Translational Research on the Mechanism of Water and Electrolyte Movements into the Cell
Reviewer and Curator: Larry H. Bernstein, MD, FACP
Introduction
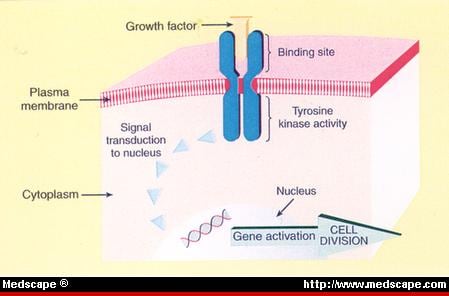
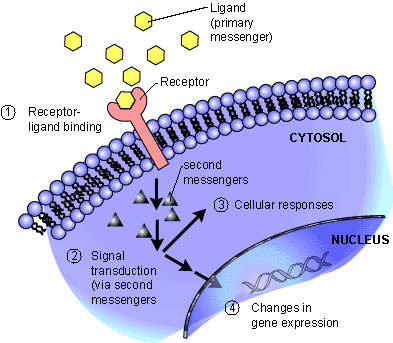
This article is the first in a three part curation covering work that has great importance to our understanding of hydration and possibly the effects of dehydration in cell physiology, and studied effects on renal function and brain, with possible implications for heart failure, myocardial contraction, heart rate, and arrhythmiagenesis. The discovery of aquaporins and the elucidation of potassium channels and selective ion conduction was jointly awarded the Nobel Prize in Chemistry in 2003 to Peter Agre, at the Johns Hopkins School of Medicine, Baltimore, and Roderick Mac Kinnon, at the Howard Hughes Medical Institute, Laboratory of Molecular Neurobiology and Biophysics, Rockefeller University, New York, NY. The transport of water, it was assumed, is associated with the movements of Na(+), K(+), Ca(2+), Mg(2+). The calmodulin kinase, rhyanodine, and calcium sparks in the Ca(2+) release from sarcolemma is covered elsewhere in cardiac contraction, skeletal muscle, smooth muscle, and neural stimulation of muscle and adrenergic release. The sodium/potassium exchange is depicted in diagrams, but not discussed. In traditional chemistry we would think in terms of a cationic and anionic balance that has to be maintained in charge equivalents on both sides of a membrane. However, the intricacies of membrane structure as well as active transporters has been delineated and has been a transformative factor in our understanding of organ function in health and disease.
Aquaporin Water Channels
We have studied the aquaporin water channels for several years, and we now understand that they explain how water crosses biological membranes. Our bodies are 70% water, and all other vertebrates, invertebrates, microbes, and plants are also primarily water. The organization of water within biological compartments is fundamental to life, and the aquaporins serve as the plumbing systems for cells. Aquaporins explain how our
brains secrete and absorb spinal fluid, how we can generate aqueous humor within our eyes, how we can secrete tears, saliva, sweat, and bile, and how our kidneys can concentrate urine so effectively. These proteins are fundamental to mammalian physiology, but they are also very important in the lives of microorganisms and plants.
It was correctly proposed in the 1920’s that water could move through the cell membrane simply by diffusing through the lipid bilayer. The current view is that the lipid bilayer has a finite permeability for water, but, in addition, a set of proteins exists that we now refer to as “aquaporins.” Their existence was suggested by a group of pioneers in the water transport field who preceded us by decades – people including Arthur K. Solomon in Boston, Alan Finkelstein in New York, Robert Macey in Berkeley, Gheorghe Benga in Romania, Guillermo Whittembury in Venezuela, Mario Parisi in Argentina – who by biophysical methods predicted that water channels must exist in certain cell types with high water permeability such as renal tubules, salivary glands, and red cells (reviewed by Finkelstein, 1987).
The difference between diffusional and channel-mediated water perme-ability is fairly distinct. Diffusion is a low capacity, bidirectional movement of water that occurs in all cell membranes, whereas the membranes of a subset of cells with aquaporin proteins have very high capacity for permeation by water.
This permeability is selective, since water (H O) crosses through the membranes with almost no resistance, while acid, the hydronium ion (H O ) does not permeate the proteins. This distinction is essential to life. The movement of water is directed by osmotic gradients, so aquaporins are not pumps or exchangers. They form a simple pore that allows water to rapidly pass through membranes by osmosis. There are also other differences between diffusion and channel-mediated water transport. No inhibitors are known for simple diffusion. In contrast, mercurials were discovered by Robert Macey to inhibit water transport in red cells but water permeability was restored by treatment with reducing agents (Macey and Farmer, 1970). These observations predicted that water channels must be proteins with sulfhydryls and characteristically low Arrhenius activation energy.
A number of investigators using ver y logical approaches attempted to identify the water channel molecule; identification proved a very difficult prolem. Isotopic mercurials labeled several membrane proteins – the anion exchanger (band 3). Solomon and a group of several proteins (band 4.5) by Benga. None of the proteins were isolated, reconstituted, and shown to transport watter (reviewed by Agre et al., 1993a).
DISCOVERY OF AQP1
The field was essentially stuck, but following the well known scientific approach known as “sheer blind luck,” we stumbled upon the protein. Looking through our notebooks for the earliest studies that showed there was such a protein water channel. We were at that time attempting to raise antibodies in rabbits to the denatured partially purified Rh polypeptide. The rabbits vigorously produced antibodies, but we failed to recognize initially that our antibody did not react with the core Rh polypeptide that migrated at 32 kDa, seen clearly by silver staining of sodium dodecyl sulfate polyacryamide electrophoresis gels (SDS-PAGE). Instead, our antibodies reacted only with a 28 kDa polypeptide. The 28 kDa was an unrelated protein. Silver staining of SDS-PAGE migration of the isolated protein revealed a discrete band of 28 kDa in detergent insoluble extracts (it failed to stain with the conventional protein stains such as Coomassie blue). The protein was then purified in large amounts from human red cell membranes (Denker et al., 1988; Smith and Agre, 1991). The 28 kDa protein was strikingly abundant. With approximately copies per red cell, it was one of the major proteins in the membrane. The protein had features suggesting that it was a tetrameric membrane-spanning protein – suggesting that it was a channel, but a channel for what? The purified protein also provided us the N-terminal amino acid sequence that we used for cDNA cloning. Using our antibody, we looked at several other tissues and found the protein is also strikingly abundant in human kidney. We observed staining over the apical and basolateral membranes of proximal renal tubules and the descending thin limb of the loops of Henle, but we were still frustrated by our failure to recognize what the protein’s function might be. My clinical mentor, John C. Parker (1935–1993) at the University of North Carolina at Chapel Hill, was the first to suggest to me that red cells and renal tubules were exceedingly permeable to water. He recommended that we consider a role in membrane water transport. While John did not live to see our later studies, he did live to see our initial discovery and we celebrated together.
Postdoctoral fellow Gregor y Preston cloned the cDNA from an erythroid brary (Preston and Agre, 1991). The coding region corresponded to a 269 amino acid polypeptide, predicted by hydropathy analysis to have six bilayer-spanning domains. Interestingly, the amino terminal half (repeat-1) and the carboxy terminal half of the molecule (repeat-2) were genetically related – about 20% identical. Two loops B and E were more highly related to each other, and each contained the signature motif – asparagine, proline, alanine (NPA) [Fig. 1]. Examining the genetics database, we recognized several sequence-related DNAs from diverse sources: lens of cow eyes, brains of fruit flies, bacteria, and plants. Nevertheless, none was functionally defined.
Figure 1. Membrane orientation of AQP1 predicted from primary amino acid sequence. Two tandem repeats each have three bilayer-spanning domains; the repeats are oriented 180˚ with respect to each other. The loops B and E each contain the conser ved motif, Asn- Pro-Ala (NPA)
These clues heightened our suspicion that the 28 kDa protein was a transporter, so we tested for possible water transport function with our colleague Bill Guggino at Johns Hopkins. We used oocytes the frog Xenopus laevis, a useful model, since frog oocytes have very low water permeability. Control oocytes were injected with water alone; oocytes were injected with 2 ng of cRNA encoding our protein. After days of protein synthesis, the oocytes appeared essentially identical. Then we stressed the oocytes by transferring them to distilled water, and an amazing difference was immediately apparent. Having exceedingly low water perme-
ability, the control oocytes failed to swell. In contrast, the test oocytes were highly permeable to water and exploded like popcorn [Fig. 2] (Preston et al., 1992). The protein was christened “aquaporin” and is now officially designated “AQP1,” the first functionally defined water channel protein (Agre et al., 1993b).
Figure 2. Functional expression of AQP1 water channels in Xenopus laevis oocytes. Control oocyte (left) was injected with water; AQP1 oocyte (right) was injected with cRNA. The oocytes were transferred to hypotonic buffer. After 30 seconds (top) the AQP1 oocyte has begun to swell; after 3 minutes (bottom), the AQP1 oocyte has exploded. Modified and reprinted from Science with permission (Preston et al., 1992).
We confirmed the function of this protein by studying the purified AQP1 reconstituted into synthetic lipid vesicles of ~0.1 micron diameter prepared by our colleague Suresh Ambudkar at Johns Hopkins (Zeidel et al., 1992). These simple membrane vesicles were examined by freeze fracture electron microscopy by our colleague Arvid Maunsbach, from the University of Aarhus. When lipid was reconstituted without protein, the membrane surfaces were smooth; however, membranes reconstituted with AQP1 contained many intramembraneous particles 0.01 micron diameter (Zeidel et al., 1994). We tested the membranes for water permeability in collaboration with Mark Zeidel at Har vard Medical School. Using stopped flow transfer to hypertonic buffer, the simple liposomes shrank, reaching equilibrium in about one half
second; this is believed to represent the baseline water permeability. When membranes reconstituted with AQP1 were examined, the shrinking occurred much more rapidly, reaching equilibrium in about 20 milliseconds. The channel-mediated flow of water was confirmed, since it was inhibited with mercurials. We calculated the Arrhenius activation energy (<5 kcal/mol), and we determined the unit permeability to be ~3×10 water molecules per subunit per second. Importantly, we attempted to measure proton permeation of AQP1, but despite massive water permeability, acid permeation was not detected. These studies verified that we had, in fact, isolated the long-sought water channel protein.
STRUCTURE OF AQP1
Subsequent efforts were devoted to identifying the mercurial inhibitory site predicted by the studies of Macey. Mercurials react with free sulfhydryls in the amino acid cysteine. Four cysteines are found in the AQP1 polypeptide, but only the residue in loop E (Cys-189 proximal to the second NPA motif) is inhibited by mercurials. We altered the AQP1 sequence by site-directed mutagenesis and expressed the recombinants in oocytes for water permeability studies. Mutation of this residue to serine (Cys-189-Ser) resulted in full water permeability without mercurial inhibition. When we then replaced the alanine in the corresponding position of loop B with a cysteine (Ala-73-Cys), the protein exhibited mercurial sensitive water permeability (Preston et al., 1993). Substitutions elsewhere in the AQP1 failed to produce this behavior. This suggested to us that loops B and E in opposite parts of the molecule must somehow form the aqueous pore. The model that we concocted turned out to be schematically correct and was termed “the hourglass.” The ancient timepiece allows sand to run from upper chamber to lower chamber; if inverted, the sand will flow in the opposite direction. Six bilayer spanning domains were predicted to surround a central domain containing loop B, dipping into the membrane from the cytoplasmic surface, and loop E, dipping into the membrane from the extracellular surface [Fig. 3 left and right].
Figure 3. Hourglass model for membrane topology of AQP1 subunit.
Left panel – Schematic folding of loops B and E overlap within the lipid bilayer to form a single aqueous pathway.
Right panel – Ribbon model of three dimensional structure of AQP1 subunit confirms hourglass with single aqueous pathway. Modified and reprinted with permission from Jour-
nal of Biological Chemistr y (Jung et al., 1994b) and Journal of Clinical Investigation (Kozono et al., 2002).
The overlap of loops B and E was predicted to form a single aqueous pore through the center of the molecule with the NPA motifs juxtaposed and mercurial inhibitory site alongside (Jung et al., 1994b). The AQP1 protein tetrameric with a central pore in each subunit. Thus, AQP1 is structurally like ion channel proteins where four subunits surround a single central as discussed by Rod MacKinnon in his lecture.
We then sought to establish the high resolution structure of AQP1 in collaboration with Andreas Engel and his group at the Biozentrum in Basel. We were later joined with Yoshinori Fujiyoshi and his group at Kyoto University. Human red cell AQP1 protein was purified by Barb Smith in our lab; Andreas’s student Tom Walz reconstituted it into synthetic membranes at very high protein concentrations. Under these conditions, the AQP1 protein forms remarkably symmetrical arrays referred to as membrane crystals. By measuring the water permeability, we confirmed that the function was 100% retained, giving us confidence that the structure we deduced would be the biologically relevant structure (Walz et al., 1994).
Figure 4. Functional representation for selective water flow through AQP1 subunit and residues involved in human disease.
Left panel – Schematic of sagittal cross-section of AQP1 reveals bulk water in extracellu- lar and intracellular vestibules of hourglass. These are separated by a 20Å span where water passes in single file with transient interactions with pore-lining residues that prevent hy- drogen bonding between water molecules (bold colors). Two structures are believed to pre- vent permeation by protons (H O ): electrostatic repulsion is created by a fixed positive
charge from pore-lining arginine (R195) at a 2.8Å narrowing in the channel; water dipole reorientation occurs from simultaneous hydrogen bonding of water molecule with side chains of two asparagines residues in NPA motifs (N192 and N76). Two partial positive charges at the center of the channel result from orientation of two non-membrane span- ning alpha helices distal to the NPA motifs
THE AQUAPORIN AND AQUAGLYCEROPORIN PROTEIN FAMILY
While we were pursuing studies of AQP1, several other research groups from around the world became interested in what is now known to be a large family of related proteins. The combined efforts of these labs have led to the molecular identification of 12 mammalian aquaporin homologs, and several hundred related proteins have been recognized in other vertebrates as well as invertebrates, plants, and unicellular micro-organisms. The mammalian homologs may be loosely clustered into two subsets [Fig. 5]. The first is referred to as “classical aquaporins”, since they were initially considered to be exclusive water pores. The second is referred to as “aquaglyceroporins”, since they are permeated by water plus glycerol. Interestingly, E. coli has one member of
each – AqpZ (Calamita et al., 1995), and GlpF, isolated by other investigators much earlier. Together, the mammalian aquaporins and aquaglyceroporins are now known to contribute to multiple physiological processes that occur during our daily lives.
Figure 5. Human aquaporin gene family contains two subsets. Homologs freely permeated by water (classical aquaporins, blue) or water and glycerol (aquaglyceroporins, yellow) are represented. E. coli has one aquaporin (AqpZ) and one aquaglyceroporin (GlpF). Reprinted with permission from Journal of Physiology (Agre et al., 2002)
The remainder of the Nobel Lecture (2003) can be found at the Nobel Prize site. This portion is sufficient to cover the genesis and advancement of the water transport discovery.
Urinary Excretion of Aquaporin-2 Water Channel Differentiates Psychogenic Polydipsia from Central Diabetes Insipidus
T Saito, San-e Ishikawa, T Ito, H Oda, F Ando, … and T Saito Division of Endocrinology and Metabolism (Ta.S., S.I., F.A., Mi.H., S.N., To.S.), Department of Medicine, Jichi Medical School, Tochigi 329-0498; and Departments of Medicine and Psychiatry (T.I., H.O., Ma.H.), Tokyo Metropolitan Matsuzawa Hospital, Tokyo, Jp
correspondence to: San-e Ishikawa, M.D., Division of Endocrinology and Metabolism, Department of Medicine, Jichi Medical School, Tochigi 329-0498, Japan. E-mail: saneiskw@jichi.ac.jp.
http://jcem.endojournals.org/full/84/6/2235The present study was undertaken to determine whether urinary excretion of aquaporin-2 (AQP-2) water channel under ad libitum water intake is of value to differentiate polyuria caused by psychogenic polydipsia from central diabetes insipidus. A 30-min urine collection was made at 0900 h in 3 groups of: 11 patients with central diabetes insipidus (22–68 yr old), 10 patients with psychogenic polydipsia (28–60 yr old), and 15 normal subjects (21–38 yr old). In the patients with central diabetes insipidus, the plasma arginine vasopressin level was low despite hyperosmolality, resulting in hypotonic urine. Urinary excretion of AQP-2 was 37 ± 15 fmol/mg creatinine, a value one-fifth less than that in the normal subjects. In the patients with psychogenic polydipsia, plasma arginine vasopressin and urinary osmolality were as low as those in the patients with central diabetes insipidus. However, urinary excretion of AQP-2 of 187 ± 45 fmol/mg creatinine was not decreased, and its excretion was equal to that in the normal subjects. These results indicate that urinary excretion of AQP-2, under ad libitum water drinking, participates in the differentiation of psychogenic polydipsia from central diabetes insipidus.
PSYCHOGENIC polydipsia causes a marked polyuria with hypotonic urine (1, 2). Arginine vasopressin (AVP) secretion is suppressed by hypoosmolality caused by excess intake of water. Suppression of AVP release obliges us to differentiate psychogenic polydipsia from central diabetes insipidus. Osmotic stimulation tests have been carried out to determine the reserve function of the posterior pituitary gland. Plasma AVP levels increase in response to an increase in plasma osmolality (Posm) in patients with psychogenic polydipsia but not in those with central diabetes insipidus.
In response to AVP, concentrated urine is produced by water reabsorption across the renal collecting duct (3, 4). Aquaporin-2 (AQP-2) is an AVP-regulated water channel of the collecting duct; it is translocated from the cytoplasmic vesicles to the apical plasma membranes by shuttle trafficking when the cells are stimulated by AVP (5, 6, 7), and it is again redistributed into the cytoplasmic vesicles after removal of AVP stimulation (8). Also, AQP-2 is, in part, excreted into the urine (9, 10). We demonstrated that urinary excretion of AQP-2 is of great value in diagnosing central diabetes insipidus in the hypertonic saline infusion test and impaired water excretion in the acute oral water load test (11, 12). The present study was undertaken to determine whether urinary excretion of AQP-2, under ad libitum water intake, is a useful tool for diagnosing psychogenic polydipsia.
Subjects and study design
Three groups of subjects were examined in the present study.
[1] 11 patients who had been diagnosed as having idiopathic central diabetes insipidus. They had taken 1-deamino-8-D-AVP (DDAVP) intranasally, twice a day, and discontinued the DDAVP therapy 24 h before the study.
[2] 10 patients were diagnosed as having psychogenic polydipsia. They had been treated for psychiatric disorders, including schizophrenia, atypical psychiatric disorder, and chronic alcoholism.
[3] 15 normal volunteers, with ages ranging from 21–38 yr. (the age range of [1] and [2] reached 60)
All the subjects drank water ad libitum, and 30-min urine collection was made and blood drawn at 0900 h. Urine samples were subjected to measurements of urinary osmolality (Uosm) and urinary excretion of creatinine and AQP-2. Blood samples were used to measure Posm and plasma AVP levels. Uosm and Posm were measured by freezing-point depression (Model 3W2, Advanced Instruments, Needham Height, MA). Urinary creatinine was measured with an automatic clinical analyzer (Model 736, Hitachi Co., Tokyo, Jp). Plasma AVP levels were determined by RIA using AVP RIA kits (Mitsubishi Chemistry, Tokyo, Jp) (13). Urinary excretion of AQP-2 was measured as described below.
RIA of AQP-2
The RIA of urinary AQP-2 was performed by the method described in our previous reports (11, 12). Urinary AQP-2-like immunoreactivity was measured by a specific RIA that used the polyclonal antibody against a synthetic portion (Tyr0-AQP-2[ V257-A271]) of the C-terminal of human AQP-2 raised in rabbits. A synthetic peptide [Tyr0-AQP-2 (V257-A271)] was radioiodinated with iodine-125 (New England Nuclear, Boston, MA) by the chloramine-T method. All samples were analyzed in duplicate. The intra- and interassay coefficients of variation were less than 10%. The minimal detectable quantity of AQP-2 was 0.86 pmol/tube, and an amount equivalent to 6.9 pmol/tube caused 50% inhibition of binding of the radiolabeled ligand.
Results
In the patients with central diabetes insipidus, the plasma AVP level was low despite hyperosmolality of 297.8 ± 3.4 mosmol/kg H2O, resulting in hypotonic urine (Fig. 1⇓). Urinary excretion of AQP-2 was one-fifth less in the patients with central diabetes insipidus than in the normal subjects. AQP-2 is the AVP-dependent water channel of collecting duct cells and is recycling between the cytoplasmic vesicles and the apical plasma membranes in the cells (5, 6, 7, 8). AQP-2 is partly excreted into the urine, which is approximately 3% of AQP-2 in the collecting duct cells (14). In normal subjects, urinary excretion of AQP-2 is changeable in a wide range in physiological conditions (11). Because urinary excretion of AQP-2 has a positive correlation with plasma AVP levels in normal subjects (11), the reduced urinary excretion of AQP-2 was in concert with the impaired secretion of AVP in central diabetes insipidus.
Figure 1.
Posm, plasma AVP (Pavp), Uosm, and urinary excretion of AQP-2 (UAQP-2), under ad libitum water drinking, in 15 normal subjects (NL, •), 11 patients with central diabetes insipidus (CDI, ○) and 10 patients with psychogenic polydipsia (PP, □). *, P < 0.01; **, P < 0.05 vs. the normal subjects. Value are means ± sem.
In the patients with psychogenic polydipsia, Uosm was as low as that in the patients with central diabetes insipidus (Fig. 1⇑). The plasma AVP level was low because of the reduced Posm, which was derived from an exaggerated intake of water. Urinary excretion of AQP-2, however, was not decreased; and rather, its excretion kept the normal range. The relationship between plasma AVP levels and urinary excretion of AQP-2 is shown in Fig. 2⇓. The urinary excretion of AQP-2 in the patients with psychogenic polydipsia was dissociated from the positive correlation between plasma AVP and urinary excretion of AQP-2 in the normal subjects.
Figure 2.
Relationship between plasma AVP levels and UAQP-2. •, Normal subjects (n = 15); ○, patients with central diabetes insipidus (n = 11); □, patients with psychogenic polydipsia (n = 10). Values are means ± sem.
Discussion
The present study demonstrated the clinical tool, of urinary excretion of AQP-2, in differentiating psychogenic polydipsia from central diabetes insipidus. What is involved in the marked difference in urinary excretion of AQP-2 in these two disorders? There is a possibility that, as patients with psychogenic polydipsia reduce water intake during sleep, antidiuresis may occur periodically at night and the production of AQP-2 be somewhat restored. Because approximately 3% of AQP-2 in collecting duct cells is excreted into the urine, urinary excretion of AQP-2 may keep relatively high, despite hypotonic urine. The difference may come from the periodicity of water intake in a day, in the patients with psychogenic polydipsia. As a whole, these changes may disrupt the positive relationship between urinary excretion of AQP-2 and plasma AVP levels. At the present time, however, other factors involved in urinary excretion of AQP-2 remain undetermined.
In conclusion, urinary excretion of AQP-2, under ad libitum water drinking, participates in the differentiation of polyuria caused by psychogenic polydipsia from central diabetes insipidus.
References
Jose CI, Perez-Cruet J. 1979 Incidence and morbidity of self-induced water intoxication in state mental hospital patients. Am J Psychiatry. 136:221–222. Medline
Goldman MB, Luchins DJ, Robertson GL. 1988 Mechanisms of altered water metabolism in psychiatric patients with polydipsia and hyponatremia. N Engl J Med. 318:397–403. Medline
Ishikawa S. 1993 Cellular action of arginine vasopressin in the kidney. Endocr J. 40:373–386. Medline
Fushimi K, Uchida S, Hara Y, Hirata Y, Marumo F, Sasaki S. 1993 Cloning and expression of apical membrane water channel of rat kidney collecting tubule. Nature. 361:549–552. CrossRefMedline
Sasaki S, Fushimi K, Saito H, et al. 1994 Cloning, characterization and chromosomal mapping of human aquaporin of collecting duct. J Clin Invest. 93:1250–1256.
Nielsen S, DiGiovanni SR, Christensen EI, Knepper MA, Harris HW. 1993 Cellular and subcellular immunolocalization of vasopressin-regulated water channel in rat kidney. Proc Natl Acad Sci USA. 90:11663–11667. Abstract/FREE Full Text
Saito T, Ishikawa S, Sasaki S, et al. 1997 Alteration in water channel AQP-2 by removal of AVP stimulation in collecting duct cells of dehydrated rats. Am J Physiol. 272:F183–F191.
Kanno K, Sasaki S, Ishikawa S, et al. 1995 Urinary excretion of aquaporin-2 in patients with diabetes insipidus. N Engl J Med. 332:1540–1545. CrossRefMedline
Elliot S, Goldsmith P, Knepper MA, Haughey M, Olson B. 1996 Urinary excretion of aquaporin-2 in humans: a potential marker of collecting duct responsiveness to vasopressin. J Am Soc Nephrol. 7:403–409. Abstract
Saito T, Ishikawa S, Sasaki S, et al. 1997 Urinary excretion of aquaporin-2 in the diagnosis of central diabetes insipidus. J Clin Endocrinol Metab. 82:1823–1827. Abstract/FREE Full Text
Saito T, Ishikawa S, Ando F, et al. 1998 Exaggerated urinary excretion of aquaporin-2 in the pathological state of impaired water excretion dependent upon arginine vasopressin. J Clin Endocrinol Metab. 83:4034–4040. Abstract/FREE Full Text
Terris J, Ecelbarger CA, Nielsen S, Knepper MA. 1996 Long-term regulation of four renal aquaporins in rats. Am J Physiol. 271:F414–F422. Clin Endocrinology & Metabolism 1999; 84(6):2235-2237
http://dx.doi.org/10.1210/jc.84.6.2235Comparison of cardiovascular aquaporin-1 changes during water restriction between 25- and 50-day-old rats.
Netti VA, Vatrella MC, Chamorro MF, Rosón MI, Zotta E, Fellet AL, Balaszczuk AM.
Cátedra de Fisiología, Facultad de Farmacia y Bioquímica, Universidad de Buenos Aires, IQUIMEFA, CONICET, Junín 956, C1113AAD, Buenos Aires, Argentina, vnetti@conicet.gov.ar.
Eur J Nutr. Apr 27, 2013
Aquaporin-1 (AQP1) is the predominant water channel in the heart, linked to cardiovascular homeostasis. Our aim was to study cardiovascular AQP1 distribution and protein levels during osmotic stress and subsequent hydration during postnatal growth.
Rats aged 25 and 50 days were divided in: 3d-WR: water restriction 3 days; 3d-WAL: water ad libitum 3 days; 6d-WR+ORS: water restriction 3 days + oral rehydration solution (ORS) 3 days; and 6d-WAL: water ad libitum 6 days. AQP1 was evaluated by immunohistochemistry and western blot in left ventricle, right atrium and thoracic aorta.
Water restriction induced a hypohydration state in both age groups (40 and 25 % loss of body weight in 25- and 50-day-old rats, respectively), reversible with ORS therapy. Cardiac AQP1 was localized in the endocardium and endothelium in both age groups, being evident in cardiomyocytes membrane only in 50-day-old 3d-WR group, which presented increased protein levels of AQP1; no changes were observed in the ventricle of pups. In vascular tissue, AQP1 was present in the smooth muscle of pups; in the oldest group, it was found in the endothelium, increasing after rehydration in smooth muscle. No differences were observed between control groups 3d-WAL and 6d-WAL of both ages.
Our findings suggest that cardiovascular AQP1 can be differentially regulated in response to hydration status in vivo, being this response dependent on postnatal growth. The lack of adaptive mechanisms of mature animals in young pups may indicate an important role of this water channel in maintaining fluid balance during hypovolemic state.
Clinical application of aquaporin research: aquaporin-1 in the peritoneal membrane
Nishino T, Devuyst O.
Division of Renal Care Unit, Second Department of Internal Medicine, Nagasaki University School of Medicine, Nagasaki, Jp
Peritoneal dialysis (PD) is an established mode of renal replacement therapy based on the exchange of fluid and solutes between blood and a dialysate that has been instilled in the peritoneal cavity. The dialysis process involves osmosis, as well as diffusive and convective transports through the highly vascularized peritoneal membrane. The membrane contains ultrasmall pores responsible for the selective transport of water across the capillary endothelium. The distribution of the water channel aquaporin-1 (AQP1), as well as its molecular structure ensuring an exquisite selectivity for water, fit with the characteristics of the ultrasmall pore. Peritoneal transport studies using AQP1 knockout mice demonstrated that the osmotic water flux across the peritoneal membrane is mediated by AQP1. This water transport accounts for 50% of the ultrafiltration during PD. Treatment with high-dose corticosteroids upregulates the expression of AQP1 in peritoneal capillaries, resulting in increased water transport and ultrafiltration in rats. These data illustrate the potential of the peritoneal membrane as an experimental model in the investigation of the role of AQP1 in the endothelium. They emphasize the critical role of AQP1 during PD and suggest that manipulating AQP1 expression could be clinically useful in PD patients.
Stoenoiu MS, Ni J, Verkaeren C, Debaix H, Jonas JC, Lameire N, Verbavatz JM, Devuyst O.
Division of Nephrology and ENDO Unit, Université Catholique de Louvain Medical School, Brussels, Belgium
J Am Soc Nephrol. Mar 2003; 14(3):555-565.
The water channel aquaporin-1 (AQP1) is the molecular counterpart of the ultrasmall pore responsible for transcellular water permeability during peritoneal dialysis (PD). This water permeability accounts for up to 50% of ultrafiltration (UF) during a hypertonic dwell, and its loss can be a major clinical problem for PD patients. By analogy with the lung, the hypothesis was tested that corticosteroids may increase AQP1 expression in the peritoneal membrane (PM) and improve water permeability and UF in rats. First, the expression and distribution of the glucocorticoid receptor (GR) in the PM and capillary endothelium was documented. Time-course and dose-response analyses showed that a daily IM injection of dexamethasone (1 or 4 mg/kg) for 5 d induced an approximately twofold increase in the expression of AQP1 at the mRNA and protein levels. The GR antagonist RU-486 completely inhibited the dexamethasone effect. The functional counterpart of the increased AQP1 expression was a significant increase in sodium sieving and net UF across the PM, contrasting with a lack of effect on the osmotic gradient and permeability for small solutes. The latter observation reflected the lack of effect of corticosteroids on nitric oxide synthase (NOS) activity and endothelial NOS isoform expression in the PM. In conclusion, corticosteroids induce AQP1 expression in the capillary endothelium of the PM, which is reflected by increased transcellular water permeability and UF. These data emphasize the critical role of AQP1 during PD and suggest that pharmacologic regulation of AQP1 may provide a target for manipulating water permeability across the PM.
Aquaporins: relevance to cerebrospinal fluid physiology and therapeutic potential in hydrocephalus
Owler BK, Pitham T, Wang D.
Kids Neurosurgical Research Unit, Children’s Hospital at Westmead, Westmead NSW 2145, Australia. brian@sydneyneurosurgeon.com.au.
The discovery of a family of membrane water channel proteins called aquaporins, and the finding that aquaporin 1 was located in the choroid plexus, has prompted interest in the role of aquaporins in cerebrospinal fluid (CSF) production and consequently hydrocephalus. While the role of aquaporin 1 in choroidal CSF production has been demonstrated, the relevance of aquaporin 1 to the pathophysiology of hydrocephalus remains debated. This has been further hampered by the lack of a non-toxic specific pharmacological blocking agent for aquaporin 1. In recent times aquaporin 4, the most abundant aquaporin within the brain itself, which has also been shown to have a role in brain water physiology and relevance to brain oedema in trauma and tumours, has become an alternative focus of attention for hydrocephalus research. This review summarises current knowledge and concepts in relation to aquaporins, specifically aquaporin 1 and 4, and hydrocephalus. It also examines the relevance of aquaporins as potential therapeutic targets in hydrocephalus and other CSF circulation disorders.
PMID: 20860832 PMCID: PMC2949735
Pathophysiology of the aquaporin water channels
King LS, Agre P.
Department of Medicine, Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, USA.
Annu Rev Physiol. 1996; 58:619-48.
Discovery of aquaporin water channel proteins has provided insight into the molecular mechanism of membrane water permeability. The distribution of known mammalian aquaporins predicts roles in physiology and disease.
Aquaporin-1 mediates proximal tubule fluid reabsorption, secretion of aqueous humor and cerebrospinal fluid, and lung water homeostasis.
Aquaporin-2 mediates vasopressin-dependent renal collecting duct water permeability; mutations or downregulation can cause nephrogenic diabetes insipidus.
Aquaporin-3 in the basolateral membrane of the collecting duct provides an exit pathway for reabsorbed water.
Aquaporin-4 is abundant in brain and probably participates in reabsorption of cerebrospinal fluid, osmoregulation, and regulation of brain edema.
Aquaporin-5 mediates fluid secretion in salivary and lacrimal glands and is abundant in alveolar epithelium of the lung.
Specific regulation of membrane water permeability will likely prove important to understanding edema formation and fluid balance in both normal physiology and disease.
Discovery of aquaporins: a breakthrough in research on renal water transport
van Lieburg AF, Knoers NV, Deen PM.
Department of Pediatrics, University of Nijmegen, The Netherlands.
Pediatr Nephrol. Apr 1995; 9(2):228-34.
Several membranes of the kidney are highly water permeable, thereby enabling this organ to retain large quantities of water. Recently, the molecular identification of water channels responsible for this high water permeability has finally been accomplished. At present, four distinct renal water channels have been identified, all members of the family of major intrinsic proteins.
Aquaporin 1 (AQP1), aquaporin 2 (AQP2) and the mercury-insensitive water channel (MIWC) are water-selective channel proteins, whereas the fourth,
Aquaporin 3 (AQP3), permits transport of urea and glycerol as well. Furthermore, a putative renal water channel (WCH3) has been found.
AQP1 is expressed in apical and basolateral membranes of proximal tubules and descending limbs of Henle,
AQP2 predominantly in apical membranes of principal and inner medullary collecting duct cells and
AQP3 in basolateral membranes of kidney collecting duct cells.
MIWC is expressed in the inner medulla of the kidney and has been suggested to be localised in the vasa recta.
The human genes encoding AQP1 and AQP2 have been cloned, permitting deduction of their amino acid sequence, prediction of their two-dimensional structure by hydropathy analysis, speculations on their way of functioning and DNA analysis in patients with diseases possibly caused by mutant aquaporins. Mutations in the AQP1 gene were recently detected in clinically normal individuals, a finding which contradicts the presumed vital importance of this protein. Mutations in the AQP2 gene were shown to cause autosomal recessive nephrogenic diabetes insipidus. The renal unresponsiveness to arginine vasopressin, which characterises this disease, is in accordance with the assumption that AQP2 is the effector protein of the renal vasopressin pathway.(ABSTRACT TRUNCATED AT 250 WORDS)
Selectivity of the renal collecting duct water channel aquaporin-3
Echevarría M, Windhager EE, Frindt G.
Depart Physiol Biophys, Cornell University Medical College, New York, NY
J Biol Chem. Oct 11, 1996; 271(41):25079-82.
Aquaporin-3 (AQP3) is a water channel found in the basolateral cell membrane of principal cells of the renal collecting tubule as well as in other epithelia. To examine the selectivity of AQP3, the permeability to water (Pf), urea (Pur), and glycerol (Pgly) of Xenopus oocytes injected with cRNA encoding AQP3 was measured. Oocytes injected with cRNA encoding either human or rat aquaporin-1 (AQP1) were used as controls. Although both aquaporins permit water flow across the cell membrane, only AQP3 was permeable to glycerol and urea (Pgly > Pur). The uptake of glycerol into oocytes expressing AQP3 was linear up to 165 mM. For AQP3 the Arrhenius energy of activation for Pf was 3 kcal/mol, whereas for Pgly and Pur it was >12 kcal/mol. The sulfhydryl reagent p-chloromercuriphenylsulfonate (1 mM) abolished Pf of AQP3, whereas it did not affect Pgly. In addition, phloretin (0.1 mM) inhibited Pf of AQP3 by 35%, whereas it did not alter Pgly or Pur. We conclude that water does not share the same pathway with glycerol or urea in AQP3 and that this aquaporin, therefore, forms a water-selective channel.
The aquaporin family of water channels in kidney
Agre P, Nielsen S.
Depart of Med, Johns Hopkins University School of Medicine, Baltimore, MD
Nephrologie. 1996;17(7):409-15.
The longstanding puzzle of membrane water-permeability was advanced by discovery of a new class of proteins known as the “aquaporins” (AQPs). First identified in red blood cells, AQP1 was shown to function as a water channel when expressed in Xenopus oocytes or when pure AQP1 protein was reconstituted into synthetic membranes. Analysis of the primary sequence revealed that the two halves of the AQP1 polypeptide are tandem repeats; site directed mutagenesis studies indicate that the repeats may fold into an obversely symmetric structure which resembles an hourglass. Electron crystallography elucidated the tetrameric organization of AQP1, and functional studies suggest that each tetramer contains multiple functionally independent aqueous pores.
AQP1 is abundant in the apical and basolateral membranes of renal proximal tubules and descending thin limbs, and is also present in multiple extra renal tissues.
AQP2 is expressed only in the principal cells of renal collecting duct where it is the predominant vasopressin (ADH, antidiuretic hormone) regulated water channel. AQP2 is localized in the apical membrane and in intracellular vesicles which are targeted to the apical plasma membranes when stimulated by ADH. Humans with mutations in genes encoding AQP1 and AQP2 exhibit contrasting clinical phenotypes.
AQP3 resides in the basolateral membranes of renal collecting duct principal cells providing an exit pathway for water;
AQP4 is abundant in brain where it may function as the hypothalamic osmoreceptor responsible for secretion of ADH. Continued analysis of the aquaporins is providing detailed molecular insight into the fundamental physiological problems of water balance and disorders of water balance.
Aquaporins in the kidney: from molecules to medicine
Nielsen S, Frøkiaer J, Marples D, Kwon TH, Agre P, Knepper MA.
The Water and Salt Res Center, Anatomy and Exper Clin Res Institutes, University of Aarhus, Aarhus, Denmark. sn@ana.au.dk
The molecular identity of membrane water channels long-standing biophysical question of how water crosses long remained elusive until the pioneering discovery of biological membranes specifically, and provided insight, at the molecular level, of AQP1 by Agre and colleagues around 1989 –1991, The discovery of aquaporin-1 (AQP1) answered the long-standing biophysical question of how water specifically crosses biological membranes. In the kidney, at least seven aquaporins are expressed at distinct sites. AQP1 is extremely abundant in the proximal tubule and descending thin limb and is essential for urinary concentration. AQP2 is exclusively expressed in the principal cells of the connecting tubule and collecting duct and is the predominant vasopressin-regulated water channel. AQP3 and AQP4 are both present in the basolateral plasma membrane of collecting duct principal cells and represent exit pathways for water reabsorbed apically via AQP2. Studies in patients and transgenic mice have demonstrated that both AQP2 and AQP3 are essential for urinary concentration.
Since the discovery of aquaporins, major efforts have been aimed at elucidating their structural organization. Hydropathy analysis of the deduced amino acid sequence of AQP1 led to the prediction that the protein resides primarily within the lipid bilayer (191), consistent with the initial studies of AQP1 in red cell membranes (46). AQP1 contains an internal repeat with the NH – and the first provided a molecular answer to the long-standing COOH-terminal halves being sequence related and each
containing the signature motif Asn-Pro-Ala (NPA) (181,252). This is consistent with earlier observations on the homologous major intrinsic protein from lens, (MIP, nowreferred to as AQP0). When evaluated by hydropathy analysis, six bilayer-spanning domains are apparent (Fig.1); however, the apparent interhelical loops B and E also exhibit significant hydrophobicity. Critical to the topology is the location of loop C which connects the two halves of the molecule. Preston et al. (194) demonstrated that loop C resides at the extracellular surface of the oocytes, confirming the obverse sym-metry of the NH – and COOH-terminal halves of the mol-lar surface of the oocytes, confirming the obverse symmetry of the NH – and COOH-terminal halves of the mol-ecule.The structural organization of other aquaporins such as bacterial aquaporin-Z and plant aquaporins have also been deduced. How can water channels avoid passage of protons (H O )? As predicted, loops B and E are associated by Van der Waals interactions between the two NPA motifs. Free hydrogen bonding occurs in the column of water within the pore, except at the very center where a single water molecule transiently reorients to bond with the two asparagines residues of the NPA motif. This results in minimum resistance to the flow of water, thus permitting kidneys to perform their important physiological roles of reabsorbing water while excreting acid.
FIG. 1. A: schematic representation of the structural organization of aquaporin-1 (AQP1) monomers in the membrane (top and bottom). Aquaporins have six membrane-spanning regions, both intracellular NH and COOH termini, and internal tandem repeats that, presumably, are due to an ancient gene duplication (top). The topology is consistent with an obverse symmetry for the two similar NH – and COOH- 2 terminal halves (bottom). The tandem repeat structure with two asparagine-proline-alanine (NPA) sequences has been proposed to form tight turn structures that interact in the membrane to form the pathway for translocation of water across the plasma membrane. Of the five loops in AQP1, the B and E loops dip into the lipid bilayer, and it has been proposed that they form “hemichannels” that connect between the leaflets to form a single aqueous pathway within a symmetric structure that resembles an “hourglass.” B: AQP1 is a multisubunit oligomer that is organized as a tetrameric assembly of four identical polypeptide subunits with a large glycan attached to only one.
Discovery and Biophysical Characterization of the First Molecular Water Channel AQP1 Expression of AQP1 in X. laevis oocytes by Preston et al. (192) demonstrated that AQP1-expressing oocytes exhibited remarkably high osmotic water permeability (P
cm/s), causing the cells to swell rapidly and explode in hypotonic buffer. The osmotically induced swelling of oocytes expressing AQP1 occurs with a low activation energy and is reversibly inhibited by HgCl or other mercurials. Only inward water flow (swelling) was examined, but it was predicted that the direction of water flow through AQP1 is determined by the orientation of the osmotic gradient. Consistent with this, it was later demonstrated that AQP1-expressing oocytes swell in hyposmolar buffers but shrink in hyperosmolar buffers (160). Swelling of oocytes expressing AQP1 occurs with a low activation energy and is reversibly inhibited by HgCl or other mercurials. Only inward water flow (swelling) was examined, but it was predicted that the direction of water flow through AQP1 is determined by the orientation of the osmotic gradient. Consistent with this, it was later demonstrated that AQP1-expressing oocytes swell in hyposmolar buffers but shrink in hyperosmolar buffers (160).
Over the past 4 years a series of studies have explored the issues of selectivity and polytransport function of aquaporins. This has led to a division of aquaporins (4) into a group that transports water relatively selectively (the “orthodox” set or “aquaporins”) and a group of water channels that also conduct glycerol and other small solutes in addition to water (the “cocktail” set or aquaglyceroporins). This appears to represent an ancient phylogenetic divergence between glycerol transporters and pure water channels (185). Recently, it has become clear that transport properties are even more diverse, since AQP6 has been demonstrated to conduct anions as well (263), and it has also been demonstrated that aquaporins can be regulated by gating, as discussed below.
The signal transduction pathways have been described thoroughly in previous reviews. cAMP levels in collecting duct principal cells are increased by binding of vasopressin to V2 receptors. The synthesis of cAMP by adenylate cyclase is stimulated by a V2 receptor-coupled heterotrimeric GTP-bind-ing protein, Gs. Gs interconverts between an inactive responses to vasopressin. In this study it was demonstrated that changes in AQP2 labeling density of the apical plasma membrane correlated closely with the water permeability in the same tubules, while there were reciprocal changes in the intracellular labeling for AQP2. In vivo studies using normal rats or vasopressin-deficient Brattleboro rats also showed a marked increase in apical plasma membrane labeling of AQP2 in response to vasopressin or dDAVP treatment. The acute treatment of rats with vasopressin V2-receptor antagonist or acute water loading (to reduce endogenous vasopressin levels, both reducing vasopressin action, resulted in a prominent internalization of AQP2 from the apical plasma membrane to small intracellular vesicles further underscoring the role of AQP2 trafficking in the regulation of collecting duct water permeability.
PGE2 inhibits vasopressin-induced water permeability by reducing cAMP levels. In preliminary studies, Zelenina et al. investigated the effect of PGE2 on PKA phosphorylation of AQP2 in kidney papilla, and the results suggest that the action of prostaglandins is associated with retrieval of AQP2 from the plasma membrane, but that this appears to be independent of AQP2 phosphorylation by PKA. Phosphorylation of AQP2 by other kinases, e.g., protein kinase C or casein kinase II, may potentially participate in regulation of AQP2 trafficking (Fig. 9C). Phosphorylation of other cytoplasmic or vesicular regulatory proteins may also be involved. These issues remain to be investigated directly.
Since the fundamentals of the shuttle hypothesis have been confirmed, interest has turned to the cellular mechanisms mediating the vasopressin-induced transfer of AQP2 to the apical plasma membrane. The shuttle hypothesis has a number of features whose molecular basis remains poorly understood. First, AQP2 is delivered in a relatively rapid and coordinated fashion, and vesicles move from a distribution throughout the cell to the apical region of the cell in response to vasopressin stimulation. Furthermore, AQP2 is delivered specifically to the apical plasma membrane. Finally, AQP2-bearing vesicles fuse with the apical plasma membrane in response to vasopressin, but not to a significant degree in the absence of stimulation (e.g., in vasopressin-deficient Brattleboro rats where < 5% of total AQP2 is present in the apical plasma membrane. Thus there must be some kind of a “clamp” preventing fusion in the unstimulated state and/or a “trigger” when activation occurs.
The coordinated delivery of AQP2-bearing vesicles to the apical part of the cell appears to depend on the translocation of the vesicles along the cytoskeletal elements. In particular, the microtubular network has been implicated in this process, since chemical disruption of microtubules inhibits the increase in permeability both in the toad bladder and in the mammalian collecting duct. Because microtubule-disruptive agents inhibit the development of the hydrosmotic response to vaso-pressin, but have no effect on the maintenance of an established response, and because they have been reported to slow the development of the response without affecting the final permeability in toad bladders , it has been deduced that microtubules appear to be involved in the coordinated delivery of water channels, without being involved in the actual insertion process.
In addition to increasing cAMP levels in collecting duct principal cells, vasopressin acting through the V2 receptor has also been demonstrated to transiently increase intracellular Ca2+. The increase occurs in the absence of activation of the phosphoinositide signaling pathway and has recently been demonstrated to be due to activation of ryanodine-sensitive calcium release channels in the collecting duct cells. Buffering intracellular calcium with BAPTA or inhibition of calmodulin completely blocked the water permeability response to vasopressin in isolated perfused inner medullary collecting ducts, suggesting a critical role for calcium at some step in the process of AQP2 vesicle trafficking.
In addition to the acute regulation of collecting duct water permeability brought about by the trafficking of AQP2 described above, it is now clear that there are longer term adaptational changes that modulate this acute response. These occur during prolonged changes in body hydration status and form an appropriate physiological response to such challenges. However, similar long term changes also appear to be important in a wide variety of pathological conditions, and an understanding of the mechanisms involved in these adaptational responses may provide the basis both for a better understanding of, and for potential therapeutic approaches to, pathological disorders of water balance. Microtubules are polar structures, arising from microtubule organizing centers (MTOCs), at which their minus ends are anchored, and with the plus ends growing away “into” the cell. In fibroblastic cells, there is a single MTOC in the perinuclear region, and the plus ends project to the periphery of the cell. However, there is increasing evidence that in polarized epithelia microtubules arise from multiple MTOCs in the apical region, with their plus ends projecting down toward the basolateral membrane. If this is the case in collecting duct cells, and there is some evidence that it is , then a minus end-directed motor protein such as dynein would be expected to be involved in the movement of vesicles toward the apical plasma membrane. Recently, it has been shown that dynein is present in the kidney of several mammalian species and that both dynein and dynactin, a protein complex believed to mediate the interaction of dynein with vesicles, associate with AQP2-bearing vesicles. It seems likely that dynein may drive the microtubule-dependent delivery of AQP2-bearing vesicles toward the apical plasma membrane.
The apical part of the collecting duct principal cells contains a prominent terminal web made up of actin filaments. These also appear to be involved in the hydrosmotic response, since disruption of microfilaments with cytochalasins inhibits the response in the toad bladder. Cytochalasins can also inhibit an established response, and even the offset of the response. From this it has been concluded that microfilaments are probably involved in the final movement of vesicles through the terminal web, their fusion with the plasma membrane, and the subsequent endocytic retrieval of the water channels. Interestingly, vasopressin itself causes actin depolymerization, suggesting that reorganization of the terminal web is an important part of the cellular response to vasopressin, a conclusion reached on morphological grounds by DiBona.
The problem of delivering vesicles to a particular domain and allowing them to fuse when, and only when, a signal arrives is conceptually very similar to the situation in the neuronal synapse. It therefore seemed possible that a molecular apparatus similar to the SNAP/SNARE system described there might be present in the collecting duct principal cells. There are specific proteins on the vesicles (vSNAREs) and the target plasma membrane (tSNAREs) that interact with components of a fusion complex to induce fusion of the vesicles only with the required target membrane. The process is thought to be regulated by other protein components that sense the signal for fusion (i.e., increased calcium in the synapse). Several groups have now shown that vSNAREs such as VAMP-2 are present in the collecting duct principal cells and colocalize with AQP2 in the same vesicles .
A putative tSNARE, SNAP23, has been found in collecting duct principal cells both in the apical plasma membrane and in AQP2-bearing vesicles. Some soluble components of the fusion complex, including NEM-sensitive factor (NSF) and a-soluble NSF-associated protein (SNAP), have also been identified in these cells. Thus it seems likely that the exocytic insertion of AQP2 is indeed controlled by a set of proteins similar to those involved in synaptic transmission, although considerable work remains to be done in isolating and characterizing the components, their regulation, and prime physiological function.
Body water balance is tightly regulated by vasopressin, and multiple studies now have underscored the essential roles of AQP2 in this.
Vasopressin regulates acutely the water permeability of the kidney collecting duct by trafficking of AQP2 from intracellular vesicles to the apical plasma membrane.
The long-term adaptational changes in body water balance are controlled in part by regulated changes in AQP2 and AQP3 expression levels. Lack of functional AQP2 is seen in primary forms of diabetes insipidus, and reduced expression and targeting are seen in several diseases associated with urinary concentrating defects such as acquired nephrogenic diabetes insipidus, postobstructive polyuria, as well as acute and chronic renal failure. In contrast, in conditions with water retention such as severe congestive heart failure, pregnancy, and syndrome of inappropriate antidiuretic hormone secretion, both AQP2 expression levels and apical plasma membrane targetting are increased, suggesting a role for AQP2 in the development of water retention. Continued analysis of the aquaporins is providing detailed molecular insight into the fundamental physiology and pathophysiology of water balance and water balance disorders.
Three additional aquaporins are present in the kidney. AQP6 is present in intracellular vesicles in collecting duct intercalated cells, and AQP8 is present intracellularly at low abundance in proximal tubules and collecting duct principal cells, but the physiological function of these two channels remains undefined. AQP7 is abundant in the brush border of proximal tubule cells and is likely to be involved in proximal tubule water reabsorption.
Fluid transport across leaky epithelia: central role of the tight junction and supporting role of aquaporins.
Fischbarg J.
Institute of Cardiology Research , A. C. Taquini, University of Buenos Aires and National Council for Scientific and Technical Investigations, Buenos Aires, Argentina. jf20@columbia.edu
The mechanism of epithelial fluid transport remains unsolved, which is partly due to inherent experimental difficulties. However, a preparation with which our laboratory works, the corneal endothelium, is a simple leaky secretory epithelium in which we have made some experimental and theoretical headway. As we have reported, transendothelial fluid movements can be generated by electrical currents as long as there is tight junction integrity. The direction of the fluid movement can be reversed by current reversal or by changing junctional electrical charges by polylysine. Residual endothelial fluid transport persists even when no anions (hence no salt) are being transported by the tissue and is only eliminated when all local recirculating electrical currents are. The notion that transepithelial movement of water depends on the movement of electrolytes arises from a finding by Peter Curran and Arthur K. Solomon that transintestinal water flow (“solvent” flow) depended on the transport of NaCl (“solute” flux) by that layer. That gave birth to the question of how the flow of solute (or “salt”) is linked to the movement of solvent (or “fluid”), or in the short jargon of the field, how solute-solvent coupling arises.
To be noted, gradientless flow is different from transepithelial osmosis a` la Dutrochet. In this last one, in the presence of an osmotic gradient across an epithelial layer, water obligingly traverses the layer. This is well exemplified by the kidney collecting duct, a tight epithelium for which we accept nowadays that the water goes across both cell plasma membranes in series, traversing their aquaporins. There is also the special case of the anuran skin epithelia, whose intercellular junctions are tight, and which water also appears to traverse through cell membrane aquaporins. As a rule, epithelia specialized to transport fluid do so in the absence of any external osmotic gradient across their layers; that is, fluid is transported between compartments of similar osmolarity. That gave birth to the question of how the flow of solute (or “salt”) is linked to the movement of solvent (or “fluid”), or in the short jargon of the field, how solute-solvent coupling arises.
The progression of the ideas on fluid transport is linked to those in a parallel field, that of water channels. After early advances in their characterization and isolation, they were molecularly identified by Peter Agre and co-workers in the early 1990s, who termed them aquaporins (AQPs). It was subsequently determined that AQPs were present in many fluid transporting epithelia and were also present in water-permeable kidney segments while absent in relatively water-impermeable ones . By then, the measurements of osmotic permeabilities of epithelial cell membranes had been refined using video microscopy techniques. The laboratories of Kenneth Spring (working on gallbladders) and of the Welling brothers (working on kidney proximal tubule) found rather high osmotic permeability (or “filtration” permeability, Pf) values (Persson and Spring: 550 and 1,200 pm/s for the apical and baso-lateral membranes, respectively; Welling: -300 pm/s). Both laboratories suggested that, given such high Pf values, a few milliosmoles of osmotic pressure difference across the cell boundaries would suffice to drive the transported fluids through the cells.
There had been all along experimental evidence for the diverging view that fluid transport across leaky epithelia took place via paracellular, transjunctional water flow. That contrary evidence came from the laboratories of Adrian Hill using gallbladder, John Pappenheimer and his fellow James Madara using intestine, and Guillermo Whittembury and Gerhard Malnic using kidney proximal tubule. The contrary view of paracellular flow had remained a minority opinion. Still and all, these “rebels” stood their ground, led by an utterly unconvinced Adrian Hill. Considering the divergent views, Kenneth Spring and colleagues decided to take the bull by the horns and use confocal microscopy to look for evidence for or against transjunctional water flow in epithelia.
Paracellular, transjunctional fluid flow in an absorbing epithelium would lead to significant dilution of a paracellular fluorescent marker trapped in the intercellular spaces, which in turn would be detectable by the optical sectioning methods they mastered; all very elegant, for sure.
And so we come to the paper Spring and colleagues published in May of 1998 reporting that they had found no transjunctional water flow in cultured Madin-Darby canine kidney (MDCK) cell layers. Understandably, their statement had a very large impact. And yet, only some months afterwards, this notion had to be revised as it became clear that the preparation they had chosen presumably transported little if any water. By Spring’s own admission in October of the same 1998, “ . . . the fluid transport rate of MDCK cells is only about 1% of that of the renal proximal tubule… ” To spell out the obvious, little or no fluid transport means no transjunctional (or trans-cellular) water flow either, so in perspective, the findings of Spring and colleagues (“absence of junctional flow”) bring no surprise and have no bearing on the issue of the route of fluid flow in general.
After the demise of the 1998 paper above, doubts about local osmosis continued to be fueled. Adrian Hill had been joined in his criticism of it by Thomas Zeuthen and Ernest Wright. In particular, Zeuthen and co-workers had developed an alternative model for transcellular water transfer based on molecular cotransport through transporters. Predictably, Hill’s views were newly sought out. In a thorough review written with his wife and colleague Bruria Shachar-Hill, they restated the evidence from theirs and collaborating laboratories for junctional flow for Necturus and rabbit gallbladder, Necturus intestine, Rhodnius Malpighian tubule, and rat and rabbit salivary gland. In addition, they gave a convincing account of the evidence consistent with junctional water flow for renal proximal tubule, exocrine gland (salivary, lacrimal), and small intestine. Here we will simply call attention to those arguments and will concentrate on other arguments plus additional evidence of our own.
By the end of the 1990s, Alan Verkman’s laboratory had been investigating the physiological effects of knocking out AQPs in mice. The deletion of AQPs resulted in drastic decreases of cell membrane osmotic permeability, but only in rather mild decreases in rates of fluid transport, and this last to boot only in tissues that transported fluid at high rates. Verkman and colleagues generally discuss those results in a guarded manner, underlining the role of aquaporins as routes for cell water permeability without making pronouncements on the mechanism of transtissue fluid transport. Yet, paraphrasing the comments by Hill and colleagues in another cogent review, the effects seen in the AQP knockouts are sometimes difficult to explain, and not commensurate with the deletion of what would be hypothetically a major route for transcellular transtissue water transfer.
Perhaps the existence and the location of electrogenic transporters and channels are telling us something very fundamental about the function of these layers. There does not seem to be an explanation of why epithelia in general, and specifically leaky epithelia, would have evolved to have an electrical potential difference across the layer. In principle, salts could simply be transported neutrally. In a similar vein, apical Na channels that allow Na to leak back into the cell would not make sense if the task of an epithelial cell would be to transport salt from the serosal (basal) to the luminal (apical) side. However, both of these apparent incongruencies suddenly make sense if the raison d’être of these epithelia is to perform tasks such as electro-osmosis. The electrical potential might not be an evolutionary leftover but a central feature. The Na channel would not be apical by accident but to help build up the local current meant for electro-osmosis. As mentioned above, aside from the corneal endothelium , there is evidence for electro-osmosis in small intestine, kidney proximal tubule, and frog skin glands. Hence, it would be desirable if the presence of electro-osmosis would be explored in other fluid-transporting epithelia.
Electro-osmotic coupling would result in somewhat (perhaps 30%) hypotonic emerging fluid. This entails that the fluid left behind at the intercellular spaces might be correspondingly hypertonic. Such osmolarity difference in turn might be sensed by the cell and trigger mechanisms that would affect sites for regulation at basolateral and apical sites for HCO3 and Na transports, and perhaps also at the junction so as to modify the characteristics of the coupling. It is conceivable that such regulation might take place with some degree of periodicity. There may be a role for AQP1 in this regulation, which would explain the mild effects seen on fluid transport in this and other preparations in experiments done with AQP1 null cells. This would explain what has been noted by Verkman and colleagues, namely, that effects of AQP deletion are more pronounced in epithelia that generate higher rates of fluid transport. Thus AQP deletion reduced near-isosmolar fluid transport in kidney proximal tubule and salivary gland, where fluid transport is rapid, but not in lung, lacrimal gland, sweat gland, or corneal endothelium where fluid transport is relatively slow.
Aquaporin (AQP) 1 is the only AQP present in these cells, and its deletion in AQP1 null mice significantly affects cell osmotic permeability (by > 40%) but fluid transport much less ( > 20%), which militates against the presence of sizable water movements across the cell. In contrast, AQP1 null mice cells have reduced regulatory volume decrease (only 60% of control), which suggests a possible involvement of AQP1 in either the function or the expression of volume-sensitive membrane channels/transporters. A mathematical model of corneal endothelium we have developed correctly predicts experimental results only when paracellular electro-osmosis is assumed rather than transcellular local osmosis. Our evidence therefore suggests that the fluid is transported across this layer via the paracellular route by a mechanism that we attribute to electro-osmotic coupling at the junctions. From our findings we have developed a novel paradigm for this preparation that includes
1) paracellular fluid flow;
2) a crucial role for the junctions;
3) hypotonicity of the primary secretion; and
4) an AQP role in regulation rather than as a significant water pathway.
These elements are remarkably similar to those proposed by the laboratory of Adrian Hill for fluid transport across other leaky epithelia.
Related articles in Pharmaceutical Intelligence:
Part I: Identification of Biomarkers that are Related to the Actin Cytoskeleton
Larry H Bernstein, MD, FCAP
http://pharmaceuticalintelligence.com/2012/12/10/identification-of-biomarkers-that-are-related-to-the-actin-cytoskeleton/
Part II: Role of Calcium, the Actin Skeleton, and Lipid Structures in Signaling and Cell Motility
Larry H. Bernstein, MD, FCAP, Stephen Williams, PhD and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/08/26/role-of-calcium-the-actin-skeleton-and-lipid-structures-in-signaling-and-cell-motility/
Part III: Renal Distal Tubular Ca2+ Exchange Mechanism in Health and Disease
Larry H. Bernstein, MD, FCAP, Stephen J. Williams, PhD
and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/09/02/renal-distal-tubular-ca2-exchange-mechanism-in-health-and-disease/
Part IV: The Centrality of Ca(2+) Signaling and Cytoskeleton Involving Calmodulin Kinases and Ryanodine Receptors in Cardiac Failure, Arterial Smooth Muscle, Post-ischemic Arrhythmia, Similarities and Differences, and Pharmaceutical Targets
Larry H Bernstein, MD, FCAP, Justin Pearlman, MD, PhD, FACC and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/09/08/the-centrality-of-ca2-signaling-and-cytoskeleton-involving-calmodulin-kinases-and-ryanodine-receptors-in-cardiac-failure-arterial-smooth-muscle-post-ischemic-arrhythmia-similarities-and-differen/
Part V: Heart, Vascular Smooth Muscle, Excitation-Contraction Coupling (E-CC), Cytoskeleton, Cellular Dynamics and Ca2 Signaling
Larry H Bernstein, MD, FCAP, Justin Pearlman, MD, PhD, FACC and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/08/26/heart-smooth-muscle-excitation-contraction-coupling-cytoskeleton-cellular-dynamics-and-ca2-signaling/
Part VI: Calcium Cycling (ATPase Pump) in Cardiac Gene Therapy: Inhalable Gene Therapy for Pulmonary Arterial Hypertension and Percutaneous Intra-coronary Artery Infusion for Heart Failure: Contributions by Roger J. Hajjar, MD
Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/08/01/calcium-molecule-in-cardiac-gene-therapy-inhalable-gene-therapy-for-pulmonary-arterial-hypertension-and-percutaneous-intra-coronary-artery-infusion-for-heart-failure-contributions-by-roger-j-hajjar/
Part VII: Cardiac Contractility & Myocardium Performance: Ventricular Arrhythmiasand Non-ischemic Heart Failure – Therapeutic Implications for Cardiomyocyte Ryanopathy (Calcium Release-related Contractile Dysfunction) and Catecholamine Responses
Justin Pearlman, MD, PhD, FACC, Larry H Bernstein, MD, FCAP and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/08/28/cardiac-contractility-myocardium-performance-ventricular-arrhythmias-and-non-ischemic-heart-failure-therapeutic-implications-for-cardiomyocyte-ryanopathy-calcium-release-related-contractile/
Part VIII: Disruption of Calcium Homeostasis: Cardiomyocytes and Vascular Smooth Muscle Cells: The Cardiac and Cardiovascular Calcium Signaling Mechanism
Justin Pearlman, MD, PhD, FACC, Larry H Bernstein, MD, FCAP and Aviva Lev-Ari, PhD, RN
http://pharmaceuticalintelligence.com/2013/09/12/disruption-of-calcium-homeostasis-cardiomyocytes-and-vascular-smooth-muscle-cells-the-cardiac-and-cardiovascular-calcium-signaling-mechanism/
Part IX: Calcium-Channel Blockers, Calcium Release-related Contractile Dysfunction (Ryanopathy) and Calcium as Neurotransmitter Sensor
Justin Pearlman, MD, PhD, FACC, Larry H Bernstein, MD, FCAP and Aviva Lev-Ari, PhD, RN
Part X: Synaptotagmin functions as a Calcium Sensor: How Calcium Ions Regulate the fusion of vesicles with cell membranes during Neurotransmission
Larry H Bernstein, MD, FCAP and Aviva Lev-Ari, PhD, RN











Read Full Post »






























































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}