Healthcare analytics, AI solutions for biological big data, providing an AI platform for the biotech, life sciences, medical and pharmaceutical industries, as well as for related technological approaches, i.e., curation and text analysis with machine learning and other activities related to AI applications to these industries.
Article SELECTION from Collection of Aviva Lev-Ari, PhD, RN Scientific Articles on PULSE on LinkedIn.com for Training Small Language Models (SLMs) in Domain-aware Content of Medical, Pharmaceutical, Life Sciences and Healthcare by 15 Subjects Matter
Article selection: Aviva Lev-Ari, PhD, RN
#1 – February 20, 2016
Contributions to Personalized and Precision Medicine & Genomic Research
Nobel Prize in Chemistry 2024 to David Baker, Demis Hassabis and John M. Jumper
Reporter: Aviva Lev-Ari, PhD, RN
UPDATED on 10/22/2024
ProteinMPNN, which is now available free on the open-source software repository GitHub, will give researchers the tools to make unlimited new designs. “The challenge, of course … is what are you going to design?” Baker says.
In a second Nobel win for AI, the Royal Swedish Academy of Sciences has awarded half the 2024 prize in chemistry to Demis Hassabis, the cofounder and CEO of Google DeepMind, and John M. Jumper, a director at the same company, for their work on using artificial intelligence to predict the structures of proteins. The other half goes to David Baker, a professor of biochemistry at the University of Washington, for his work on computational protein design. The winners will share a prize pot of 11 million Swedish kronor ($1 million).
The potential impact of this research is enormous. Proteins are fundamental to life, but understanding what they do involves figuring out their structure—a very hard puzzle that once took months or years to crack for each type of protein. By cutting down the time it takes to predict a protein’s structure, computational tools such as those developed by this year’s award winners are helping scientists gain a greater understanding of how proteins work and opening up new avenues of research and drug development. The technology could unlock more efficient vaccines, speed up research on cures for cancer, or lead to completely new materials.
Hassabis and Jumper created AlphaFold, which in 2020 solved a problem scientists have been wrestling with for decades: predicting the three-dimensional structure of a protein from a sequence of amino acids. The AI tool has since been used to predict the shapes of all proteins known to science.
“I’ve dedicated my career to advancing AI because of its unparalleled potential to improve the lives of billions of people,” said Demis Hassabis. “AlphaFold has already been used by more than two million researchers to advance critical work, from enzyme design to drug discovery. I hope we’ll look back on AlphaFold as the first proof point of AI’s incredible potential to accelerate scientific discovery,” he added.
Baker has created several AI tools for designing and predicting the structure of proteins, such as a family of programs called Rosetta. In 2022, his lab created an open-source AI tool called ProteinMPNN that could help researchers discover previously unknown proteins and design entirely new ones. It helps researchers who have an exact protein structure in mind find amino acid sequences that fold into that shape.
Most recently, in late September, Baker’s lab announced it had developed custom molecules that allow scientists to precisely target and eliminate proteins associated with diseases in living cells.
“[Proteins] evolved over the course of evolution to solve the problems that organisms faced during evolution. But we face new problems today, like covid. If we could design proteins that were as good at solving new problems as the ones that evolved during evolution are at solving old problems, it would be really, really powerful,” Baker told MIT Technology Review in 2022.
born 1962 in Seattle, WA, USA. PhD 1989 from University of California, Berkeley, CA, USA. Professor at University of Washington, Seattle, WA, USA and Investigator, Howard Hughes Medical Institute, USA.
University of Washington, Seattle, WA, USA
Howard Hughes Medical Institute, USA
Demis Hassabis “for protein structure prediction”
born 1976 in London, UK. PhD 2009 from University College London, UK. CEO of Google DeepMind, London, UK.
Google DeepMind, London, UK
John M. Jumper “for protein structure prediction”
born 1985 in Little Rock, AR, USA. PhD 2017 from University of Chicago, IL, USA. Senior Research Scientist at Google DeepMind, London, UK.
Google DeepMind, London, UK
The Nobel Prize in Chemistry 2024 is about proteins, life’s ingenious chemical tools. David Baker has succeeded with the almost impossible feat of building entirely new kinds of proteins. Demis Hassabis and John Jumper have developed an AI model to solve a 50-year-old problem: predicting proteins’ complex structures. These discoveries hold enormous potential.
“One of the discoveries being recognised this year concerns the construction of spectacular proteins. The other is about fulfilling a 50-year-old dream: predicting protein structures from their amino acid sequences. Both of these discoveries open up vast possibilities,” says Heiner Linke, Chair of the Nobel Committee for Chemistry.
Proteins generally consist of 20 different amino acids, which can be described as life’s building blocks. In 2003, David Baker succeeded in using these blocks to design a new protein that was unlike any other protein. Since then, his research group has produced one imaginative protein creation after another, including proteins that can be used as pharmaceuticals, vaccines, nanomaterials and tiny sensors.
The second discovery concerns the prediction of protein structures. In proteins, amino acids are linked together in long strings that fold up to make a three-dimensional structure, which is decisive for the protein’s function. Since the 1970s, researchers had tried to predict protein structures from amino acid sequences, but this was notoriously difficult. However, four years ago, there was a stunning breakthrough.
In 2020, Demis Hassabis and John Jumper presented an AI model called AlphaFold2. With its help, they have been able to predict the structure of virtually all the 200 million proteins that researchers have identified. Since their breakthrough, AlphaFold2 has been used by more than two million people from 190 countries. Among a myriad of scientific applications, researchers can now better understand antibiotic resistance and create images of enzymes that can decompose plastic.
Life could not exist without proteins. That we can now predict protein structures and design our own proteins confers the greatest benefit to humankind.
@@@@
This year’s Nobel Prize laureates in chemistry Demis Hassabis and John Jumper have developed an AI model to solve a 50-year-old problem: predicting proteins’ complex structures.
In 2020, Hassabis and Jumper presented an AI model called AlphaFold2. With its help, they have been able to predict the structure of virtually all the 200 million proteins that researchers have identified. Since their breakthrough, AlphaFold2 has been used by more than two million people from 190 countries. Among a myriad of scientific applications, researchers can now better understand antibiotic resistance and create images of enzymes that can decompose plastic.
Eight Subcellular Pathologies driving Chronic Metabolic Diseases – Methods for Mapping Bioelectronic Adjustable Measurements as potential new Therapeutics: Impact on Pharmaceuticals in Use
In this curation we wish to present two breaking through goals:
Goal 1:
Exposition of a new direction of research leading to a more comprehensive understanding of Metabolic Dysfunctional Diseases that are implicated in effecting the emergence of the two leading causes of human mortality in the World in 2023: (a) Cardiovascular Diseases, and (b) Cancer
Goal 2:
Development of Methods for Mapping Bioelectronic Adjustable Measurements as potential new Therapeutics for these eight subcellular causes of chronic metabolic diseases. It is anticipated that it will have a potential impact on the future of Pharmaceuticals to be used, a change from the present time current treatment protocols for Metabolic Dysfunctional Diseases.
According to Dr. Robert Lustig, M.D, an American pediatric endocrinologist. He is Professor emeritus of Pediatrics in the Division of Endocrinology at the University of California, San Francisco, where he specialized in neuroendocrinology and childhood obesity, there are eight subcellular pathologies that drive chronic metabolic diseases.
These eight subcellular pathologies can’t be measured at present time.
In this curation we will attempt to explore methods of measurement for each of these eight pathologies by harnessing the promise of the emerging field known as Bioelectronics.
Unmeasurable eight subcellular pathologies that drive chronic metabolic diseases
Glycation
Oxidative Stress
Mitochondrial dysfunction [beta-oxidation Ac CoA malonyl fatty acid]
Insulin resistance/sensitive [more important than BMI], known as a driver to cancer development
Membrane instability
Inflammation in the gut [mucin layer and tight junctions]
Epigenetics/Methylation
Autophagy [AMPKbeta1 improvement in health span]
Diseases that are not Diseases: no drugs for them, only diet modification will help
Image source
Robert Lustig, M.D. on the Subcellular Processes That Belie Chronic Disease
These eight Subcellular Pathologies driving Chronic Metabolic Diseases are becoming our focus for exploration of the promise of Bioelectronics for two pursuits:
Will Bioelectronics be deemed helpful in measurement of each of the eight pathological processes that underlie and that drive the chronic metabolic syndrome(s) and disease(s)?
IF we will be able to suggest new measurements to currently unmeasurable health harming processes THEN we will attempt to conceptualize new therapeutic targets and new modalities for therapeutics delivery – WE ARE HOPEFUL
In the Bioelecronics domain we are inspired by the work of the following three research sources:
Michael Levin is an American developmental and synthetic biologist at Tufts University, where he is the Vannevar Bush Distinguished Professor. Levin is a director of the Allen Discovery Center at Tufts University and Tufts Center for Regenerative and Developmental Biology. Wikipedia
THE VOICE of Dr. Justin D. Pearlman, MD, PhD, FACC
PENDING
THE VOICE of Stephen J. Williams, PhD
Ten TakeAway Points of Dr. Lustig’s talk on role of diet on the incidence of Type II Diabetes
25% of US children have fatty liver
Type II diabetes can be manifested from fatty live with 151 million people worldwide affected moving up to 568 million in 7 years
A common myth is diabetes due to overweight condition driving the metabolic disease
There is a trend of ‘lean’ diabetes or diabetes in lean people, therefore body mass index not a reliable biomarker for risk for diabetes
Thirty percent of ‘obese’ people just have high subcutaneous fat. the visceral fat is more problematic
there are people who are ‘fat’ but insulin sensitive while have growth hormone receptor defects. Points to other issues related to metabolic state other than insulin and potentially the insulin like growth factors
At any BMI some patients are insulin sensitive while some resistant
Visceral fat accumulation may be more due to chronic stress condition
Fructose can decrease liver mitochondrial function
A methionine and choline deficient diet can lead to rapid NASH development
@MIT Artificial intelligence system rapidly predicts how two proteins will attach: The model called Equidock, focuses on rigid body docking — which occurs when two proteins attach by rotating or translating in 3D space, but their shapes don’t squeeze or bend
Reporter: Aviva Lev-Ari, PhD, RN
This paper introduces a novel SE(3) equivariant graph matching network, along with a keypoint discovery and alignment approach, for the problem of protein-protein docking, with a novel loss based on optimal transport. The overall consensus is that this is an impactful solution to an important problem, whereby competitive results are achieved without the need for templates, refinement, and are achieved with substantially faster run times.
Keywords:protein complexes, protein structure, rigid body docking, SE(3) equivariance, graph neural networks
Abstract: Protein complex formation is a central problem in biology, being involved in most of the cell’s processes, and essential for applications such as drug design or protein engineering. We tackle rigid body protein-protein docking, i.e., computationally predicting the 3D structure of a protein-protein complex from the individual unbound structures, assuming no three-dimensional flexibility during binding. We design a novel pairwise-independent SE(3)-equivariant graph matching network to predict the rotation and translation to place one of the proteins at the right location and the right orientation relative to the second protein. We mathematically guarantee that the predicted complex is always identical regardless of the initial placements of the two structures, avoiding expensive data augmentation. Our model approximates the binding pocket and predicts the docking pose using keypoint matching and alignment through optimal transport and a differentiable Kabsch algorithm. Empirically, we achieve significant running time improvements over existing protein docking software and predict qualitatively plausible protein complex structures despite not using heavy sampling, structure refinement, or templates.
One-sentence Summary: We perform rigid protein docking using a novel independent SE(3)-equivariant message passing mechanism that guarantees the same resulting protein complex independent of the initial placement of the two 3D structures.
MIT researchers created a machine-learning model that can directly predict the complex that will form when two proteins bind together. Their technique is between 80 and 500 times faster than state-of-the-art software methods, and often predicts protein structures that are closer to actual structures that have been observed experimentally.
This technique could help scientists better understand some biological processes that involve protein interactions, like DNA replication and repair; it could also speed up the process of developing new medicines.
“Deep learning is very good at capturing interactions between different proteins that are otherwise difficult for chemists or biologists to write experimentally. Some of these interactions are very complicated, and people haven’t found good ways to express them. This deep-learning model can learn these types of interactions from data,” says Octavian-Eugen Ganea, a postdoc in the MIT Computer Science and Artificial Intelligence Laboratory (CSAIL) and co-lead author of the paper.
Ganea’s co-lead author is Xinyuan Huang, a graduate student at ETH Zurich. MIT co-authors include Regina Barzilay, the School of Engineering Distinguished Professor for AI and Health in CSAIL, and Tommi Jaakkola, the Thomas Siebel Professor of Electrical Engineering in CSAIL and a member of the Institute for Data, Systems, and Society. The research will be presented at the International Conference on Learning Representations.
Significance of the Scientific Development by the @MIT Team
EquiDock wide applicability:
Our method can be integrated end-to-end to boost the quality of other models (see above discussion on runtime importance). Examples are predicting functions of protein complexes [3] or their binding affinity [5], de novo generation of proteins binding to specific targets (e.g., antibodies [6]), modeling back-bone and side-chain flexibility [4], or devising methods for non-binary multimers. See the updated discussion in the “Conclusion” section of our paper.
Advantages over previous methods:
Our method does not rely on templates or heavy candidate sampling [7], aiming at the ambitious goal of predicting the complex pose directly. This should be interpreted in terms of generalization (to unseen structures) and scalability capabilities of docking models, as well as their applicability to various other tasks (discussed above).
Our method obtains a competitive quality without explicitly using previous geometric (e.g., 3D Zernike descriptors [8]) or chemical (e.g., hydrophilic information) features [3]. Future EquiDock extensions would find creative ways to leverage these different signals and, thus, obtain more improvements.
Novelty of theory:
Our work is the first to formalize the notion of pairwise independent SE(3)-equivariance. Previous work (e.g., [9,10]) has incorporated only single object Euclidean-equivariances into deep learning models. For tasks such as docking and binding of biological objects, it is crucial that models understand the concept of multi-independent Euclidean equivariances.
All propositions in Section 3 are our novel theoretical contributions.
We have rewritten the Contribution and Related Work sections to clarify this aspect.
Footnote [a]: We have fixed an important bug in the cross-attention code. We have done a more extensive hyperparameter search and understood that layer normalization is crucial in layers used in Eqs. 5 and 9, but not on the h embeddings as it was originally shown in Eq. 10. We have seen benefits from training our models with a longer patience in the early stopping criteria (30 epochs for DIPS and 150 epochs for DB5). Increasing the learning rate to 2e-4 is important to speed-up training. Using an intersection loss weight of 10 leads to improved results compared to the default of 1.
Bibliography:
[1] Protein-ligand blind docking using QuickVina-W with inter-process spatio-temporal integration, Hassan et al., 2017
[2] GNINA 1.0: molecular docking with deep learning, McNutt et al., 2021
[3] Protein-protein and domain-domain interactions, Kangueane and Nilofer, 2018
[4] Side-chain Packing Using SE(3)-Transformer, Jindal et al., 2022
[5] Contacts-based prediction of binding affinity in protein–protein complexes, Vangone et al., 2015
[6] Iterative refinement graph neural network for antibody sequence-structure co-design, Jin et al., 2021
[7] Hierarchical, rotation-equivariant neural networks to select structural models of protein complexes, Eismann et al, 2020
[8] Protein-protein docking using region-based 3D Zernike descriptors, Venkatraman et al., 2009
[9] SE(3)-transformers: 3D roto-translation equivariant attention networks, Fuchs et al, 2020
[10] E(n) equivariant graph neural networks, Satorras et al., 2021
[11] Fast end-to-end learning on protein surfaces, Sverrisson et al., 2020
Dr. Ahmad Masri and Dr. C Michael Gibson discuss outpatient worsening heart failure in transthyretin amyloid cardiomyopathy in the Attribute-CM trial, May 29, 2026
The following are key points to remember from this update on the treatment of cardiac transthyretin amyloidosis:
Transthyretin (TTR) is a highly conserved protein involved in transportation of thyroxine (T4) and retinol-binding protein. TTR is synthesized mostly by the liver and is rich in beta strands with an intrinsic propensity to aggregate into insoluble amyloid fibers, which deposit within tissue leading to the development of TTR-related amyloidosis (ATTR). ATTR can follow the deposition of either variant TTR (ATTRv, previously known as mutant ATTR) or wild type TTR (ATTRwt).
Cardiac ATTR has a favorable survival rate compared to light chain (AL) amyloidosis, with a median survival of 75 versus 11 months. However, ATTR cardiomyopathy is a progressive disorder but newer therapeutic options include tafamidis (positive phase 3 clinical trial), and possibly patisiran and inotersen.
Inhibition of the Synthesis of Mutated Transthyretin
Liver transplantation removes the source of mutated TTR molecules and prolongs survival, with a 20-year survival of 55.3%. However, tissue accumulation of TTR can continue after liver transplantation because TTR amyloid fibers promote subsequent deposition of ATTRwt. Combined liver–heart transplantation is feasible in younger patients with ATTRv cardiomyopathy and a small series suggests better prognosis than cardiac transplantation.
Inhibition of TTR gene expression: Patisiran is a small interfering RNA blocking the expression of both variant and wt TTR. On the basis of the APOLLO trial, it was approved for therapy of adults with ATTRv-related polyneuropathy both in the United States and European Union. In this trial, patisiran promoted favorable myocardial remodeling based on echocardiographic and N-terminal B-type natriuretic peptide (NT-BNP) changes (this effect was not demonstrated for inotersen) and is still under investigation for tafamidis.
Antisense oligonucleotides inotersen inhibits the production of both variant and wt TTR. Based on the findings of the NEURO-TTR trial, the Food and Drug Administration (FDA) approved this agent for patients with ATTRv-related polyneuropathy. In the NEURO-TTR trial, cardiomyopathy was present in 63%, but the study was not powered to measure effects of inotersen on heart disease. Inotersen can cause thrombocytopenia and must be used cautiously with bleeding risk.
Tetramer Stabilization
Selective stabilizers include tafamidis and AG10. Tafamidis is a benzoxazole and a small molecule that inhibits the dissociation of TTR tetramers by binding the T4-binding sites. The phase ATTR-ACT study showed that when comparing the pooled tafamidis arms (80 and 20 mg) with the placebo arm, tafamidis was associated with lower all-cause mortality than placebo (78 of 264 [29.5%] vs. 76 of 177 [42.9%]; hazard ratio, 0.70; 95% confidence interval, 0.51-0.96) and a lower rate of cardiovascular hospitalizations. Based on the results of the ATTR-ACT trial, it has received Breakthrough Therapy designation from the FDA for treatment of ATTR cardiomyopathy.
Nonselective agents: Diflunisal, a nonsteroidal anti-inflammatory drug, is reported to stabilize TTR tetramers. More studies are needed to confirm its clinical efficacy.
Inhibition of Oligomer Aggregation and Oligomer Disruption
Epigallocatechin gallate is the most abundant catechin in green tea. One single-center open-label 12-month study did not show survival benefits or any change in echocardiographic parameters or NT-BNP compared to baseline.
Degradation and Reabsorption of Amyloid Fibers
Doxycycline-taurosodeoxycholic acid (TUDCA) has been evaluated in two small studies and the results appear to be modest. More data are needed to confirm its efficacy.
Antibodies targeting serum amyloid P protein or amyloid fibrils: Patient enrollment for miridesap followed by anti-SAP antibodies was suspended, and this approach is not being evaluated currently. However, a monoclonal antibody designed to specifically target TTR amyloid deposits (PRX004) has entered clinical evaluation, with an ongoing phase 1 study on ATTRv.
Supportive Treatment of Cardiac Involvement
Drug therapies: Although angiotensin-converting enzyme (ACE) inhibitors/angiotensin-receptor blockers (ARBs) and beta-blockers may have been poorly tolerated in the ATTR-ACT trial, 30% of the patients were on ACE inhibitors/ARBs. There are no data with digoxin in TTR amyloid, and non-dihydropyridine calcium channel blockers are contraindicated due to negative inotropy.
Implantable cardioverter-defibrillators (ICDs): In one study, which included 53 patients with amyloid, ICD shocks occurred exclusively in the AL amyloid group and none in the TTR amyloid patients. Higher defibrillation thresholds and complication rates are of concern.
Cardiac pacing: In a large series of ATTRv-related polyneuropathy (n = 262), a pacemaker was implanted in 110 patients with His ventricular interval >700 ms. The authors recommend that any conduction disturbance on 12-lead electrocardiogram (ECG) warrants further investigation with Holter monitoring to determine candidacy for a pacemaker.
Left ventricular assist device (LVAD): Although an LVAD is technically feasible, it is associated with high short-term mortality and worse outcomes than in dilated cardiomyopathy.
Cardiac transplantation: This is a valuable option for patients with end-stage heart failure when significant extracardiac disease is excluded. In one study with 10 patients, only episodes of amyloid recurrence occurred.
This is an outstanding overview of this topic and recommended reading for anyone who cares for patients with cardiac transthyretin amyloid.
First-Ever Evidence that Patisiran Reduces Pathogenic, Misfolded TTR Monomers and Oligomers in FAP Patients
We reported data from our ongoing Phase 2 open-label extension (OLE) study of patisiran, an investigational RNAi therapeutic targeting transthyretin (TTR) for the treatment of TTR-mediated amyloidosis (ATTR amyloidosis) patients with familial amyloidotic polyneuropathy (FAP). Alnylam scientists and collaborators from The Scripps Research Institute and Misfolding Diagnostics, Inc. were able to measure the effects of patisiran on pathogenic, misfolded TTR monomers and oligomers in FAP patients. Results showed a rapid and sustained reduction in serum non-native conformations of TTR (NNTTR) of approximately 90%. Since NNTTR is pathogenic in ATTR amyloidosis and the level of NNTTR reduction correlated with total TTR knockdown, these results provide direct mechanistic evidence supporting the therapeutic hypothesis that TTR knockdown has the potential to result in clinical benefit. Furthermore, complete 12-month data from all 27 patients that enrolled in the patisiran Phase 2 OLE study showed sustained mean maximum reductions in total serum TTR of 91% for over 18 months and a mean 3.1-point decrease in mNIS+7 at 12 months, which compares favorably to an estimated increase in mNIS+7 of 13 to 18 points at 12 months based upon analysis of historical data sets in untreated FAP patients with similar baseline characteristics. Importantly, patisiran administration continues to be generally well tolerated out to 21 months of treatment.
We are encouraged by these new data that provide continued support for our hypothesis that patisiran has the potential to halt neuropathy progression in patients with FAP. If these results are replicated in a randomized, double-blind, placebo-controlled study, we believe that patisiran could emerge as an important treatment option for patients suffering from this debilitating, progressive and life-threatening disease.
Hereditary ATTR Amyloidosis with Polyneuropathy (hATTR-PN)
ATTR amyloidosis is a progressive, life-threatening disease caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in multiple organs, but primarily in the peripheral nerves and heart. ATTR amyloidosis can lead to significant morbidity, disability, and mortality. The TTR protein is produced primarily in the liver and is normally a carrier for retinol binding protein – one of the vehicles used to transport vitamin A around the body. Mutations in the TTR gene cause misfolding of the protein and the formation of amyloid fibrils that typically contain both mutant and wild-type TTR that deposit in tissues such as the peripheral nerves and heart, resulting in intractable peripheral sensory neuropathy, autonomic neuropathy, and/or cardiomyopathy.
ATTR represents a major unmet medical need with significant morbidity and mortality. There are over 100 reported TTR mutations; the particular TTR mutation and the site of amyloid deposition determine the clinical manifestations of the disease whether it is predominantly symptoms of neuropathy or cardiomyopathy.
Specifically, hereditary ATTR amyloidosis with polyneuropathy (hATTR-PN), also known as familial amyloidotic polyneuropathy (FAP), is an inherited, progressive disease leading to death within 5 to 15 years. It is due to a mutation in the transthyretin (TTR) gene, which causes misfolded TTR proteins to accumulate as amyloid fibrils predominantly in peripheral nerves and other organs. hATTR-PN can cause sensory, motor, and autonomic dysfunction, resulting in significant disability and death.
It is estimated that hATTR-PN, also known as FAP, affects approximately 10,000 people worldwide. Patients have a life expectancy of 5 to 15 years from symptom onset, and the only treatment options for early stage disease are liver transplantation and TTR stabilizers such as tafamidis (approved in Europe) and diflunisal. Unfortunately liver transplantation has limitations, including limited organ availability as well as substantial morbidity and mortality. Furthermore, transplantation eliminates the production of mutant TTR but does not affect wild-type TTR, which can further deposit after transplantation, leading to cardiomyopathy and worsening of neuropathy. There is a significant need for novel therapeutics to treat patients who have inherited mutations in the TTR gene.
Our ATTR program is the lead effort in our Genetic Medicine Strategic Therapeutic Area (STAr) product development and commercialization strategy, which is focused on advancing innovative RNAi therapeutics toward genetically defined targets for the treatment of rare diseases with high unmet medical need. We are developing patisiran (ALN-TTR02), an intravenously administered RNAi therapeutic, to treat the hATTR-PN form of the disease.
Patisiran for the Treatment hATTR-PN
APOLLO Phase 3 Trial
In 2012, Alnylam entered into an exclusive alliance with Genzyme, a Sanofi company, to develop and commercialize RNAi therapeutics, including patisiran and revusiran, for the treatment of ATTR amyloidosis in Japan and the broader Asian-Pacific region. In early 2014, this relationship was extended as a significantly broader alliance to advance RNAi therapeutics as genetic medicines. Under this new agreement, Alnylam will lead development and commercialization of patisiran in North America and Europe while Genzyme will develop and commercialize the product in the rest of world.
Hereditary ATTR Amyloidosis with Cardiomyopathy (hATTR-CM)
ATTR amyloidosis is a progressive, life-threatening disease caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in multiple organs, but primarily in the peripheral nerves and heart. ATTR amyloidosis can lead to significant morbidity, disability, and mortality. The TTR protein is produced primarily in the liver and is normally a carrier for retinol binding protein – one of the vehicles used to transport vitamin A around the body. Mutations in the TTR gene cause misfolding of the protein and the formation of amyloid fibrils that typically contain both mutant and wild-type TTR that deposit in tissues such as the peripheral nerves and heart, resulting in intractable peripheral sensory neuropathy, autonomic neuropathy, and/or cardiomyopathy.
ATTR represents a major unmet medical need with significant morbidity and mortality. There are over 100 reported TTR mutations; the particular TTR mutation and the site of amyloid deposition determine the clinical manifestations of the disease, whether it is predominantly symptoms of neuropathy or cardiomyopathy.
Specifically, hereditary ATTR amyloidosis with cardiomyopathy (hATTR-CM), also known as familial amyloidotic cardiomyopathy (FAC), is an inherited, progressive disease leading to death within 2 to 5 years. It is due to a mutation in the transthyretin (TTR) gene, which causes misfolded TTR proteins to accumulate as amyloid fibrils primarily in the heart. Hereditary ATTR amyloidosis with cardiomyopathy can result in heart failure and death.
While the exact numbers are not known, it is estimated hATTR-CM, also known as FAC affects at least 40,000 people worldwide. hATTR-CM is fatal within 2 to 5 years of diagnosis and treatment is currently limited to supportive care. Wild-type ATTR amyloidosis (wtATTR amyloidosis), also known as senile systemic amyloidosis, is a nonhereditary, progressive disease leading to death within 2 to 5 years. It is caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in the heart. Wild-type ATTR amyloidosis can cause cardiomyopathy and result in heart failure and death. There are no approved therapies for the treatment of hATTR-CM or SSA; hence there is a significant unmet need for novel therapeutics to treat these patients.
Our ATTR program is the lead effort in our Genetic Medicine Strategic Therapeutic Area (STAr) product development and commercialization strategy, which is focused on advancing innovative RNAi therapeutics toward genetically defined targets for the treatment of rare diseases with high unmet medical need. We are developing revusiran (ALN-TTRsc), a subcutaneously administered RNAi therapeutic for the treatment of hATTR-CM.
Revusiran for the Treatment of hATTR-CM
ENDEAVOUR Phase 3 Trial
In 2012, Alnylam entered into an exclusive alliance with Genzyme, a Sanofi company, to develop and commercialize RNAi therapeutics, including patisiran and revusiran, for the treatment of ATTR amyloidosis in Japan and the broader Asian-Pacific region. In early 2014, this relationship was extended as a broader alliance to advance RNAi therapeutics as genetic medicines. Under this new agreement, Alnylam and Genzyme have agreed to co-develop and co-commercialize revusiran in North America and Europe, with Genzyme developing and commercializing the product in the rest of world. This broadened relationship on revusiran is aimed at expanding and accelerating the product’s global value.
We presented pre-clinical data with ALN-TTRsc02, an investigational RNAi therapeutic targeting transthyretin (TTR) for the treatment of TTR-mediated amyloidosis (ATTR amyloidosis). In pre-clinical studies, including those in non-human primates (NHPs), ALN-TTRsc02 achieved potent and highly durable knockdown of serum TTR of up to 99% with multi-month durability achieved after just a single dose, supportive of a potentially once quarterly dose regimen. Results from studies comparing TTR knockdown activity of ALN-TTRsc02 to that of revusiran showed that ALN-TTRsc02 has a markedly superior TTR knockdown profile. Further, in initial rat toxicology studies, ALN-TTRsc02 was found to be generally well tolerated with no significant adverse events at doses as high as 100 mg/kg.
Heart failure with a preserved ejection fraction (HFPEF) is a clinical syndrome that has no pharmacologic therapies approved for this use to date. In light of failed medicines, cardiologists have refocused treatment strategies based on the theory that HFPEF is a heterogeneous clinical syndrome with different etiologies. Classification of HFPEF according to etiologic subtype may, therefore, identify cohorts with treatable pathophysiologic mechanisms and may ultimately pave the way forward for developing meaningful HFPEF therapies.1
A wealth of data now indicates that amyloid infiltration is an important mechanism underlying HFPEF. Inherited mutations in transthyretin cardiac amyloidosis (ATTRm) or the aging process in wild-type disease (ATTRwt) cause destabilization of the transthyretin (TTR) protein into monomers or oligomers, which aggregate into amyloid fibrils. These insoluble fibrils accumulate in the myocardium and result in diastolic dysfunction, restrictive cardiomyopathy, and eventual congestive heart failure (Figure 1). In an autopsy study of HFPEF patients, almost 20% without antemortem suspicion of amyloid had left ventricular (LV) TTR amyloid deposition.2 Even more resounding evidence for the contribution of TTR amyloid to HFPEF was a study in which 120 hospitalized HFPEF patients with LV wall thickness ≥12 mm underwent technetium-99m 3,3-diphosphono-1,2-propranodicarboxylic acid (99mTc-DPD) cardiac imaging,3,4 a bone isotope known to have high sensitivity and specificity for diagnosing TTR cardiac amyloidosis.5,6 Moderate-to-severe myocardial uptake indicative of TTR cardiac amyloid deposition was detected in 13.3% of HFPEF patients who did not have TTR gene mutations. Therefore, TTR cardiac amyloid deposition, especially in older adults, is not rare, can be easily identified, and may contribute to the underlying pathophysiology of HFPEF.
Figure 1
As no U.S. Food and Drug Administration-approved drugs are currently available for the treatment of HFPEF or TTR cardiac amyloidosis, the development of medications that attenuate or prevent TTR-mediated organ toxicity has emerged as an important therapeutic goal. Over the past decade, a host of therapies and therapeutic drug classes have emerged in clinical trials (Table 1), and these may herald a new direction for treating HFPEF secondary to TTR amyloid.
Table 1
TTR Silencers (siRNA and Antisense Oligonucleotides)
siRNA
Ribonucleic acid interference (RNAi) has surfaced as an endogenous cellular mechanism for controlling gene expression. Small interfering RNAs (siRNAs) delivered into cells can disrupt the production of target proteins.7,8 A formulation of lipid nanoparticle and triantennary N-acetylgalactosamine (GalNAc) conjugate that delivers siRNAs to hepatocytes is currently in clinical trials.9 Prior research demonstrated these GalNAc-siRNA conjugates result in robust and durable knockdown of a variety of hepatocyte targets across multiple species and appear to be well suited for suppression of TTR gene expression and subsequent TTR protein production.
The TTR siRNA conjugated to GalNAc, ALN-TTRSc, is now under active investigation as a subcutaneous injection in phase 3 clinical trials in patients with TTR cardiac amyloidosis.10 Prior phase 2 results demonstrated that ALN-TTRSc was generally well tolerated in patients with significant TTR disease burden and that it reduced both wild-type and mutant TTR gene expression by a mean of 87%. Harnessing RNAi technology appears to hold great promise for treating patients with TTR cardiac amyloidosis. The ability of ALN-TTRSc to lower both wild-type and mutant proteins may provide a major advantage over liver transplantation, which affects the production of only mutant protein and is further limited by donor shortage, cost, and need for immunosuppression.
Antisense Oligonucleotides
Antisense oligonucleotides (ASOs) are under clinical investigation for their ability to inhibit hepatic expression of amyloidogenic TTR protein. Currently, the ASO compound, ISIS-TTRRx, is under investigation in a phase 3 multicenter, randomized, double-blind, placebo-controlled clinical trial in patients with familial amyloid polyneuropathy (FAP).11 The primary objective is to evaluate its efficacy as measured by change in neuropathy from baseline relative to placebo. Secondary measures will evaluate quality of life (QOL), modified body mass index (mBMI) by albumin, and pharmacodynamic effects on retinol binding protein. Exploratory objectives in a subset of patients with LV wall thickness ≥13 mm without a history of persistent hypertension will examine echocardiographic parameters, N-terminal pro–B-type natriuretic peptide (NT-proBNP), and polyneuropathy disability score relative to placebo. These data will facilitate analysis of the effect of antisense oligonucleotide-mediated TTR suppression on the TTR cardiac phenotype with a phase 3 trial anticipated to begin enrollment in 2016.
TTR Stabilizers (Diflunisal, Tafamidis)
Diflunisal
Several TTR-stabilizing agents are in various stages of clinical trials. Diflunisal, a traditionally used and generically available nonsteroidal anti-inflammatory drug (NSAID), binds and stabilizes familial TTR variants against acid-mediated fibril formation in vitro and is now in human clinical trials.12,13 The use of diflunisal in patients with TTR cardiac amyloidosis is controversial given complication of chronic inhibition of cyclooxygenase (COX) enzymes, including gastrointestinal bleeding, renal dysfunction, fluid retention, and hypertension that may precipitate or exacerbate heart failure in vulnerable individuals.14-17 In TTR cardiac amyloidosis, an open-label cohort study suggested that low-dose diflunisal with careful monitoring along with a prophylactic proton pump inhibitor could be safely administered to compensated patients.18 An association was observed, however, between chronic diflunisal use and adverse changes in renal function suggesting that advanced kidney disease may be prohibitive in diflunisal therapy.In FAP patients with peripheral or autonomic neuropathy randomized to diflunisal or placebo, diflunisal slowed progression of neurologic impairment and preserved QOL over two years of follow-up.19 Echocardiography demonstrated cardiac involvement in approximately 50% of patients.20 Longer-term safety and efficacy data over an average 38 ± 31 months in 40 Japanese patients with hereditary ATTR amyloidosis who were not candidates for liver transplantation showed that diflunisal was mostly well tolerated.12 The authors cautioned the need for attentive monitoring of renal function and blood cell counts. Larger multicenter collaborations are needed to determine diflunisal’s true efficacy in HFPEF patients with TTR cardiac amyloidosis.
Tafamidis
Tafamidis is under active investigation as a novel compound that binds to the thyroxine-binding sites of the TTR tetramer, inhibiting its dissociation into monomers and blocking the rate-limiting step in the TTR amyloidogenesis cascade.21 The TTR compound was shown in an 18-month double-blind, placebo-controlled trial to slow progression of neurologic symptoms in patients with early-stage ATTRm due to the V30M mutation.22 When focusing on cardiomyopathy in a phase 2, open-label trial, tafamidis also appeared to effectively stabilize TTR tetramers in non-V30M variants, wild-type and V122I, as well as biochemical and echocardiographic parameters.23,24 Preliminary data suggests that clinically stabilized patients had shorter disease duration, lower cardiac biomarkers, less myocardial thickening, and higher EF than those who were not stabilized, suggesting early institution of therapy may be beneficial. A phase 3 trial has completed enrollment and will evaluate the efficacy, safety, and tolerability of tafamidis 20 or 80 mg orally vs. placebo.25 This will contribute to long-term safety and efficacy data needed to determine the therapeutic effects of tafamidis among ATTRm variants.
Amyloid Degraders (Doxycycline/TUDCA and Anti-SAP Antibodies)
Doxycycline/TUDCA
While silencer and stabilizer drugs are aimed at lowering amyloidogenic precursor protein production, they cannot remove already deposited fibrils in an infiltrated heart. Removal of already deposited fibrils by amyloid degraders would be an important therapeutic strategy, particularly in older adults with heavily infiltrated hearts reflected by thick walls, HFPEF, systolic heart failure, and restrictive cardiomyopathy. Combined doxycycline and tauroursodeoxycholic acid (TUDCA) disrupt TTR amyloid fibrils and appeared to have an acceptable safety profile in a small phase 2 open-label study among 20 TTR patients. No serious adverse reactions or clinical progression of cardiac or neuropathic involvement was observed over one year.26 An active phase 2, single-center, open-label, 12-month study will assess primary outcome measures including mBMI, neurologic impairment score, and NT-proBNP.27 Another phase 2 study is examining the tolerability and efficacy of doxycycline/TUDCA over an 18-month period in patients with TTR amyloid cardiomyopathy.28 Additionally, a study in patients with TTR amyloidosis is ongoing to determine the effect of doxycycline alone on neurologic function, cardiac biomarkers, echocardiographic parameters, modified body mass index, and autonomic neuropathy.29
Anti-SAP Antibodies
In order to safely clear established amyloid deposits, the role of the normal, nonfibrillar plasma glycoprotein present in all human amyloid deposits, serum amyloid P component (SAP), needs to be more clearly understood.30 In mice with amyloid AA type deposits, administration of antihuman SAP antibody triggered a potent giant cell reaction that removed massive visceral amyloid deposits without adverse effects.31 In humans with TTR cardiac amyloidosis, anti-SAP antibody treatments could be feasible because the bis-D proline compound, CPHPC, is capable of clearing circulating human SAP, which allow anti-SAP antibodies to reach residual deposited SAP. In a small, open-label, single-dose-escalation, phase 1 trial involving 15 patients with systemic amyloidosis, none of whom had clinical evidence of cardiac amyloidosis, were treated with CPHPC followed by human monoclonal IgG1 anti-SAP antibody.32 No serious adverse events were reported and amyloid deposits were cleared from the liver, kidney, and lymph node. Anti-SAP antibodies hold promise as a potential amyloid therapy because of their potential to target all forms of amyloid deposits across multiple tissue types.
Mutant or wild-type TTR cardiac amyloidoses are increasingly recognized as a cause of HFPEF. Clinicians need to be aware of this important HFPEF etiology because the diverse array of emerging disease-modifying agents for TTR cardiac amyloidosis in human clinical trials has the potential to herald a new era for the treatment of HFPEF.
References
Maurer MS, Mancini D. HFpEF: is splitting into distinct phenotypes by comorbidities the pathway forward? J Am Coll Cardiol 2014;64:550-2.
Mohammed SF, Mirzoyev SA, Edwards WD, et al. Left ventricular amyloid deposition in patients with heart failure and preserved ejection fraction. JACC Heart Fail 2014;2:113-22.
González-López E, Gallego-Delgado M, Guzzo-Merello G, et al. Wild-type transthyretin amyloidosis as a cause of heart failure with preserved ejection fraction. Eur Heart J 2015.
Castano A, Bokhari S, Maurer MS. Unveiling wild-type transthyretin cardiac amyloidosis as a significant and potentially modifiable cause of heart failure with preserved ejection fraction. Eur Heart J 2015 Jul 28. [Epub ahead of print]
Rapezzi C, Merlini G, Quarta CC, et al. Systemic cardiac amyloidoses: disease profiles and clinical courses of the 3 main types. Circulation 2009;120:1203-12.
Bokhari S, Castano A, Pozniakoff T, Deslisle S, Latif F, Maurer MS. (99m)Tc-pyrophosphate scintigraphy for differentiating light-chain cardiac amyloidosis from the transthyretin-related familial and senile cardiac amyloidoses. Circ Cardiovasc Imaging 2013;6:195-201.
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998;391:806-11.
Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001;411:494-8.
Kanasty R, Dorkin JR, Vegas A, Anderson D. Delivery materials for siRNA therapeutics. Nature Mater 2013;12:967-77.
U.S. National Institutes of Health. Phase 2 Study to Evaluate ALN-TTRSC in Patients With Transthyretin (TTR) Cardiac Amyloidosis (ClinicalTrials.gov website). 2014. Available at: https://www.clinicaltrials.gov/ct2/show/NCT01981837. Accessed 8/19/2015.
U.S. National Institutes of Health. Efficacy and Safety of ISIS-TTRRx in Familial Amyloid Polyneuropathy (Clinical Trials.gov Website. 2013. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01737398. Accessed 8/19/2015.
Sekijima Y, Dendle MA, Kelly JW. Orally administered diflunisal stabilizes transthyretin against dissociation required for amyloidogenesis. Amyloid 2006;13:236-49.
Tojo K, Sekijima Y, Kelly JW, Ikeda S. Diflunisal stabilizes familial amyloid polyneuropathy-associated transthyretin variant tetramers in serum against dissociation required for amyloidogenesis. Neurosci Res 2006;56:441-9.
Epstein M. Non-steroidal anti-inflammatory drugs and the continuum of renal dysfunction. J Hypertens Suppl 2002;20:S17-23.
Wallace JL. Pathogenesis of NSAID-induced gastroduodenal mucosal injury. Best Pract Res Clin Gastroenterol 2001;15:691-703.
Mukherjee D, Nissen SE, Topol EJ. Risk of cardiovascular events associated with selective COX-2 inhibitors. JAMA 2001;286:954-9.
Page J, Henry D. Consumption of NSAIDs and the development of congestive heart failure in elderly patients: an underrecognized public health problem. Arch Intern Med 2000;160:777-84.
Castano A, Helmke S, Alvarez J, Delisle S, Maurer MS. Diflunisal for ATTR cardiac amyloidosis. Congest Heart Fail 2012;18:315-9.
Berk JL, Suhr OB, Obici L, et al. Repurposing diflunisal for familial amyloid polyneuropathy: a randomized clinical trial. JAMA 2013;310:2658-67.
Quarta CCF, Solomon RH Suhr SD, et al. The prevalence of cardiac amyloidosis in familial amyloidotic polyneuropathy with predominant neuropathy: The Diflunisal Trial. International Symposium on Amyloidosis 2014:88-9.
Hammarstrom P, Jiang X, Hurshman AR, Powers ET, Kelly JW. Sequence-dependent denaturation energetics: A major determinant in amyloid disease diversity. Proc Natl Acad Sci U S A 2002;99 Suppl 4:16427-32.
Coelho T, Maia LF, Martins da Silva A, et al. Tafamidis for transthyretin familial amyloid polyneuropathy: a randomized, controlled trial. Neurology 2012;79:785-92.
Merlini G, Plante-Bordeneuve V, Judge DP, et al. Effects of tafamidis on transthyretin stabilization and clinical outcomes in patients with non-Val30Met transthyretin amyloidosis. J Cardiovasc Transl Res 2013;6:1011-20.
Maurer MS, Grogan DR, Judge DP, et al. Tafamidis in transthyretin amyloid cardiomyopathy: effects on transthyretin stabilization and clinical outcomes. Circ Heart Fail 2015;8:519-26.
U.S. National Institutes of Health. Safety and Efficacy of Tafamidis in Patients With Transthyretin Cardiomyopathy (ATTR-ACT) (ClinicalTrials.gov website). 2014. Available at: http://www.clinicaltrials.gov/show/NCT01994889. Accessed 8/19/2015.
Obici L, Cortese A, Lozza A, et al. Doxycycline plus tauroursodeoxycholic acid for transthyretin amyloidosis: a phase II study. Amyloid 2012;19 Suppl 1:34-6.
U.S. National Institutes of Health. Safety, Efficacy and Pharmacokinetics of Doxycycline Plus Tauroursodeoxycholic Acid in Transthyretin Amyloidosis (ClinicalTrials.gov website). 2011. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01171859. Accessed 8/19/2015.
U.S. National Institutes of Health. Tolerability and Efficacy of a Combination of Doxycycline and TUDCA in Patients With Transthyretin Amyloid Cardiomyopathy (ClinicalTrials.gov website). 2013. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01855360. Accessed 8/19/2015.
U.S. National Institutes of Health. Safety and Effect of Doxycycline in Patients With Amyloidosis (ClinicalTrials.gov website).2015. Available at: https://clinicaltrials.gov/ct2/show/NCT01677286. Accessed 8/19/2015.
Pepys MB, Dash AC. Isolation of amyloid P component (protein AP) from normal serum as a calcium-dependent binding protein. Lancet 1977;1:1029-31.
Bodin K, Ellmerich S, Kahan MC, et al. Antibodies to human serum amyloid P component eliminate visceral amyloid deposits. Nature 2010;468:93-7.
Richards DB, Cookson LM, Berges AC, et al. Therapeutic Clearance of Amyloid by Antibodies to Serum Amyloid P Component. N Engl J Med 2015;373:1106-14.
…
The Acid-Mediated Denaturation Pathway of Transthyretin Yields a Conformational Intermediate That Can Self-Assemble into Amyloid†
Transthyretin (TTR) amyloid fibril formation is observed during partial acid denaturation and while refolding acid-denatured TTR, implying that amyloid fibril formation results from the self-assembly of a conformational intermediate. The acid denaturation pathway of TTR has been studied in detail herein employing a variety of biophysical methods to characterize the intermediate(s) capable of amyloid fibril formation. At physiological concentrations, tetrameric TTR remains associated from pH 7 to pH 5 and is incapable of amyloid fibril formation. Tetrameric TTR dissociates to a monomer in a process that is dependent on both pH and protein concentration below pH 5. The extent of amyloid fibril formation correlates with the concentration of the TTR monomer having an altered, but defined, tertiary structure over the pH range of 5.0−3.9. The inherent Trp fluorescence-monitored denaturation curve of TTR exhibits a plateau over the pH range where amyloid fibril formation is observed (albeit at a higher concentration), implying that a steady-state concentration of the amyloidogenic intermediate with an altered tertiary structure is being detected. Interestingly, 1-anilino-8-naphthalenesulfonate fluorescence is at a minimum at the pH associated with maximal amyloid fibril formation (pH 4.4), implying that the amyloidogenic intermediate does not have a high extent of hydrophobic surface area exposed, consistent with a defined tertiary structure. Transthyretin has two Trp residues in its primary structure, Trp-41 and Trp-79, which are conveniently located far apart in the tertiary structure of TTR. Replacement of each Trp with Phe affords two single Trp containing variants which were used to probe local pH-dependent tertiary structural changes proximal to these chromophores. The pH-dependent fluorescence behavior of the Trp-79-Phe mutant strongly suggests that Trp-41 is located near the site of the tertiary structural rearrangement that occurs in the formation of the monomeric amyloidogenic intermediate, likely involving the C-strand−loop−D-strand region. Upon further acidification of TTR (below pH 4.4), the structurally defined monomeric amyloidogenic intermediate begins to adopt alternative conformations that are not amyloidogenic, ultimately forming an A-state conformation below pH 3 which is also not amyloidogenic. In summary, analytical equilibrium ultracentrifugation, SDS−PAGE, far- and near-UV CD, fluorescence, and light scattering studies suggest that the amyloidogenic intermediate is a monomeric predominantly β-sheet structure having a well-defined tertiary structure.
Prevention of Transthyretin Amyloid Disease by Changing Protein Misfolding Energetics
Per Hammarström*, R. Luke Wiseman*, Evan T. Powers, Jeffery W. Kelly†+ Author Affiliations
Genetic evidence suggests that inhibition of amyloid fibril formation by small molecules should be effective against amyloid diseases. Known amyloid inhibitors appear to function by shifting the aggregation equilibrium away from the amyloid state. Here, we describe a series of transthyretin amyloidosis inhibitors that functioned by increasing the kinetic barrier associated with misfolding, preventing amyloidogenesis by stabilizing the native state. The trans-suppressor mutation, threonine 119 → methionine 119, which is known to ameliorate familial amyloid disease, also functioned through kinetic stabilization, implying that this small-molecule strategy should be effective in treating amyloid diseases.
Rational design of potent human transthyretin amyloid disease inhibitors
Thomas Klabunde1,2, H. Michael Petrassi3, Vibha B. Oza3, Prakash Raman3, Jeffery W. Kelly3 & James C. Sacchettini1
The human amyloid disorders, familial amyloid polyneuropathy, familial amyloid cardiomyopathy and senile systemic amyloidosis, are caused by insoluble transthyretin (TTR) fibrils, which deposit in the peripheral nerves and heart tissue. Several nonsteroidal anti-inflammatory drugs and structurally similar compounds have been found to strongly inhibit the formation of TTR amyloid fibrils in vitro. These include flufenamic acid, diclofenac, flurbiprofen, and resveratrol. Crystal structures of the protein–drug complexes have been determined to allow detailed analyses of the protein–drug interactions that stabilize the native tetrameric conformation of TTR and inhibit the formation of amyloidogenic TTR. Using a structure-based drug design approach ortho-trifluormethylphenyl anthranilic acid and N-(meta-trifluoromethylphenyl) phenoxazine 4,6-dicarboxylic acid have been discovered to be very potent and specific TTR fibril formation inhibitors. This research provides a rationale for a chemotherapeutic approach for the treatment of TTR-associated amyloid diseases.
First European consensus for diagnosis, management, and treatment of transthyretin familial amyloid polyneuropathy
Adams, Davida; Suhr, Ole B.b; Hund, Ernstc; Obici, Laurad; Tournev, Ivailoe,f; Campistol, Josep M.g; Slama, Michel S.h; Hazenberg, Bouke P.i; Coelho, Teresaj; from the European Network for TTR-FAP (ATTReuNET)
Purpose of review: Early and accurate diagnosis of transthyretin familial amyloid polyneuropathy (TTR-FAP) represents one of the major challenges faced by physicians when caring for patients with idiopathic progressive neuropathy. There is little consensus in diagnostic and management approaches across Europe.
Recent findings: The low prevalence of TTR-FAP across Europe and the high variation in both genotype and phenotypic expression of the disease means that recognizing symptoms can be difficult outside of a specialized diagnostic environment. The resulting delay in diagnosis and the possibility of misdiagnosis can misguide clinical decision-making and negatively impact subsequent treatment approaches and outcomes.
Summary: This review summarizes the findings from two meetings of the European Network for TTR-FAP (ATTReuNET). This is an emerging group comprising representatives from 10 European countries with expertise in the diagnosis and management of TTR-FAP, including nine National Reference Centres. The current review presents management strategies and a consensus on the gold standard for diagnosis of TTR-FAP as well as a structured approach to ongoing multidisciplinary care for the patient. Greater communication, not just between members of an individual patient’s treatment team, but also between regional and national centres of expertise, is the key to the effective management of TTR-FAP.
Transthyretin familial amyloid polyneuropathy (TTR-FAP) is a highly debilitating and irreversible neurological disorder presenting symptoms of progressive sensorimotor and autonomic neuropathy [1▪,2▪,3]. TTR-FAP is caused by misfolding of the transthyretin (TTR) protein leading to protein aggregation and the formation of amyloid fibrils and, ultimately, to amyloidosis (commonly in the peripheral and autonomic nervous system and the heart) [4,5]. TTR-FAP usually proves fatal within 7–12 years from the onset of symptoms, most often due to cardiac dysfunction, infection, or cachexia [6,7▪▪].
The prevalence and disease presentation of TTR-FAP vary widely within Europe. In endemic regions (northern Portugal, Sweden, Cyprus, and Majorca), patients tend to present with a distinct genotype in large concentrations, predominantly a Val30Met substitution in the TTR gene [8–10]. In other areas of Europe, the genetic footprint of TTR-FAP is more varied, with less typical phenotypic expression [6,11]. For these sporadic or scattered cases, a lack of awareness among physicians of variable clinical features and limited access to diagnostic tools (i.e., pathological studies and genetic screening) can contribute to high rates of misdiagnosis and poorer patient outcomes [1▪,11]. In general, early and late-onset variants of TTR-FAP, found within endemic and nonendemic regions, present several additional diagnostic challenges [11,12,13▪,14].
Delay in the time to diagnosis is a major obstacle to the optimal management of TTR-FAP. With the exception of those with a clearly diagnosed familial history of FAP, patients still invariably wait several years between the emergence of first clinical signs and accurate diagnosis [6,11,14]. The timely initiation of appropriate treatment is particularly pertinent, given the rapidity and irreversibility with which TTR-FAP can progress if left unchecked, as well as the limited effectiveness of available treatments during the later stages of the disease [14]. This review aims to consolidate the existing literature and present an update of the best practices in the management of TTR-FAP in Europe. A summary of the methods used to achieve a TTR-FAP diagnosis is presented, as well as a review of available treatments and recommendations for treatment according to disease status.
Patients with TTR-FAP can present with a range of symptoms [11], and care should be taken to acquire a thorough clinical history of the patient as well as a family history of genetic disease. Delay in diagnosis is most pronounced in areas where TTR-FAP is not endemic or when there is no positive family history [1▪]. TTR-FAP and TTR-familial amyloid cardiomyopathy (TTR-FAC) are the two prototypic clinical disease manifestations of a broader disease spectrum caused by an underlying hereditary ATTR amyloidosis [19]. In TTR-FAP, the disease manifestation of neuropathy is most prominent and definitive for diagnosis, whereas cardiomyopathy often suggests TTR-FAC. However, this distinction is often superficial because cardiomyopathy, autonomic neuropathy, vitreous opacities, kidney disease, and meningeal involvement all may be present with varying severity for each patient with TTR-FAP.
Among early onset TTR-FAP with usually positive family history, symptoms of polyneuropathy present early in the disease process and usually predominate throughout the progression of the disease, making neurological testing an important diagnostic aid [14]. Careful clinical examination (e.g., electromyography with nerve conduction studies and sympathetic skin response, quantitative sensation test, quantitative autonomic test) can be used to detect, characterize, and scale the severity of neuropathic abnormalities involving small and large nerve fibres [10]. Although a patient cannot be diagnosed definitively with TTR-FAP on the basis of clinical presentation alone, symptoms suggesting the early signs of peripheral neuropathy, autonomic dysfunction, and cardiac conduction disorders or infiltrative cardiomyopathy are all indicators that further TTR-FAP diagnostic investigation is warranted. Late-onset TTR-FAP often presents as sporadic cases with distinct clinical features (e.g., milder autonomic dysfunction) and can be more difficult to diagnose than early-onset TTR-FAP (Table 2) [1▪,11,12,13▪,14,20].
Genetic testing is carried out to allow detection of specific amyloidogenic TTR mutations (Table 1), using varied techniques depending on the expertise and facilities available in each country (Table S2, http://links.lww.com/CONR/A39). A targeted approach to detect a specific mutation can be used for cases belonging to families with previous diagnosis. In index cases of either endemic and nonendemic regions that do not have a family history of disease, are difficult to confirm, and have atypical symptoms, TTR gene sequencing is required for the detection of both predicted and new amyloidogenic mutations [26,27].
Following diagnosis, the neuropathy stage and systemic extension of the disease should be determined in order to guide the next course of treatment (Table 4) [3,30,31]. The three stages of TTR-FAP severity are graded according to a patient’s walking disability and degree of assistance required [30]. Systemic assessment, especially of the heart, eyes, and kidney, is also essential to ensure all aspects of potential impact of the disease can be detected [10].
The goals of cardiac investigations are to detect serious conduction disorders with the risk of sudden death and infiltrative cardiomyopathy. Electrocardiograms (ECG), Holter-ECG, and intracardiac electrophysiology study are helpful to detect conduction disorders. Echocardiograms, cardiac magnetic resonance imaging, scintigraphy with bone tracers, and biomarkers (e.g., brain natriuretic peptide, troponin) can all help to diagnose infiltrative cardiomyopathy[10]. An early detection of cardiac abnormalities has obvious benefits to the patient, given that the prophylactic implantation of pacemakers was found to prevent 25% of major cardiac events in TTR-FAP patients followed up over an average of 4 years [32▪▪]. Assessment of cardiac denervation with 123-iodine meta-iodobenzylguanidine is a powerful prognostic marker in patients diagnosed with FAP [33].
…..
Tafamidis
Tafamidis is a first-in-class therapy that slows the progression of TTR amyloidogenesis by stabilizing the mutant TTR tetramer, thereby preventing its dissociation into monomers and amyloidogenic and toxic intermediates [55,56]. Tafamidis is currently indicated in Europe for the treatment of TTR amyloidosis in adult patients with stage I symptomatic polyneuropathy to delay peripheral neurological impairment [57].
In an 18-month, double-blind, placebo-controlled study of patients with early-onset Val30Met TTR-FAP, tafamidis was associated with a 52% lower reduction in neurological deterioration (P = 0.027), a preservation of nerve function, and TTR stabilization versus placebo [58▪▪]. However, only numerical differences were found for the coprimary endpoints of neuropathy impairment [neuropathy impairment score in the lower limb (NIS-LL) responder rates of 45.3% tafamidis vs 29.5% placebo; P = 0.068] and quality of life scores [58▪▪]. A 12-month, open-label extension study showed that the reduced rates of neurological deterioration associated with tafamidis were sustained over 30 months, with earlier initiation of tafamidis linking to better patient outcomes (P = 0.0435) [59▪]. The disease-slowing effects of tafamidis may be dependent on the early initiation of treatment. In an open-label study with Val30Met TTR-FAP patients with late-onset and advanced disease (NIS-LL score >10, mean age 56.4 years), NIS-LL and disability scores showed disease progression despite 12 months of treatment with tafamidis, marked by a worsening of neuropathy stage in 20% and the onset of orthostatic hypotension in 22% of patients at follow-up [60▪].
Tafamidis is not only effective in patients exhibiting the Val30Met mutation; it also has proven efficacy, in terms of TTR stabilization, in non-Val30Met patients over 12 months [61]. Although tafamidis has demonstrated safe use in patients with TTR-FAP, care should be exercised when prescribing to those with existing digestive problems (e.g., diarrhoea, faecal incontinence) [60▪].
Diflunisal is a nonsteroidal anti-inflammatory drug (NSAID) that, similar to tafamidis, slows the rate of amyloidogenesis by preventing the dissociation, misfolding, and misassembly of the mutated TTR tetramer [62,63]. Off-label use has been reported for patients with stage I and II disease, although diflunisal is not currently licensed for the treatment of TTR-FAP.
Evidence for the clinical effectiveness of diflunisal in TTR-FAP derives from a placebo-controlled, double-blind, 24-month study in 130 patients with clinically detectable peripheral or autonomic neuropathy[64▪]. The deterioration in NIS scores was significantly more pronounced in patients receiving placebo compared with those taking diflunisal (P = 0.001), and physical quality of life measures showed significant improvement among diflunisal-treated patients (P = 0.001). Notable during this study was the high rate of attrition in the placebo group, with 50% more placebo-treated patients dropping out of this 2-year study as a result of disease progression, advanced stage of the disease, and varied mutations.
One retrospective analysis of off-label use of diflunisal in patients with TTR-FAP reported treatment discontinuation in 57% of patients because of adverse events that were largely gastrointestinal [65]. Conclusions on the safety of diflunisal in TTR-FAP will depend on further investigations on the impact of known cardiovascular and renal side-effects associated with the NSAID drug class [66,67].
Disease related changes in proteomics, protein folding, protein-protein interaction, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 1: Next Generation Sequencing (NGS)
Disease related changes in proteomics, protein folding, protein-protein interaction
Curator: Larry H. Bernstein, MD, FCAP
LPBI
Frankenstein Proteins Stitched Together by Scientists
The Frankenstein monster, stitched together from disparate body parts, proved to be an abomination, but stitched together proteins may fare better. They may, for example, serve specific purposes in medicine, research, and industry. At least, that’s the ambition of scientists based at the University of North Carolina. They have developed a computational protocol called SEWING that builds new proteins from connected or disconnected pieces of existing structures. [Wikipedia]
Unlike Victor Frankenstein, who betrayed Promethean ambition when he sewed together his infamous creature, today’s biochemists are relatively modest. Rather than defy nature, they emulate it. For example, at the University of North Carolina (UNC), researchers have taken inspiration from natural evolutionary mechanisms to develop a technique called SEWING—Structure Extension With Native-substructure Graphs. SEWING is a computational protocol that describes how to stitch together new proteins from connected or disconnected pieces of existing structures.
“We can now begin to think about engineering proteins to do things that nothing else is capable of doing,” said UNC’s Brian Kuhlman, Ph.D. “The structure of a protein determines its function, so if we are going to learn how to design new functions, we have to learn how to design new structures. Our study is a critical step in that direction and provides tools for creating proteins that haven’t been seen before in nature.”
Traditionally, researchers have used computational protein design to recreate in the laboratory what already exists in the natural world. In recent years, their focus has shifted toward inventing novel proteins with new functionality. These design projects all start with a specific structural “blueprint” in mind, and as a result are limited. Dr. Kuhlman and his colleagues, however, believe that by removing the limitations of a predetermined blueprint and taking cues from evolution they can more easily create functional proteins.
Dr. Kuhlman’s UNC team developed a protein design approach that emulates natural mechanisms for shuffling tertiary structures such as pleats, coils, and furrows. Putting the approach into action, the UNC team mapped 50,000 stitched together proteins on the computer, and then it produced 21 promising structures in the laboratory. Details of this work appeared May 6 in the journal Science, in an article entitled, “Design of Structurally Distinct Proteins Using Strategies Inspired by Evolution.”
“Helical proteins designed with SEWING contain structural features absent from other de novo designed proteins and, in some cases, remain folded at more than 100°C,” wrote the authors. “High-resolution structures of the designed proteins CA01 and DA05R1 were solved by x-ray crystallography (2.2 angstrom resolution) and nuclear magnetic resonance, respectively, and there was excellent agreement with the design models.”
Essentially, the UNC scientists confirmed that the proteins they had synthesized contained the unique structural varieties that had been designed on the computer. The UNC scientists also determined that the structures they had created had new surface and pocket features. Such features, they noted, provide potential binding sites for ligands or macromolecules.
“We were excited that some had clefts or grooves on the surface, regions that naturally occurring proteins use for binding other proteins,” said the Science article’s first author, Tim M. Jacobs, Ph.D., a former graduate student in Dr. Kuhlman’s laboratory. “That’s important because if we wanted to create a protein that can act as a biosensor to detect a certain metabolite in the body, either for diagnostic or research purposes, it would need to have these grooves. Likewise, if we wanted to develop novel therapeutics, they would also need to attach to specific proteins.”
Currently, the UNC researchers are using SEWING to create proteins that can bind to several other proteins at a time. Many of the most important proteins are such multitaskers, including the blood protein hemoglobin.
Histone Mutation Deranges DNA Methylation to Cause Cancer
In some cancers, including chondroblastoma and a rare form of childhood sarcoma, a mutation in histone H3 reduces global levels of methylation (dark areas) in tumor cells but not in normal cells (arrowhead). The mutation locks the cells in a proliferative state to promote tumor development. [Laboratory of Chromatin Biology and Epigenetics at The Rockefeller University]
They have been called oncohistones, the mutated histones that are known to accompany certain pediatric cancers. Despite their suggestive moniker, oncohistones have kept their oncogenic secrets. For example, it has been unclear whether oncohistones are able to cause cancer on their own, or whether they need to act in concert with additional DNA mutations, that is, mutations other than those affecting histone structures.
While oncohistone mechanisms remain poorly understood, this particular question—the oncogenicity of lone oncohistones—has been resolved, at least in part. According to researchers based at The Rockefeller University, a change to the structure of a histone can trigger a tumor on its own.
This finding appeared May 13 in the journal Science, in an article entitled, “Histone H3K36 Mutations Promote Sarcomagenesis Through Altered Histone Methylation Landscape.” The article describes the Rockefeller team’s study of a histone protein called H3, which has been found in about 95% of samples of chondoblastoma, a benign tumor that arises in cartilage, typically during adolescence.
The Rockefeller scientists found that the H3 lysine 36–to–methionine (H3K36M) mutation impairs the differentiation of mesenchymal progenitor cells and generates undifferentiated sarcoma in vivo.
After the scientists inserted the H3 histone mutation into mouse mesenchymal progenitor cells (MPCs)—which generate cartilage, bone, and fat—they watched these cells lose the ability to differentiate in the lab. Next, the scientists injected the mutant cells into living mice, and the animals developed the tumors rich in MPCs, known as an undifferentiated sarcoma. Finally, the researchers tried to understand how the mutation causes the tumors to develop.
The scientists determined that H3K36M mutant nucleosomes inhibit the enzymatic activities of several H3K36 methyltransferases.
“Depleting H3K36 methyltransferases, or expressing an H3K36I mutant that similarly inhibits H3K36 methylation, is sufficient to phenocopy the H3K36M mutation,” the authors of the Science study wrote. “After the loss of H3K36 methylation, a genome-wide gain in H3K27 methylation leads to a redistribution of polycomb repressive complex 1 and de-repression of its target genes known to block mesenchymal differentiation.”
Essentially, when the H3K36M mutation occurs, the cell becomes locked in a proliferative state—meaning it divides constantly, leading to tumors. Specifically, the mutation inhibits enzymes that normally tag the histone with chemical groups known as methyls, allowing genes to be expressed normally.
In response to this lack of modification, another part of the histone becomes overmodified, or tagged with too many methyl groups. “This leads to an overall resetting of the landscape of chromatin, the complex of DNA and its associated factors, including histones,” explained co-author Peter Lewis, Ph.D., a professor at the University of Wisconsin-Madison and a former postdoctoral fellow in laboratory of C. David Allis, Ph.D., a professor at Rockefeller.
The finding—that a “resetting” of the chromatin landscape can lock the cell into a proliferative state—suggests that researchers should be on the hunt for more mutations in histones that might be driving tumors. For their part, the Rockefeller researchers are trying to learn more about how this specific mutation in histone H3 causes tumors to develop.
“We want to know which pathways cause the mesenchymal progenitor cells that carry the mutation to continue to divide, and not differentiate into the bone, fat, and cartilage cells they are destined to become,” said co-author Chao Lu, Ph.D., a postdoctoral fellow in the Allis lab.
Once researchers understand more about these pathways, added Dr. Lewis, they can consider ways of blocking them with drugs, particularly in tumors such as MPC-rich sarcomas—which, unlike chondroblastoma, can be deadly. In fact, drugs that block these pathways may already exist and may even be in use for other types of cancers.
“One long-term goal of our collaborative team is to better understand fundamental mechanisms that drive these processes, with the hope of providing new therapeutic approaches,” concluded Dr. Allis.
Histone H3K36 mutations promote sarcomagenesis through altered histone methylation landscape
Missense mutations (that change one amino acid for another) in histone H3 can produce a so-called oncohistone and are found in a number of pediatric cancers. For example, the lysine-36–to-methionine (K36M) mutation is seen in almost all chondroblastomas. Lu et al. show that K36M mutant histones are oncogenic, and they inhibit the normal methylation of this same residue in wild-type H3 histones. The mutant histones also interfere with the normal development of bone-related cells and the deposition of inhibitory chromatin marks.
Several types of pediatric cancers reportedly contain high-frequency missense mutations in histone H3, yet the underlying oncogenic mechanism remains poorly characterized. Here we report that the H3 lysine 36–to–methionine (H3K36M) mutation impairs the differentiation of mesenchymal progenitor cells and generates undifferentiated sarcoma in vivo. H3K36M mutant nucleosomes inhibit the enzymatic activities of several H3K36 methyltransferases. Depleting H3K36 methyltransferases, or expressing an H3K36I mutant that similarly inhibits H3K36 methylation, is sufficient to phenocopy the H3K36M mutation. After the loss of H3K36 methylation, a genome-wide gain in H3K27 methylation leads to a redistribution of polycomb repressive complex 1 and de-repression of its target genes known to block mesenchymal differentiation. Our findings are mirrored in human undifferentiated sarcomas in which novel K36M/I mutations in H3.1 are identified.
Mitochondria? We Don’t Need No Stinking Mitochondria!
Diagram comparing typical eukaryotic cell to the newly discovered mitochondria-free organism. [Karnkowska et al., 2016, Current Biology 26, 1–11]
The organelle that produces a significant portion of energy for eukaryotic cells would seemingly be indispensable, yet over the years, a number of organisms have been discovered that challenge that biological pretense. However, these so-called amitochondrial species may lack a defined organelle, but they still retain some residual functions of their mitochondria-containing brethren. Even the intestinal eukaryotic parasite Giardia intestinalis, which was for many years considered to be mitochondria-free, was proven recently to contain a considerably shriveled version of the organelle.
Now, an international group of scientists has released results from a new study that challenges the notion that mitochondria are essential for eukaryotes—discovering an organism that resides in the gut of chinchillas that contains absolutely no trace of mitochondria at all.
“In low-oxygen environments, eukaryotes often possess a reduced form of the mitochondrion, but it was believed that some of the mitochondrial functions are so essential that these organelles are indispensable for their life,” explained lead study author Anna Karnkowska, Ph.D., visiting scientist at the University of British Columbia in Vancouver. “We have characterized a eukaryotic microbe which indeed possesses no mitochondrion at all.”
Mysterious Eukaryote Missing Mitochondria
Researchers uncover the first example of a eukaryotic organism that lacks the organelles.
Monocercomonoides sp. PA203VLADIMIR HAMPL, CHARLES UNIVERSITY, PRAGUE, CZECH REPUBLIC
Scientists have long thought that mitochondria—organelles responsible for energy generation—are an essential and defining feature of a eukaryotic cell. Now, researchers from Charles University in Prague and their colleagues are challenging this notion with their discovery of a eukaryotic organism,Monocercomonoides species PA203, which lacks mitochondria. The team’s phylogenetic analysis, published today (May 12) in Current Biology,suggests that Monocercomonoides—which belong to the Oxymonadida group of protozoa and live in low-oxygen environments—did have mitochondria at one point, but eventually lost the organelles.
“This is quite a groundbreaking discovery,” said Thijs Ettema, who studies microbial genome evolution at Uppsala University in Sweden and was not involved in the work.
“This study shows that mitochondria are not so central for all lineages of living eukaryotes,” Toni Gabaldonof the Center for Genomic Regulation in Barcelona, Spain, who also was not involved in the work, wrote in an email to The Scientist. “Yet, this mitochondrial-devoid, single-cell eukaryote is as complex as other eukaryotic cells in almost any other aspect of cellular complexity.”
Charles University’s Vladimir Hampl studies the evolution of protists. Along with Anna Karnkowska and colleagues, Hampl decided to sequence the genome of Monocercomonoides, a little-studied protist that lives in the digestive tracts of vertebrates. The 75-megabase genome—the first of an oxymonad—did not contain any conserved genes found on mitochondrial genomes of other eukaryotes, the researchers found. It also did not contain any nuclear genes associated with mitochondrial functions.
“It was surprising and for a long time, we didn’t believe that the [mitochondria-associated genes were really not there]. We thought we were missing something,” Hampl told The Scientist. “But when the data kept accumulating, we switched to the hypothesis that this organism really didn’t have mitochondria.”
Because researchers have previously not found examples of eukaryotes without some form of mitochondria, the current theory of the origin of eukaryotes poses that the appearance of mitochondria was crucial to the identity of these organisms.
“We now view these mitochondria-like organelles as a continuum from full mitochondria to very small . Some anaerobic protists, for example, have only pared down versions of mitochondria, such as hydrogenosomes and mitosomes, which lack a mitochondrial genome. But these mitochondrion-like organelles perform essential functions of the iron-sulfur cluster assembly pathway, which is known to be conserved in virtually all eukaryotic organisms studied to date.
Yet, in their analysis, the researchers found no evidence of the presence of any components of this mitochondrial pathway.
Like the scaling down of mitochondria into mitosomes in some organisms, the ancestors of modernMonocercomonoides once had mitochondria. “Because this organism is phylogenetically nested among relatives that had conventional mitochondria, this is most likely a secondary adaptation,” said Michael Gray, a biochemist who studies mitochondria at Dalhousie University in Nova Scotia and was not involved in the study. According to Gray, the finding of a mitochondria-deficient eukaryote does not mean that the organelles did not play a major role in the evolution of eukaryotic cells.
To be sure they were not missing mitochondrial proteins, Hampl’s team also searched for potential mitochondrial protein homologs of other anaerobic species, and for signature sequences of a range of known mitochondrial proteins. While similar searches with other species uncovered a few mitochondrial proteins, the team’s analysis of Monocercomonoides came up empty.
“The data is very complete,” said Ettema. “It is difficult to prove the absence of something but [these authors] do a convincing job.”
To form the essential iron-sulfur clusters, the team discovered that Monocercomonoides use a sulfur mobilization system found in the cytosol, and that an ancestor of the organism acquired this system by lateral gene transfer from bacteria. This cytosolic, compensating system allowed Monocercomonoides to lose the otherwise essential iron-sulfur cluster-forming pathway in the mitochondrion, the team proposed.
“This work shows the great evolutionary plasticity of the eukaryotic cell,” said Karnkowska, who participated in the study while she was a postdoc at Charles University. Karnkowska, who is now a visiting researcher at the University of British Columbia in Canada, added: “This is a striking example of how far the evolution of a eukaryotic cell can go that was beyond our expectations.”
“The results highlight how many surprises may await us in the poorly studied eukaryotic phyla that live in under-explored environments,” Gabaldon said.
Ettema agreed. “Now that we’ve found one, we need to look at the bigger picture and see if there are other examples of eukaryotes that have lost their mitochondria, to understand how adaptable eukaryotes are.”
Karnkowska et al., “A eukaryote without a mitochondrial organelle,” Current Biology,doi:10.1016/j.cub.2016.03.053, 2016.
•Monocercomonoides sp. is a eukaryotic microorganism with no mitochondria
•The complete absence of mitochondria is a secondary loss, not an ancestral feature
•The essential mitochondrial ISC pathway was replaced by a bacterial SUF system
The presence of mitochondria and related organelles in every studied eukaryote supports the view that mitochondria are essential cellular components. Here, we report the genome sequence of a microbial eukaryote, the oxymonad Monocercomonoides sp., which revealed that this organism lacks all hallmark mitochondrial proteins. Crucially, the mitochondrial iron-sulfur cluster assembly pathway, thought to be conserved in virtually all eukaryotic cells, has been replaced by a cytosolic sulfur mobilization system (SUF) acquired by lateral gene transfer from bacteria. In the context of eukaryotic phylogeny, our data suggest that Monocercomonoides is not primitively amitochondrial but has lost the mitochondrion secondarily. This is the first example of a eukaryote lacking any form of a mitochondrion, demonstrating that this organelle is not absolutely essential for the viability of a eukaryotic cell.
This method catches a bait protein together with its associated protein partners in virus-like particles that are budded from human cells. Like this, cell lysis is not needed and protein complexes are preserved during purification.
With his feet in both a proteomics lab and an interactomics lab, VIB/UGent professor Sven Eyckerman is well aware of the shortcomings of conventional approaches to analyze protein complexes. The lysis conditions required in mass spectrometry–based strategies to break open cell membranes often affect protein-protein interactions. “The first step in a classical study on protein complexes essentially turns the highly organized cellular structure into a big messy soup”, Eyckerman explains.
Inspired by virus biology, Eyckerman came up with a creative solution. “We used the natural process of HIV particle formation to our benefit by hacking a completely safe form of the virus to abduct intact protein machines from the cell.” It is well known that the HIV virus captures a number of host proteins during its particle formation. By fusing a bait protein to the HIV-1 GAG protein, interaction partners become trapped within virus-like particles that bud from mammalian cells. Standard proteomic approaches are used next to reveal the content of these particles. Fittingly, the team named the method ‘Virotrap’.
The Virotrap approach is exceptional as protein networks can be characterized under natural conditions. By trapping protein complexes in the protective environment of a virus-like shell, the intact complexes are preserved during the purification process. The researchers showed the method was suitable for detection of known binary interactions as well as mass spectrometry-based identification of novel protein partners.
Virotrap is a textbook example of bringing research teams with complementary expertise together. Cross-pollination with the labs of Jan Tavernier (VIB/UGent) and Kris Gevaert (VIB/UGent) enabled the development of this platform.
Jan Tavernier: “Virotrap represents a new concept in co-complex analysis wherein complex stability is physically guaranteed by a protective, physical structure. It is complementary to the arsenal of existing interactomics methods, but also holds potential for other fields, like drug target characterization. We also developed a small molecule-variant of Virotrap that could successfully trap protein partners for small molecule baits.”
Kris Gevaert: “Virotrap can also impact our understanding of disease pathways. We were actually surprised to see that this virus-based system could be used to study antiviral pathways, like Toll-like receptor signaling. Understanding these protein machines in their natural environment is essential if we want to modulate their activity in pathology.“
Trapping mammalian protein complexes in viral particles
Cell lysis is an inevitable step in classical mass spectrometry–based strategies to analyse protein complexes. Complementary lysis conditions, in situ cross-linking strategies and proximal labelling techniques are currently used to reduce lysis effects on the protein complex. We have developed Virotrap, a viral particle sorting approach that obviates the need for cell homogenization and preserves the protein complexes during purification. By fusing a bait protein to the HIV-1 GAG protein, we show that interaction partners become trapped within virus-like particles (VLPs) that bud from mammalian cells. Using an efficient VLP enrichment protocol, Virotrap allows the detection of known binary interactions and MS-based identification of novel protein partners as well. In addition, we show the identification of stimulus-dependent interactions and demonstrate trapping of protein partners for small molecules. Virotrap constitutes an elegant complementary approach to the arsenal of methods to study protein complexes.
Proteins mostly exert their function within supramolecular complexes. Strategies for detecting protein–protein interactions (PPIs) can be roughly divided into genetic systems1 and co-purification strategies combined with mass spectrometry (MS) analysis (for example, AP–MS)2. The latter approaches typically require cell or tissue homogenization using detergents, followed by capture of the protein complex using affinity tags3 or specific antibodies4. The protein complexes extracted from this ‘soup’ of constituents are then subjected to several washing steps before actual analysis by trypsin digestion and liquid chromatography–MS/MS analysis. Such lysis and purification protocols are typically empirical and have mostly been optimized using model interactions in single labs. In fact, lysis conditions can profoundly affect the number of both specific and nonspecific proteins that are identified in a typical AP–MS set-up. Indeed, recent studies using the nuclear pore complex as a model protein complex describe optimization of purifications for the different proteins in the complex by examining 96 different conditions5. Nevertheless, for new purifications, it remains hard to correctly estimate the loss of factors in a standard AP–MS experiment due to washing and dilution effects during treatments (that is, false negatives). These considerations have pushed the concept of stabilizing PPIs before the actual homogenization step. A classical approach involves cross-linking with simple reagents (for example, formaldehyde) or with more advanced isotope-labelled cross-linkers (reviewed in ref. 2). However, experimental challenges such as cell permeability and reactivity still preclude the widespread use of cross-linking agents. Moreover, MS-generated spectra of cross-linked peptides are notoriously difficult to identify correctly. A recent lysis-independent solution involves the expression of a bait protein fused to a promiscuous biotin ligase, which results in labelling of proteins proximal to the activity of the enzyme-tagged bait protein6. When compared with AP–MS, this BioID approach delivers a complementary set of candidate proteins, including novel interaction partners7, 8. Such particular studies clearly underscore the need for complementary approaches in the co-complex strategies.
The evolutionary stress on viruses promoted highly condensed coding of information and maximal functionality for small genomes. Accordingly, for HIV-1 it is sufficient to express a single protein, the p55 GAG protein, for efficient production of virus-like particles (VLPs) from cells9, 10. This protein is highly mobile before its accumulation in cholesterol-rich regions of the membrane, where multimerization initiates the budding process11. A total of 4,000–5,000 GAG molecules is required to form a single particle of about 145 nm (ref. 12). Both VLPs and mature viruses contain a number of host proteins that are recruited by binding to viral proteins. These proteins can either contribute to the infectivity (for example, Cyclophilin/FKBPA13) or act as antiviral proteins preventing the spreading of the virus (for example, APOBEC proteins14).
We here describe the development and application of Virotrap, an elegant co-purification strategy based on the trapping of a bait protein together with its associated protein partners in VLPs that are budded from the cell. After enrichment, these particles can be analysed by targeted (for example, western blotting) or unbiased approaches (MS-based proteomics). Virotrap allows detection of known binary PPIs, analysis of protein complexes and their dynamics, and readily detects protein binders for small molecules.
Concept of the Virotrap system
Classical AP–MS approaches rely on cell homogenization to access protein complexes, a step that can vary significantly with the lysis conditions (detergents, salt concentrations, pH conditions and so on)5. To eliminate the homogenization step in AP–MS, we reasoned that incorporation of a protein complex inside a secreted VLP traps the interaction partners under native conditions and protects them during further purification. We thus explored the possibility of protein complex packaging by the expression of GAG-bait protein chimeras (Fig. 1) as expression of GAG results in the release of VLPs from the cells9, 10. As a first PPI pair to evaluate this concept, we selected the HRAS protein as a bait combined with the RAF1 prey protein. We were able to specifically detect the HRAS–RAF1 interaction following enrichment of VLPs via ultracentrifugation (Supplementary Fig. 1a). To prevent tedious ultracentrifugation steps, we designed a novel single-step protocol wherein we co-express the vesicular stomatitis virus glycoprotein (VSV-G) together with a tagged version of this glycoprotein in addition to the GAG bait and prey. Both tagged and untagged VSV-G proteins are probably presented as trimers on the surface of the VLPs, allowing efficient antibody-based recovery from large volumes. The HRAS–RAF1 interaction was confirmed using this single-step protocol (Supplementary Fig. 1b). No associations with unrelated bait or prey proteins were observed for both protocols.
Figure 1: Schematic representation of the Virotrap strategy.
Expression of a GAG-bait fusion protein (1) results in submembrane multimerization (2) and subsequent budding of VLPs from cells (3). Interaction partners of the bait protein are also trapped within these VLPs and can be identified after purification by western blotting or MS analysis (4).
Virotrap for the detection of binary interactions
We next explored the reciprocal detection of a set of PPI pairs, which were selected based on published evidence and cytosolic localization15. After single-step purification and western blot analysis, we could readily detect reciprocal interactions between CDK2 and CKS1B, LCP2 and GRAP2, and S100A1 and S100B (Fig. 2a). Only for the LCP2 prey we observed nonspecific association with an irrelevant bait construct. However, the particle levels of the GRAP2 bait were substantially lower as compared with those of the GAG control construct (GAG protein levels in VLPs; Fig. 2a, second panel of the LCP2 prey). After quantification of the intensities of bait and prey proteins and normalization of prey levels using bait levels, we observed a strong enrichment for the GAG-GRAP2 bait (Supplementary Fig. 2).
…..
Virotrap for unbiased discovery of novel interactions
For the detection of novel interaction partners, we scaled up VLP production and purification protocols (Supplementary Fig. 5 and Supplementary Note 1 for an overview of the protocol) and investigated protein partners trapped using the following bait proteins: Fas-associated via death domain (FADD), A20 (TNFAIP3), nuclear factor-κB (NF-κB) essential modifier (IKBKG), TRAF family member-associated NF-κB activator (TANK), MYD88 and ring finger protein 41 (RNF41). To obtain specific interactors from the lists of identified proteins, we challenged the data with a combined protein list of 19 unrelated Virotrap experiments (Supplementary Table 1 for an overview). Figure 3 shows the design and the list of candidate interactors obtained after removal of all proteins that were found in the 19 control samples (including removal of proteins from the control list identified with a single peptide). The remaining list of confident protein identifications (identified with at least two peptides in at least two biological repeats) reveals both known and novel candidate interaction partners. All candidate interactors including single peptide protein identifications are given in Supplementary Data 2 and also include recurrent protein identifications of known interactors based on a single peptide; for example, CASP8 for FADD and TANK for NEMO. Using alternative methods, we confirmed the interaction between A20 and FADD, and the associations with transmembrane proteins (insulin receptor and insulin-like growth factor receptor 1) that were captured using RNF41 as a bait (Supplementary Fig. 6). To address the use of Virotrap for the detection of dynamic interactions, we activated the NF-κB pathway via the tumour necrosis factor (TNF) receptor (TNFRSF1A) using TNFα (TNF) and performed Virotrap analysis using A20 as bait (Fig. 3). This resulted in the additional enrichment of receptor-interacting kinase (RIPK1), TNFR1-associated via death domain (TRADD), TNFRSF1A and TNF itself, confirming the expected activated complex20.
Figure 3: Use of Virotrap for unbiased interactome analysis
Lysis conditions used in AP–MS strategies are critical for the preservation of protein complexes. A multitude of lysis conditions have been described, culminating in a recent report where protein complex stability was assessed under 96 lysis/purification protocols5. Moreover, the authors suggest to optimize the conditions for every complex, implying an important workload for researchers embarking on protein complex analysis using classical AP–MS. As lysis results in a profound change of the subcellular context and significantly alters the concentration of proteins, loss of complex integrity during a classical AP–MS protocol can be expected. A clear evolution towards ‘lysis-independent’ approaches in the co-complex analysis field is evident with the introduction of BioID6 and APEX25 where proximal proteins, including proteins residing in the complex, are labelled with biotin by an enzymatic activity fused to a bait protein. A side-by-side comparison between classical AP–MS and BioID showed overlapping and unique candidate binding proteins for both approaches7, 8, supporting the notion that complementary methods are needed to provide a comprehensive view on protein complexes. This has also been clearly demonstrated for binary approaches15 and is a logical consequence of the heterogenic nature underlying PPIs (binding mechanism, requirement for posttranslational modifications, location, affinity and so on).
In this report, we explore an alternative, yet complementary method to isolate protein complexes without interfering with cellular integrity. By trapping protein complexes in the protective environment of a virus-like shell, the intact complexes are preserved during the purification process. This constitutes a new concept in co-complex analysis wherein complex stability is physically guaranteed by a protective, physical structure. A comparison of our Virotrap approach with AP–MS shows complementary data, with specific false positives and false negatives for both methods (Supplementary Fig. 7).
The current implementation of the Virotrap platform implies the use of a GAG-bait construct resulting in considerable expression of the bait protein. Different strategies are currently pursued to reduce bait expression including co-expression of a native GAG protein together with the GAG-bait protein, not only reducing bait expression but also creating more ‘space’ in the particles potentially accommodating larger bait protein complexes. Nevertheless, the presence of the bait on the forming GAG scaffold creates an intracellular affinity matrix (comparable to the early in vitro affinity columns for purification of interaction partners from lysates26) that has the potential to compete with endogenous complexes by avidity effects. This avidity effect is a powerful mechanism that aids in the recruitment of cyclophilin to GAG27, a well-known weak interaction (Kd=16 μM (ref. 28)) detectable as a background association in the Virotrap system. Although background binding may be increased by elevated bait expression, weaker associations are readily detectable (for example, MAL—MYD88-binding study; Fig. 2c).
The size of Virotrap particles (around 145 nm) suggests limitations in the size of the protein complex that can be accommodated in the particles. Further experimentation is required to define the maximum size of proteins or the number of protein complexes that can be trapped inside the particles.
….
In conclusion, Virotrap captures significant parts of known interactomes and reveals new interactions. This cell lysis-free approach purifies protein complexes under native conditions and thus provides a powerful method to complement AP–MS or other PPI data. Future improvements of the system include strategies to reduce bait expression to more physiological levels and application of advanced data analysis options to filter out background. These developments can further aid in the deployment of Virotrap as a powerful extension of the current co-complex technology arsenal.
New Autism Blood Biomarker Identified
Researchers at UT Southwestern Medical Center have identified a blood biomarker that may aid in earlier diagnosis of children with autism spectrum disorder, or ASD
In a recent edition of Scientific Reports, UT Southwestern researchers reported on the identification of a blood biomarker that could distinguish the majority of ASD study participants versus a control group of similar age range. In addition, the biomarker was significantly correlated with the level of communication impairment, suggesting that the blood test may give insight into ASD severity.
“Numerous investigators have long sought a biomarker for ASD,” said Dr. Dwight German, study senior author and Professor of Psychiatry at UT Southwestern. “The blood biomarker reported here along with others we are testing can represent a useful test with over 80 percent accuracy in identifying ASD.”
ASD1 – was 66 percent accurate in diagnosing ASD. When combined with thyroid stimulating hormone level measurements, the ASD1-binding biomarker was 73 percent accurate at diagnosis
A Search for Blood Biomarkers for Autism: Peptoids
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by impairments in social interaction and communication, and restricted, repetitive patterns of behavior. In order to identify individuals with ASD and initiate interventions at the earliest possible age, biomarkers for the disorder are desirable. Research findings have identified widespread changes in the immune system in children with autism, at both systemic and cellular levels. In an attempt to find candidate antibody biomarkers for ASD, highly complex libraries of peptoids (oligo-N-substituted glycines) were screened for compounds that preferentially bind IgG from boys with ASD over typically developing (TD) boys. Unexpectedly, many peptoids were identified that preferentially bound IgG from TD boys. One of these peptoids was studied further and found to bind significantly higher levels (>2-fold) of the IgG1 subtype in serum from TD boys (n = 60) compared to ASD boys (n = 74), as well as compared to older adult males (n = 53). Together these data suggest that ASD boys have reduced levels (>50%) of an IgG1 antibody, which resembles the level found normally with advanced age. In this discovery study, the ASD1 peptoid was 66% accurate in predicting ASD.
….
Peptoid libraries have been used previously to search for autoantibodies for neurodegenerative diseases19 and for systemic lupus erythematosus (SLE)21. In the case of SLE, peptoids were identified that could identify subjects with the disease and related syndromes with moderate sensitivity (70%) and excellent specificity (97.5%). Peptoids were used to measure IgG levels from both healthy subjects and SLE patients. Binding to the SLE-peptoid was significantly higher in SLE patients vs. healthy controls. The IgG bound to the SLE-peptoid was found to react with several autoantigens, suggesting that the peptoids are capable of interacting with multiple, structurally similar molecules. These data indicate that IgG binding to peptoids can identify subjects with high levels of pathogenic autoantibodies vs. a single antibody.
In the present study, the ASD1 peptoid binds significantly lower levels of IgG1 in ASD males vs. TD males. This finding suggests that the ASD1 peptoid recognizes antibody(-ies) of an IgG1 subtype that is (are) significantly lower in abundance in the ASD males vs. TD males. Although a previous study14 has demonstrated lower levels of plasma IgG in ASD vs. TD children, here, we additionally quantified serum IgG levels in our individuals and found no difference in IgG between the two groups (data not shown). Furthermore, our IgG levels did not correlate with ASD1 binding levels, indicating that ASD1 does not bind IgG generically, and that the peptoid’s ability to differentiate between ASD and TD males is related to a specific antibody(-ies).
ASD subjects underwent a diagnostic evaluation using the ADOS and ADI-R, and application of the DSM-IV criteria prior to study inclusion. Only those subjects with a diagnosis of Autistic Disorder were included in the study. The ADOS is a semi-structured observation of a child’s behavior that allows examiners to observe the three core domains of ASD symptoms: reciprocal social interaction, communication, and restricted and repetitive behaviors1. When ADOS subdomain scores were compared with peptoid binding, the only significant relationship was with Social Interaction. However, the positive correlation would suggest that lower peptoid binding is associated with better social interaction, not poorer social interaction as anticipated.
The ADI-R is a structured parental interview that measures the core features of ASD symptoms in the areas of reciprocal social interaction, communication and language, and patterns of behavior. Of the three ADI-R subdomains, only the Communication domain was related to ASD1 peptoid binding, and this correlation was negative suggesting that low peptoid binding is associated with greater communication problems. These latter data are similar to the findings of Heuer et al.14 who found that children with autism with low levels of plasma IgG have high scores on the Aberrant Behavior Checklist (p < 0.0001). Thus, peptoid binding to IgG1 may be useful as a severity marker for ASD allowing for further characterization of individuals, but further research is needed.
It is interesting that in serum samples from older men, the ASD1 binding is similar to that in the ASD boys. This is consistent with the observation that with aging there is a reduction in the strength of the immune system, and the changes are gender-specific25. Recent studies using parabiosis26, in which blood from young mice reverse age-related impairments in cognitive function and synaptic plasticity in old mice, reveal that blood constituents from young subjects may contain important substances for maintaining neuronal functions. Work is in progress to identify the antibody/antibodies that are differentially binding to the ASD1 peptoid, which appear as a single band on the electrophoresis gel (Fig. 4).
……..
The ADI-R is a structured parental interview that measures the core features of ASD symptoms in the areas of reciprocal social interaction, communication and language, and patterns of behavior. Of the three ADI-R subdomains, only the Communication domain was related to ASD1 peptoid binding, and this correlation was negative suggesting that low peptoid binding is associated with greater communication problems. These latter data are similar to the findings of Heuer et al.14 who found that children with autism with low levels of plasma IgG have high scores on the Aberrant Behavior Checklist (p < 0.0001). Thus, peptoid binding to IgG1 may be useful as a severity marker for ASD allowing for further characterization of individuals, but further research is needed.
Titration of IgG binding to ASD1 using serum pooled from 10 TD males and 10 ASD males demonstrates ASD1’s ability to differentiate between the two groups. (B)Detecting IgG1 subclass instead of total IgG amplifies this differentiation. (C) IgG1 binding of individual ASD (n=74) and TD (n=60) male serum samples (1:100 dilution) to ASD1 significantly differs with TD>ASD. In addition, IgG1 binding of older adult male (AM) serum samples (n=53) to ASD1 is significantly lower than TD males, and not different from ASD males. The three groups were compared with a Kruskal-Wallis ANOVA, H = 10.1781, p<0.006. **p<0.005. Error bars show SEM. (D) Receiver-operating characteristic curve for ASD1’s ability to discriminate between ASD and TD males.
Association between peptoid binding and ADOS and ADI-R subdomains
Higher scores in any domain on the ADOS and ADI-R are indicative of more abnormal behaviors and/or symptoms. Among ADOS subdomains, there was no significant relationship between Communication and peptoid binding (z = 0.04, p = 0.966), Communication + Social interaction (z = 1.53, p = 0.127), or Stereotyped Behaviors and Restrictive Interests (SBRI) (z = 0.46, p = 0.647). Higher scores on the Social Interaction domain were significantly associated with higher peptoid binding (z = 2.04, p = 0.041).
Among ADI-R subdomains, higher scores on the Communication domain were associated with lower levels of peptoid binding (z = −2.28, p = 0.023). There was not a significant relationship between Social Interaction (z = 0.07, p = 0.941) or Restrictive/Repetitive Stereotyped Behaviors (z = −1.40, p = 0.162) and peptoid binding.
Computational Model Finds New Protein-Protein Interactions
Researchers at University of Pittsburgh have discovered 500 new protein-protein interactions (PPIs) associated with genes linked to schizophrenia.
Using a computational model they developed, researchers at the University of Pittsburgh School of Medicine have discovered more than 500 new protein-protein interactions (PPIs) associated with genes linked to schizophrenia. The findings, published online in npj Schizophrenia, a Nature Publishing Group journal, could lead to greater understanding of the biological underpinnings of this mental illness, as well as point the way to treatments.
There have been many genome-wide association studies (GWAS) that have identified gene variants associated with an increased risk for schizophrenia, but in most cases there is little known about the proteins that these genes make, what they do and how they interact, said senior investigator Madhavi Ganapathiraju, Ph.D., assistant professor of biomedical informatics, Pitt School of Medicine.
“GWAS studies and other research efforts have shown us what genes might be relevant in schizophrenia,” she said. “What we have done is the next step. We are trying to understand how these genes relate to each other, which could show us the biological pathways that are important in the disease.”
Each gene makes proteins and proteins typically interact with each other in a biological process. Information about interacting partners can shed light on the role of a gene that has not been studied, revealing pathways and biological processes associated with the disease and also its relation to other complex diseases.
Dr. Ganapathiraju’s team developed a computational model called High-Precision Protein Interaction Prediction (HiPPIP) and applied it to discover PPIs of schizophrenia-linked genes identified through GWAS, as well as historically known risk genes. They found 504 never-before known PPIs, and noted also that while schizophrenia-linked genes identified historically and through GWAS had little overlap, the model showed they shared more than 100 common interactors.
“We can infer what the protein might do by checking out the company it keeps,” Dr. Ganapathiraju explained. “For example, if I know you have many friends who play hockey, it could mean that you are involved in hockey, too. Similarly, if we see that an unknown protein interacts with multiple proteins involved in neural signaling, for example, there is a high likelihood that the unknown entity also is involved in the same.”
Dr. Ganapathiraju and colleagues have drawn such inferences on protein function based on the PPIs of proteins, and made their findings available on a website Schizo-Pi. This information can be used by biologists to explore the schizophrenia interactome with the aim of understanding more about the disease or developing new treatment drugs.
Schizophrenia interactome with 504 novel protein–protein interactions
(GWAS) have revealed the role of rare and common genetic variants, but the functional effects of the risk variants remain to be understood. Protein interactome-based studies can facilitate the study of molecular mechanisms by which the risk genes relate to schizophrenia (SZ) genesis, but protein–protein interactions (PPIs) are unknown for many of the liability genes. We developed a computational model to discover PPIs, which is found to be highly accurate according to computational evaluations and experimental validations of selected PPIs. We present here, 365 novel PPIs of liability genes identified by the SZ Working Group of the Psychiatric Genomics Consortium (PGC). Seventeen genes that had no previously known interactions have 57 novel interactions by our method. Among the new interactors are 19 drug targets that are targeted by 130 drugs. In addition, we computed 147 novel PPIs of 25 candidate genes investigated in the pre-GWAS era. While there is little overlap between the GWAS genes and the pre-GWAS genes, the interactomes reveal that they largely belong to the same pathways, thus reconciling the apparent disparities between the GWAS and prior gene association studies. The interactome including 504 novel PPIs overall, could motivate other systems biology studies and trials with repurposed drugs. The PPIs are made available on a webserver, called Schizo-Pi at http://severus.dbmi.pitt.edu/schizo-pi with advanced search capabilities.
Schizophrenia (SZ) is a common, potentially severe psychiatric disorder that afflicts all populations.1 Gene mapping studies suggest that SZ is a complex disorder, with a cumulative impact of variable genetic effects coupled with environmental factors.2 As many as 38 genome-wide association studies (GWAS) have been reported on SZ out of a total of 1,750 GWAS publications on 1,087 traits or diseases reported in the GWAS catalog maintained by the National Human Genome Research Institute of USA3 (as of April 2015), revealing the common variants associated with SZ.4 The SZ Working Group of the Psychiatric Genomics Consortium (PGC) identified 108 genetic loci that likely confer risk for SZ.5 While the role of genetics has been clearly validated by this study, the functional impact of the risk variants is not well-understood.6,7 Several of the genes implicated by the GWAS have unknown functions and could participate in possibly hitherto unknown pathways.8 Further, there is little or no overlap between the genes identified through GWAS and ‘candidate genes’ proposed in the pre-GWAS era.9
Interactome-based studies can be useful in discovering the functional associations of genes. For example,disrupted in schizophrenia 1 (DISC1), an SZ related candidate gene originally had no known homolog in humans. Although it had well-characterized protein domains such as coiled-coil domains and leucine-zipper domains, its function was unknown.10,11 Once its protein–protein interactions (PPIs) were determined using yeast 2-hybrid technology,12 investigators successfully linked DISC1 to cAMP signaling, axon elongation, and neuronal migration, and accelerated the research pertaining to SZ in general, and DISC1 in particular.13 Typically such studies are carried out on known protein–protein interaction (PPI) networks, or as in the case of DISC1, when there is a specific gene of interest, its PPIs are determined by methods such as yeast 2-hybrid technology.
Knowledge of human PPI networks is thus valuable for accelerating discovery of protein function, and indeed, biomedical research in general. However, of the hundreds of thousands of biophysical PPIs thought to exist in the human interactome,14,15 <100,000 are known today (Human Protein Reference Database, HPRD16 and BioGRID17 databases). Gold standard experimental methods for the determination of all the PPIs in human interactome are time-consuming, expensive and may not even be feasible, as about 250 million pairs of proteins would need to be tested overall; high-throughput methods such as yeast 2-hybrid have important limitations for whole interactome determination as they have a low recall of 23% (i.e., remaining 77% of true interactions need to be determined by other means), and a low precision (i.e., the screens have to be repeated multiple times to achieve high selectivity).18,19Computational methods are therefore necessary to complete the interactome expeditiously. Algorithms have begun emerging to predict PPIs using statistical machine learning on the characteristics of the proteins, but these algorithms are employed predominantly to study yeast. Two significant computational predictions have been reported for human interactome; although they have had high false positive rates, these methods have laid the foundation for computational prediction of human PPIs.20,21
We have created a new PPI prediction model called High-Confidence Protein–Protein Interaction Prediction (HiPPIP) model. Novel interactions predicted with this model are making translational impact. For example, we discovered a PPI between OASL and DDX58, which on validation showed that an increased expression of OASL could boost innate immunity to combat influenza by activating the RIG-I pathway.22 Also, the interactome of the genes associated with congenital heart disease showed that the disease morphogenesis has a close connection with the structure and function of cilia.23Here, we describe the HiPPIP model and its application to SZ genes to construct the SZ interactome. After computational evaluations and experimental validations of selected novel PPIs, we present here 504 highly confident novel PPIs in the SZ interactome, shedding new light onto several uncharacterized genes that are associated with SZ.
We developed a computational model called HiPPIP to predict PPIs (see Methods and Supplementary File 1). The model has been evaluated by computational methods and experimental validations and is found to be highly accurate. Evaluations on a held-out test data showed a precision of 97.5% and a recall of 5%. 5% recall out of 150,000 to 600,000 estimated number of interactions in the human interactome corresponds to 7,500–30,000 novel PPIs in the whole interactome. Note that, it is likely that the real precision would be higher than 97.5% because in this test data, randomly paired proteins are treated as non-interacting protein pairs, whereas some of them may actually be interacting pairs with a small probability; thus, some of the pairs that are treated as false positives in test set are likely to be true but hitherto unknown interactions. In Figure 1a, we show the precision versus recall of our method on ‘hub proteins’ where we considered all pairs that received a score >0.5 by HiPPIP to be novel interactions. In Figure 1b, we show the number of true positives versus false positives observed in hub proteins. Both these figures also show our method to be superior in comparison to the prediction of membrane-receptor interactome by Qi et al’s.24 True positives versus false positives are also shown for individual hub proteins by our method in Figure 1cand by Qi et al’s.23 in Figure 1d. These evaluations showed that our predictions contain mostly true positives. Unlike in other domains where ranked lists are commonly used such as information retrieval, in PPI prediction the ‘false positives’ may actually be unlabeled instances that are indeed true interactions that are not yet discovered. In fact, such unlabeled pairs predicted as interactors of the hub gene HMGB1 (namely, the pairs HMGB1-KL and HMGB1-FLT1) were validated by experimental methods and found to be true PPIs (See the Figures e–g inSupplementary File 3). Thus, we concluded that the protein pairs that received a score of ⩾0.5 are highly confident to be true interactions. The pairs that receive a score less than but close to 0.5 (i.e., in the range of 0.4–0.5) may also contain several true PPIs; however, we cannot confidently say that all in this range are true PPIs. Only the PPIs predicted with a score >0.5 are included in the interactome.
Computational evaluation of predicted protein–protein interactions on hub proteins: (a) precision recall curve. (b) True positive versus false positives in ranked lists of hub type membrane receptors for our method and that by Qi et al. True positives versus false positives are shown for individual membrane receptors by our method in (c) and by Qi et al. in (d). Thick line is the average, which is also the same as shown in (b). Note:x-axis is recall in (a), whereas it is number of false positives in (b–d). The range of y-axis is observed by varying the threshold from 1.0–0 in (a), and to 0.5 in (b–d).
SZ interactome
By applying HiPPIP to the GWAS genes and Historic (pre-GWAS) genes, we predicted over 500 high confidence new PPIs adding to about 1400 previously known PPIs.
Schizophrenia interactome: network view of the schizophrenia interactome is shown as a graph, where genes are shown as nodes and PPIs as edges connecting the nodes. Schizophrenia-associated genes are shown as dark blue nodes, novel interactors as red color nodes and known interactors as blue color nodes. The source of the schizophrenia genes is indicated by its label font, where Historic genes are shown italicized, GWAS genes are shown in bold, and the one gene that is common to both is shown in italicized and bold. For clarity, the source is also indicated by the shape of the node (triangular for GWAS and square for Historic and hexagonal for both). Symbols are shown only for the schizophrenia-associated genes; actual interactions may be accessed on the web. Red edges are the novel interactions, whereas blue edges are known interactions. GWAS, genome-wide association studies of schizophrenia; PPI, protein–protein interaction.
We have made the known and novel interactions of all SZ-associated genes available on a webserver called Schizo-Pi, at the addresshttp://severus.dbmi.pitt.edu/schizo-pi. This webserver is similar to Wiki-Pi33 which presents comprehensive annotations of both participating proteins of a PPI side-by-side. The difference between Wiki-Pi which we developed earlier, and Schizo-Pi, is the inclusion of novel predicted interactions of the SZ genes into the latter.
Despite the many advances in biomedical research, identifying the molecular mechanisms underlying the disease is still challenging. Studies based on protein interactions were proven to be valuable in identifying novel gene associations that could shed new light on disease pathology.35 The interactome including more than 500 novel PPIs will help to identify pathways and biological processes associated with the disease and also its relation to other complex diseases. It also helps identify potential drugs that could be repurposed to use for SZ treatment.
Functional and pathway enrichment in SZ interactome
When a gene of interest has little known information, functions of its interacting partners serve as a starting point to hypothesize its own function. We computed statistically significant enrichment of GO biological process terms among the interacting partners of each of the genes using BinGO36 (see online at http://severus.dbmi.pitt.edu/schizo-pi).
Protein aggregation and aggregate toxicity: new insights into protein folding, misfolding diseases and biological evolution
Massimo Stefani · Christopher M. Dobson
Abstract The deposition of proteins in the form of amyloid fibrils and plaques is the characteristic feature of more than 20 degenerative conditions affecting either the central nervous system or a variety of peripheral tissues. As these conditions include Alzheimer’s, Parkinson’s and the prion diseases, several forms of fatal systemic amyloidosis, and at least one condition associated with medical intervention (haemodialysis), they are of enormous importance in the context of present-day human health and welfare. Much remains to be learned about the mechanism by which the proteins associated with these diseases aggregate and form amyloid structures, and how the latter affect the functions of the organs with which they are associated. A great deal of information concerning these diseases has emerged, however, during the past 5 years, much of it causing a number of fundamental assumptions about the amyloid diseases to be reexamined. For example, it is now apparent that the ability to form amyloid structures is not an unusual feature of the small number of proteins associated with these diseases but is instead a general property of polypeptide chains. It has also been found recently that aggregates of proteins not associated with amyloid diseases can impair the ability of cells to function to a similar extent as aggregates of proteins linked with specific neurodegenerative conditions. Moreover, the mature amyloid fibrils or plaques appear to be substantially less toxic than the prefibrillar aggregates that are their precursors. The toxicity of these early aggregates appears to result from an intrinsic ability to impair fundamental cellular processes by interacting with cellular membranes, causing oxidative stress and increases in free Ca2+ that eventually lead to apoptotic or necrotic cell death. The ‘new view’ of these diseases also suggests that other degenerative conditions could have similar underlying origins to those of the amyloidoses. In addition, cellular protection mechanisms, such as molecular chaperones and the protein degradation machinery, appear to be crucial in the prevention of disease in normally functioning living organisms. It also suggests some intriguing new factors that could be of great significance in the evolution of biological molecules and the mechanisms that regulate their behaviour.
The genetic information within a cell encodes not only the specific structures and functions of proteins but also the way these structures are attained through the process known as protein folding. In recent years many of the underlying features of the fundamental mechanism of this complex process and the manner in which it is regulated in living systems have emerged from a combination of experimental and theoretical studies [1]. The knowledge gained from these studies has also raised a host of interesting issues. It has become apparent, for example, that the folding and unfolding of proteins is associated with a whole range of cellular processes from the trafficking of molecules to specific organelles to the regulation of the cell cycle and the immune response. Such observations led to the inevitable conclusion that the failure to fold correctly, or to remain correctly folded, gives rise to many different types of biological malfunctions and hence to many different forms of disease [2]. In addition, it has been recognised recently that a large number of eukaryotic genes code for proteins that appear to be ‘natively unfolded’, and that proteins can adopt, under certain circumstances, highly organised multi-molecular assemblies whose structures are not specifically encoded in the amino acid sequence. Both these observations have raised challenging questions about one of the most fundamental principles of biology: the close relationship between the sequence, structure and function of proteins, as we discuss below [3].
It is well established that proteins that are ‘misfolded’, i.e. that are not in their functionally relevant conformation, are devoid of normal biological activity. In addition, they often aggregate and/or interact inappropriately with other cellular components leading to impairment of cell viability and eventually to cell death. Many diseases, often known as misfolding or conformational diseases, ultimately result from the presence in a living system of protein molecules with structures that are ‘incorrect’, i.e. that differ from those in normally functioning organisms [4]. Such diseases include conditions in which a specific protein, or protein complex, fails to fold correctly (e.g. cystic fibrosis, Marfan syndrome, amyotonic lateral sclerosis) or is not sufficiently stable to perform its normal function (e.g. many forms of cancer). They also include conditions in which aberrant folding behaviour results in the failure of a protein to be correctly trafficked (e.g. familial hypercholesterolaemia, α1-antitrypsin deficiency, and some forms of retinitis pigmentosa) [4]. The tendency of proteins to aggregate, often to give species extremely intractable to dissolution and refolding, is of course also well known in other circumstances. Examples include the formation of inclusion bodies during overexpression of heterologous proteins in bacteria and the precipitation of proteins during laboratory purification procedures. Indeed, protein aggregation is well established as one of the major difficulties associated with the production and handling of proteins in the biotechnology and pharmaceutical industries [5].
Considerable attention is presently focused on a group of protein folding diseases known as amyloidoses. In these diseases specific peptides or proteins fail to fold or to remain correctly folded and then aggregate (often with other components) so as to give rise to ‘amyloid’ deposits in tissue. Amyloid structures can be recognised because they possess a series of specific tinctorial and biophysical characteristics that reflect a common core structure based on the presence of highly organised βsheets [6]. The deposits in strictly defined amyloidoses are extracellular and can often be observed as thread-like fibrillar structures, sometimes assembled further into larger aggregates or plaques. These diseases include a range of sporadic, familial or transmissible degenerative diseases, some of which affect the brain and the central nervous system (e.g. Alzheimer’s and Creutzfeldt-Jakob diseases), while others involve peripheral tissues and organs such as the liver, heart and spleen (e.g. systemic amyloidoses and type II diabetes) [7, 8]. In other forms of amyloidosis, such as primary or secondary systemic amyloidoses, proteinaceous deposits are found in skeletal tissue and joints (e.g. haemodialysis-related amyloidosis) as well as in several organs (e.g. heart and kidney). Yet other components such as collagen, glycosaminoglycans and proteins (e.g. serum amyloid protein) are often present in the deposits protecting them against degradation [9, 10, 11]. Similar deposits to those in the amyloidoses are, however, found intracellularly in other diseases; these can be localised either in the cytoplasm, in the form of specialised aggregates known as aggresomes or as Lewy or Russell bodies or in the nucleus (see below).
The presence in tissue of proteinaceous deposits is a hallmark of all these diseases, suggesting a causative link between aggregate formation and pathological symptoms (often known as the amyloid hypothesis) [7, 8, 12]. At the present time the link between amyloid formation and disease is widely accepted on the basis of a large number of biochemical and genetic studies. The specific nature of the pathogenic species, and the molecular basis of their ability to damage cells, are however, the subject of intense debate [13, 14, 15, 16, 17, 18, 19, 20]. In neurodegenerative disorders it is very likely that the impairment of cellular function follows directly from the interactions of the aggregated proteins with cellular components [21, 22]. In the systemic non-neurological diseases, however, it is widely believed that the accumulation in vital organs of large amounts of amyloid deposits can by itself cause at least some of the clinical symptoms [23]. It is quite possible, however, that there are other more specific effects of aggregates on biochemical processes even in these diseases. The presence of extracellular or intracellular aggregates of a specific polypeptide molecule is a characteristic of all the 20 or so recognised amyloid diseases. The polypeptides involved include full length proteins (e.g. lysozyme or immunoglobulin light chains), biological peptides (amylin, atrial natriuretic factor) and fragments of larger proteins produced as a result of specific processing (e.g. the Alzheimer βpeptide) or of more general degradation [e.g. poly(Q) stretches cleaved from proteins with poly(Q) extensions such as huntingtin, ataxins and the androgen receptor]. The peptides and proteins associated with known amyloid diseases are listed in Table 1. In some cases the proteins involved have wild type sequences, as in sporadic forms of the diseases, but in other cases these are variants resulting from genetic mutations associated with familial forms of the diseases. In some cases both sporadic and familial diseases are associated with a given protein; in this case the mutational variants are usually associated with early-onset forms of the disease. In the case of the neurodegenerative diseases associated with the prion protein some forms of the diseases are transmissible. The existence of familial forms of a number of amyloid diseases has provided significant clues to the origins of the pathologies. For example, there are increasingly strong links between the age at onset of familial forms of disease and the effects of the mutations involved on the propensity of the affected proteins to aggregate in vitro. Such findings also support the link between the process of aggregation and the clinical manifestations of disease [24, 25].
The presence in cells of misfolded or aggregated proteins triggers a complex biological response. In the cytosol, this is referred to as the ‘heat shock response’ and in the endoplasmic reticulum (ER) it is known as the ‘unfolded protein response’. These responses lead to the expression, among others, of the genes for heat shock proteins (Hsp, or molecular chaperone proteins) and proteins involved in the ubiquitin-proteasome pathway [26]. The evolution of such complex biochemical machinery testifies to the fact that it is necessary for cells to isolate and clear rapidly and efficiently any unfolded or incorrectly folded protein as soon as it appears. In itself this fact suggests that these species could have a generally adverse effect on cellular components and cell viability. Indeed, it was a major step forward in understanding many aspects of cell biology when it was recognised that proteins previously associated only with stress, such as heat shock, are in fact crucial in the normal functioning of living systems. This advance, for example, led to the discovery of the role of molecular chaperones in protein folding and in the normal ‘housekeeping’ processes that are inherent in healthy cells [27, 28]. More recently a number of degenerative diseases, both neurological and systemic, have been linked to, or shown to be affected by, impairment of the ubiquitin-proteasome pathway (Table 2). The diseases are primarily associated with a reduction in either the expression or the biological activity of Hsps, ubiquitin, ubiquitinating or deubiquitinating enzymes and the proteasome itself, as we show below [29, 30, 31, 32], or even to the failure of the quality control mechanisms that ensure proper maturation of proteins in the ER. The latter normally leads to degradation of a significant proportion of polypeptide chains before they have attained their native conformations through retrograde translocation to the cytosol [33, 34].
….
It is now well established that the molecular basis of protein aggregation into amyloid structures involves the existence of ‘misfolded’ forms of proteins, i.e. proteins that are not in the structures in which they normally function in vivo or of fragments of proteins resulting from degradation processes that are inherently unable to fold [4, 7, 8, 36]. Aggregation is one of the common consequences of a polypeptide chain failing to reach or maintain its functional three-dimensional structure. Such events can be associated with specific mutations, misprocessing phenomena, aberrant interactions with metal ions, changes in environmental conditions, such as pH or temperature, or chemical modification (oxidation, proteolysis). Perturbations in the conformational properties of the polypeptide chain resulting from such phenomena may affect equilibrium 1 in Fig. 1 increasing the population of partially unfolded, or misfolded, species that are much more aggregation-prone than the native state.
Fig. 1 Overview of the possible fates of a newly synthesised polypeptide chain. The equilibrium ① between the partially folded molecules and the natively folded ones is usually strongly in favour of the latter except as a result of specific mutations, chemical modifications or partially destabilising solution conditions. The increased equilibrium populations of molecules in the partially or completely unfolded ensemble of structures are usually degraded by the proteasome; when this clearance mechanism is impaired, such species often form disordered aggregates or shift equilibrium ② towards the nucleation of pre-fibrillar assemblies that eventually grow into mature fibrils (equilibrium ③). DANGER! indicates that pre-fibrillar aggregates in most cases display much higher toxicity than mature fibrils. Heat shock proteins (Hsp) can suppress the appearance of pre-fibrillar assemblies by minimising the population of the partially folded molecules by assisting in the correct folding of the nascent chain and the unfolded protein response target incorrectly folded proteins for degradation.
……
Little is known at present about the detailed arrangement of the polypeptide chains themselves within amyloid fibrils, either those parts involved in the core βstrands or in regions that connect the various β-strands. Recent data suggest that the sheets are relatively untwisted and may in some cases at least exist in quite specific supersecondary structure motifs such as β-helices [6, 40] or the recently proposed µ-helix [41]. It seems possible that there may be significant differences in the way the strands are assembled depending on characteristics of the polypeptide chain involved [6, 42]. Factors including length, sequence (and in some cases the presence of disulphide bonds or post-translational modifications such as glycosylation) may be important in determining details of the structures. Several recent papers report structural models for amyloid fibrils containing different polypeptide chains, including the Aβ40 peptide, insulin and fragments of the prion protein, based on data from such techniques as cryo-electron microscopy and solid-state magnetic resonance spectroscopy [43, 44]. These models have much in common and do indeed appear to reflect the fact that the structures of different fibrils are likely to be variations on a common theme [40]. It is also emerging that there may be some common and highly organised assemblies of amyloid protofilaments that are not simply extended threads or ribbons. It is clear, for example, that in some cases large closed loops can be formed [45, 46, 47], and there may be specific types of relatively small spherical or ‘doughnut’ shaped structures that can result in at least some circumstances (see below).
…..
The similarity of some early amyloid aggregates with the pores resulting from oligomerisation of bacterial toxins and pore-forming eukaryotic proteins (see below) also suggest that the basic mechanism of protein aggregation into amyloid structures may not only be associated with diseases but in some cases could result in species with functional significance. Recent evidence indicates that a variety of micro-organisms may exploit the controlled aggregation of specific proteins (or their precursors) to generate functional structures. Examples include bacterial curli [52] and proteins of the interior fibre cells of mammalian ocular lenses, whose β-sheet arrays seem to be organised in an amyloid-like supramolecular order [53]. In this case the inherent stability of amyloid-like protein structure may contribute to the long-term structural integrity and transparency of the lens. Recently it has been hypothesised that amyloid-like aggregates of serum amyloid A found in secondary amyloidoses following chronic inflammatory diseases protect the host against bacterial infections by inducing lysis of bacterial cells [54]. One particularly interesting example is a ‘misfolded’ form of the milk protein α-lactalbumin that is formed at low pH and trapped by the presence of specific lipid molecules [55]. This form of the protein has been reported to trigger apoptosis selectively in tumour cells providing evidence for its importance in protecting infants from certain types of cancer [55]. ….
Amyloid formation is a generic property of polypeptide chains ….
It is clear that the presence of different side chains can influence the details of amyloid structures, particularly the assembly of protofibrils, and that they give rise to the variations on the common structural theme discussed above. More fundamentally, the composition and sequence of a peptide or protein affects profoundly its propensity to form amyloid structures under given conditions (see below).
Because the formation of stable protein aggregates of amyloid type does not normally occur in vivo under physiological conditions, it is likely that the proteins encoded in the genomes of living organisms are endowed with structural adaptations that mitigate against aggregation under these conditions. A recent survey involving a large number of structures of β-proteins highlights several strategies through which natural proteins avoid intermolecular association of β-strands in their native states [65]. Other surveys of protein databases indicate that nature disfavours sequences of alternating polar and nonpolar residues, as well as clusters of several consecutive hydrophobic residues, both of which enhance the tendency of a protein to aggregate prior to becoming completely folded [66, 67].
……
Precursors of amyloid fibrils can be toxic to cells
It was generally assumed until recently that the proteinaceous aggregates most toxic to cells are likely to be mature amyloid fibrils, the form of aggregates that have been commonly detected in pathological deposits. It therefore appeared probable that the pathogenic features underlying amyloid diseases are a consequence of the interaction with cells of extracellular deposits of aggregated material. As well as forming the basis for understanding the fundamental causes of these diseases, this scenario stimulated the exploration of therapeutic approaches to amyloidoses that focused mainly on the search for molecules able to impair the growth and deposition of fibrillar forms of aggregated proteins. ….
Structural basis and molecular features of amyloid toxicity
The presence of toxic aggregates inside or outside cells can impair a number of cell functions that ultimately lead to cell death by an apoptotic mechanism [95, 96]. Recent research suggests, however, that in most cases initial perturbations to fundamental cellular processes underlie the impairment of cell function induced by aggregates of disease-associated polypeptides. Many pieces of data point to a central role of modifications to the intracellular redox status and free Ca2+ levels in cells exposed to toxic aggregates [45, 89, 97, 98, 99, 100, 101]. A modification of the intracellular redox status in such cells is associated with a sharp increase in the quantity of reactive oxygen species (ROS) that is reminiscent of the oxidative burst by which leukocytes destroy invading foreign cells after phagocytosis. In addition, changes have been observed in reactive nitrogen species, lipid peroxidation, deregulation of NO metabolism [97], protein nitrosylation [102] and upregulation of heme oxygenase-1, a specific marker of oxidative stress [103]. ….
Results have recently been reported concerning the toxicity towards cultured cells of aggregates of poly(Q) peptides which argues against a disease mechanism based on specific toxic features of the aggregates. These results indicate that there is a close relationship between the toxicity of proteins with poly(Q) extensions and their nuclear localisation. In addition they support the hypotheses that the toxicity of poly(Q) aggregates can be a consequence of altered interactions with nuclear coactivator or corepressor molecules including p53, CBP, Sp1 and TAF130 or of the interaction with transcription factors and nuclear coactivators, such as CBP, endowed with short poly(Q) stretches ([95] and references therein)…..
Concluding remarks
The data reported in the past few years strongly suggest that the conversion of normally soluble proteins into amyloid fibrils and the toxicity of small aggregates appearing during the early stages of the formation of the latter are common or generic features of polypeptide chains. Moreover, the molecular basis of this toxicity also appears to display common features between the different systems that have so far been studied. The ability of many, perhaps all, natural polypeptides to ‘misfold’ and convert into toxic aggregates under suitable conditions suggests that one of the most important driving forces in the evolution of proteins must have been the negative selection against sequence changes that increase the tendency of a polypeptide chain to aggregate. Nevertheless, as protein folding is a stochastic process, and no such process can be completely infallible, misfolded proteins or protein folding intermediates in equilibrium with the natively folded molecules must continuously form within cells. Thus mechanisms to deal with such species must have co-evolved with proteins. Indeed, it is clear that misfolding, and the associated tendency to aggregate, is kept under control by molecular chaperones, which render the resulting species harmless assisting in their refolding, or triggering their degradation by the cellular clearance machinery [166, 167, 168, 169, 170, 171, 172, 173, 175, 177, 178].
Misfolded and aggregated species are likely to owe their toxicity to the exposure on their surfaces of regions of proteins that are buried in the interior of the structures of the correctly folded native states. The exposure of large patches of hydrophobic groups is likely to be particularly significant as such patches favour the interaction of the misfolded species with cell membranes [44, 83, 89, 90, 91, 93]. Interactions of this type are likely to lead to the impairment of the function and integrity of the membranes involved, giving rise to a loss of regulation of the intracellular ion balance and redox status and eventually to cell death. In addition, misfolded proteins undoubtedly interact inappropriately with other cellular components, potentially giving rise to the impairment of a range of other biological processes. Under some conditions the intracellular content of aggregated species may increase directly, due to an enhanced propensity of incompletely folded or misfolded species to aggregate within the cell itself. This could occur as the result of the expression of mutational variants of proteins with decreased stability or cooperativity or with an intrinsically higher propensity to aggregate. It could also occur as a result of the overproduction of some types of protein, for example, because of other genetic factors or other disease conditions, or because of perturbations to the cellular environment that generate conditions favouring aggregation, such as heat shock or oxidative stress. Finally, the accumulation of misfolded or aggregated proteins could arise from the chaperone and clearance mechanisms becoming overwhelmed as a result of specific mutant phenotypes or of the general effects of ageing [173, 174].
The topics discussed in this review not only provide a great deal of evidence for the ‘new view’ that proteins have an intrinsic capability of misfolding and forming structures such as amyloid fibrils but also suggest that the role of molecular chaperones is even more important than was thought in the past. The role of these ubiquitous proteins in enhancing the efficiency of protein folding is well established [185]. It could well be that they are at least as important in controlling the harmful effects of misfolded or aggregated proteins as in enhancing the yield of functional molecules.
Nutritional Status is Associated with Faster Cognitive Decline and Worse Functional Impairment in the Progression of Dementia: The Cache County Dementia Progression Study1
Nutritional status may be a modifiable factor in the progression of dementia. We examined the association of nutritional status and rate of cognitive and functional decline in a U.S. population-based sample. Study design was an observational longitudinal study with annual follow-ups up to 6 years of 292 persons with dementia (72% Alzheimer’s disease, 56% female) in Cache County, UT using the Mini-Mental State Exam (MMSE), Clinical Dementia Rating Sum of Boxes (CDR-sb), and modified Mini Nutritional Assessment (mMNA). mMNA scores declined by approximately 0.50 points/year, suggesting increasing risk for malnutrition. Lower mMNA score predicted faster rate of decline on the MMSE at earlier follow-up times, but slower decline at later follow-up times, whereas higher mMNA scores had the opposite pattern (mMNA by time β= 0.22, p = 0.017; mMNA by time2 β= –0.04, p = 0.04). Lower mMNA score was associated with greater impairment on the CDR-sb over the course of dementia (β= 0.35, p < 0.001). Assessment of malnutrition may be useful in predicting rates of progression in dementia and may provide a target for clinical intervention.
Shared Genetic Risk Factors for Late-Life Depression and Alzheimer’s Disease
Background: Considerable evidence has been reported for the comorbidity between late-life depression (LLD) and Alzheimer’s disease (AD), both of which are very common in the general elderly population and represent a large burden on the health of the elderly. The pathophysiological mechanisms underlying the link between LLD and AD are poorly understood. Because both LLD and AD can be heritable and are influenced by multiple risk genes, shared genetic risk factors between LLD and AD may exist. Objective: The objective is to review the existing evidence for genetic risk factors that are common to LLD and AD and to outline the biological substrates proposed to mediate this association. Methods: A literature review was performed. Results: Genetic polymorphisms of brain-derived neurotrophic factor, apolipoprotein E, interleukin 1-beta, and methylenetetrahydrofolate reductase have been demonstrated to confer increased risk to both LLD and AD by studies examining either LLD or AD patients. These results contribute to the understanding of pathophysiological mechanisms that are common to both of these disorders, including deficits in nerve growth factors, inflammatory changes, and dysregulation mechanisms involving lipoprotein and folate. Other conflicting results have also been reviewed, and few studies have investigated the effects of the described polymorphisms on both LLD and AD. Conclusion: The findings suggest that common genetic pathways may underlie LLD and AD comorbidity. Studies to evaluate the genetic relationship between LLD and AD may provide insights into the molecular mechanisms that trigger disease progression as the population ages.
Association of Vitamin B12, Folate, and Sulfur Amino Acids With Brain Magnetic Resonance Imaging Measures in Older Adults: A Longitudinal Population-Based Study
Importance Vitamin B12, folate, and sulfur amino acids may be modifiable risk factors for structural brain changes that precede clinical dementia.
Objective To investigate the association of circulating levels of vitamin B12, red blood cell folate, and sulfur amino acids with the rate of total brain volume loss and the change in white matter hyperintensity volume as measured by fluid-attenuated inversion recovery in older adults.
Design, Setting, and Participants The magnetic resonance imaging subsample of the Swedish National Study on Aging and Care in Kungsholmen, a population-based longitudinal study in Stockholm, Sweden, was conducted in 501 participants aged 60 years or older who were free of dementia at baseline. A total of 299 participants underwent repeated structural brain magnetic resonance imaging scans from September 17, 2001, to December 17, 2009.
Main Outcomes and Measures The rate of brain tissue volume loss and the progression of total white matter hyperintensity volume.
Results In the multi-adjusted linear mixed models, among 501 participants (300 women [59.9%]; mean [SD] age, 70.9 [9.1] years), higher baseline vitamin B12 and holotranscobalamin levels were associated with a decreased rate of total brain volume loss during the study period: for each increase of 1 SD, β (SE) was 0.048 (0.013) for vitamin B12 (P < .001) and 0.040 (0.013) for holotranscobalamin (P = .002). Increased total homocysteine levels were associated with faster rates of total brain volume loss in the whole sample (β [SE] per 1-SD increase, –0.035 [0.015]; P = .02) and with the progression of white matter hyperintensity among participants with systolic blood pressure greater than 140 mm Hg (β [SE] per 1-SD increase, 0.000019 [0.00001]; P = .047). No longitudinal associations were found for red blood cell folate and other sulfur amino acids.
Conclusions and Relevance This study suggests that both vitamin B12 and total homocysteine concentrations may be related to accelerated aging of the brain. Randomized clinical trials are needed to determine the importance of vitamin B12supplementation on slowing brain aging in older adults.
Notes from Kurzweill
This vitamin stops the aging process in organs, say Swiss researchers
A potential breakthrough for regenerative medicine, pending further studies
Improved muscle stem cell numbers and muscle function in NR-treated aged mice: Newly regenerated muscle fibers 7 days after muscle damage in aged mice (left: control group; right: fed NR). (Scale bar = 50 μm). (credit: Hongbo Zhang et al./Science) http://www.kurzweilai.net/images/improved-muscle-fibers.png
EPFL researchers have restored the ability of mice organs to regenerate and extend life by simply administering nicotinamide riboside (NR) to them.
NR has been shown in previous studies to be effective in boosting metabolism and treating a number of degenerative diseases. Now, an article by PhD student Hongbo Zhang published in Science also describes the restorative effects of NR on the functioning of stem cells for regenerating organs.
As in all mammals, as mice age, the regenerative capacity of certain organs (such as the liver and kidneys) and muscles (including the heart) diminishes. Their ability to repair them following an injury is also affected. This leads to many of the disorders typical of aging.
Mitochondria —> stem cells —> organs
To understand how the regeneration process deteriorates with age, Zhang teamed up with colleagues from ETH Zurich, the University of Zurich, and universities in Canada and Brazil. By using several biomarkers, they were able to identify the molecular chain that regulates how mitochondria — the “powerhouse” of the cell — function and how they change with age. “We were able to show for the first time that their ability to function properly was important for stem cells,” said Auwerx.
Under normal conditions, these stem cells, reacting to signals sent by the body, regenerate damaged organs by producing new specific cells. At least in young bodies. “We demonstrated that fatigue in stem cells was one of the main causes of poor regeneration or even degeneration in certain tissues or organs,” said Zhang.
How to revitalize stem cells
Which is why the researchers wanted to “revitalize” stem cells in the muscles of elderly mice. And they did so by precisely targeting the molecules that help the mitochondria to function properly. “We gave nicotinamide riboside to 2-year-old mice, which is an advanced age for them,” said Zhang.
“This substance, which is close to vitamin B3, is a precursor of NAD+, a molecule that plays a key role in mitochondrial activity. And our results are extremely promising: muscular regeneration is much better in mice that received NR, and they lived longer than the mice that didn’t get it.”
Parallel studies have revealed a comparable effect on stem cells of the brain and skin. “This work could have very important implications in the field of regenerative medicine,” said Auwerx. This work on the aging process also has potential for treating diseases that can affect — and be fatal — in young people, like muscular dystrophy (myopathy).
So far, no negative side effects have been observed following the use of NR, even at high doses. But while it appears to boost the functioning of all cells, it could include pathological ones, so further in-depth studies are required.
Abstract of NAD+ repletion improves mitochondrial and stem cell function and enhances life span in mice
Adult stem cells (SCs) are essential for tissue maintenance and regeneration yet are susceptible to senescence during aging. We demonstrate the importance of the amount of the oxidized form of cellular nicotinamide adenine dinucleotide (NAD+) and its impact on mitochondrial activity as a pivotal switch to modulate muscle SC (MuSC) senescence. Treatment with the NAD+ precursor nicotinamide riboside (NR) induced the mitochondrial unfolded protein response (UPRmt) and synthesis of prohibitin proteins, and this rejuvenated MuSCs in aged mice. NR also prevented MuSC senescence in the Mdx mouse model of muscular dystrophy. We furthermore demonstrate that NR delays senescence of neural SCs (NSCs) and melanocyte SCs (McSCs), and increased mouse lifespan. Strategies that conserve cellular NAD+ may reprogram dysfunctional SCs and improve lifespan in mammals.
Discriminating the gene target of a distal regulatory element from other nearby transcribed genes is a challenging problem with the potential to illuminate the causal underpinnings of complex diseases. We present TargetFinder, a computational method that reconstructs regulatory landscapes from diverse features along the genome. The resulting models accurately predict individual enhancer–promoter interactions across multiple cell lines with a false discovery rate up to 15 times smaller than that obtained using the closest gene. By evaluating the genomic features driving this accuracy, we uncover interactions between structural proteins, transcription factors, epigenetic modifications, and transcription that together distinguish interacting from non-interacting enhancer–promoter pairs. Most of this signature is not proximal to the enhancers and promoters but instead decorates the looping DNA. We conclude that complex but consistent combinations of marks on the one-dimensional genome encode the three-dimensional structure of fine-scale regulatory interactions.
On observing schizophrenia from a clinical point of view up to its molecular basis, one may conclude that this is likely to be one of the most complex human disorders to be characterized in all aspects. Such complexity is the reflex of an intricate combination of genetic and environmental components that influence brain functions since pre-natal neurodevelopment, passing by brain maturation, up to the onset of disease and disease establishment. The perfect function of tissues, organs, systems, and finally the organism depends heavily on the proper functioning of cells. Several lines of evidence, including genetics, genomics, transcriptomics, neuropathology, and pharmacology, have supported the idea that dysfunctional cells are causative to schizophrenia. Together with the above-mentioned techniques, proteomics have been contributing to understanding the biochemical basis of schizophrenia at the cellular and tissue level through the identification of differentially expressed proteins and consequently their biochemical pathways, mostly in the brain tissue but also in other cells. In addition, mass spectrometry-based proteomics have identified and precisely quantified proteins that may serve as biomarker candidates to prognosis, diagnosis, and medication monitoring in peripheral tissue. Here, we review all data produced by proteomic investigation in the last 5 years using tissue and/or cells from schizophrenic patients, focusing on postmortem brain tissue and peripheral blood serum and plasma. This information has provided integrated pictures of the biochemical systems involved in the pathobiology, and has suggested potential biomarkers, and warrant potential targets to alternative treatment therapies to schizophrenia.
Schizophrenia is a complex neuropsychiatric disorder that produces severe symptoms and significant lifelong disability, causing massive personal and societal burden.1,2 About 1% of the world’s population is affected by schizophrenia.3 Despite the strong genetic component, showing increasing risks for those related to schizophrenic patients,4 and the known role of environment as a trigger, schizophrenia signs and symptoms have unknown etiology. Currently, the disease diagnosis is essentially clinically defined by observed signs of psychosis, which often include paranoid delusions and auditory hallucinations,5 with onset during late adolescence and/or early adulthood.
Pharmacological treatments are available for schizophrenia; yet, most of the currently used antipsychotic medications were discovered in the 1950s, or are a variation of those medications, and since then no new major drug class has been introduced to the clinic. In addition, efficacy of medication is poor, and only about 40% of schizophrenic patients respond effectively to initial treatment with antipsychotics.6,7 Unfortunately, comprehensive studies on molecular mechanisms of schizophrenia have been scant; hence, current treatments are only partly beneficial to a subset of symptoms. The response to drugs is heterogenous, mainly because of individual variations of the disease, in addition to scarce knowledge on its pathophysiology, impairing both diagnosis and adequate treatment selection.8,9
Heterogenic and multifactorial aspects of schizophrenia have always hindered biochemical characterization studies and delayed the establishment of preclinical models of the disease.10 Several studies, including postmortem, imaging, pharmacological, and genetic studies, reported common traces of the disease, such as synaptic deficits, abnormal neural network, and changes in neurotransmission, involving dopamine, glutamate, and gamma-aminobutyric acid.2,11,12,13 Additional abnormalities, such as aberrant inflammatory responses, oligodendrocyte alterations, epigenetic changes, mitochondrial dysfunction, and reactive oxygen species (ROS) imbalance, are often described in schizophrenia.14,15,16
A complex cross talk between genetic and environmental factors during neurogenesis is responsible for promoting differences of gene and protein expression in schizophrenia, causing abnormal processes during neurodevelopment.2 Recent studies found reinforcement of genes associated with the major hypotheses of glutamatergic neurotransmission, such as DRD2 (dopamine receptor D2)—the main target of antipsychotic drugs17—among other potential targets, involving perturbation of specific neurotransmitter systems or pathways, which are yet to be studied. The complexity of schizophrenia reinforces the need to unravel molecular mechanisms, as those insights have been shown to be essential in identifying and validating drug targets and biomarkers.9 Therefore, unraveling models with relevance to the cause and onset of schizophrenia is essential toward improving treatments and outcomes for those with the disorder.
Here we review the advances of proteomics on schizophrenia research, toward a better understanding of disease mechanisms and response to treatment, and the efforts toward the discovery of biomarkers for diagnosis and disease evolution.
The role of proteomics in schizophrenia research
In the past century, psychiatric research was dedicated to understanding the nature of several disorders, including action of psychotherapeutics. It was also shown that schizophrenia is a highly heritable disease, indicating a strong genetic influence and an estimated heritability of 80–85%,18,19 more likely with a polygenic basis.20 Since the beginning of the twenty-first century, revolution of genomic technologies has allowed a deeper understanding of the genetic basis of diseases, and several genetic findings on psychiatric disorders have been reported,21 unraveling candidate genes linked to risk factors of psychiatric disorders, such as DISC1 (disrupted in schizophrenia 1),22 involved in neuronal development and synapse formation.23,24 In fact, the International Schizophrenia Consortium (ISC) found indication for a polygenic contribution to schizophrenia.25 While candidate gene studies are beneficial, in cases with a not yet well-understood biology, such as schizophrenia, a single gene only adds a small phenotype effect to the multifactorial etiology of the disease.20,26,27
Since 2008, genomic technology innovations have led to a better understanding of psychiatric disorders, providing information about numerous genes that have a role in brain development.21 Recent advances of next-generation sequencing have facilitated a higher coverage and sample throughput of schizophrenia studies.28,29,30Furthermore, international collaborations, which increased the number of participant subjects and samples, have combined efforts to provide deeper insight from comprehensive biological data sets, such as the Psychiatric GWAS (genome-wide association studies) Consortium (PGC; http://pgc.unc.edu).31 Most recently, two main studies, reporting comprehensive GWAS analysis, were able to identify 13 (ref. 27) and 108 schizophrenia-associated risk loci,17 the latter being the largest GWAS study on schizophrenia to date, with up to 36,989 cases and 113,075 controls. Unbiased GWAS,17,27,32,33 indicating genetic regions (loci) that contribute to disease susceptibility, and structural variation studies, such as copy number variants,30,34 are the main identification sources of gene variants with small effects on disease phenotype.35 For instance, copy number variants, including deletions and duplications of several DNA segments, confer significant risk increase in alleles of schizophrenia genome up to 10–25-fold.9,34,36Several of those findings support the leading etiological hypothesis of the disorder, and point to functionally related targets, such as DRD2, miRNA-137, N-methyl-D-aspartate receptor (NMDAR) complex, or calcium channel subunits.17,30,36,37 Information on genetic variations as a base will increase knowledge on mechanisms of schizophrenia and other psychiatric disorders.
Deciphering the human genome was a revolution in genetics, and the anticipated next step was to decode RNA complexity to understand how information was delivered, and its variety between individuals. Development of large-scale transcriptome analyses, such as cDNA microarrays, Serial Analysis of Gene Expression, and the analyses of Expressed Sequence Tag, and more recently the advance of whole transcriptome shotgun sequencing (or RNA-Seq), providing the presence and quantification of RNA at a given time in a genome, allowed a deeper insight into the dynamics of an organism. Transcriptome analyses revealed RNA implication in psychiatric diseases,38,39 including abnormalities resulting from alternative splicing, in addition to messenger RNA transcripts, such as total RNA and small RNA, including micro-RNA.40 Those abnormalities were observed in several biological processes, such as synaptic and mitochondrial/energetic function,41,42,43 cytoskeleton,44 immune and inflammation response,45,46,47 and the myelination pathway.48 Although not yet fully understood, the more the pieces of the puzzle discovered, the more comprehensive the pathology network becomes.
Genomic and transcriptomic studies generated significant data, although these changes cannot yet be translated into biomarkers. The main limitation of genetic approaches in schizophrenia is extrapolation to functional protein expression, as proteins undergo several modifications from transcription to posttranslation, and transcript abundance cannot really predict protein levels either in normal conditions or in response to stress, such as diseases.49,50 Therefore, proteomic techniques are being increasingly used in screening for identification of biomarkers in schizophrenia,51,52 providing several insights into the pathophysiology of the disease. Proteomics can show global expression of proteins or protein groups, and is more complex than genomics as it can change from each cell type at any given time or state.49 Also a high-throughput method, proteomic studies detect fewer expressed proteins than a transcriptomic detects expressed genes, but protein expression provides a precise functional profile and presents an unbiased current physiological state as a reflex of the complex interaction of gene versus environment. The importance of those interactions has been increasing in the research of schizophrenia and other neurological diseases.10,41,53
Regarding research into schizophrenia, numerous studies have investigated the proteome of postmortem brain tissue, including several brain regions such as the dorsolateral prefrontal cortex,41,54,55 frontal cortex,56thalamus,57 anterior cingulate cortex,58,59,60hippocampus,61,62 corpus callosum,63 and insular cortex.64 Postmortem brain tissue has yielded many valuable insights into the pathophysiology of schizophrenia, but less information on disease onset and development. Thus, other tissues and cells have been tested, providing data from naive patients as well, such as from cerebrospinal fluid (CSF),65,66,67,68blood serum and plasma,69,70,71,72,73 liver,68,74 and fibroblasts,75 which can be biopsied from living patients, among others,76,77aiming to reveal more about potential biomarkers of discovery and monitoring of the disease.
Proteomic methodologies used in schizophrenia research
A proteome comprises the entire set of proteins in a biological system (cell, tissue, or organism) in a particular state, at a given time.78 The need to understand all proteins derived from almost 20,000 genes identified by the Human Genome Project turns molecular biology studies toward proteomics. Because of the progress of mass spectrometry techniques, more fine and high-throughput methods are available, supporting the identification of hundreds (or thousands) of proteins in a single biological sample. In 2014, two major consortiums have delivered a draft of the human proteome,79,80 with a large-scale data set covering 84–92% of the protein-coded genes annotated for the human genome. The more information annotated on protein knowledge databases, the more unknown causes of diseases and biomarker identification can be performed.
In the first decade of proteomics, the main quantitative methods used were gel-based, such as two-dimensional gel electrophoresis (2DE), including the fluorescent two-dimensional differential gel electrophoresis (2D-DIGE). Despite its recognized usefulness,81 gel-based techniques have been consistently replaced by gel-free techniques with the introduction of the concept of shotgun proteomics, which employs basically liquid chromatography followed by mass spectrometry (LC/MS).82 The large scale was possible only because of the development of proteomics based on mass spectrometry, which offers insights into protein abundance, expression profiles according to cell type, posttranslational modifications, and protein–protein interactions, and the possibility to study modifications at the protein level.83
2DE was first described in 1975,84 and after intense enhancements in the 1980s85,86 it became widely used in the separation of complex protein mixtures according to their isoelectric point, in the first dimension by isoelectric focusing, and according to their molecular weight (MW), in the second dimension, by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. This separation leads to a protein profile comprising several spots, each of which, in theory, represents a single protein, providing information about intact proteins and isoforms. Protein visualization techniques include common post-run methods, such as Coomassie blue or silver staining, and also pre-labeling of samples with fluorescent dies, such as in the 2D-DIGE.87 Image analysis of the latter provides a more sensitive quantification method, as up to 10-fold lower amount of samples can be applied. In addition, 2D-DIGE allows co-running of different samples in the same gel, labeled with distinct fluorescent dies (i.e., Cy3, Cy2, Cy5), and might also include an internal control for cross-gel comparison purposes. Those techniques have significantly improved in the previous years with respect to reproducibility and robustness, allowing better comparison between samples and across different laboratories.
Furthermore, mass spectrometry (MS) revolutionized proteomic studies when combined with the 2DE/2D-DIGE workflow, improving sensitivity for identification of differentially expressed proteins, by measuring molecular mass-to-charge ratio of ions (m/z).88 Protein spots, excised from the gel, are digested (i.e., trypsin) and masses of these peptides measured on MS instruments, providing a peptide mass fingerprint of each protein, which is then compared with an in silico-digested database. Further fragmentation of each peptide, performed on an MS/MS instrument, provides the sequence of that peptide, assisting in protein identification. Disadvantages of 2DE/MS combination include the incompatibility to very low or very high molecular weight or isoelectric point, in addition to those proteins with low abundance, which will not be spotted.89 Nevertheless, several proteome studies in schizophrenia were performed using proteomic screening approaches such as 2DE/2D-DIGE, providing large-scale data on the pathophysiology of the disease.41,55,56,58,61,66,90
Hence, schizophrenia and other psychiatric disorder studies have intensively used shotgun proteomics for the analysis of peptides and proteins for profiling, and for quantification of protein modification analysis.54,59,72,75,77,91 For shotgun proteomics, proteins are first digested into peptides (i.e., using trypsin, as previously), which are next separated by high-performance liquid chromatography online-connected to a hybrid MS, providing a gel-free proteomic system (LC-MS/MS). Shotgun proteomics lead to the possibility of identifying more proteins, increasing sampling of low abundant and extreme-sized proteins.82 Most proteomic studies on schizophrenia have used label-free methods for quantification,59,75,92 which assume chromatographic peak areas correlated to the concentration of peptides,93and is one of the simplest ways to compare proteomics, allowing comparison among several samples at once.94Nevertheless, both in vitro and in vivo stable isotopic labeling methods are available in shotgun proteomics, for quantification accuracy of protein concentrations simultaneously in several biological samples. In vitroapproaches include isobaric tags for relative and absolute quantification95 and isotope-coded protein labeling, whereas in vivo metabolic methods, such as stable isotope labeling by amino acids in cell culture96 and stable isotope labeling in mammals, have been used in proteomic quantification.97 Some have been applied to neuropsychiatric disorders, in postmortem brains and CSF,54,57,98 and in animal and cell studies99,100 on proteomic research.
The power to identify and quantify proteins and protein sets at high resolution, among multiple samples, is essential to understand large case studies in biomedical research. Recent advances have been made in MS-based techniques, such as selected reaction monitoring/multiple reaction monitoring, which has just emerged as a promising technology for a more precise MS-based quantification of targeted protein,101,102 and was awarded Nature’s Method of the Year in 2012 on biological research methods.103 Selected reaction monitoring is specific, accurate, and sensitive, as it selects proteotypic peptides—those that uniquely identify the targeted protein—for its analysis, which might overcome several current validation issues, such as semiquantitative western blotting techniques, availability, and specificity.104 Furthermore, this ability to quantify specific proteins across several samples is particularly interesting with regard to biomarkers, as clinical validation of biomarker signatures for a given disease must be tested over a large sample set to achieve satisfactory statistical power. Indeed, proteomic studies in psychiatric disorders slowly start to validate pathways and biological functions that were found differentially expressed by selected reaction monitoring.105,106,107,108
Likewise, other proteomic methodologies have been extensively applied to schizophrenia research in order to discover and validate biomarkers, such as multiplex immunoassays,69 which use multiplexed dye-coded microspheres of selected protein sets, thus providing profile studies of cytokines, growth factors, or metabolic pathways, from blood serum or CSF samples.70,109,110,111Aiming to reach the broader spectrum of protein visualization, concerns regarding the possibility to obtain sub-proteomes (using fractionation methods)112 by depleting high-abundant proteins or enriching a group of proteins in a sample should be part of the design and technique choice. Protein separation and quantification using SELDI-TOF-MS ProteinChip analysis or metal ion affinity chromatography to select proteins from a mixture have been used in schizophrenia research lately.65,68,72 Regardless of the protein analysis method, study design and sample preparation choice are crucial steps in proteomic studies. Platforms using reduced number of analytes, but a broader number of clinical samples, provide a precise statistical interpretation.
Indeed, statistics and bioinformatics are of extreme importance for proteomic studies, as different types of assays (2DE, shotgun-MS, or multiplex immunoassays) are required to precisely quantify changes in expression of hundreds (or thousands) of proteins. Therefore, those fields are improving, together with the development of new tools and methods for proteomic analysis, offering better algorithms and image analysis tools, in order to provide a more robust analysis from the growing number of data generated.
What do proteomics tell us about schizophrenia?
Proteomic technologies, mostly focusing on mass spectrometric analysis, are a valuable tool in psychiatric research. A simple search on PubMed using the terms ‘proteomics or proteome and schizophrenia’ provides a total of 218 articles since the first article on proteomics of schizophrenia in the beginning of the 2000s.56 Out of them, 124 articles (and growing) were published within the last 5 years (2010–2014) on human and animal studies, including some reviews, showing considerable increase in awareness of the importance of proteomics in the study of schizophrenia. We have focused, for the purpose of the review, on proteomic studies on human samples of schizophrenia patients compared with controls, from the last 5 years.
These studies, which are summarized in Table 1, have been using proteomic screening approaches such as shotgun-MS (10/23), 2DE/DIGE (7/23), and multiplex immunoassays (10/23), alone or combined. Although postmortem brains are the main studied tissue in schizophrenia research,57,58,59,61,113 influences of chronic medication or sample heterogeneity and age have impaired some interpretation of the molecular differences found in postmortem brain tissue of schizophrenia patients compared with control subjects.114Thus, current studies have been mostly focusing on more accessible peripheral tissues, with a preference for blood serum and plasma,72,73,109,115 and CSF,65,66 although there are studies on skin fibroblasts75 and saliva as well.76Those have become the main tissues used in proteomic studies of schizophrenia because of the possibility of multiple sampling, thus providing better characterization of disease onset, development, and response to treatment. This broader characterization could lead to a more complete understanding of the disease and to development of diagnostic/prognostic biomarkers. Indeed, an analysis of proteins that are common to brain, CSF, and blood samples from at least two studies presented in Table 1, using Ingenuity Pathway Analysis (IPA, Ingenuity Systems, Qiagen, Redwood, CA, USA; www.ingenuity.com—Figure 1), shows biomarker candidates of psychiatric disorders and their interactions, and is further discussed.
Table 1: Human proteomic studies from the last 5 years of different tissues and cells in schizophrenic patients
Figure 1 Protein network of regulated proteins in schizophrenia brain, CSF, and blood samples, analyzed by ingenuity pathways knowledge database. ALDOC, aldolase C; CSF, cerebrospinal fluid; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Differentially expressed proteins in schizophrenia proteomic studies have been found to be involved in neuronal transmission, synaptic plasticity, and neurites outgrowth, including several cytoskeletal constituents. Most significant proteomic changes included downregulation of neuroreceptors such as NMDA receptors and alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA), in addition to glutamatergic signaling molecules, such as neurofilaments (NEFL and NEFM), glutamate-ammonia ligase (GLUL), and guanine nucleotide-binding proteins (G proteins) (GNB1), or dihydropyrimidinase-related protein 2 (DPYSL2), which are involved in synaptic function, axon guidance, and signal transduction impairment in schizophrenia.59,113NEFL, in addition to its role in neuronal morphogenesis, is directly associated with NMDA receptors. NMDAR hypofunction was associated with neurotransmitter dysfunction in NR1 transgenic mice,105 with variations in bioactive peptides and proteins. As GLUL is responsible for removing glutamate from neuronal synapses, it is most likely involved in glutamate imbalance in schizophrenia.2
Other proteins related to NMDA functionality and synaptic plasticity, such as MAPK3, SYNPO, CYFIP2, VDAC, CAMK2B, PRDX1, and ESYT, were also observed differentially expressed in postsynaptic density-enriched samples of postmortem brain tissues.59 Data corresponding to a genomic study of schizophrenia34found an excess of copy number variants in schizophrenia, confirming several of the proteins differentially regulated with functions in the postsynaptic membrane.
Calcium homeostasis and signaling
Calcium signaling has also been found to be differentially regulated in schizophrenia proteomic studies.54,76,113,116Calcium is a pivotal metabolite for the dopaminergic hypothesis in schizophrenia, mainly because it has a central role in the function of dopamine receptors D1 and D2.117 Proteins such as calmodulin (CALM1, CALM2), calcium/calmodulin-dependent protein kinase II (CAMK2B, CAMK2D, CAMK2G), voltage-dependent anion channels (VDAC1, VDAC2), and the plasma membrane calcium-transporting ATPase 4 (PMCA-4) are some of the calcium-related proteins found downregulated in the brains of schizophrenia patients.59,116 Some proteins were found differentially expressed in secretion fluids of schizophrenic patients—for example, calmodulin-like proteins and the S100 family of calcium-binding proteins (S100A6, S100A12)—such as in eccrine sweat118 and saliva.76 Complementing these findings, S100B was found downregulated in the nuclear proteome of schizophrenia corpus callosum.119 In addition, calcium activated differential expression of calmodulin-dependent protein kinase II (CAMK2), and calcineurin A in phencyclidine-treated rats.113
Energy metabolism
The brain has a high glucose uptake to supply its major metabolic activity rate. Thus, one of the most consistent dysfunctions underlying the pathophysiology of schizophrenia is in energy metabolism pathways, along with mitochondrial dysfunction and oxidative stress.120,121Glucose metabolism is confirmed by hyperglycemia, impaired glucose tolerance, and/or insulin resistance in first-onset, antipsychotic, naive schizophrenic patients.110,122 Numerous proteomic studies have identified the glycolysis–gluconeogenesis pathway as being consistently disrupted both in brain and CSF,41,57,58,60,123 and is followed by peripheral tissues.77,90,113,120 The expression of proteins associated with the energy metabolism pathway, such as aldolase C (ALDOC), enolase 1 (ENO1), neuronal enolase 2 (ENO2), lactate dehydrogenase B (LDHB), phosphoglycerate mutase 1 (brain) (PGAM1), phosphoglycerate kinase 1 (PGK1), pyruvate kinase isozyme R/L [PKLR], and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), are often significantly deregulated in schizophrenic patients compared with controls.55,77,120 The most consistent differentially expressed enzyme is aldolase C (ALDOC), which was found altered in several brain samples58,61,66 and also as a marker on blood serum samples.77 Likewise, pyruvate, the final product of glycolysis, and NADPH have been quantified in lower amounts in schizophrenic samples compared with controls, in the thalamus,57 and is replicated in phencyclidine-treated rats, a model of schizophrenia research.113 Whereas schizophrenia seems to be more associated with glycolysis, major depressive disorders are likely to be more associated with oxidative phosphorylation.124
DISC1, a major risk factor of the schizophrenia-susceptibility gene candidate,22 can affect mitochondrial morphology and axonal trafficking.125,126 Alterations of mitochondria morphology were reinforced by the imbalance of the oxidative phosphorilation system, including proteins such as NADH dehydrogenases (i.e., NDUFA1, NDUFV2, NDUFS3, NDUFB5), and ATPases (ATP5B, ATP6V1B2, ATP6V1A1), which have been previously shown altered in animal models of schizoprenia,113,127,128,129 but also had significant regulation in human brains.57,59 Other molecules such as dopamine have been shown to inhibit electron transport chain complex I (NADH dehydrogenase).130
Oxidative stress
This overall imbalance of mitochondrial energy metabolism, associated with elevated calcium, leads to hazardous ROS concentrations and oxidative stress events in brain cells.131 The resultant ROS may cause oxidative damage in cellular DNA, RNA, proteins, and membrane lipids. Proteomics of the brain have shown several enzymes involved in redox activities (responsible for removing ROS and protecting cells against oxidative injury) to be differentially expressed in schizophrenia brain tissues. Proteins such as superoxide dismutase (which catalyze the dismutation of superoxide (O2−) into oxygen and hydrogen peroxide), peroxiredoxins (PRDX1, PRDX2, and PRDX3) (which are responsible for reducing hydrogen peroxide), glutathione S-transferases (i.e., GSTM3, GSTTLp28, and GSTP1) (which are a family of multifunctional enzymes involved in cellular detoxification, glutathione reduction, and neutralization of ROS), and NADPH-dependent oxidoreductases such as carbonyl reductases (CBR1 and CBR2) and quinoid dihydropteridine reductase (QDPR) (which might be involved in the NADP/NADPH imbalance observed in the thalamus) were often found regulated in brain tissue,41,57,58,59,61,113 but could also be detected to be differentially regulated in peripheral tissues such as blood and fibroblast samples .70,73,75,132 Proteomics and combined metabolomics support evidence that slight imbalance in energy glucose metabolism, disrupting mitochondria and the oxidative phosphorylation system, results in compromised ATP production and oxidative stress, which is central in the pathophysiology of schizophrenia.41,113,133
Cytoskeleton
Cytoskeleton constituents are proteins that have shown broad differential expressions in schizophrenia—namely, microtubules such as tubulins (TBA1B, TUBB2A), microfilaments such as actins (ACTG1, ACTB) and actin-binding proteins such as tropomyosins (TPM1, TPM2, TPM3, TPM4), and intermediate filaments (i.e., GFAP, vimetin) and endocytosis proteins, such as dynamin (DNM1), a protein involved in clathrin-mediated endocytosis and other vesicular trafficking processes.55,58,59,113,134,135 Such modifications impact the cellular structure, axonal function, and neurite outgrowth, influencing synaptic plasticity and metabolism, all significantly influencing disturbed cytoskeleton arrangement in schizophrenia.44 Protein components of the cytoskeleton, such as the above-mentioned neurofilaments M and L (NEFL, NEFM) and DPYSL2, a regulator of cytoskeleton remodeling, have a role in axon guidance, neuronal growth, and cell migration. Glial fibrillary acidic protein (GFAP), the major intermediate filament of astrocytes, was found to be strongly regulated in brain tissues, both up- and downexpressed, indicating a precise protein expression across the brain.55,59,61,135,136 In addition, actin was often found downregulated in brain tissues,41,61,62,75 but was upregulated in fibroblasts75 or liver74 of schizophrenic patients.
Immune system and inflammation
Several abnormalities were found in schizophrenia proteomics, including changes in immune- and inflammation-related pathways in first-onset schizophrenic patients compared with controls. Molecules such as α-defensins (DEFA1, DEFA2, DEFA3, DEFA4), migration inhibitory factor, and several interleukins (IL-1ra, IL-8, IL-10, IL-15, IL-16, IL-17, and IL-18), including growth factors such as brain-derived neurotrophic factor, have been differentially regulated in blood samples from schizophrenic patients compared with controls.70,73,76,109,115,132 In addition, extracellular calcium-binding S100A12 exhibits cytokine-like characteristics, recruiting inflammatory cells to the sites of tissue damage. Indeed, anti-inflammatory treatment with cyclooxygenase-2 (COX-2) inhibitors has shown diminished schizophrenic behavior by blocking the synthesis of proinflammatory prostaglandins.137 Multiplex immunoassay profiling studies of blood serum have found numerous components of inflammation signaling pathways.109,111,115 Levels of anti-inflammatory cytokines IL-1ra and IL-10 were decreased after treatment with atypical antipsychotics, which correlated with symptom improvement.109 In addition, profiling studies using a subset of cytokines found increased levels of interleukins (i.e., IL-1β) in the cerebrospinal fluid of first-episode schizophrenic patients, indicative of immune system activation in the brain of some patients.138 Therefore, a proper subset of those altered molecular inflammatory molecules could be included in a sensitive and specific biomarker panel, both for diagnosis and treatment follow-up response.
An overview
Diverse proteomic techniques provided non-biased screening analyses of postmortem brain tissue from schizophrenic patients, and insights into pathways affected in the disease.10,57 In addition, more accessible tissues, such as cerebrospinal fluid, blood serum and plasma, and others such as fibroblasts, liver, and urine,57,66,70,74,75,139 have complemented those findings, suggesting several proteins that could be used as biomarkers to improve diagnosis. We have not gone through the details of the role of oligodendrocytes in schizophrenia, as these were recently tackled somewhere else,140,141,142 although these are as important as all that are listed here.
Proteomic insights from naive first-onset patients’ impaired protein pathways confirm patterns of disease onset, which along with genetic predisposition could be used as biomarkers for stratification of patients, improving the diagnosis and treatment classification. Also, this valuable information can lead to a more individualized medication, selected according to specific molecular dysfunctions and phenotype observed in schizophrenic individuals. Understanding the pathways affected by medication might also lead to reliable analytical platforms to evaluate individual response to treatment in a personalized-medicine mode. Moreover, the ability to monitor levels of molecules in noninvasive body fluids, such as saliva, urine, or blood serum or plasma, is a great advance. In addition, knowledge of gene–protein pathway networks affected and impaired by the disease can give clues for the development of new and more efficient targeted drugs to those relevant pathways.51,121
Perspectives
Psychiatric disorders are one of the biggest burdens to society,3 and consequently one of the most challenging fields of medical research, with complex and multifactorial characteristics, along with genetic, neurodevelopmental, environmental, and molecular components. Hence, proteomics can add valuable insights into revealing psychiatric disorder connections, as it is closely linked to phenotype, and, by definition, proteomics constitute one of the most suitable approaches for this purpose.143
In 2010, the Human Proteome Organization (HUPO) started a project aiming to map the entire human proteome, the Human Proteome Project (HPP) initiative, with joint initiatives such as the Chromosome-centric Human Proteome Project.144,145,146 Thus, at the beginning of 2014, two extensive drafts of this map were released,79,80 showing progress in the identification of proteins from high-quality proteomic data to complement genomic annotation. The Human Brain Proteome Project (HBPP) initiative, specifically addressing the proteomic landscape of the human brain, aims to study individuals affected by neurodegenerative diseases, understanding its many different cell types and their particular structure at the cellular and tissue level.147,148 Another main focus is to untangle the human plasma proteome149 on health and disease, to support biomarker validation and development of new tools for diagnosis, disease progression, and medication efficiency, considering the confounding factors present in those body fluids.
From the schizophrenia research point of view, this are exciting news, because of the potential of information that can be extracted, as, regardless of efforts in the search for biomarkers, by investigating the transcriptome and proteome in the post-genomic era, schizophrenia is one more psychiatric disorder without a reliable marker. Those recent advances in ‘omics’ technologies, such as genomics, transcriptomics, proteomics, and metabolomics, which are not only expanding coverage and resolution but also becoming cheaper and more accessible, present new prospects for a global comprehension of biological characteristics of disease mechanisms.150
While genomic and transcriptomic technologies have achieved single-nucleotide resolution, the protein coverage of the amino-acid sequence is still restricted. State-of-the-art shotgun mass spectrometry has improved immensely, such as targeted proteomic measurements, and is useful for biomarker identification. Although the detection of some protein variants, such as differential splice products and posttranslational modifications, remains a challenge for proteomics to get a more comprehensive picture of the whole proteome using a systematic approach. This high-throughput investigation of nucleic acids, proteins, and metabolites from particular tissues and cells provides essential data, which is basic to system biology studies, in order to create integral models of cellular processes.151 Therefore, integrating biological data from omics studies to the expertise of complementary disciplines such as mathematics, physics, and computational sciences, toward better conceptual analysis and predictive models, provides new tools for understanding biological systems at different levels. Hence, we can analyze the cellular space-time and hierarchical organization,152 aiming for complete understanding of psychiatric diseases and identifying candidate biomarkers, especially before and after the onset of clinical manifestations, as well as target metabolic pathways impaired and/or affected by antipsychotics, in order to distinguish subgroups of patients who respond to medication on the basis of their molecular profiles.51
….. more
Heterochronic microRNAs in temporal specification of neural stem cells: application toward rejuvenation
Plasticity is a critical factor enabling stem cells to contribute to the development and regeneration of tissues. In the mammalian central nervous system (CNS), neural stem cells (NSCs) that are defined by their capability for self-renewal and differentiation into neurons and glia, are present in the ventricular neuroaxis throughout life. However, the differentiation potential of NSCs changes in a spatiotemporally regulated manner and these cells progressively lose plasticity during development. One of the major alterations in this process is the switch from neurogenesis to gliogenesis. NSCs initiate neurogenesis immediately after neural tube closure and then turn to gliogenesis from midgestation, which requires an irreversible competence transition that enforces a progressive reduction of neuropotency. A growing body of evidence indicates that the neurogenesis-to-gliogenesis transition is governed by multiple layers of regulatory networks consisting of multiple factors, including epigenetic regulators, transcription factors, and non-coding RNA (ncRNA). In this review, we focus on critical roles of microRNAs (miRNAs), a class of small ncRNA that regulate gene expression at the post-transcriptional level, in the regulation of the switch from neurogenesis to gliogenesis in NSCs in the developing CNS. Unraveling the regulatory interactions of miRNAs and target genes will provide insights into the regulation of plasticity of NSCs, and the development of new strategies for the regeneration of damaged CNS.
Biological process utilize amino acids almost uniquely one chiral form or the other. In terrestrial biology, only the L-amino acids is common in biological processes. If signature of life existed elsewhere in the D form it then be concluded that life had evolutionary beginning on that body. Detection of an enantiomeric excess of L over D would also be a powerful sign that life had existed on that body at one time, see link in here .
As we know powder XRD pattern of both the L- or D-amino acids are basically identical. This is primarily due to the fact that the chirality of the amino acids does not favor a specific orientation on the sample holder in the powder form. However, when the amino acids are crystallized in moisture environment, water molecule enters in the unit cell of the amino acid by changing its structure. In order to determine structural changes using x-ray diffraction, two samples of both L- and D-phenylalanine (min 98% and >99% purity, respectively; purchased from Sigma Aldrich Co. St. Louis Mo) directly on the x-ray sample holder, alumina single crystal (0001) face, see link in here. The picture below shows an example of the diffraction pattern for two specimens of D- and L- phenylalanine, fresh crystallized in pure water under the same conditions.
As we see from the picture above, the two peaks L and D do not have same position. For raw phenylalanine D and L enantiomers the laboratory x-ray diffraction pattern matches, despite the fact that Powder Diffraction File (PDF) that correspond to PDF# 11-0827 for D form and PDF# 37-1771 indicates detectable differences. This is due to a structure factor, which is invariant for both D and L forms. For fresh crystallized specimens of D and L forms, a difference can be distinguished. The diffraction peak D of D-Phenylalanine has a less d-spacing suggesting a more compact structure than L-Phenylalanine.
Comparing diffraction pattern for the entire range produced by L and D crystallized Phenylalanine in pure water, shifts occur in peak position and intensity. These differences are possibly due to stereo-specific interactions of D- and L- phenylalanine molecules with water molecules. The x-ray diffraction pattern for D and L form, a small part shown in the picture above, reflects new structural configurations such as rotation, slight deformation and/or translation of the phenylalanine molecule inside the unit cell. It is possible that D form crystallizes with more molecules of water than L form. Structural solution for freshly crystallized phenylalanine can be found using Rietveld refinement technique.
The implications of water effect on the structure of the amino acids can lead to similar effects on DNA structure as well as proteins and cells in human body.
There has always been Personalized Medicine if you consider the time a physician spends with a patient, which has dwindled. But the current recognition of personalized medicine refers to breakthrough advances in technological innovation in diagnostics and treatment that differentiates subclasses within diagnoses that are amenable to relapse eluding therapies. There are just a few highlights to consider:
We live in a world with other living beings that are adapting to a changing environmental stresses.
Nutritional resources that have been available and made plentiful over generations are not abundant in some climates.
Despite the huge impact that genomics has had on biological progress over the last century, there is a huge contribution not to be overlooked in epigenetics, metabolomics, and pathways analysis.
A Reconstructed View of Personalized Medicine
There has been much interest in ‘junk DNA’, non-coding areas of our DNA are far from being without function. DNA has two basic categories of nitrogenous bases: the purines (adenine [A] and guanine [G]), and the pyrimidines (cytosine [C], thymine [T], and no uracil [U]), while RNA contains only A, G, C, and U (no T). The Watson-Crick proposal set the path of molecular biology for decades into the 21st century, culminating in the Human Genome Project.
There is no uncertainty about the importance of “Junk DNA”. It is both an evolutionary remnant, and it has a role in cell regulation. Further, the role of histones in their relationship the oligonucleotide sequences is not understood. We now have a large output of research on noncoding RNA, including siRNA, miRNA, and others with roles other than transcription. This requires major revision of our model of cell regulatory processes. The classic model is solely transcriptional.
DNA-> RNA-> Amino Acid in a protein.
Redrawn we have
DNA-> RNA-> DNA and
DNA->RNA-> protein-> DNA.
Neverthess, there were unrelated discoveries that took on huge importance. For example, since the 1920s, the work of Warburg and Meyerhoff, followed by that of Krebs, Kaplan, Chance, and others built a solid foundation in the knowledge of enzymes, coenzymes, adenine and pyridine nucleotides, and metabolic pathways, not to mention the importance of Fe3+, Cu2+, Zn2+, and other metal cofactors. Of huge importance was the work of Jacob, Monod and Changeux, and the effects of cooperativity in allosteric systems and of repulsion in tertiary structure of proteins related to hydrophobic and hydrophilic interactions, which involves the effect of one ligand on the binding or catalysis of another, demonstrated by the end-product inhibition of the enzyme, L-threonine deaminase (Changeux 1961), L-isoleucine, which differs sterically from the reactant, L-threonine whereby the former could inhibit the enzyme without competing with the latter. The current view based on a variety of measurements (e.g., NMR, FRET, and single molecule studies) is a ‘‘dynamic’’ proposal by Cooper and Dryden (1984) that the distribution around the average structure changes in allostery affects the subsequent (binding) affinity at a distant site.
What else do we have to consider? The measurement of free radicals has increased awareness of radical-induced impairment of the oxidative/antioxidative balance, essential for an understanding of disease progression. Metal-mediated formation of free radicals causes various modifications to DNA bases, enhanced lipid peroxidation, and altered calcium and sulfhydryl homeostasis. Lipid peroxides, formed by the attack of radicals on polyunsaturated fatty acid residues of phospholipids, can further react with redox metals finally producing mutagenic and carcinogenic malondialdehyde, 4-hydroxynonenal and other exocyclic DNA adducts (etheno and/or propano adducts). The unifying factor in determining toxicity and carcinogenicity for all these metals is the generation of reactive oxygen and nitrogen species. Various studies have confirmed that metals activate signaling pathways and the carcinogenic effect of metals has been related to activation of mainly redox sensitive transcription factors, involving NF-kappaB, AP-1 and p53.
I have provided mechanisms explanatory for regulation of the cell that go beyond the classic model of metabolic pathways associated with the cytoplasm, mitochondria, endoplasmic reticulum, and lysosome, such as, the cell death pathways, expressed in apoptosis and repair. Nevertheless, there is still a missing part of this discussion that considers the time and space interactions of the cell, cellular cytoskeleton and extracellular and intracellular substrate interactions in the immediate environment.
There is heterogeneity among cancer cells of expected identical type, which would be consistent with differences in phenotypic expression, aligned with epigenetics. There is also heterogeneity in the immediate interstices between cancer cells. Integration with genome-wide profiling data identified losses of specific genes on 4p14 and 5q13 that were enriched in grade 3 tumors with high microenvironmental diversity that also substratified patients into poor prognostic groups. In the case of breast cancer, there is interaction with estrogen , and we refer to an androgen-unresponsive prostate cancer.
Finally, the interaction between enzyme and substrates may be conditionally unidirectional in defining the activity within the cell. The activity of the cell is dynamically interacting and at high rates of activity. In a study of the pyruvate kinase (PK) reaction the catalytic activity of the PK reaction was reversed to the thermodynamically unfavorable direction in a muscle preparation by a specific inhibitor. Experiments found that in there were differences in the active form of pyruvate kinase that were clearly related to the environmental condition of the assay – glycolitic or glyconeogenic. The conformational changes indicated by differential regulatory response were used to present a dynamic conformational model functioning at the active site of the enzyme. In the model, the interaction of the enzyme active site with its substrates is described concluding that induced increase in the vibrational energy levels of the active site decreases the energetic barrier for substrate induced changes at the site. Another example is the inhibition of H4 lactate dehydrogenase, but not the M4, by high concentrations of pyruvate. An investigation of the inhibition revealed that a covalent bond was formed between the nicotinamide ring of the NAD+ and the enol form of pyruvate. The isoenzymes of isocitrate dehydrogenase, IDH1 and IDH2 mutations occur in gliomas and in acute myeloid leukemias with normal karyotype. IDH1 and IDH2 mutations are remarkably specific to codons that encode conserved functionally important arginines in the active site of each enzyme. In this case, there is steric hindrance by Asp279 where the isocitrate substrate normally forms hydrogen bonds with Ser94.
Personalized medicine has been largely viewed from a lens of genomics. But genomics is only the reading frame. The living activities of cell processes are dynamic and occur at rapid rates. We have to keep in mind that personalized in reference to genotype is not complete without reconciliation of phenotype, which is the reference to expressed differences in outcomes.
Biochemistry and Dysmetabolism of Aging and Serious Illness, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 1: Next Generation Sequencing (NGS)
Biochemistry and Dysmetabolism of Aging and Serious Illness
Curator: Larry H. Bernstein, MD, FCAP
White Matter Lipids as a Ketogenic Fuel Supply in Aging Female Brain: Implications for Alzheimer’s Disease
•Mitochondrial dysfunction activates mechanisms for catabolism of myelin lipids to generate ketone bodies for ATP production.
•Mechanisms leading to ketone body driven energy production in brain coincide with stages of reproductive aging in females.
•Sequential activation of myelin catabolism pathway during aging provides multiple therapeutic targets and windows of efficacy.
The mechanisms underlying white matter degeneration, a hallmark of multiple neurodegenerative diseases including Alzheimer’s, remain unclear. Herein we provide a mechanistic pathway, spanning multiple transitions of aging, that links mitochondrial dysfunction early in aging with later age white matter degeneration. Catabolism of myelin lipids to generate ketone bodies can be viewed as an adaptive survival response to address brain fuel and energy demand. Women are at greatest risk of late-onset-AD, thus, our analyses in female brain address mechanisms of AD pathology and therapeutic targets to prevent, delay and treat AD in the sex most affected with potential relevance to men.
White matter degeneration is a pathological hallmark of neurodegenerative diseases including Alzheimer’s. Age remains the greatest risk factor for Alzheimer’s and the prevalence of age-related late onset Alzheimer’s is greatest in females. We investigated mechanisms underlying white matter degeneration in an animal model consistent with the sex at greatest Alzheimer’s risk. Results of these analyses demonstrated decline in mitochondrial respiration, increased mitochondrial hydrogen peroxide production and cytosolic-phospholipase-A2 sphingomyelinase pathway activation during female brain aging. Electron microscopic and lipidomic analyses confirmed myelin degeneration. An increase in fatty acids and mitochondrial fatty acid metabolism machinery was coincident with a rise in brain ketone bodies and decline in plasma ketone bodies. This mechanistic pathway and its chronologically phased activation, links mitochondrial dysfunction early in aging with later age development of white matter degeneration. The catabolism of myelin lipids to generate ketone bodies can be viewed as a systems level adaptive response to address brain fuel and energy demand. Elucidation of the initiating factors and the mechanistic pathway leading to white matter catabolism in the aging female brain provides potential therapeutic targets to prevent and treat demyelinating diseases such as Alzheimer’s and multiple sclerosis. Targeting stages of disease and associated mechanisms will be critical.
Age remains the greatest risk factor for developing AD (Hansson et al., 2006, Alzheimer’s, 2015). Thus, investigation of transitions in the aging brain is a reasoned strategy for elucidating mechanisms and pathways of vulnerability for developing AD. Aging, while typically perceived as a linear process, is likely composed of dynamic transition states, which can protect against or exacerbate vulnerability to AD (Brinton et al., 2015). An aging transition unique to the female is the perimenopausal to menopausal conversion (Brinton et al., 2015). The bioenergetic similarities between the menopausal transition in women and the early appearance of hypometabolism in persons at risk for AD make the aging female a rational model to investigate mechanisms underlying risk of late onset AD.
Findings from this study replicate our earlier findings that age of reproductive senescence is associated with decline in mitochondrial respiration, increased H2O2 production and shift to ketogenic metabolism in brain (Yao et al., 2010, Ding et al., 2013, Yin et al., 2015). These well established early age-related changes in mitochondrial function and shift to ketone body utilization in brain, are now linked to a mechanistic pathway that connects early decline in mitochondrial respiration and H2O2 production to activation of the cPLA2-sphingomyelinase pathway to catabolize myelin lipids resulting in WM degeneration (Fig. 12). These lipids are sequestered in lipid droplets for subsequent use as a local source of ketone body generation via astrocyte mediated beta-oxidation of fatty acids. Astrocyte derived ketone bodies can then be transported to neurons where they undergo ketolysis to generate acetyl-CoA for TCA derived ATP generation required for synaptic and cell function (Fig. 12).
Schematic model of mitochondrial H2O2 activation of cPLA2-sphingomyelinase pathway as an adaptive response to provide myelin derived fatty acids as a substrate for ketone body generation: The cPLA2-sphingomyelinase pathway is proposed as a mechanistic pathway that links an early event, mitochondrial dysfunction and H2O2, in the prodromal/preclinical phase of Alzheimer’s with later stage development of pathology, white matter degeneration. Our findings demonstrate that an age dependent deficit in mitochondrial respiration and a concomitant rise in oxidative stress activate an adaptive cPLA2-sphingomyelinase pathway to provide myelin derived fatty acids as a substrate for ketone body generation to fuel an energetically compromised brain.
Biochemical evidence obtained from isolated whole brain mitochondria confirms that during reproductive senescence and in response to estrogen deprivation brain mitochondria decline in respiratory capacity (Yao et al., 2009, Yao et al., 2010, Brinton, 2008a, Brinton, 2008b, Swerdlow and Khan, 2009). A well-documented consequence of mitochondrial dysfunction is increased production of reactive oxygen species (ROS), specifically H2O2 (Boveris and Chance, 1973, Beal, 2005, Yin et al., 2014, Yap et al., 2009). While most research focuses on the damage generated by free radicals, in this case H2O2 functions as a signaling molecule to activate cPLA2, the initiating enzyme in the cPLA2-sphingomyelinase pathway (Farooqui and Horrocks, 2006, Han et al., 2003, Sun et al., 2004). In AD brain, increased cPLA2 immunoreactivity is detected almost exclusively in astrocytes suggesting that activation of the cPLA2-sphingomyelinase pathway is localized to astrocytes in AD, as opposed to the neuronal or oligodendroglial localization that is observed during apoptosis (Sun et al., 2004, Malaplate-Armand et al., 2006, Di Paolo and Kim, 2011, Stephenson et al., 1996,Stephenson et al., 1999). In our analysis, cPLA2 (Sanchez-Mejia and Mucke, 2010) activation followed the age-dependent rise in H2O2 production and was sustained at an elevated level.
Direct and robust activation of astrocytic cPLA2 by physiologically relevant concentrations of H2O2 was confirmed in vitro. Astrocytic involvement in the cPLA2-sphingomyelinase pathway was also indicated by an increase in cPLA2 positive astrocyte reactivity in WM tracts of reproductively incompetent mice. These data are consistent with findings from brains of persons with AD that demonstrate the same striking localization of cPLA2immunoreactivity within astrocytes, specifically in the hippocampal formation (Farooqui and Horrocks, 2004). While neurons and astrocytes contain endogenous levels of cPLA2, neuronal cPLA2 is activated by an influx of intracellular calcium, whereas astrocytic cPLA2 is directly activated by excessive generation of H2O2 (Sun et al., 2004, Xu et al., 2003, Tournier et al., 1997). Evidence of this cell type specific activation was confirmed by the activation of cPLA2 in astrocytes by H2O2 and the lack of activation in neurons. These data support that astrocytic, not neuronal, cPLA2 is the cellular mediator of the H2O2 dependent cPLA2-sphingomyelinase pathway activation and provide associative evidence supporting a role of astrocytic mitochondrial H2O2 in age-related WM catabolism.
The pattern of gene expression during the shift to reproductive senescence in the female mouse hippocampus recapitulates key observations in human AD brain tissue, specifically elevation in cPLA2, sphingomyelinase and ceramidase (Schaeffer et al., 2010, He et al., 2010, Li et al., 2014). Further, up-regulation of myelin synthesis, lipid metabolism and inflammatory genes in reproductively incompetent female mice is consistent with the gene expression pattern previously reported from aged male rodent hippocampus, aged female non-human primate hippocampus and human AD hippocampus (Blalock et al., 2003, Blalock et al., 2004, Blalock et al., 2010, Blalock et al., 2011, Kadish et al., 2009, Rowe et al., 2007). In these analyses of gene expression in aged male rodent hippocampus, aged female non-human primate hippocampus and human AD hippocampus down regulation of genes related to mitochondrial function, and up-regulation in multiple genes encoding for enzymes involved in ketone body metabolism occurred (Blalock et al., 2003, Blalock et al., 2004, Blalock et al., 2010, Blalock et al., 2011, Kadish et al., 2009, Rowe et al., 2007). The comparability across data derived from aging female mouse hippocampus reported herein and those derived from male rodent brain, female nonhuman brain and human AD brain strongly suggest that cPLA2-sphingomyelinase pathway activation, myelin sheath degeneration and fatty acid metabolism leading to ketone body generation is a metabolic adaptation that is generalizable across these naturally aging models and are evident in aged human AD brain. Collectively, these data support the translational relevance of findings reported herein.
Data obtained via immunohistochemistry, electron microscopy and MBP protein analyses demonstrated an age-related loss in myelin sheath integrity. Evidence for a loss of myelin structural integrity emerged in reproductively incompetent mice following activation of the cPLA2-sphingomyelinase pathway. The unraveling myelin phenotype observed following reproductive senescence and aging reported herein is consistent with the degenerative phenotype that emerges following exposure to the chemotherapy drug bortezomib which induces mitochondrial dysfunction and increased ROS generation (Carozzi et al., 2010, Cavaletti et al., 2007,Ling et al., 2003). In parallel to the decline in myelin integrity, lipid droplet density increased. In aged mice, accumulation of lipid droplets declined in parallel to the rise in ketone bodies consistent with the utilization of myelin-derived fatty acids to generate ketone bodies. Due to the sequential relationship between WM degeneration and lipid droplet formation, we posit that lipid droplets serve as a temporary storage site for myelin-derived fatty acids prior to undergoing β-oxidation in astrocytes to generate ketone bodies.
Microstructural alterations in myelin integrity were associated with alterations in the lipid profile of brain, indicative of WM degeneration resulting in release of myelin lipids. Sphingomyelin and galactocerebroside are two main lipids that compose the myelin sheath (Baumann and Pham-Dinh, 2001). Ceramide is common to both galactocerebroside and sphingomyelin and is composed of sphingosine coupled to a fatty acid. Ceramide levels increase in aging, in states of ketosis and in neurodegeneration (Filippov et al., 2012, Blazquez et al., 1999, Costantini et al., 2005). Specifically, ceramide levels are elevated at the earliest clinically recognizable stage of AD, indicating a degree of WM degeneration early in disease progression (Di Paolo and Kim, 2011,Han et al., 2002, Costantini et al., 2005). Sphingosine is statistically significantly elevated in the brains of AD patients compared to healthy controls; a rise that was significantly correlated with acid sphingomyelinase activity, Aβ levels and tau hyperphosphorylation (He et al., 2010). In our analyses, a rise in ceramides was first observed early in the aging process in reproductively incompetent mice. The rise in ceramides was coincident with the emergence of loss of myelin integrity consistent with the release of myelin ceramides from sphingomyelin via sphingomyelinase activation. Following the rise in ceramides, sphingosine and fatty acid levels increased. The temporal sequence of the lipid profile was consistent with gene expression indicating activation of ceramidase for catabolism of ceramide into sphingosine and fatty acid during reproductive senescence. Once released from ceramide, fatty acids can be transported into the mitochondrial matrix of astrocytes via CPT-1, where β-oxidation of fatty acids leads to the generation of acetyl-CoA (Glatz et al., 2010). It is well documented that acetyl-CoA cannot cross the inner mitochondrial membrane, thus posing a barrier to direct transport of acetyl-CoA generated by β-oxidation into neurons. In response, the newly generated acetyl-CoA undergoes ketogenesis to generate ketone bodies to fuel energy demands of neurons (Morris, 2005,Guzman and Blazquez, 2004, Stacpoole, 2012). Because astrocytes serve as the primary location of β-oxidation in brain they are critical to maintaining neuronal metabolic viability during periods of reduced glucose utilization (Panov et al., 2014, Ebert et al., 2003, Guzman and Blazquez, 2004).
Once fatty acids are released from myelin ceramides, they are transported into astrocytic mitochondria by CPT1 to undergo β-oxidation. The mitochondrial trifunctional protein HADHA catalyzes the last three steps of mitochondrial β-oxidation of long chain fatty acids, while mitochondrial ABAD (aka SCHAD—short chain fatty acid dehydrogenase) metabolizes short chain fatty acids. Concurrent with the release of myelin fatty acids in aged female mice, CPT1, HADHA and ABAD protein expression as well as ketone body generation increased significantly. These findings indicate that astrocytes play a pivotal role in the response to bioenergetic crisis in brain to activate an adaptive compensatory system that activates catabolism of myelin lipids and the metabolism of those lipids into fatty acids to generate ketone bodies necessary to fuel neuronal demand for acetyl-CoA and ATP.
Collectively, these findings provide a mechanistic pathway that links mitochondrial dysfunction and H2O2generation in brain early in the aging process to later stage white matter degeneration. Astrocytes play a pivotal role in providing a mechanistic strategy to address the bioenergetic demand of neurons in the aging female brain. While this pathway is coincident with reproductive aging in the female brain, it is likely to have mechanistic translatability to the aging male brain. Further, the mechanistic link between bioenergetic decline and WM degeneration has potential relevance to other neurological diseases involving white matter in which postmenopausal women are at greater risk, such as multiple sclerosis. The mechanistic pathway reported herein spans time and is characterized by a progression of early adaptive changes in the bioenergetic system of the brain leading to WM degeneration and ketone body production. Translationally, effective therapeutics to prevent, delay and treat WM degeneration during aging and Alzheimer’s disease will need to specifically target stages within the mechanistic pathway described herein. The fundamental initiating event is a bioenergetic switch from being a glucose dependent brain to a glucose and ketone body dependent brain. It remains to be determined whether it is possible to prevent conversion to or reversal of a ketone dependent brain. Effective therapeutic strategies to intervene in this process require biomarkers of bioenergetic phenotype of the brain and stage of mechanistic progression. The mechanistic pathway reported herein may have relevance to other age-related neurodegenerative diseases characterized by white matter degeneration such as multiple sclerosis.
Cancer cells are characterized by perturbations of their metabolic processes. Recent observations demonstrated that the fatty acid oxidation (FAO) pathway may represent an alternative carbon source for anabolic processes in different tumors, therefore appearing particularly promising for therapeutic purposes. Because the carnitine palmitoyl transferase 1a (CPT1a) is a protein that catalyzes the rate-limiting step of FAO, here we investigated the in vitro antileukemic activity of the novel CPT1a inhibitor ST1326 on leukemia cell lines and primary cells obtained from patients with hematologic malignancies. By real-time metabolic analysis, we documented that ST1326 inhibited FAO in leukemia cell lines associated with a dose- and time-dependent cell growth arrest, mitochondrial damage, and apoptosis induction. Data obtained on primary hematopoietic malignant cells confirmed the FAO inhibition and cytotoxic activity of ST1326, particularly on acute myeloid leukemia cells. These data suggest that leukemia treatment may be carried out by targeting metabolic processes.
Suppression of detachment-induced cell death, known as anoikis, is an essential step for cancer metastasis to occur. We report here that expression of KLF12, a member of the Kruppel-like family of transcription factors, is downregulated in lung cancer cell lines that have been selected to grow in the absence of cell adhesion. Knockdown of KLF12 in parental cells results in decreased apoptosis following cell detachment from matrix. KLF12 regulates anoikis by promoting the cell cycle transition through S phase and therefore cell proliferation. Reduced expression levels of KLF12 results in increased ability of lung cancer cells to form tumours in vivo and is associated with poorer survival in lung cancer patients. We therefore identify KLF12 as a novel metastasis-suppressor gene whose loss of function is associated with anoikis resistance through control of the cell cycle.
Mol Cell. 2015 Oct 14. pii: S1097-2765(15)00764-9. doi: 10.1016/j.molcel.2015.09.025. [Epub ahead of print] PEPCK Coordinates the Regulation of Central Carbon Metabolism to Promote Cancer Cell Growth.
Phosphoenolpyruvate carboxykinase (PEPCK) is well known for its role in gluconeogenesis. However, PEPCK is also a key regulator of TCA cycle flux. The TCA cycle integrates glucose, amino acid, and lipid metabolism depending on cellular needs. In addition, biosynthetic pathways crucial to tumor growth require the TCA cycle for the processing of glucose and glutamine derived carbons. We show here an unexpected role for PEPCK in promoting cancer cell proliferation in vitro and in vivo by increasing glucose and glutamine utilization toward anabolic metabolism. Unexpectedly, PEPCK also increased the synthesis of ribose from non-carbohydrate sources, such as glutamine, a phenomenon not previously described. Finally, we show that the effects of PEPCK on glucose metabolism and cell proliferation are in part mediated via activation of mTORC1. Taken together, these data demonstrate a role for PEPCK that links metabolic flux and anabolic pathways to cancer cell proliferation.
BACKGROUND: Mutations in the Fe-S cluster-containing SDHB subunit of succinate dehydrogenase cause familial cancer syndromes. Recently the tripeptide motif L(I)YR was identified in the Fe-S recipient protein SDHB, to which the cochaperone HSC20 binds.
METHODS: In order to characterize the metabolic basis of SDH-deficient cancers we performed stable isotope-resolved metabolomics in a novel SDHB-deficient renal cell carcinoma cell line and conducted bioinformatics and biochemical screening to analyze Fe-S cluster acquisition and assembly of SDH in the presence of other cancer-causing SDHB mutations.
RESULTS:
We found that the SDHB(R46Q) mutation in UOK269 cells disrupted binding of HSC20, causing rapid degradation of SDHB. In the absence of SDHB, respiration was undetectable in UOK269 cells, succinate was elevated to 351.4±63.2 nmol/mg cellular protein, and glutamine became the main source of TCA cycle metabolites through reductive carboxylation. Furthermore, HIF1α, but not HIF2α, increased markedly and the cells showed a strong DNA CpG island methylator phenotype (CIMP). Biochemical and bioinformatic screening revealed that 37% of disease-causing missense mutations in SDHB were located in either the L(I)YR Fe-S transfer motifs or in the 11 Fe-S cluster-ligating cysteines.
CONCLUSIONS:
These findings provide a conceptual framework for understanding how particular mutations disproportionately cause the loss of SDH activity, resulting in accumulation of succinate and metabolic remodeling in SDHB cancer syndromes.
Targeting cancer cell metabolism is a promising strategy against cancer. Here, we confirmed that the anti-cancer drug carboxyamidotriazole (CAI) inhibited mitochondrial respiration in cancer cells for the first time and found a way to enhance its anti-cancer activity by further disturbing the energy metabolism. CAI promoted glucose uptake and lactate production when incubated with cancer cells. The oxidative phosphorylation (OXPHOS) in cancer cells was inhibited by CAI, and the decrease in the activity of the respiratory chain complex I could be one explanation. The anti-cancer effect of CAI was greatly potentiated when being combined with 2-deoxyglucose (2-DG). The cancer cells treated with the combination of CAI and 2-DG were arrested in G2/M phase. The apoptosis and necrosis rates were also increased. In a mouse xenograft model, this combination was well tolerated and retarded the tumor growth. The impairment of cancer cell survival was associated with significant cellular ATP decrease, suggesting that the combination of CAI and 2-DG could be one of the strategies to cause dual inhibition of energy pathways, which might be an effective therapeutic approach for a broad spectrum of tumors.
Yun, E. Mullarky, C. Lu, K. N. Bosch, A. Kavalier, K. Rivera, J. Roper, Chio, II, E. G. Giannopoulou, C. Rago, A. Muley, J. M. Asara, J. Paik, O. Elemento, Z. Chen, D. J. Pappin, L. E. Dow, N. Papadopoulos, S. S. Gross and L. C. Cantley.
The G protein-coupled receptor kinase-2 (GRK2) is upregulated in the injured heart and contributes to heart failure pathogenesis. GRK2 was recently shown to associate with mitochondria but its functional impact in myocytes due to this localization is unclear. This study was undertaken to determine the effect of elevated GRK2 on mitochondrial respiration in cardiomyocytes. Sub-fractionation of purified cardiac mitochondria revealed that basally GRK2 is found in multiple compartments. Overexpression of GRK2 in mouse cardiomyocytes resulted in an increased amount of mitochondrial-based superoxide. Inhibition of GRK2 increased oxygen consumption rates and ATP production. Moreover, fatty acid oxidation was found to be significantly impaired when GRK2 was elevated and was dependent on the catalytic activity and mitochondrial localization of this kinase. Our study shows that independent of cardiac injury, GRK2 is localized in the mitochondria and its kinase activity negatively impacts the function of this organelle by increasing superoxide levels and altering substrate utilization for energy production.
Br J Pharmacol. 2015 Oct 27. doi: 10.1111/bph.13377. [Epub ahead of print]
All-trans retinoic acid protects against doxorubicin-induced cardiotoxicity by activating the Erk2 signalling pathway.
Doxorubicin (Dox) is a powerful antineoplastic agent for treating a wide range of cancers. However, doxorubicin cardiotoxicity of the heart has largely limited its clinical use. It is known that all-trans retinoic acid (ATRA) plays important roles in many cardiac biological processes, however, the protective effects of ATRA on doxorubicin cardiotoxicity remain unknown. Here, we studied the effect of ATRA on doxorubicin cardiotoxicity and underlying mechanisms.
EXPERIMENTAL APPROACHES:
Cellular viability assays, western blotting and mitochondrial respiration analyses were employed to evaluate the cellular response to ATRA in H9c2 cells and primary cardiomyocytes. Quantitative PCR (Polymerase Chain Reaction) and gene knockdown were performed to investigate the underlying molecular mechanisms of ATRA’s effects on doxorubicin cardiotoxicity.
KEY RESULTS:
ATRA significantly inhibited doxorubicin-induced apoptosis in H9c2 cells and primary cardiomyocytes. ATRA was more effective against doxorubicin cardiotoxicity than resveratrol and dexrazoxane. ATRA also suppressed reactive oxygen species (ROS) generation, and restored the expression level of mRNA and proteins in phase II detoxifying enzyme system: Nrf2 (nuclear factor-E2-related factor 2), MnSOD (manganese superoxide dismutase), HO-1 (heme oxygenase1) as well as mitochondrial function (mitochondrial membrane integrity, mitochondrial DNA copy numbers, mitochondrial respiration capacity, biogenesis and dynamics). Both Erk1/2 (extracellular signal-regulated kinase1/2) inhibitor (U0126) and Erk2 siRNA, but not Erk1 siRNA, abolished the protective effect of ATRA against doxorubicin-induced toxicity in H9c2 cells. Remarkably, ATRA did not compromise the anticancer efficacy of doxorubicin in gastric carcinoma cells.
CONCLUSION AND IMPLICATION:
ATRA protected cardiomyocytes against doxorubicin-induced toxicity by activating the Erk2 pathway without compromising the anticancer efficacy of doxorubicin. Therefore, ATRA may be a promising candidate as a cardioprotective agent against doxorubicin cardiotoxicity.
Colak, O. Pougovkina, L. Dai, M. Tan, H. Te Brinke, H. Huang, Z. Cheng, J. Park, X. Wan, X. Liu, W. W. Yue, R. J. Wanders, J. W. Locasale, D. B. Lombard, V. C. de Boer and Y. Zhao.
Mol Cell Proteomics. 2015 Nov 1, 14 (11): 3056-71.
The insulin/insulin-like growth factor (IGF)-1 signaling pathway (ISP) plays a fundamental role in long term health in a range of organisms. Protein kinases including Akt and ERK are intimately involved in the ISP. To identify other kinases that may participate in this pathway or intersect with it in a regulatory manner, we performed a whole kinome (779 kinases) siRNA screen for positive or negative regulators of the ISP, using GLUT4 translocation to the cell surface as an output for pathway activity. We identified PFKFB3, a positive regulator of glycolysis that is highly expressed in cancer cells and adipocytes, as a positive ISP regulator. Pharmacological inhibition of PFKFB3 suppressed insulin-stimulated glucose uptake, GLUT4 translocation, and Akt signaling in 3T3-L1 adipocytes. In contrast, overexpression of PFKFB3 in HEK293 cells potentiated insulin-dependent phosphorylation of Akt and Akt substrates. Furthermore, pharmacological modulation of glycolysis in 3T3-L1 adipocytes affected Akt phosphorylation. These data add to an emerging body of evidence that metabolism plays a central role in regulating numerous biological processes including the ISP. Our findings have important implications for diseases such as type 2 diabetes and cancer that are characterized by marked disruption of both metabolism and growth factor signaling.
Skeletal muscle mitochondrial content and oxidative capacity are important determinants of muscle function and whole-body health. Mitochondrial content and function are enhanced by endurance exercise and impaired in states or diseases where muscle function is compromised, such as myopathies, muscular dystrophies, neuromuscular diseases, and age-related muscle atrophy. Hence, elucidating the mechanisms that control muscle mitochondrial content and oxidative function can provide new insights into states and diseases that affect muscle health. In past studies, we identified Perm1 (PPARGC1- and ESRR-induced regulator, muscle 1) as a gene induced by endurance exercise in skeletal muscle, and regulating mitochondrial oxidative function in cultured myotubes. The capacity of Perm1 to regulate muscle mitochondrial content and function in vivo is not yet known. In this study, we use adeno-associated viral (AAV) vectors to increase Perm1 expression in skeletal muscles of 4-wk-old mice. Compared to control vector, AAV1-Perm1 leads to significant increases in mitochondrial content and oxidative capacity (by 40-80%). Moreover, AAV1-Perm1-transduced muscles show increased capillary density and resistance to fatigue (by 33 and 31%, respectively), without prominent changes in fiber-type composition. These findings suggest that Perm1 selectively regulates mitochondrial biogenesis and oxidative function, and implicate Perm1 in muscle adaptations that also occur in response to endurance exercise.-Cho, Y., Hazen, B. C., Gandra, P. G., Ward, S. R., Schenk, S., Russell, A. P., Kralli, A. Perm1 enhances mitochondrial biogenesis, oxidative capacity, and fatigue resistance in adult skeletal muscle.
Exposure to metabolic disease during fetal development alters cellular differentiation and perturbs metabolic homeostasis, but the underlying molecular regulators of this phenomenon in muscle cells are not completely understood. To address this, we undertook a computational approach to identify cooperating partners of the myocyte enhancer factor-2 (MEF2) family of transcription factors, known regulators of muscle differentiation and metabolic function. We demonstrate that MEF2 and the serum response factor (SRF) collaboratively regulate the expression of numerous muscle-specific genes, including microRNA-133a (miR-133a). Using tandem mass spectrometry techniques, we identify a conserved phosphorylation motif within the MEF2 and SRF Mcm1 Agamous Deficiens SRF (MADS)-box that regulates miR-133a expression and mitochondrial function in response to a lipotoxic signal. Furthermore, reconstitution of MEF2 function by expression of a neutralizing mutation in this identified phosphorylation motif restores miR-133a expression and mitochondrial membrane potential during lipotoxicity. Mechanistically, we demonstrate that miR-133a regulates mitochondrial function through translational inhibition of a mitophagy and cell death modulating protein, called Nix. Finally, we show that rodents exposed to gestational diabetes during fetal development display muscle diacylglycerol accumulation, concurrent with insulin resistance, reduced miR-133a, and elevated Nix expression, as young adult rats. Given the diverse roles of miR-133a and Nix in regulating mitochondrial function, and proliferation in certain cancers, dysregulation of this genetic pathway may have broad implications involving insulin resistance, cardiovascular disease, and cancer biology.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
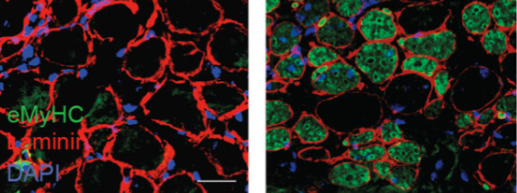{kind=link}
{kind=link}
{kind=link}
This year’s Nobel Prize laureates in chemistry Demis Hassabis and John Jumper have developed an AI model to solve a 50-year-old problem: predicting proteins’ complex structures.
In 2020, Hassabis and Jumper presented an AI model called AlphaFold2. With its help, they have been able to predict the structure of virtually all the 200 million proteins that researchers have identified. Since their breakthrough, AlphaFold2 has been used by more than two million people from 190 countries. Among a myriad of scientific applications, researchers can now better understand antibiotic resistance and create images of enzymes that can decompose plastic.
Read more about their story: https://bit.ly/4diKiJ2