-free fractions (Figure 3M, compare lanes 1 and 2), which is consistent with PCNA-independent mitogenic role of PAF in colon cancer cell proliferation (Figure 1I). Due to exclusive interaction of PAF with either PCNA or β-catenin, we asked whether Wnt signaling activation affects either PAF-β-catenin or PAF-PCNA interaction. Co-IP assays showed that, in HeLa cells, PAF-β-catenin interaction was only detected upon LiCl treatment, while PAF-EZH2 interaction remained constant. Moreover, PAF-PCNA association was decreased by LiCl or Wnt3A treatment (Figures 3N and 3O, compare lanes 3 and 4). These data suggest that Wnt signaling activation is required for PAF-β-catenin interaction. Due to absence of endogenous Wnt signaling activity in HeLa cells, we also assessed the effects of active Wnt/β-catenin signaling on PAF-PCNA binding in colon cancer cell lines that exhibit hyperactivation of Wnt signaling by genetic mutations in APC or β-catenin alleles. Surprisingly, PAF-PCNA interaction was barely detectable in colon cancer cell lines, whereas 293T and HeLa cells displayed strong PAF-PCNA association (Figure 3P), implying that active β-catenin may sequester PAF from PCNA. In binding domain mapping analysis, we also found that N-terminal and PIP regions are required for β-catenin interaction (Figure S2), suggesting that β-catenin competes with PCNA for PAF interaction. These results suggest that, upon Wnt signaling activation, PAF is conditionally associated with β-catenin transcriptional complex.
PAF activates β-catenin target genes by recruiting EZH2 to promoters
Previous studies showed that EZH2 interacts with β-catenin (Li et al., 2009; Shi et al., 2007). Also, we found that PAF is physically associated with EZH2, independently of PRC2 complex (Figure 3). These evidences prompted us to ask whether EZH2 mediates PAF-induced Wnt signaling hyperactivation. Given PAF-EZH2-β-catenin complex formation, we tested whether EZH2 is also associated with the promoters of β-catenin target genes. Intriguingly, PAF, EZH2, and β-catenin steadily co-occupied the promoters of c-Myc,Cyclin D1, and Axin2 in HCT116 cells carrying β-catenin mutation, whereas PCNA, EED, and Suz12 did not (Figure 4A), which recapitulates PRC2 complex-independent association of EZH2 with PAF (see Figures 3K and 3N). Next, we asked whether PAF, EZH2, and β-catenin are recruited to β-catenin target gene promoter upon Wnt signaling activation, as PAF-β-catenin interaction was dependent of Wnt signaling activation (Figure 3N). In HeLa cells, we found that PAF, EZH2, and β-catenin conditionally bound to TBEs in the c-Myc and Axin2 promoters, only upon LiCl treatment (Figure 4B), indicating that Wnt signaling activation is a prerequisite for PAF-β-catenin-EZH2’s promoter association. To further confirm the specificity of PAF-EZH2-β-catenin’s recruitment to β-catenin target promoters, we performed ChIP promoter scanning of 10 kb of the c-Myc promoter, and found that PAF, EZH2, and β-catenin specifically co-occupied the proximal promoter containing TBEs of the c-Myc gene (at -1037 and -459 bp) (He et al., 1998) in HCT116 cells (Figure 4C). Also, the analysis of EZH2 ChIP-sequencing data from mouse embryonic stem cells showed that EZH2 was specifically enriched in the proximal promoters of β-catenin targets (Lef1, Lgr5, c-Myc, and Axin2) (Figure 4D).
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040269/bin/nihms573362f4.gif
Figure 4 PAF promotes EZH2-β-catenin interaction
Next, we asked whether EZH2 promoter recruitment is necessary for activation of β-catenin target gene transcription. Previously, depletion of EZH2 was shown to inhibit c-Myc expression in DLD-1 colon cancer cells (Fussbroich et al., 2011). Consistently, EZH2 knockdown downregulated β-catenin target genes, Axin2and Cyclin D1 in HCT116 cells (Figure 4E), and decreased LiCl-induced β-catenin reporter activation (Figure 4F), suggesting that EZH2 is required for PAF-mediated Wnt target gene hyperactivation. These results are also supported by previous finding that EZH2 enhances β-catenin transcriptional activity by connecting β-catenin with the Med1/RNA polymerase II (Pol II) complex (Shi et al., 2007). Indeed, Med1/TRAAP220 and Pol II conditionally binds to c-Myc and Axin2 promoters in LiCl-treated HeLa cells (Figure 4G). Given that PRC2-indepednent interaction between EZH2 and PAF (Figures 3K and 3N), we asked whether EZH2’s histone methyltransferase activity is dispensable in β-catenin regulation. We utilized an EZH2 point mutant (F681I) that disrupts the contact between the EZH2 hydrophobic pocket and histone lysine residue H3K27 (Joshi et al., 2008). Ectopic expression of either EZH2 or EZH2-F681I enhanced β-catenin reporter activity (Figure 4H). Also, PAF knockdown did not change the H3K27 methylation status (H3K27me3) of proximal promoters of c-Myc, Axin2, Cyclin D1, and DCC in HCT116 cells (Figure 4I). These results indicate a methyltransferase-independent role of EZH2 in transactivating β-catenin targets.
Due to PAF’s (1) small size (111 amino acids, one α-helix), (2) lack of a specific catalytic domain, and (3) binding to both β-catenin and EZH2, PAF may facilitate the interaction between EZH2 and β-catenin through recruiting EZH2 to the promoter. We tested this using ChIP assays for EZH2 in the setting of PAF depletion. Indeed, PAF-depleted HCT116 cells displayed decreased EZH2-association at the c-Myc promoter (Figure 4J), suggesting that PAF assists or is needed to recruit EZH2 to β-catenin transcriptional complex. Also, β-catenin knockdown decreased recruitment of PAF and EZH2 to promoters (Figure 4K), showing that PAF and EZH2 occupy target promoters via β-catenin. We then asked whether PAF promotes β-catenin-EZH2 binding. In vitro binding assays showed that the addition of GST-PAF protein increased EZH2-β-catenin association (Figure 4L). Moreover, ectopic expression of PAF promoted the EZH2-β-catenin interaction in HeLa cells treated with LiCl (Figure 4M). Additionally, we tested whether Wnt signaling-induced post-translational modification of either β-catenin or PAF is required for EZH2 interaction. However, in GST pull-down assays, we found that bacterially expressed either GST-β-catenin or –PAF bound to EZH2 (Figure S3). Due to the lack of post-translational modification in GST protein expression system, these data indicate that post-translation modification of either β-catenin or PAF is not necessary for EZH2 interaction. Together, these results suggest that PAF acts as a molecular adaptor to facilitate EZH2-β-catenin complex, and subsequently enhances the transcriptional activity of the β-catenin transcriptional complex at Wnt target promoters (Figure 4N).
Intestinal tumorigenesis following PAF conditional expression
Having determined that PAF is overexpressed in colon cancer cells and hyperactivates Wnt/β-catenin signaling, we aimed to determine whether mimicking PAF overexpression drives intestinal tumorigenesis, using genetically engineered mouse models. To conditionally express PAF, we generated doxycycline (doxy)-inducible PAF transgenic mice (TetO-PAF-IRES-emGFP [iPAF]). For intestine-specific expression of PAF, we used iPAF:Villin-Cre:Rosa26-LSL-rtTA mouse strains. Villin-Cre is specifically expressed in intestinal epithelial cells (IECs), including ISCs and progenitor cells. Cre removes a floxed stop cassette (loxP-STOP-loxP [LSL]) from the Rosa26 allele and induces rtTA expression. Upon doxy treatment, rtTA drives the transcriptional activation of the tetracycline-responsive element promoter, resulting in conditional transactivation of PAF selectively in IECs. We also utilized the Rosa26-rtTA strain for ubiquitous expression of PAF (Figure 5A and Figure S4). First, we examined the effects of PAF induction on IEC proliferation using a crypt organoid culture system (Figure S5A). Intriguingly, PAF conditional expression (2 weeks) induced expansion of the crypt organoids (Figures 5B and 5C), which recapitulates the mitogenic function of PAF (Figure 1). In mouse, IEC-specific PAF expression (iPAF:Villin-Cre:Rosa26-LSL-rtTA; 2 months) developed adenoma in both small intestine and colon (Figure 5D). Also, microscopic analysis using hematoxylin and eosin (H&E) staining showed aberrant IEC growth and crypt foci formation (Figures 5E and 5F), with disorganized epithelial cell arrangements (Figure S5B). Consistently, PAF-induced IEC hyperproliferation was manifested by increased Ki67 expression, a mitotic marker (Figure 5G). Importantly, these lesions exhibited the upregulation of CD44, a β-catenin target gene, whereas CD44 was expressed strictly in the crypts of normal intestine (Figure 5H). Next, we examined whether PAF directly hyperactivates Wnt/β-catenin in vivo using BAT-gal, a β-catenin reporter transgenic mouse carrying multiple TBEs followed by a LacZ reporter. To quantify the early effects of PAF on β-catenin activity, we treated mice with doxy for 1 week, and found that short-term induction of PAF increased β-catenin transcriptional activity as represented by enhanced X-gal staining (Figure 5I). Moreover, conditional PAF expression upregulated the β-catenin target genes, Axin2, Lgr5, CD44, Cyclin D1, and c-Myc in crypt organoids (Figure 5J). Additionally, mice ubiquitously expressing PAF exhibited intestinal hypertrophy (Figure S5C), which is similar to that induced by R-Spondin1, a secreted Wnt agonist (Kim et al., 2005). These data strongly suggest that PAF expression is sufficient to initiate intestinal tumorigenesis via Wnt signaling hyperactivation.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4040269/bin/nihms573362f5.gif
Figure 5 Induction of intestinal neoplasia by PAF expression
Herein we reveal the unexpected role of PAF in modulating Wnt/β-catenin signaling. PAF enhances the transcription of Wnt targets by recruiting EZH2 to the β-catenin transcriptional complex. This is similar to the mechanism by which Lgl/BCL9 binds to β-catenin and thereby recruits the PHD-finger protein Pygopus, to bridge the β-catenin/TCF complex to Med12 and Med13 (Carrera et al., 2008). Importantly, due to specific overexpression of PAF in cancer cells, our studies identified an additional layer of the regulatory mechanism of β-catenin target gene transactivation.
In cancer cells, the upregulation of EZH2 contributes to tumorigenesis through the epigenetic repression of various genes including tumor suppressor genes, Wnt antagonists, and DNA repair genes (Chang et al., 2011; Cheng et al., 2011; Kondo et al., 2008). Our results propose a noncanonical function of EZH2 in activating β-catenin target genes in conjunction with PAF. Consistently, recent study also suggests methyltransferase activity-independent function of EZH2 in gene activation (Xu et al., 2012). Moreover, this non-canonical role of EZH2 is supported by several lines of evidence: (a) EZH2 transactivates β-catenin target genes (Li et al., 2009; Shi et al., 2007) (Figures 4E and 4F); (b) EZH2 overexpression in murine mammary epithelium induces ductal hyperplasia (Li et al., 2009), which phenocopies that in a ∆Nβ-catenin (constitutively active form of β-catenin) mouse model (Imbert et al., 2001); (c) EZH2 occupies β-catenin target promoters (Figures 4A-D); and (d) EZH2’s methyltransferase activity is dispensable for β-catenin target activation (Figures 4H and 4I). Moreover, similar to PAF expression in the AER (Figure 2A), EZH2 is also specifically expressed there to maintain of Hox cluster gene transcription (Wyngaarden et al., 2011). Thus, it is plausible that EZH2 and PAF cooperatively control Hox gene activation in the developing limb. Interestingly, despite the presence of a physical and functional connection between Bmi1 and EZH2 in H3K27me3-mediated gene repression, EZH2 is expressed only in crypt IECs including ISCs (Figure S6), whereas Bmi1 is expressed in ISCs at position 4 (Figure 3J), implying a Bmi1-independent role for EZH2 in gene regulation. These results demonstrate the novel function of EZH2 in β-catenin target gene activation, independent of the histone methyltransferase activity of EZH2.
Previously, we found that TERT, a catalytic subunit of telomerase, positively modulates Wnt signaling (Park et al., 2009), elucidating a non-telomeric function of telomerase in development and cancer. Here our results propose that one component of DNA damage bypass process also functions in regulating Wnt signaling, dependent of context. In cancer, PAF overexpression may play a dual role in inducing (a) cell hyperproliferation (via Wnt signaling hyperactivation) and (b) the accumulation of mutations arising from DNA lesion bypass (by PAF-mediated TLS) (Povlsen et al., 2012). Importantly, PAF is only expressed in cancer cells, but not in normal epithelial cells. Thus, upon DNA damage, instead of cell growth arrest to permit high-fidelity DNA repair, the PAF overexpression in cancer cells is likely to induce DNA lesion bypass by facilitating TLS. However, in the setting of Wnt signaling deregulation, nuclear β-catenin sequesters PAF from PCNA and utilize PAF as a co-factor of transcriptional complex, which induces Wnt signaling hyperactivation and possibly lead to increased mutagenesis.
We observed that PAF marked the stemness of ISCs and mouse embryonic stem cells (Figure S7), implicating its roles in stem cell regulation under physiological conditions. In a previous study, a PAFgermline knockout mouse model displayed defects in hematopoietic stem cell self-renewal (Amrani et al., 2011), suggesting a crucial role of PAF in stem cell maintenance and activation. In the intestine, β-catenin activation in Lgr5-positive or Bmi1-positive cells is sufficient to develop intestinal adenoma (Barker et al., 2009; Sangiorgi and Capecchi, 2008), suggesting an essential role of tissue stem cells in tumor initiation. Considering PAF expression in Bmi1-positive ISCs, PAF upregulation in ISCs likely hyperactivates the Wnt/β-catenin signaling and contributes to intestinal tumor initiation.
Despite the critical role of Wnt signaling in early vertebrate, development PAF germline knockout mice are viable (Amrani et al., 2011). It is noteworthy that, whereas deletion of any core component in the Wnt signaling pathway causes embryonic lethality, mice with germline knockout of Wnt signaling modulators, including Nkd1/2, Pygo1/2, and BCL9/9-2, exhibit no lethal phenotypes (Deka et al., 2010; Schwab et al., 2007; Zhang et al., 2007). This may result from the robustness of Wnt signaling during embryogenesis because of functional compensation not only via the presence of multiple Wnt signaling regulators per se but also via other types of signaling crosstalk. Therefore, as described previously in pRb studies (Sage et al., 2003), acute deletion of PAF in a conditional knockout mouse model may disrupt the developmental balance or tissue homeostasis, and then reveal the full spectrum of the physiological and pathological roles of PAF in tumorigenesis. Taken together, our findings reveal unexpected function of PAF and EZH2 in modulating Wnt signaling, and highlight the impacts of PAF-induced Wnt signaling deregulation on tumorigenesis.
11.1.9 PAF Makes It EZ(H2) for β-Catenin Transactivation
Xinjun Zhang1 and Xi He1
Mol Cell. 2013 Oct 24; 52(2)
http://dx.doi.org:/10.1016/j.molcel.2013.10.008.
In this issue of Molecular Cell, Park and colleagues (Jung et al., 2013) show that PAF (PCNA-associatedfactor) binds to and hyperactivates transcriptional function of β-catenin in colon cancer cells by recruiting EZH2 to the coactivator complex. PAF-β-catenin and PAF-PCNA interactions are competitive, raising the question of whether β-catenin might regulate PCNA-dependent DNA replication and repair.
Wnt signaling through stabilization of transcription co-activator β-catenin plays critical roles in animal development and tissue homeostasis, and its deregulation is involved in myriad human diseases including cancer (Clevers and Nusse, 2012). Notably, most colorectal cancers (CRCs) have elevated β-catenin signaling caused by mutations of Wnt pathway components such as the tumor suppressor APC (Adenomatosis polyposis coli) and β-catenin itself (Clevers and Nusse, 2012). Much effort has focused on studying β-catenin-dependent transactivation in CRCs, including the current study by Park and colleagues that identifies PAF as an unexpected β-catenin co-activator (Jung et al., 2013).
PAF, for PCNA (proliferating cell nuclear antigen)-associated factor (also known as KIAA0101 or p15PAF), is an interacting partner of PCNA (Yu et al., 2001). PCNA has a key role in DNA replication and repair by assembling various DNA polymerase and repair complexes at the replication fork (Mailand et al., 2013). Dynamic regulation of PAF abundance and/or interaction with PCNA appears to be important for engaging DNA damage repair and bypass pathways (Emanuele et al., 2011; Povlsen et al., 2012). PAF is overexpressed in many types of cancers and required for cell proliferation (e.g., Yu et al., 2001).
In the current study (Jung et al., 2013), Jung et al. show that PAF is overexpressed in CRCs in a manner that parallels expression of Axin2, an established Wnt/β-catenin target gene. PAF knockdown inhibits CRC proliferation, and this effect is independent of PAF-PCNA interaction and can be rescued by a PAF mutant that does not binds to PCNA or by β-catenin overexpression. PAF knockdown downregulates the expression of Wnt/β-catenin target genes Cyclin D1, c-Myc, and Axin2 in a CRC line, leading the authors to hypothesize that PAF participates in Wnt/β-catenin signaling. Indeed PAF knockdown reduces, and its overexpression augments, Wnt/β-catenin responsive TOPFLASH reporter and target gene expression induced by Wnt3a or by pharmacological agents that stabilize β-catenin. In Xenopus embryos, PAF synergizes with β-catenin to induce Wnt target gene expression and axis duplication (a hallmark of Wnt/β-catenin activation). In mouse embryos, PAF is highly expressed in regions known for Wnt/β-catenin signaling such as the apical ectodermal ridge of the limb bud. Therefore PAF appears to be a positive regulator of Wnt/β-catenin signaling in CRCs and vertebrate embryos.
PAF does not affect β-catenin protein levels and is localized in the nucleus. Protein binding assays show that PAF interacts, directly or indirectly, with β-catenin (via the Armadillo-repeat domain) and its DNA-bound partner TCF (T Cell factor). Indeed PAF is associated with promoters of Wnt/β-catenin target genes in chromatin in CRC cells. Interestingly in the mouse intestine, the PAF protein is enriched in Bmi (B lymphoma Mo-MLV insertion region 1 homolog)-positive stem cells (at the “+4” position) (Sangiorgi and Capecchi, 2008). Bmi1 is a component of Polycomb Repressive Complex 1 (PRC1), which, together with the PRC2 complex that modifies Histone H3, has critical functions in transcriptional epigenetic silencing. Previous studies have suggested that a core PRC2 component, EZH2 (enhancer of zeste homolog 2), is a partner and paradoxically a co-activator of β-catenin, acting in a manner that is independent of EZH2’s methyltransferase activity (Li et al., 2009; Shi et al., 2007). Jung et al. found that PAF indeed interacts with both Bmi1 and EZH2, but not other PRC2 components, and EZH2 overexpression augments β-catenin transcriptional activity. PAF, EZH2, and β-catenin are found to co-occupy promoters of several Wnt/β-catenin target genes in CRC and mouse ES cells, and PAF depletion decreases EZH2 association with the c-Myc promoter, and β-catenin depletion decreases the association of both PAF and EZH2 with the promoter. Thus the β-catenin-PAF-EZH2 complex appears to constitute a chain of co-activators (Figure 1), and indeed PAF, which binds to both β-catenin and EZH2, enhances β-catenin-EZH2 co-immunoprecipitation. Together with an earlier study (Shi et al., 2007), these results suggest a model that PAF brings EZH2 and the associated RNA polymerase II Mediator complex to β-catenin target genes for transactivation in CRCs (Figure 1). Consistent with this model, transgenic overexpression of PAF in the mouse intestine induces β-catenin-dependent target and reporter gene expression, intestinal overgrowth, and adenoma formation in vivo and crypt organoid expansion in vitro, resembling Wnt/β-catenin signaling activation in the gastrointestinal tract.

ceb2-catenin-transactivation-nihms532034f1
β-catenin transactivation
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3848709/bin/nihms532034f1.jpg
Figure 1 β-catenin transactivation mediated by PAF and EZH2 in the G1 phase and a speculative role of β-catenin in modulating PAF-PCNA-dependent DNA replication and repair/bypass pathways in the S phase.
PAF and EZH2 represent newer additions to β-catenin’s plethora of co-activators (Mosimann et al., 2009), which offer multiple routes to engage the basal transcription apparatus. These co-activators may have partially redundant and/or context-dependent functions for numerous Wnt/β-catenin-dependent gene programs. Mouse mutants that lack an individual β-catenin co-activator are often viable (MacDonald et al., 2009; Mosimann et al., 2009). Paf−/− mice are viable but exhibit defects in hematopoietic stem cell properties (Amrani et al., 2011). PAF is also expressed in self-renewing mouse ES cells but the expression is downregulated upon ES cell differentiation (Jung et al., 2013). Whether PAF has a general role in self-renewal of embryonic and adult stem cells through its role in β-catenin signaling or DNA replication and repair pathways remains to be investigated.
PAF-β-catenin interaction is observed under Wnt stimulation, likely as a consequence of β-catenin accumulation (Jung et al., 2013). In some cell types PAF is ubiquitinated and degraded by the anaphase promoting complex and thus exhibits the lowest level in the G1 phase of the cell cycle (Emanuele et al., 2011). In these cells PAF may have a limited role as a co-activator for β-catenin-dependent transcription, which primarily occurs in G1. But in CRC and other cancers where PAF is overexpressed, PAF may have a prominent role as a β-catenin co-activator.
PAF-PCNA interaction is well documented (e.g., Yu et al., 2001). Surprisingly however, in CRCs with high levels of β-catenin, PAF-PCNA interaction is barely detectable (Jung et al., 2013). Conversely, in cells where the basal level of Wnt/β-catenin signaling is low, PAF-PCNA interaction is detected but is diminished by Wnt3a or pharmacological agents that stabilize β-catenin (Jung et al., 2013). PAF seems to interact with β-catenin and PCNA via an overlapping domain (although this remains to be better defined), offering a possible explanation why PAF-β-catenin and PAF-PCNA complexes appear to be mutually exclusive (Jung et al., 2013). This may simply reflect the fact that PAF-β-catenin (for RNA transcription) and PAF-PCNA (for DNA replication/repair) complexes act in G1 and S, respectively (Figure 1). However, when β-catenin levels are high in Wnt-stimulated cells or in CRCs, one may speculate that β-catenin accumulation could inhibit PAF-PCNA complex formation in the S phase, thereby enabling Wnt/β-catenin signaling to modulate PAF-PCNA-dependent DNA replication and repair/bypass pathways (Figure 1). This would constitute an unsuspected role for Wnt/β-catenin signaling in genomic stability beyond its established transcriptional function and could have implications to tumorigenesis.
- Amrani YM, Gill J, Matevossian A, Alonzo ES, Yang C, Shieh JH, Moore MA, Park CY, Sant’Angelo DB, Denzin LK. J Exp Med. 2011;208:1757–1765. [PMC free article] [PubMed]
- Clevers H, Nusse R. Cell. 2012;149:1192–1205. [PubMed]
- Emanuele MJ, Ciccia A, Elia AE, Elledge SJ. Proc Natl Acad Sci USA. 2011;108:9845–9850.[PMC free article] [PubMed]
- Jung H-Y, Jun S, Lee M, Kim H-C, Wang X, Ji H, McCrea PD, Park J-I. Molecular Cell. 2013 this issue, *bxs. [PMC free article] [PubMed]
- Li X, Gonzalez ME, Toy K, Filzen T, Merajver SD, Kleer CG. Am J Pathol. 2009;175:1246–1254.[PMC free article] [PubMed]
- MacDonald BT, Tamai K, He X. Dev Cell. 2009;17:9–26. [PMC free article] [PubMed]
- Mailand N, Gibbs-Seymour I, Bekker-Jensen S. Nat Rev Mol Cell Biol. 2013;14:269–282.[PubMed]
11.1.10 PI3K.AKT.mTOR pathway as a therapeutic target in ovarian cancer
Li H1, Zeng J, Shen K.
Arch Gynecol Obstet. 2014 Dec; 290(6):1067-78
http://dx.doi.org:/10.1007/s00404-014-3377-3
Background: Ovarian cancer is one of the major causes of death in women worldwide. Despite improvements in conventional treatment approaches, such as surgery and chemotherapy, a majority of patients with advanced ovarian cancer experience relapse and eventually succumb to the disease; the outcome of patients remains poor. Hence, new therapeutic strategies are urgently required. The phosphatidylinositol 3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) is activated in approximately 70 % of ovarian cancers, resulting in hyperactive signaling cascades that relate to cellular growth, proliferation, survival, metabolism, and angiogenesis. Consistent with this, a number of clinical studies are focusing on PI3K pathway as an attractive target in the treatment of ovarian cancer. In this review, we present an overview of PI3K pathway as well as its pathological aberrations reported in ovarian cancer. We also discuss inhibitors of PI3K pathway that are currently under clinical investigations and the challenges these inhibitors face in future clinical utility.Methods: PubMed was searched for articles of relevance to ovarian cancer and the PI3K pathway. In addition, the ClinicalTrials.gov was also scanned for data on novel therapeutic inhibitors targeting the PI3K pathway. Results: Genetic aberrations at different levels of PI3K pathway are frequently observed in ovarian cancer, resulting in hyperactivation of this pathway. The alterations of this pathway make the PI3K pathway an attractive therapeutic target in ovarian cancer. Currently, several inhibitors of PI3K pathway, such as PI3K/AKT inhibitors, rapamycin analogs for mTOR inhibition, and dual PI3K/mTOR inhibitors are in clinical testing in patients with ovarian cancer. Conclusions: PI3K pathway inhibitors have shown great promise in the treatment of ovarian cancer. However, further researches on selection patients that respond to PI3K inhibitors and exploration of effective combinatorial therapies are required to improve the management of ovarian cancer.
Fig.1. Inputs from receptor tyrosine kinases (RTKs) and G protein-coupled receptors (GPCR) to class I PI3K.
Fig. 2. Schematic representation of the PI3K/AKT/mTOR signaling pathway.
Fig.3. PI3K/AKT/mTOR inhibitors.
AKT inhibitors
AKT inhibitors can be grouped into three classes including lipid based phosphatidylinositol (PI) analogs, ATP-competitive inhibitors, and allosteric inhibitors. Perifosine, which is the most clinically studied AKT inhibitor, is a lipid-based PIanalog that targets the pleckstrin homology domain of AKT, preventing its translocation to the cell membrane. Amongthe three classes of AKT inhibitors, allosteric AKT inhibitors display highly specific selectivity for AKT isoforms. Considering the genetic background of ovarian cancer, allosteric AKT inhibitors such as MK2206 that can target both AKT1 and AKT2 might be the best agents for treating ovarian cancer.In clinical trials, AKT inhibitors have shown similar toxicities to those caused by PI3K inhibitors, such as hyperglycemia, rashes, stomatitis, and gastrointestinal side effects [25].
mTOR inhibitors
Rapamycin and its analogs Rapamycin (sirolimus), a potent inhibitor of mTORC1, was first isolated in 1975 from the bacterium Streptomyces hygroscopicus. Rapamycin inhibits mTORC1 by first binding to the intracellular protein FK506 binding protein 12 (FKBP12). The resultant rapamycin–FKBP12 complex then binds to the FKBP12–rapamycin-binding domain (FRB) of mTORC1 and inhibits the serine/threonine kinase activity of mTORC1 via an allosteric mechanism. In contrast to mTORC1, the rapamycin–FKBP12 complex cannot interact with the FRB domain of mTORC2, and thus,mTORC2 is generally resistant to rapamycin treatment [12]. As rapamycin displays very poor water solubility, which limits its clinical use, several soluble ester analogs of rapamycin (rapalogs) have been developed [12]. Currently, these analogs include temsirolimus, everolimus, and ridaforolimus. Temsirolimus and everolimus are formulated for intravenous and oral administration, respectively. Ridaforolimus was initially developed as an intravenous formulation, but an oral formulation was subsequently produced [12,28]. Clinically, rapalogs are generally well tolerated, with the most common side effects including stomatitis, rashes, fatigue, hyperglycemia, hyperlipidemia, hypercholesterolemia, and myelosuppression [3,12,25].
ATP-competitive inhibitors
Different from rapalogs, ATP-competitive inhibitors do not require co-factors such as FKBP12 to bind to mTOR. By competingwith ATP for theATP-binding sites of mTOR, this class of mTOR inhibitors can inhibit the kinase activity of both mTORC1 and mTORC2. Although there is a concern that the simultaneous inhibition of mTORC1 and mTORC2 might result in greater toxicities in normal tissues, ATP-competitive mTOR inhibitors have been shown to display stronger anti-proliferative activity than rapalogs across a broad range of cancers includingovarian cancer [12,15].
Metformin
Metformin,the most commonly prescribed oral anti-diabetic agent, has been shown to reduce the incidence of malignancies in patients with diabetes. The activation of 5′ adenosine monophosphateactivated protein kinase (AMPK) by metformin plays an important role in mediating the drug’s effects. AMPK activation results in the phosphorylation and activation of TSC2, which exerts inhibitory effects on mTORC1. Metformin-induced AMPK activation also reduces AKT activity by inhibiting insulin receptor substrate 1 (IRS-1). Ultimately, AMPK activation results in the inhibition of the PI3K/AKT/mTOR signaling pathway, making metformin an effective treatment for cancer [28].
mTORC1 inhibitors mTORC1 Dual PI3K/mTOR inhibitors
PI3K inhibitors Class I PI3K mTORC2
AKT inhibitors AKT mTORC ½ inhibitors
PI3K inhibitors
Pan-class I PI3K inhibitors Pan-class IPI3K inhibitors can inhibit the kinase activity ofall 4 isoforms of classI PI3K.The main advantage of pan-class IPI3K inhibitors is that most cancer cells express multiple PI3K isoforms with redundant oncogenic signaling functions. Early clinical trials have suggested that the most common toxicitiesof pan-class IPI3K inhibitors are hyperglycemia, skin toxicities, stomatitis, and gastrointestinal side effects. Of these, hyperglycemia is likely to be a mechanism-based toxicity given the well described role of PI3K in insulin receptor signaling [3,25].
Isoform-selective PI3K inhibitors
This class of agents target the specific PI3K p110 isoforms involved in particular types of cancer. The p110α isoform (which is encoded by the PIK3CA gene) is a frequent genetic driver (PIK3CA mutations) of ovarian cancer, whereas p110β activity is known to be essential in cancer cells lacking PTEN. As for the p110δ isoform, it plays a fundamental role in the survival of normal B cells and is implicated in malignancies affecting this lineage. Thus, the main theoretical advantage of these inhibitors is that they have the potential to completely block the relevant target whilst causing limited toxicities compared with pan-PI3K inhibitors. Consistent withthese findings, preclinical studies have detected significant activities of PI3Kα inhibitor in tumors exhibiting PIK3CA mutations, PI3Kβ inhibitors in tumors with PTEN loss, and PI3Kδ inhibitors in hematologic malignancies. In addition, PI3Kδ inhibitors have already shown very promising activity in patients with chronic lymphocytic leukemia [26].
Dual PI3K/mTOR inhibitors
Structural similarities between the ATP-binding domain of p110 and the catalytic domain of mTOR have led to the development of a class of agents that inhibit both class I PI3K and mTORC1/2. Theoretically, dual mTOR/PI3K inhibitors should lead to more complete suppression of the PI3K/AKT/mTOR pathway than targeting either component independently.In agreement with this, in preclinical studies of ovarian cancer dual PI3K/mTOR inhibitors were found to exhibit greater in vitro and in vivo anti-tumor activity than mTOR inhibitors alone [27]. The safety profile of these inhibitors is similar to that of pan-PI3K inhibitors, with common adverse events including nausea, diarrhea, fatigue, and vomiting [3,25].
11.1.11 Endogenous, hyperactive Rac3 controls proliferation of breast cancer cells by a p21-activated kinase-dependent pathway
Mira JP1, Benard V, Groffen J, Sanders LC, Knaus UG.
Proc Natl Acad Sci U S A. 2000 Jan 4; 97(1):185-9.
Uncontrolled cell proliferation is a major feature of cancer. Experimental cellular models have implicated some members of the Rho GTPase family in this process. However, direct evidence for active Rho GTPases in tumors or cancer cell lines has never been provided. In this paper, we show that endogenous, hyperactive Rac3 is present in highly proliferative human breast cancer-derived cell lines and tumor tissues. Rac3 activity results from both its distinct subcellular localization at the membrane and altered regulatory factors affecting the guanine nucleotide state of Rac3. Associated with active Rac3 was deregulated, persistent kinase activity of two isoforms of the Rac effector p21-activated kinase (Pak) and of c-Jun N-terminal kinase (JNK). Introducing dominant-negative Rac3 and Pak1 fragments into a breast cancer cell line revealed that active Rac3 drives Pak and JNK kinase activities by two separate pathways. Only the Rac3-Pak pathway was critical for DNA synthesis, independently of JNK. These findings identify Rac3 as a consistently active Rho GTPase in human cancer cells and suggest an important role for Rac3 and Pak in tumor growth.
Uncontrolled cell proliferation is a major feature of cancer. Experimental cellular models have implicated some members of the Rho GTPase family in this process. However, direct evidence for active Rho GTPases in tumors or cancer cell lines has never been provided. In this paper, we show that endogenous, hyperactive Rac3 is present in highly proliferative human breast cancer-derived cell lines and tumor tissues. Rac3 activity results from both its distinct subcellular localization at the membrane and altered regulatory factors affecting the guanine nucleotide state of Rac3. Associated with active Rac3 was deregulated, persistent kinase activity of two isoforms of the Rac effector p21-activated kinase (Pak) and of c-Jun N-terminal kinase (JNK). Introducing dominant-negative Rac3 and Pak1 fragments into a breast cancer cell line revealed that active Rac3 drives Pak and JNK kinase activities by two separate pathways. Only the Rac3–Pak pathway was critical for DNA synthesis, independently of JNK. These findings identify Rac3 as a consistently active Rho GTPase in human cancer cells and suggest an important role for Rac3 and Pak in tumor growth.
Rac proteins are members of the Rho GTPase family and act as molecular switches in regulating a variety of biological response pathways, including cell motility, gene transcription, cell transformation, and cell-cycle progression (1). The Rac family includes Rac1, the myeloid-lineage-specific Rac2, and the recently cloned Rac3 proteins (2). Rac3 differs from Rac1 and Rac2 in two domains, the insert region and the C terminus, which influence transformation (3, 4), interaction with guanine nucleotide exchange factors (GEFs) (5, 6), and subcellular localization (7, 8). Small GTPases, including Rac, cycle between an inactive GDP-bound state and an active GTP-bound state. Two classes of regulatory factors, GTPase-activating proteins (GAPs) and GEFs, determine by their opposing effects the ratio of GDP versus GTP, which is bound to the GTPase (1). GAP proteins increase the intrinsic rate of GTP hydrolysis, rendering the GTPase inactive, whereas GEFs enhance the exchange of bound GDP for GTP, thereby activating the protein. Active Rac regulates distinct downstream signaling pathways by interacting with specific effector proteins, including a family of serine-threonine protein kinases termed Paks (p21-activated kinases) (9–11).
Apart from its well documented role in cytoskeletal rearrangements in growth factor-stimulated cells (12), Rac1 is required for Ras-induced malignant transformation and is involved in transcription and growth control (1, 13, 14). Recently, the importance of the Rac effector Pak in cell transformation has been highlighted by inhibiting RasV12- and Rac1V12-induced transformation of Rat-1 fibroblasts with a catalytically inactive form of Pak (15, 16). The involvement of Rac1 in driving cell-cycle progression through the G1 phase and stimulating DNA synthesis has been shown by introducing dominant-active and -negative Rac1 mutants into fibroblasts (17, 18). However, the signaling pathways used by Rac to control mitogenesis and proliferation still remain poorly understood. Overexpression of constitutively active Rac-effector-domain mutants in fibroblasts indicated that although Rac1 mediated cyclin D1 transcription by Pak in NIH 3T3 cells (19), Pak was not involved in the DNA synthesis of Swiss 3T3 cells (20). Accumulating evidence, however, suggests higher complexity where Pak-binding proteins, such as the GEF Pix, contribute to the Rac–Pak interaction in vivo and influence subsequent cellular functions (21–23).
All biological functions listed above have been attributed to Rac1 in experimental cell systems using overexpression or microinjection of mutant forms. Endogenously active Rho GTPases, including Rac, have not yet been observed. In this paper, we describe a consistently active Rac3 GTPase leading to hyperactivity of its effector protein kinase, Pak, in human breast cancer-derived epithelial cell lines. Analysis of growth properties and DNA synthesis revealed that both proteins are required to convey the highly proliferative phenotype displayed by these cells.
Highly Proliferating Cancer Cells Contain Hyperactive Rac3.
Comparison of growth rates among several breast cancer cell lines showed that three lines (MDA-MB 435, T47D, and MCF 7) grew faster under normal and low-serum conditions (Fig. (Fig.1).1). Interestingly, in contrast to MDA-MD 231 and Hs578T cells, these three highly proliferative cell lines do not possess mutated Ras (28, 29). To assess whether Rho GTPases drive this cellular phenotype, we determined whether these cell lines contained active GTP-bound Rac or Cdc42. We used a recently described assay, the PBD-pulldown assay (24), which is based on the specific binding of the GTP-bound forms of Rac and Cdc42 to the PBD of Pak (10). Neither active Rac1 (Fig. (Fig.22A) nor active Cdc42 (data not shown) could be detected in any of the cell lysates obtained from serum-starved cells. However, both proteins were detected if the PBD-pulldown assay was performed with in vitro guanosine 5′-[γ-thio]triphosphate (GTP[γS])-loaded cell lysates, confirming that Rac1 and Cdc42 were present in their inactive GDP-bound forms in these cells (Fig. (Fig.22A for Rac1). Next we wanted to determine whether active Rac3 was present in breast cancer cell lines. Because Rac3 effectors have not yet been characterized, we demonstrated by overlay binding and kinase assays that Rac3 bound to and activated Pak as efficiently as Rac1 (data not shown). We verified that the PBD-pulldown assay specifically detected the active GTP-bound form of Rac3 (GTP[γS]-loaded Rac3wt or Rac3V12, Fig. Fig.22B) and not the inactive form. To probe for Rac3 protein in breast cell lysates, a Rac3-specific antibody was used. GST-PBD-pulldown experiments from cell lysates revealed the presence of hyperactive Rac3 in highly proliferative cell lines (MDA-MB 435, T47D, and MCF 7), but not in normal breast cell lines or in less proliferative breast cancer cells (Fig. (Fig.22C). Additionally, as indicated by the virtual absence of Rac3 in the supernatant of the PBD pulldown, all the Rac3 protein present in these cell lines was active (Fig. (Fig.22C). To demonstrate that consistent Rac3 activation is not limited to cell lines, we performed an initial screening of human metastatic breast cancer tissues and found active Rac3 in one of three samples, underlining the potential clinical relevance of the cellular findings (Fig. (Fig.22D).

Differential growth rates of human breast cell lines. pq0104939001
Differential growth rates of human breast cell lines.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC26637/bin/pq0104939001.jpg
Figure 1 Differential growth rates of human breast cell lines. Human breast cell lines, including HMEC 184 (○), MDA-MB 231 (▵), Hs578T (□), MDA-MB 435 (●), T47D (▴), and MCF 7 (♦), were grown in 10% serum (A) or 0.5% serum (B) conditions. The cells were split in duplicate over 6-well plates at 5 × 105 cells per well and counted daily with a hemocytometer for 4 days. Data shown in A and B are representative of three independent experiments.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC26637/bin/pq0104939002.jpg
Figure 2 Active Rac3 is present in highly proliferative cell lines and in human breast cancer tissue. (A and C) Cell lysates from serum-starved breast cancer cell lines without (A and C) or after (+) GTP[γS] loading (A) were incubated with 10 μg of GST-PBD. Active Rac proteins (PBD pulldown) were detected by immunoblot with anti-Rac1 (A) or anti-Rac3 antibodies (C). Blotting of PBD supernatants revealed the GDP-bound form of Rac3 in lysates. Equal amounts of Rac3 protein were detected by immunoblot (IB) in all cell lines. (B) A PBD-pulldown assay of extracts from HeLa cells expressing Myc-Rac3wt or -Rac3 mutants, followed by an anti-Myc immunoblot, detected only active Rac3 (GTP[γS] loading or Rac3V12). (D) PBD pulldown of lysates obtained from three different human metastatic breast cancer tissues, followed by anti-Rac1 and anti-Rac3 immunoblots, revealed active Rac3 in tissue 1. (E) PBD pulldown of lysates derived from MDA-MB 435 and MDA-MB 231 cells expressing LacZ control or Myc-Rac3wt without or after in vitro GTP[γS] loading. Consistent activation of Myc-Rac3wt occurred only in MDA-MB 435 cells. (F) Subcellular localization of Rac1 and Rac3. Cytosol (c) and membranes (m) were obtained after nitrogen cavitation and fractionation of breast cancer cell lines and immunoblotted with anti-Rac1 and anti-Rac3 antibodies. All blots are representative of at least three experiments.
Subcellular Localization and GTPase-Regulatory Factors Influence Rac3 Activity.
Constitutive activation of Ras proteins in cancer cells is often caused by activating point mutations at the switch I or II regions (29). cDNA cloning and complete sequence analysis of full-length Rac3 did not reveal any mutations in the breast cell lines studied and did not explain the observed Rac3 activation. GTPase-regulatory proteins such as GEFs and GAPs, which are usually regulated by upstream stimuli, control cycling between the active and inactive forms of Rac. To confirm the presence of an altered regulatory mechanism involved in Rac3 activation, we used the PBD-pulldown assay to analyze the activation state of Myc-tagged Rac3wt transfected into either MDA-MB 231, a cell line harboring only GDP-Rac3, or MDA-MB 435, a cell line that contains endogenous, active GTP-Rac3. Fig. Fig.22E shows that activated Myc-Rac3 was detected only in the MDA-MB 435 cell line, confirming that the regulation of the GDP/GTP state of Rac3 was altered in these cells. We then investigated several upstream stimuli that have been shown to affect GTPase-regulatory proteins (28, 30–32). We excluded the possibility of an autocrine growth-stimulatory loop by culturing MDA-MB 231 cells with the conditioned medium from MDA-MB 435, which did not affect the Rac3 activation state (data not shown). Treatment of cell cultures with phosphatidylinositol 3-kinase or tyrosine kinase inhibitors, including wortmannin, LY294002, and genistein, did not decrease Rac3 activation (data not shown). At this point, we speculated that an oncogenic, Rac3-specific GEF is present in certain breast cancer cells. GEFs possess a pleckstrin homology domain that is essential for membrane localization and for their oncogenic properties (5, 33). Analysis of the subcellular localization of the Rac family members revealed that Rac3 is located in the membranes of breast epithelial cell lines, independently of its activation state (Fig. (Fig.22F). In contrast, endogenous Rac1 in its inactive GDP-bound state was essentially cytosolic (Fig. (Fig.22F). Thus, the distinct localization of Rac3 and Rac1 may contribute to their different activation states in certain breast cancer cell lines. It is conceivable that the highly proliferative cell lines (Fig. (Fig.1)1) express a constitutively active, membrane-bound Rho GEF that activates adjacent Rac3 protein. This hypothesis was further supported by using an hydroxymethylglutaryl-CoA reductase inhibitor, lovastatin, that interferes with isoprenoid synthesis and thereby with posttranslational processing of GTPases. Unprocessed Rac3 from lovastatin-treated MDA-MB 435 cells was predominantly cytosolic and inactive (GDP-Rac3) (data not shown). The requirement of membrane localization for consistent Rac3 activity was further supported by using a Rac3S189 mutant. Replacing cysteine-189 of the CAAX box with serine abolishes isoprenoid incorporation, rendering the GTPase cytosolic. This Rac3 mutant remained in its inactive GDP-bound state when transfected into MDA-MB 435 cells (data not shown).
Several Rho GTPase-regulating GEFs have been identified (5), including the Rac1-specific GEF Tiam-1, which has been linked to tumors such as invasive T-lymphomas (34). Although Tiam-1 is expressed in virtually all tissues, no evidence of oncogenic truncations or alternative splicing of Tiam-1 transcripts has been found (35). A variation of Tiam-1 transcript levels in certain cancer cell lines might lead to overexpression and possibly activation of Tiam-1 protein. However, the activation state of Rac3 protein in the cell lines used in this study does not seem to correlate with Tiam-1 expression levels as reported by Habets et al. (35). Hyperactivity of Rac3 in cancer cells could also result from an absent or dysfunctional Rac3-specific GAP protein. By accelerating the intrinsic GTP hydrolysis rate, GAPs render the GTPase inactive and act as tumor suppressors. Deletion or mutations in the RasGAP gene NF1 and the RhoGAP homologs bcr and DLC-1 have been reported in cancer cells (36, 37).
Active Rac3 Drives Epithelial Cell Proliferation.
To study whether active Rac3 could account for the high proliferation rate of certain breast cancer cells, we expressed a constitutively active Rac3 mutant (Rac3V12) in normal mammary epithelial cells (HMEC 184) that contain only GDP-Rac3 (Fig. (Fig.22C). Rac3V12 expression significantly increased the incorporation of BrdUrd into nascent DNA (Fig. (Fig.3),3), emphasizing that transfection of active Rac3 drives epithelial cell proliferation.

Rac3V12 induces DNA synthesis in human mammary epithelial cells pq0104939003
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC26637/bin/pq0104939003.jpg
Rac3V12 induces DNA synthesis in human mammary epithelial cells
Figure 3 Rac3V12 induces DNA synthesis in human mammary epithelial cells. HMEC 184 cells, infected with recombinant LacZ or Rac3V12 Semliki Forest virus, were allowed to express protein for 14 h in serum-free medium containing 10 μM BrdUrd. Cells were fixed and stained with anti-Myc antibody for Myc-Rac3V12 expression level (Upper) or with FITC-conjugated anti-BrdUrd antibody for BrdUrd incorporation (Lower). The presence of bright fluorescent nuclei indicates BrdUrd-positive cells. The percentage was calculated after counting 400 cells in each of three independent experiments.
Hyperactive Pak and c-Jun Kinases in Cancer Cells.
The signaling cascade utilized by Rac proteins to control cell proliferation still remains to be identified (1, 9), but might involve Paks. We analyzed Pak activity in cell lysates derived from serum-starved breast cancer cell lines by using in-gel kinase assays and by usingin vitro kinase assays after immunoprecipitation with Pak-specific antibodies. Pak activity was increased 4- to 6-fold in the three cell lines containing active Rac3 (Fig. (Fig.44A). This increased kinase activity was mainly associated with the Pak2 isoform, which can phosphorylate and positively regulate Raf-1 activity, another key component in cell proliferation (38–40).
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC26637/bin/pq0104939004.jpg
Figure 4 Rac3 activates Pak and JNK by two different pathways. (A) Breast cancer cell lysates from serum-starved cells were analyzed for Pak and JNK activities. Pak activities in cell lysates were analyzed by in-gel kinase assays. JNK activity was determined by …
Intracellular Rac-regulated signaling pathways impinge on distinct mitogen-activated protein kinase cascades. Constitutively active Rac has been shown to positively regulate the activity of the stress-activated kinases JNK and p38 (1). Moreover, ERK activity can be indirectly stimulated by Rac or mediated by crosstalk between the distinct mitogen-activated protein kinase cascades (1, 41). Determination of distinct mitogen-activated protein and stress-activated protein kinase activities in the breast cell lines studied here showed that consistent Rac3 and Pak kinase activities were associated with enhanced JNK activity (Fig. (Fig.44A). In contrast, no correlation existed between p38 or ERK kinase activities and active Rac3 or Pak (data not shown).
Rac3 Triggers Pak and JNK Activities by Separate Pathways.
To determine whether the highly proliferative phenotype of breast cancer cells depends directly on a consistently active Rac3-Pak-JNK cascade, we used virus-mediated protein expression in MDA-MB 435 cells to examine the ability of Rac3 and Paks to control JNK activation and cellular proliferation. The importance of Pak as an effector protein in Rac-mediated activation of JNK is still controversial and seems to be cell-type-dependent (42). Expression of the PBD domain, which controls the activity of both Rac and Pak (21), completely inhibited Pak and JNK stimulation (Fig. (Fig.44B). The mutation of leucine to phenylalanine at position 107 of the PBD domain suppresses the autoinhibitory function of the PBD (21). Thus, PBD F107 will act only to sequester active Rac3 and blocks its ability to bind and activate endogenous effectors. Expression of either dominant-negative Rac3N17 or PBD F107 almost completely blocked Pak and JNK activities, demonstrating that Rac3 is upstream of these proteins (Fig. (Fig.44B). Moreover, Pak kinase activity can be inhibited independently of Rac3 by overexpressing the kinase autoinhibitory domain, PID, which does not interact with Rac (21, 43). Transfection of PID into MDA-MB 435 cells dramatically inhibited Pak activity as expected, but did not decrease JNK activation (Fig. (Fig.44B). Our results indicate that in MDA-MB 435 cells, consistent stimulation of JNK by Rac3 is independent of PAK activity and that Rac3 initiates two different pathways involving Pak and JNK, respectively.
Rac3 and Pak Are Both Required for Breast Cancer Cell Proliferation.
We subsequently determined which of these two Rac3 pathways promoted the increased cell proliferation in breast cancer cell lines with hyperactive Rac3. We studied the consequence of expressing inhibitory Rac mutants or Pak fragments on DNA synthesis. LacZ-expressing MDA-MB 435 cells still proliferated in low-serum conditions and 35% incorporated BrdUrd (Fig. (Fig.5).5). This percentage increased to 50% when Rac3wt, which will be partially activated in these cells (Fig. (Fig.22E), is expressed (Fig. (Fig.55 Bottom Right). In contrast, expression of inhibitory proteins, including Rac3N17 or the PBD that suppressed Pak and JNK activation (Fig. (Fig.44B), almost completely blocked S-phase entry, as indicated by the absence of BrdUrd incorporation (Fig. (Fig.5).5). Expression of the PID that inhibited Pak kinase activity without affecting JNK stimulation (Fig. (Fig.44B) also arrested proliferation in MDA-MB 435 cells (Fig. (Fig.5).5). These experiments emphasize the crucial role of active Rac3 for DNA synthesis in breast cancer cell lines and demonstrate that Pak kinase activity is necessary for Rac3-induced proliferation.

Rac3 mediates proliferation in MDA-MB 435 cells pq0104939005
Rac3 mediates proliferation in MDA-MB 435 cells
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC26637/bin/pq0104939005.jpg
Figure 5 Rac3 mediates proliferation in MDA-MB 435 cells by a Pak-dependent pathway. MDA-MB 435 cells growing in 0.5% FBS were infected with Semliki Forest virus encoding for LacZ, Rac3N17, Pak1-PBD, Pak1-PBD F107, Pak1-PID, or Rac3wt. After 12 to14 h of protein expression in serum-free medium, 20 μM BrdUrd was added for 20 min before the cells were fixed and stained with anti-Myc antibody and phalloidin for expression (Top) or with FITC-conjugated anti-BrdUrd antibody for BrdUrd incorporation (Lower five micrographs). The presence of bright fluorescent nuclei indicates BrdUrd-positive cells. The percentage was calculated after counting 400 cells in each of four independent experiments.
Our results establish the persistent activation of a small Rho GTPase, Rac3, and the effector kinase Pak in human breast cancer cells. In contrast to Rac1, endogenous Rac3 is localized at the plasma membrane in both guanine nucleotide states. It seems likely that a Rac3 regulatory protein is altered or deleted in highly proliferating cancer cells, and that its specificity toward Rac3 results from the adjacent location of both proteins at the membrane and/or from discrete Rac3 domains, which convey a specific interaction. The cytoskeletal phenotypes of serum-starved breast cancer cells, such as ruffles or lamellipodia typical of Rac1 protein activation, did not seem to correlate with the GDP versus GTP state of endogenous Rac3. This may suggest that Rac family members are specialized in certain cellular functions, as already reported for Rac2 in leukocyte phagocytosis (44) and now demonstrated by us for Rac3 in cancer cell proliferation. Our studies establish further that endogenous, active Rac3 is essential for breast cancer cell proliferation via a Pak-dependent pathway. Paks have been shown to directly phosphorylate Raf kinase, which binds to retinoblastoma protein and regulates its function (45), and to interact with cyclin-dependent kinases to up-regulate cyclin D1 expression (46). Initial screening of various human cancer-derived cell lines revealed the presence of hyperactive Rac3 and Pak kinase in other types of highly proliferating tumors (data not shown). Further investigations, primarily in animal models and clinical settings, will be necessary to assess whether loss of Rac3 and Pak regulation correlates with certain breast tumor stages and is accompanied by specific alterations in cell-cycle regulators. Approaches to inhibit Rac3 or Pak activity would then open a new avenue for cancer therapeutics.
11.1.12 Curcumin-could-reduce-the-monomer-of-ttr-with-tyr114cys-mutation via autophagy in cell model of familial amyloid polyneuropathy.
Li H1, Zhang Y1, Cao L1, Xiong R1, Zhang B1, Wu L1, Zhao Z1, Chen SD2
Drug Des Devel Ther. 2014 Oct 31; 8:2121-8
http://dx.doi.org:/10.2147/DDDT.S70866.
Transthyretin (TTR) familial amyloid polyneuropathy (FAP) is an autosomal dominant inherited neurodegenerative disorder caused by various mutations in the transthyretin gene. We aimed to identify the mechanisms underlying TTR FAP with Tyr114Cys (Y114C) mutation. Our study showed that TTR Y114C mutation led to an increase in monomeric TTR and impaired autophagy. Treatment with curcumin resulted in a significant decrease of monomeric TTR by recovering autophagy. Our research suggests that impairment of autophagy might be involved in the pathogenesis of TTR FAP with Y114C mutation, and curcumin might be a potential therapeutic approach for TTR FAP.
Transthyretin (TTR) familial amyloid polyneuropathy (FAP) is an autosomal dominant inherited disease, characterized clinically by progressive sensory, motor, and autonomic impairment, which typically lead to death around a decade after diagnosis.1 Since the first identification of TTR with Val30Met mutation (TTR V30M), the most common gene mutation in FAP patients, more than 100 TTR mutations have been found to cause FAP.2 However, the detailed pathogenesis underlying TTR FAP remains undefined. Previous studies of the TTR V30M mutant have shown that misfolding and self-aggregation of TTR are implicated in the pathogenesis of TTR FAP involving abnormal endoplasmic reticulum (ER) stress.3
Corresponding to the various TTR gene mutations and a wide range of geographical distributions, FAP presents diverse characteristics in genotype-phenotype in different regions. We have recently published the first report of a TTR Tyr114Cys (TTR Y114C) mutation in a Chinese family with TTR FAP.4 Compared with TTR V30M, the TTR Y114C mutation showed different clinical manifestations, and was also observed in a Japanese family.5,6 This suggests that the pathogenesis of the TTR Y114C and TTR V30M mutations might be different. Studies focused on monomer generation and tetramer depolymerization have been performed.1,2 However, the mechanisms underlying the clearing of the abnormally increased monomer are unknown.
Autophagy is the major lysosomal pathway via which cells degrade intracytoplasmic protein. It is widely accepted that autophagy plays a key role in the process of amyloid deposition in certain neurodegenerative diseases, including alpha-synuclein, beta peptides, tau oligomers, and misfolded prion protein.7 Therefore, autophagy may be involved in degradation of the TTR monomer in TTR FAP.
Curcumin and its analogs have demonstrated a protective effect in many diseases involving antimicrobial, antitubercular,8 and anticancer mechanisms,9 and they can also modulate innate immunity.10 Of note, curcumin has been shown to promote autophagy.11 Therefore, we hypothesized that autophagy might be involved in the pathogenetic mechanism of the TTR Y114C mutation in TTR FAP and curcumin might have potential therapeutic role in this disease. In this study, we aimed to identify the role of autophagy in the pathogenetic mechanism of TTR FAP and to assess the therapeutic effect of curcumin in the disease.
TTR Y114C mutation led to increased monomeric TTR and impaired autophagy in vitro
To investigate the alteration of monomeric TTR with different mutations, we generated HEK293T cell lines with wild-type TTR, TTR Y114C, and stable overexpression of TTR V30M. Wild-type TTR represented the normal control and TTR V30M represented the positive control. Western blotting analysis of the TTR level in the cells when cultured for 24 hours showed that the monomer of TTR Y114C and TTR V30M was increased by approximately 2.3 times and 2.78 times, respectively, compared with wild-type TTR (Figure 1A and B). Mutation of TTR Y114C was related to the increase in monomeric TTR, as well as the mutation of TTR V30M.

Changes in autophagy and endoplasmic reticulum stress related to wild-type TTR, TTR V30M, and TTR Y114C dddt-8-2121Fig1
Changes in autophagy and endoplasmic reticulum stress related to wild-type TTR, TTR V30M, and TTR Y114C
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4222630/bin/dddt-8-2121Fig1.jpg
Figure 1 Changes in autophagy and endoplasmic reticulum stress related to wild-type TTR, TTR V30M, and TTR Y114C.
Next we investigated the activation of several markers associated with ER stress, including ER-resident chaperone BiP and p-eIF2α. Our results showed the levels of BiP and p-eIF2α is higher in TTR V30M than those in wild-type TTR. In contrast, BiP and p-eIF2α levels in TTR Y114C were similar to those in wild-type TTR (Figure 1A and C), indicating ER stress might not be the main pathogenetic mechanism for the TTR Y114C mutation. We then investigated whether autophagy plays a role in the mechanism of TTR Y114C mutation. LC3-II is well known to be a robust marker of autophagosomes, and immunofluorescent staining of LC3-II can be used to assay for autophagosome formation. A high ratio of LC3-II to LC3-I would indicate induction of autophagy. Our results revealed that the ratio of LC3-II/I was markedly decreased for TTR Y114C, but less suppressed for TTR V30M (Figure 1A and D). Likewise, a significant decrease in LC3-II immunoreactivity was detected in TTR Y114C (Figure 1E). The results of Western blotting and immunofluorescence indicated that autophagy in TTR Y114C was significantly downregulated. Therefore, impaired autophagy might be responsible for the pathogenesis of TTR Y114C mutation.
Curcumin decreased monomeric TTR by promoting autophagy
The effects of curcumin were investigated in TTR Y114C and wild-type TTR stable overexpressed HEK293T cells. Curcumin did not show toxic effects in the stable overexpressed cell lines at curcumin concentrations below 10 µM (Figure 2A and B). We chose 5 µM as the experimental concentration, because it is the minimal effective concentration of curcumin in these cell lines. Further, we wanted to determine whether curcumin could decrease monomeric TTR by promoting autophagy at the minimal effective concentration. Therefore, we used curcumin (2.5 µM and 5 µM) as a protective agent to assess whether it could decrease monomeric TTR with mutation by promoting autophagy. Quantification of LC3-II and LC3-I indicated markedly higher activation of LC3 in TTR Y114C treated with curcumin 5 µM for 24 hours (Figure 2D). In contrast, treatment with curcumin at different concentrations could not activate LC3 in wild-type TTR (Figure 2C, E). We next examined the ratio of monomers to tetramers in TTR Y114C, which was significantly decreased after 24 hours of treatment with 5 µM curcumin compared with no treatment with curcumin (Figure 2D and F). However, for wild-type TTR, the ratio of monomers to tetramers was unchanged after treatment with curcumin (Figure 2C and E). These results indicate that treatment with curcumin 5 µM for 24 hours was able to decrease the monomer in the TTR Y114C mutation by promoting autophagy.

Curcumin decreased monomeric TTR by promoting autophagy dddt-8-2121Fig2
Curcumin decreased monomeric TTR by promoting autophagy
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4222630/bin/dddt-8-2121Fig2.jpg
Figure 2 Curcumin decreased monomeric TTR by promoting autophagy.
Protective effect of curcumin on TTR Y114C could be partially blocked by 3-MA
To further validate whether the decrease in monomer by curcumin in our experiments was mediated by autophagy, 3-MA, an inhibitor of autophagosome formation, was implied to negatively regulate autophagy. 3-MA (1 mM) was added to the cell culture medium 2 hours before curcumin and incubated for 24 hours. Analysis of LC3, tetrameric TTR, and monomeric TTR from TTR Y114C revealed that 3-MA partly reversed the LC3 II activation induced by curcumin and increased the monomer of TTR Y114C (Figure 3). These results confirm that curcumin induced the decrease in the TTR Y114C monomer by promoting the autophagy pathway.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4222630/bin/dddt-8-2121Fig3.jpg
Figure 3 Protective effect of curcumin on TTR Y114C could be partially blocked by 3-MA.
Discussion
TTR FAP is a severe autosomal dominant inherited disease, for which the treatment options are limited. Liver transplantation performed early in the course of the disease is the only therapeutic strategy known to stabilize this neuropathy.1,13 More recently, tafamidis meglumine, a potent inhibitor of misfolding and deposition of mutated TTR, has completed an 18-month, placebo-controlled Phase II/III clinical trial for the treatment of FAP.14 However, in June 2012, the US Food and Drug Administration Peripheral and Central Nervous System Drugs Advisory Committee rejected this drug, stating a lack of convincing data supporting its efficacy.15 Hence, it is important to identify the pathogenetic mechanism of FAP to find an alternative effective treatment strategy.
Accumulating studies focused on the TTR mutation gene and protein have provided insights into the pathogenesis of TTR FAP, including decreased stability of TTR tetramers, conformational change in the crystal structure of variant TTR, altered kinetics of denaturation, and disturbing endoplasmic ER quality control system.1,16–18 Previous studies have demonstrated that increased levels of ER stress are correlated with extracellular TTR deposition. Two ER stress markers, BiP and p-eIF2α, have been observed to be present and upregulated in the salivary gland tissue of FAP patients.3 However, the precise molecular mechanisms underlying TTR FAP and its phenotypic heterogeneity are not yet fully understood.
Our current study investigated whether the two mutations, TTR Y114C and TTR V30M, share the same pathogenesis and evaluated the effect of pathogenic mutations on the clearance of the monomer. Our results show that the ratio of LC3-II/I was markedly decreased, while BiP and p-eIF2α levels remained constant in TTR Y114C when compared with wild-type TTR and TTR 30M. The results of our research indicate the impaired autophagy contributed to the TTR Y114C mutation, but not ER stress. This observation indicates that abnormal accumulation of TTR caused by a different mutation might be cleared by different pathways, and more studies are necessary to confirm whether this difference applies to other TTR mutations.
Curcumin is known to have neuroprotective properties through a variety of mechanisms.8–11 Our research indicates that curcumin decreased the monomeric TTR by promoting autophagy, and without toxic effects. Moreover, this protective effect of curcumin on TTR Y114C could be partially blocked by 3-MA. Pullakhandam et al showed that curcumin binds to wild-type TTR and prevents urea-induced perturbations in the tertiary structure of TTR in vitro.19 Recently, Ferreira et al reported that dietary curcumin modulated TTR amyloidogenicity.20 Therefore, curcumin might be an effective therapy for FAP involving multiple molecular pathways.
Overall, our findings show that abnormal accumulation of TTR caused by different mutations might be cleared in different ways, and curcumin might be an effective therapy for FAP by promoting autophagy. Further studies are necessary to determine whether this phenomenon exists in other TTR mutations.
Stephen Williams, PhD
For PI3K and related inhibitors of PI3K/AKT/mTOR i would refer you to two people who should be in the discussion of this signaling pathway and PI3K/AKT inhibitors used for chemotherapy. The first is Dr. Mien-Chie Hung and the second is Dr. Gordon Mills. They both had been at MD Anderson and developed some of the first inhibitors as well as the earliest discoveries of overactivity of PI3K/AKT in ovarian cancer.
Next the field had never progressed any inhibitors past Stage II as there has been some serious toxicities seen in preclinical phases (most long term tox studies are done after patients are enrolled in phase I).
I would refer to three papers
Discovery of GSK2126458, a Highly Potent Inhibitor of PI3K and the Mammalian Target of Rapamycin http://pubs.acs.org/doi/abs/10.1021/ml900028r
A new mutational AKTivation in the PI3K pathwayhttp://www.researchgate.net/publication/6146395_A_new_mutational_AKTivation_in_the_PI3K_pathway
These will show how inhibitors of certain isoforms of PI3K (namely delta) had to be developed to circumvent some of the severe toxicity seen with the earliest inhibitors (wortmanin and LY294002.
Also
Take your PIK: phosphatidylinositol 3-kinase inhibitors race through the clinic and toward cancer therapy http://mct.aacrjournals.org/content/8/1/1.full
Targeting the phosphoinositide 3-kinase (PI3K) pathway in cancerhttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC3142564/
Development of PI3K Inhibitors in Breast Cancer http://www.onclive.com/publications/contemporary-oncology/2014/November-2014/Development-of-PI3K-Inhibitors-in-Breast-Cancer by Aggerwal nice review
Phosphatidylinositol 3-kinase (PI3K) inhibitors as cancer therapeuticshttp://www.ncbi.nlm.nih.gov/pmc/articles/PMC3843585/ will explain about some of the toxicities and describes the one PI3K that has made it to phase II
Most of them have failed and I believe now are being thought as an adjuvant not front line therapy
Read Full Post »





















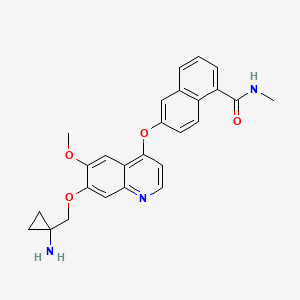












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Aurelian