Integrins, Cadherins, Signaling and the Cytoskeleton
Curator: Larry H. Bernstein, MD, FCAP
We have reviewed the cytoskeleton, cytoskeleton pores and ionic translocation under lipids. We shall now look at this again, with specific attention to proteins, transporters and signaling.
Integrins and extracellular matrix in mechanotransduction
Lindsay Ramage
Queen’s Medical Research Institute, University of Edinburgh,
Edinburgh, UK
Cell Health and Cytoskeleton 2012; 4: 1–9
Integrins are a family of cell surface receptors which
- mediate cell–matrix and cell–cell adhesions.
Among other functions they provide an important
- mechanical link between the cells external and intracellular environments while
- the adhesions that they form also have critical roles in cellular signal-transduction.
Cell–matrix contacts occur at zones in the cell surface where
- adhesion receptors cluster and when activated
- the receptors bind to ligands in the extracellular matrix.
The extracellular matrix surrounds the cells of tissues and forms the
- structural support of tissue which is particularly important in connective tissues.
Cells attach to the extracellular matrix through
- specific cell-surface receptors and molecules
- including integrins and transmembrane proteoglycans.
Integrins work alongside other proteins such as
- cadherins,
- immunoglobulin superfamily
- cell adhesion molecules,
- selectins, and
- syndecans
to mediate
- cell–cell and
- cell–matrix interactions and communication.
Activation of adhesion receptors triggers the formation of matrix contacts in which
- bound matrix components,
- adhesion receptors,
- and associated intracellular cytoskeletal and signaling molecules
form large functional, localized multiprotein complexes.
Cell–matrix contacts are important in a variety of different cell and
tissue properties including
- embryonic development,
- inflammatory responses,
- wound healing,
- and adult tissue homeostasis.
This review summarizes the roles and functions of integrins and extracellular matrix proteins in mechanotransduction.
Integrins are a family of αβ heterodimeric receptors which act as
- cell adhesion molecules
- connecting the ECM to the actin cytoskeleton.
The actin cytoskeleton is involved in the regulation of
- cell motility,
- cell polarity,
- cell growth, and
- cell survival.
The integrin family consists of around 25 members which are composed of differing
- combinations of α and β subunits.
The combination of αβ subunits determines
- binding specificity and
- signaling properties.
In mammals around 19 α and eight β subunits have been characterized.
Both α and β integrin subunits contain two separate tails, which
- penetrate the plasma membrane and possess small cytoplasmic domains which facilitate
- the signaling functions of the receptor.
There is some evidence that the β subunit is the principal
site for
- binding of cytoskeletal and signaling molecules,
whereas the α subunit has a regulatory role. The integrin
tails
- link the ECM to the actin cytoskeleton within the cell and with cytoplasmic proteins,
such as talin, tensin, and filamin. The extracellular domains of integrin receptors bind the ECM ligands.
The ECM is a complex mixture of matrix molecules, including -glycoproteins, collagens, laminins, glycosaminoglycans, proteoglycans,
and nonmatrix proteins, – including growth factors.
These can be categorized as insoluble molecules within the ECM, soluble molecules, and/or matrix-associated biochemicals, such as systemic hormones or growth factors and cytokines that act locally.
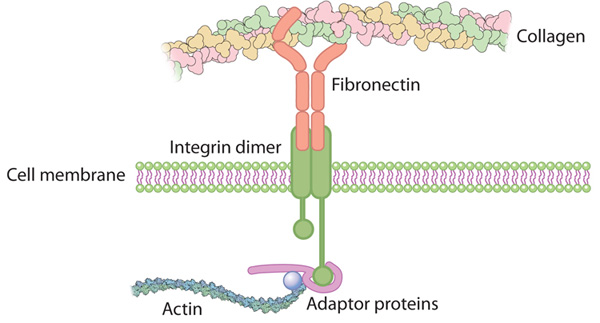
The integrin receptor formed from the binding of α and β subunits is shaped like a globular head supported by two rod-like legs (Figure 1). Most of the contact between the two subunits occurs in the head region, with the intracellular tails of the subunits forming the legs of the receptor.6 Integrin recognition of ligands is not constitutive but is regulated by alteration of integrin affinity for ligand binding. For integrin binding to ligands to occur the integrin must be primed and activated, both of which involve conformational changes to the receptor.
The integrins are composed of well-defined domains used for protein–protein interactions. The α-I domains of α integrin subunits comprise the ligand binding sites. X-ray crystallography has identified an α-I domain within the β subunit and a β propeller domain within the α subunit which complex to form the ligand-binding head of the integrin.
The use of activating and conformation-specific antibodies also suggests that the β chain is extended in the active integrin. It has since been identified that the hybrid domain in the β chain is critical for integrin activation, and a swing-out movement of this leg activates integrins.
http://www.ks.uiuc.edu/Publications/Stories/tcbg_ytt/pdfs/dbp6.pdf
DBP6: Integrin

Integrin

Integrin.large
Linking integrin conformation to function
Figure Integrin binding to extracellular matrix (ECM). Conformational changes to integrin structure and clustering of subunits which allow enhanced function of the receptor.

integrin coupled to F-actin via linker
http://dx.dio.org:/integrin-coupled-to-f-actin-via-linker-nrm3896-f4.jpg
Integrin extracellular binding activity is regulated from inside the cell and binding to the ECM induces signals that are transmitted into the cell.15 This bidirectional signaling requires
- dynamic,
- spatially, and
- temporally regulated formation and
- disassembly of multiprotein complexes that
form around the short cytoplasmic tails of integrins.
Ligand binding to integrin family members leads to clustering of integrin molecules in the plasma membrane and recruitment of actin filaments and intracellular signaling molecules to the cytoplasmic domain of the integrins. This forms focal adhesion complexes which are able to maintain
- not only adhesion to the ECM
- but are involved in complex signaling pathways
which include establishing
- cell polarity,
- directed cell migration, and
- maintaining cell growth and survival.
Initial activation through integrin adhesion to matrix recruits up to around 50 diverse signaling molecules
- to assemble the focal adhesion complex
- which is capable of responding to environmental stimuli efficiently.
Mapping of the integrin
- adhesome binding and signaling interactions
identified a network of 156 components linked together which can be modified by 690 interactions.
The binding of the adaptor protein talin to the β subunit cytoplasmic tail is known to have a key role in integrin activation. This is thought to occur through the disruption of
- inhibitory interactions between α and β subunit cytoplasmic tails.
Talin also binds
- to actin and to cytoskeletal and signaling proteins.
This allows talin to directly link activated integrins
to signaling events and the cytoskeleton.
Genetic programming occurs with the binding of integrins to the ECM
Signal transduction pathway activation arising from integrin-
ECM binding results in changes in gene expression of cells
and leads to alterations in cell and tissue function. Various
different effects can arise depending on the
- cell type,
- matrix composition, and
- integrins activated.
One way in which integrin expression is important in genetic programming is in the fate and differentiation of stem cells.
Osteoblast differentiation occurs through ECM interactions
with specific integrins
- to initiate intracellular signaling pathways leading to osteoblast-specific gene expression
- disruption of interactions between integrins and collagen;
- fibronectin blocks osteoblast differentiation and
Disruption of α2 integrin prevents osteoblast differentiation, and activation of the transcription factor
- osteoblast-specific factor 2/core-binding factor α1.
It was found that the ECM-integrin interaction induces osteoblast-specific factor 2/core-binding factor α1 to
- increase its activity as a transcriptional enhancer
- rather than increasing protein levels.
It was also found that modification of α2 integrin alters
- induction of the osteocalcin promoter;
- inhibition of α2 prevents activation of the osteocalcin promoter,
- overexpression enhanced osteocalcin promoter activity.
It has been suggested that integrin-type I collagen interaction is necessary for the phosphorylation and activation of osteoblast-specific transcription factors present in committed osteoprogenitor cells.
A variety of growth factors and cytokines have been shown to be important in the regulation of integrin expression and function in chondrocytes. Mechanotransduction in chondrocytes occurs through several different receptors and ion channels including integrins. During osteoarthritis the expression of integrins by chondrocytes is altered, resulting in different cellular transduction pathways which contribute to tissue pathology.
In normal adult cartilage, chondrocytes express α1β1, α10β1 (collagen receptors), α5β1, and αvβ5 (fibronectin) receptors. During mechanical loading/stimulation of chondrocytes there is an influx of ions across the cell membrane resulting from activation of mechanosensitive ion channels which can be inhibited by subunit-specific anti-integrin blocking antibodies or RGD peptides. Using these strategies it was identified that α5β1 integrin is a major mechanoreceptor in articular chondrocyte responses to mechanical loading/stimulation.
Osteoarthritic chondrocytes show a depolarization response to 0.33 Hz stimulation in contrast to the hyperpolarization response of normal chondrocytes. The mechanotransduction pathway in chondrocytes derived from normal and osteoarthritic cartilage both involve recognition of the mechanical stimulus by integrin receptors resulting in the activation of integrin signaling pathways leading to the generation of a cytokine loop. Normal and osteoarthritic chondrocytes show differences at multiple stages of the mechanotransduction cascade (Figure 3). Early events are similar; α5β1 integrin and stretch activated ion channels are activated and result in rapid tyrosine phosphorylation events. The actin cytoskeleton is required for the integrin-dependent Mechanotransduction leading to changes in membrane potential in normal but not osteoarthritic chondrocytes.
Cell–matrix interactions are essential for maintaining the integrity of tissues. An intact matrix is essential for cell survival and proliferation and to allow efficient mechanotransduction and tissue homeostasis. Cell–matrix interactions have been extensively studied in many tissues and this knowledge is being used to develop strategies to treat pathology. This is particularly important in tissues subject to abnormal mechanical loading, such as musculoskeletal tissues. Integrin-ECM interactions are being used to enhance tissue repair mechanisms in these tissues through differentiation of progenitor cells for in vitro and in vivo use. Knowledge of how signaling cascades are differentially regulated in response to physiological and pathological external stimuli (including ECM availability and mechanical loading/stimulation) will enable future strategies to be developed to prevent and treat the progression of pathology associated with integrin-ECM interactions.
Cellular adaptation to mechanical stress: role of integrins, Rho, cytoskeletal tension and mechanosensitive ion channels
- Matthews, DR. Overby, R Mannix and DE. Ingber
1Vascular Biology Program, Departments of Pathology and Surgery, Children’s Hospital, and 2Department of Pediatrics, Massachusetts General Hospital, Harvard Medical School, Boston, MA J Cell Sci 2006; 119: 508-518. http://dx.doi.org:/10.1242/jcs.02760
To understand how cells sense and adapt to mechanical stress, we applied tensional forces to magnetic microbeads bound to cell-surface integrin receptors and measured changes in bead isplacement with sub-micrometer resolution using optical microscopy. Cells exhibited four types of mechanical responses: (1) an immediate viscoelastic response;
(2) early adaptive behavior characterized by pulse-to-pulse attenuation in response to oscillatory forces;
(3) later adaptive cell stiffening with sustained (>15 second) static stresses; and
(4) a large-scale repositioning response with prolonged (>1 minute) stress.
Importantly, these adaptation responses differed biochemically. The immediate and early responses were affected by
- chemically dissipating cytoskeletal prestress (isometric tension), whereas
- the later adaptive response was not.
The repositioning response was prevented by
- inhibiting tension through interference with Rho signaling,
similar to the case of the immediate and early responses, but it was also prevented by
- blocking mechanosensitive ion channels or
- by inhibiting Src tyrosine kinases.
All adaptive responses were suppressed by cooling cells to 4°C to slow biochemical remodeling. Thus, cells use multiple mechanisms to sense and respond to static and dynamic changes in the level of mechanical stress applied to integrins.
Microtubule-Stimulated ADP Release, ATP Binding, and Force Generation In Transport Kinesins
J Atherton, I Farabella, I-Mei Yu, SS Rosenfeld, A Houdusse, M Topf, CA Moores
1Institute of Structural and Molecular Biology, Department of Biological Sciences, Birkbeck College, University of London, London, United Kingdom; 2Structural Motility, Institut Curie, Centre National de la Recherche Scientifique, Paris, France; 3Department of Cancer Biology, Lerner Research Institute, Cleveland Clinic, Cleveland, United States
eLife 2014;3:e03680. http://dx.doi.org:/10.7554/eLife.03680
Kinesins are a large family of microtubule (MT)-based motors that play important roles in many cellular activities including
- mitosis,
- motility, and
- intracellular transport
Their involvement in a range of pathological processes also highlights their significance as therapeutic targets and the importance of understanding the molecular basis of their function They are defined by their motor domains that contain both the microtubule (MT) and ATP binding sites. Three ATP binding motifs—the P-loop, switch I, switch II–are highly conserved among kinesins, myosin motors, and small GTPases. They share a conserved mode of MT binding such that MT binding, ATP binding, and hydrolysis are functionally coupled for efficient MT-based work.
The interior of a cell is a hive of activity, filled with proteins and other items moving from one location to another. A network of filaments called microtubules forms tracks along which so-called motor proteins carry these items. Kinesins are one group of motor proteins, and a typical kinesin protein has one end (called the ‘motor domain’) that can attach itself to the microtubules.
The other end links to the cargo being carried, and a ‘neck’ connects the two. When two of these proteins work together, flexible regions of the neck allow the two motor domains to move past one another, which enable the kinesin to essentially walk along a microtubule in a stepwise manner.
Atherton et al. use a technique called cryo-electron microscopy to study—in more detail than previously seen—the structure of the motor domains of two types of kinesin called kinesin-1 and kinesin-3. Images were taken at different stages of the cycle used by the motor domains to extract the energy from ATP molecules. Although the two kinesins have been thought to move along the microtubule tracks in different ways, Atherton et al. find that the core mechanism used by their motor domains is the same.
When a motor domain binds to the microtubule, its shape changes, first stimulating release of the breakdown products of ATP from the previous cycle. This release makes room for a new ATP molecule to bind. The structural changes caused by ATP binding are relatively small but produce larger changes in the flexible neck region that enable individual motor domains within a kinesin pair to co-ordinate their movement and move in a consistent direction. This mechanism involves tight coupling between track binding and fuel usage and makes kinesins highly efficient motors.
A number of kinesins drive long distance transport of cellular cargo with dimerisation allowing them to take multiple 8 nm ATP-driven steps toward MT plus ends. Their processivity depends on communication between the two motor domains, which is achieved via the neck linker that connects each motor domain to the dimer-forming coiled-coil
Kinesins are a superfamily of microtubule-based
- ATP-powered motors, important for multiple, essential cellular functions.
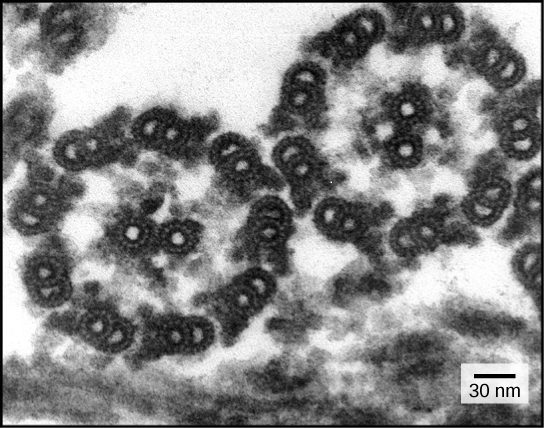
How microtubule binding stimulates their ATPase and controls force generation is not understood. To address this fundamental question, we visualized microtubule-bound kinesin-1 and kinesin-3 motor domains at multiple steps in their ATPase cycles—including their nucleotide-free states—at ∼7 Å resolution using cryo-electron microscopy.
All our reconstructions have, as their asymmetric unit, a triangle-shaped motor domain bound to an αβ-tubulin dimer within the MT lattice (Figure 1). The structural comparisons below are made with respect to the MT surface, which, at the resolution of our structures (∼7 Å, Table 1), is the same (CCC > 0.98 for all). As is well established across the superfamily, the major and largely invariant point of contact between kinesin motor domains and the MT is helix-α4, which lies at the tubulin intradimer interface (Figure 1C, Kikkawa et al., 2001).
However, multiple conformational changes are seen throughout the rest of each domain in response to bound nucleotide (Figure 1D). Below, we describe the conformational changes in functionally important regions of each motor domain starting with the nucleotide-binding site, from which all other conformational changes emanate.
The nucleotide-binding site (Figure 2) has three major elements: (1) the P-loop (brown) is visible in all our reconstructions;
(2) loop9 (yellow, contains switch I) undergoes major conformational changes through the ATPase cycle; and
(3) loop11 (red, contains switch II) that connects strand-β7 to helix-α4,
the conformation and flexibility of which is determined by MT binding and motor nucleotide state.
Movement and extension of helix-α6 controls neck linker docking
the N-terminus of helix-α6 is closely associated with elements of the nucleotide binding site suggesting that its conformation alters in response to different nucleotide states. In addition, because the orientation of helix-α6 with respect to helix-α4 controls neck linker docking and because helix-α4 is held against the MT during the ATPase cycle,
- conformational changes in helix-α6 control movement of the neck linker.
Mechanical amplification and force generation involves conformational changes across the motor domain
A key conformational change in the motor domain following Mg-ATP binding is peeling of the central β-sheet from the C-terminus of helix-α4 increasing their separation (Figure 3—figure supplement 2); this is required to accommodate rotation of helix-α6 and consequent neck linker docking (Figure 3B–E).
Peeling of the central β-sheet has previously been proposed to arise from tilting of the entire motor domain relative to static MT contacts, pivoting around helix-α4 (the so-called ‘seesaw’ model; Sindelar, 2011). Specifically, this model predicts that the major difference in the motor before and after Mg-ATP binding would be the orientation of the motor domain with respect to helix-α4.
Kinesin mechanochemistry and the extent of mechanistic conservation within the motor superfamily are open questions, critical to explain how MT binding, and ATP binding and hydrolysis drive motor activity. Our structural characterisation of two transport motors now allows us to propose a model that describes the roles of mechanochemical elements that together drive conserved MT-based motor function.
Model of conserved MT-bound kinesin mechanochemistry. Loop11/N-terminus of helix-α4 is flexible in ADP-bound kinesin in solution, the neck linker is also flexible while loop9 chelates ADP. MT binding is sensed by loop11/helix-α4 N-terminus, biasing them towards more ordered conformations.
We propose that this favours crosstalk between loop11 and loop9, stimulating ADP release. In the NN conformation, both loop11 and loop9 are well ordered and primed to favour ATP binding, while helix-α6—which is required for mechanical amplification–is closely associated with the MT on the other side of the motor domain. ATP binding draws loop11 and loop9 closer together; causing
(1) tilting of most of the motor domain not contacting the MT towards the nucleotide-binding site,
(2) rotation, translation, and extension of helix-α6 which we propose contributes to force generation, and
(3) allows neck linker docking and biases movement of the 2nd head towards the MT plus end.
In both motors, microtubule binding promotes
- ordered conformations of conserved loops that
- stimulate ADP release,
- enhance microtubule affinity and
- prime the catalytic site for ATP binding.
ATP binding causes only small shifts of these nucleotide-coordinating loops but induces
- large conformational changes elsewhere that
- allow force generation and
- neck linker docking towards the microtubule plus end.
Family-specific differences across the kinesin–microtubule interface account for the
- distinctive properties of each motor.
Our data thus provide evidence for a
conserved ATP-driven
- mechanism for kinesins and
- reveal the critical mechanistic contribution of the microtubule interface.
Phosphorylation at endothelial cell–cell junctions: Implications for VE-cadherin function
I Timmerman, PL Hordijk, JD van Buul
Cell Health and Cytoskeleton 2010; 2: 23–31
Endothelial cell–cell junctions are strictly regulated in order to
- control the barrier function of endothelium.
Vascular endothelial (VE)-cadherin is one of the proteins that is crucial in this process. It has been reported that
- phosphorylation events control the function of VE-cadherin.
This review summarizes the role of VE-cadherin phosphorylation in the regulation of endothelial cell–cell junctions and highlights how this affects vascular permeability and leukocyte extravasation.
The vascular endothelium is the inner lining of blood vessels and
- forms a physical barrier between the vessel lumen and surrounding tissue;
- controlling the extravasation of fluids,
- plasma proteins and leukocytes.
Changes in the permeability of the endothelium are tightly regulated. Under basal physiological conditions, there is a continuous transfer of substances across the capillary beds. In addition the endothelium can mediate inducible,
- transient hyperpermeability
- in response to stimulation with inflammatory mediators,
- which takes place primarily in postcapillary venules.
However, when severe, inflammation may result in dysfunction of the endothelial barrier in various parts of the vascular tree, including large veins, arterioles and capillaries. Dysregulated permeability is observed in various pathological conditions, such as tumor-induced angiogenesis, cerebrovascular accident and atherosclerosis.
Two fundamentally different pathways regulate endothelial permeability,
- the transcellular and paracellular pathways.
Solutes and cells can pass through the body of endothelial cells via the transcellular pathway, which includes
- vesicular transport systems, fenestrae, and biochemical transporters.
The paracellular route is controlled by
- the coordinated opening and closing of endothelial junctions and
- thereby regulates traffic across the intercellular spaces between endothelial cells.
Endothelial cells are connected by
- tight, gap and
- adherens junctions,
of which the latter, and particularly the adherens junction component,
- vascular endothelial (VE)-cadherin,
- are of central importance for the initiation and stabilization of cell–cell contacts.
Although multiple adhesion molecules are localized at endothelial junctions, blocking the adhesive function of VE-cadherin using antibodies is sufficient to disrupt endothelial junctions and to increase endothelial monolayer permeability both in vitro and in vivo. Like other cadherins, VE-cadherin mediates adhesion via homophilic, calcium-dependent interactions.
This cell–cell adhesion
- is strengthened by binding of cytoplasmic proteins, the catenins,
- to the C-terminus of VE-cadherin.
VE-cadherin can directly bind β-catenin and plakoglobin, which
- both associate with the actin binding protein α-catenin.
Initially, α-catenin was thought to directly anchor cadherins to the actin cytoskeleton, but recently it became clear that
- α-catenin cannot bind to both β-catenin and actin simultaneously.
Data using purified proteins show that
- monomeric α-catenin binds strongly to cadherin-bound β-catenin;
- in contrast to the dimer which has a higher affinity for actin filaments,
- indicating that α-catenin might function as a molecular switch regulating cadherin-mediated cell–cell adhesion and actin assembly.
Thus, interactions between the cadherin complex and the actin cytoskeleton are more complex than previously thought. Recently, Takeichi and colleagues reported that
- the actin binding protein EPLIN (epithelial protein lost in neoplasm)
- can associate with α-catenin and thereby
- link the E-cadherin–catenin complex to the actin cytoskeleton.
Although this study was performed in epithelial cells,
- an EPLIN-like molecule might serve as
- a bridge between the cadherin–catenin complex and
- the actin cytoskeleton in endothelial cells.
Next to β-catenin and plakoglobin, p120-catenin also binds directly to the intracellular tail of VE-cadherin.
Numerous lines of evidence indicate that
- p120-catenin promotes VE-cadherin surface expression and stability at the plasma membrane.
Different models are proposed that describe how p120-catenin regulates cadherin membrane dynamics, including the hypothesis
- that p120-catenin functions as a ‘cap’ that prevents the interaction of VE-cadherin
- with the endocytic membrane trafficking machinery.
In addition, p120-catenin might regulate VE-cadherin internalization through interactions with small GTPases. Cytoplasmic p120-catenin, which is not bound to VE-cadherin, has been shown to
- decrease RhoA activity,
- elevate active Rac1 and Cdc42, and thereby is thought
- to regulate actin cytoskeleton organization and membrane trafficking.
The intact cadherin-catenin complex is required for proper functioning of the adherens junction. Mutant forms of VE-cadherin which
- lack either the β-catenin, plakoglobin or p120 binding regions reduce the strength of cell–cell adhesion.
Moreover, our own results showed that
- interfering with the interaction between α-catenin and β-catenin,
- using a cell-permeable peptide which encodes the binding site in α-catenin for β-catenin,
- resulted in an increased permeability of the endothelial monolayer.
Several mechanisms may be involved in the regulation of the organization and function of the cadherin–catenin complex, including endocytosis of the complex, VE-cadherin cleavage and actin cytoskeleton reorganization. The remainder of this review primarily focuses on the
- role of tyrosine phosphorylation in the control of VE-cadherin-mediated cell–cell adhesion.
Regulation of the adhesive function of VE-cadherin by tyrosine phosphorylation
It is a widely accepted concept that tyrosine phosphorylation of components of the VE–cadherin-catenin complex
- Correlates with the weakening of cell–cell adhesion.
One of the first reports that supported this idea showed that the level of phosphorylation of VE-cadherin was
- high in loosely confluent endothelial cells, but
- low in tightly confluent monolayers,
when intercellular junctions are stabilized.
In addition, several conditions that induce tyrosine phosphorylation
of adherens junction components, like
- v-Src transformation
- and inhibition of phosphatase activity by pervanadate,
have been shown to shift cell–cell adhesion from a strong to a weak state. More physiologically relevant;
permeability-increasing agents such as
- histamine,
- tumor necrosis factor-α (TNF-α),
- thrombin,
- platelet-activating factor (PAF) and
- vascular endothelial growth factor (VEGF)
increase tyrosine phosphorylation of various components of the cadherin–catenin complex.
A general idea has emerged that
- tyrosine phosphorylation of the VE-cadherin complex
- leads to the uncoupling of VE-cadherin from the actin cytoskeleton
- through dissociation of catenins from the cadherin.
However, tyrosine phosphorylation of VE-cadherin is required for efficient transmigration of leukocytes.
This suggests that VE-cadherin-mediated cell–cell contacts
- are not just pushed open by the migrating leukocytes, but play
- a more active role in the transmigration process.
A schematic overview of leukocyte adhesion-induced signals leading to VE-cadherin phosphorylation

Regulation of the integrity of endothelial cell–cell contacts by phosphorylation of VE-cadherin
Regulation of the integrity of endothelial cell–cell contacts by phosphorylation of VE-cadherin.
Notes: A) Permeability-inducing agents such as thrombin, histamine and VEGF, induce tyrosine phosphorylation (pY) of VE-cadherin and the associated catenins. Although the specific consequences of catenin tyrosine phosphorylation in endothelial cells are still unknown, VE-cadherin tyrosine phosphorylation results in opening of the cell–cell junctions (indicated by arrows) and enhanced vascular permeability. How tyrosine phosphorylation affects VE-cadherin adhesiveness is not yet well understood; disrupted binding of catenins, which link the cadherin to the actin cytoskeleton, may be involved. VEGF induces phosphorylation of VE-cadherin at specific residues, Y658 and Y731, which have been reported to regulate p120-catenin and β-catenin binding, respectively. Moreover, VEGF stimulation results in serine phosphorylation (pSer) of VE-cadherin, specifically at residue S665, which leads to its endocytosis. B) Adhesion of leukocytes to endothelial cells via ICAM-1 increases endothelial permeability by inducing phosphorylation of VE-cadherin on tyrosine residues. Essential mediators, such as the kinases Pyk2 and Src, and signaling routes involving reactive oxygen species (ROS) and Rho, have been shown to act downstream of ICAM-1. Different tyrosine residues within the cytoplasmic domain of VE-cadherin are involved in the extravasation of neutrophils and lymphocytes, including Y658 and Y731. (β: β-catenin, α: α-catenin, γ: γ-catenin/plakoglobin).
N-glycosylation status of E-cadherin controls cytoskeletal dynamics through the organization of distinct β-catenin- and γ-catenin-containing AJs
BT Jamal, MN Nita-Lazar, Z Gao, B Amin, J Walker, MA Kukuruzinska
Cell Health and Cytoskeleton 2009; 1: 67–80
N-glycosylation of E-cadherin has been shown to inhibit cell–cell adhesion. Specifically, our recent studies have provided evidence that the reduction of E-cadherin N-glycosylation promoted the recruitment of stabilizing components, vinculin and serine/ threonine protein phosphatase 2A (PP2A), to adherens junctions (AJs) and enhanced the association of AJs with the actin cytoskeleton. Here, we examined the details of how
- N-glycosylation of E-cadherin affected the molecular organization of AJs and their cytoskeletal interactions.
Using the hypoglycosylated E-cadherin variant, V13, we show that
- V13/β-catenin complexes preferentially interacted with PP2A and with the microtubule motor protein dynein.
This correlated with dephosphorylation of the microtubule-associated protein tau, suggesting that
- increased association of PP2A with V13-containing AJs promoted their tethering to microtubules.
On the other hand, V13/γ-catenin complexes associated more with vinculin, suggesting that they
- mediated the interaction of AJs with the actin cytoskeleton.
- N-glycosylation driven changes in the molecular organization of AJs were physiologically significant because transfection of V13 into A253 cancer cells, lacking both mature AJs and tight junctions (TJs), promoted the formation of stable AJs and enhanced the function of TJs to a greater extent than wild-type E-cadherin.
These studies provide the first mechanistic insights into how N-glycosylation of E-cadherin drives changes in AJ composition through
- the assembly of distinct β-catenin- and γ-catenin-containing scaffolds that impact the interaction with different cytoskeletal components.
Cytoskeletal Basis of Ion Channel Function in Cardiac Muscle
Matteo Vatta, and Georgine Faulkner,
1 Departments of Pediatrics (Cardiology), Baylor College of Medicine, Houston, TX 2 Department of Reproductive and Developmental Sciences, University of Trieste, Trieste, Italy
3 Muscular Molecular Biology Unit, International Centre for Genetic Engineering and Biotechnology, Padriciano, Trieste, Italy
Future Cardiol. 2006 July 1; 2(4): 467–476. http://dx.doi.org:/10.2217/14796678.2.4.467
The heart is a force-generating organ that responds to
- self-generated electrical stimuli from specialized cardiomyocytes.
This function is modulated
- by sympathetic and parasympathetic activity.
In order to contract and accommodate the repetitive morphological changes induced by the cardiac cycle, cardiomyocytes
- depend on their highly evolved and specialized cytoskeletal apparatus.
Defects in components of the cytoskeleton, in the long term,
- affect the ability of the cell to compensate at both functional and structural levels.
In addition to the structural remodeling,
- the myocardium becomes increasingly susceptible to altered electrical activity leading to arrhythmogenesis.
The development of arrhythmias secondary to structural remodeling defects has been noted, although the detailed molecular mechanisms are still elusive. Here I will review
- the current knowledge of the molecular and functional relationships between the cytoskeleton and ion channels
and, I will discuss the future impact of new data on molecular cardiology research and clinical practice.
Myocardial dysfunction in the end-stage failing heart is very often associated with increasing
- susceptibility to ventricular tachycardia (VT) and ventricular fibrillation (VF),
both of which are common causes of sudden cardiac death (SCD).
Among the various forms of HF,
myocardial remodeling due to ischemic cardiomyopathy (ICM) or dilated cardiomyopathy (DCM)
- is characterized by alterations in baseline ECG,
which includes the
- prolongation of the QT interval,
- as well as QT dispersion,
- ST-segment elevation, and
- T-wave abnormalities,
especially during exercise. In particular, subjects with
severe left ventricular chamber dilation such as in DCM can have left bundle branch block (LBBB), while right bundle branch block (RBBB) is more characteristic of right ventricular failure. LBBB and RBBB have both been repeatedly associated with AV block in heart failure.
The impact of volume overload on structural and electro-cardiographic alterations has been noted in cardiomyopathy patients treated with left ventricular assist device (LVAD) therapy, which puts the heart at mechanical rest. In LVAD-treated subjects,
- QRS- and both QT- and QTc duration decreased,
- suggesting that QRS- and QT-duration are significantly influenced by mechanical load and
- that the shortening of the action potential duration contributes to the improved contractile performance after LVAD support.
Despite the increasing use of LVAD supporting either continuous or pulsatile blood flow in patients with severe HF, the benefit of this treatment in dealing with the risk of arrhythmias is still controversial.
Large epidemiological studies, such as the REMATCH study, demonstrated that the
- employment of LVAD significantly improved survival rate and the quality of life, in comparison to optimal medical management.
An early postoperative period study after cardiac unloading therapy in 17 HF patients showed that in the first two weeks after LVAD implantation,
- HF was associated with a relatively high incidence of ventricular arrhythmias associated with QTc interval prolongation.
In addition, a recent retrospective study of 100 adult patients with advanced HF, treated with an axial-flow HeartMate LVAD suggested that
- the rate of new-onset monomorphic ventricular tachycardia (MVT) was increased in LVAD treated patients compared to patients given only medical treatment,
while no effect was observed on the development of polymorphic ventricular tachycardia (PVT)/ventricular fibrillation (VF).
The sarcomere
The myocardium is exposed to severe and continuous biomechanical stress during each contraction-relaxation cycle. When fiber tension remains uncompensated or simply unbalanced,
- it may represent a trigger for arrhythmogenesis caused by cytoskeletal stretching,
- which ultimately leads to altered ion channel localization, and subsequent action potential and conduction alterations.
Cytoskeletal proteins not only provide the backbone of the cellular structure, but they also
- maintain the shape and flexibility of the different sub-cellular compartments, including the
- plasma membrane,
- the double lipid layer, which defines the boundaries of the cell and where
- ion channels are mainly localized.
The interaction between the sarcomere, which is the basic for the passive force during diastole and for the restoring force during systole. Titin connects
- the Z-line to the M-line of the sarcomeric structure
(Figure 1).
In addition to the strategic
- localization and mechanical spring function,
- titin is a length-dependent sensor during
- stretch and promotes actin-myosin interaction
Titin is stabilized by the cross-linking protein
- telethonin (T-Cap), which localizes at the Z-line and is also part of titin sensor machinery (Figure 1).
The complex protein interactions in the sarcomere entwine telethonin to other
- Z-line components through the family of the telethonin-binding proteins of the Z-disc, FATZ, also known as calsarcin and myozenin.
FATZ binds to
- calcineurin,
- γ-filamin as well as the
- spectrin-like repeats (R3–R4) of α-actinin-2,
the major component of the Z-line and a pivotal
- F-actin cross-linker (Figure 1).contractile unit of striated muscles, and
- the sarcolemma,
the plasma membrane surrendering the muscle fibers in skeletal muscle and the muscle cell of the cardiomyocyte,
- determines the mechanical plasticity of the cell, enabling it to complete and re-initiate each contraction-relaxation cycle.
At the level of the sarcomere,
- actin (thin) and myosin (thick) filaments generate the contractile force,
while other components such as titin, the largest protein known to date, are responsible for
- the passive force during diastole and for the restoring force during systole, and (titin).
- the Z-line to the M-line of the sarcomeric structure
(Figure 1).
In addition to the strategic
- localization and mechanical spring function,
- it acts as a length-dependent sensor during stretch and
- promotes actin-myosin interaction.
Stabilized by the cross-linking protein telethonin (T-Cap),
- titin localizes at the Z-line and is
- part of titin sensor machinery
Another cross-linker of α-actinin-2 in the complex Z-line scaffold is
- the Z-band alternatively spliced PDZ motif protein (ZASP),
- which has an important role in maintaining Z-disc stability
in skeletal and cardiac muscle (Figure 1).
ZASP contains a PDZ motif at its N-terminus,
- which interacts with C-terminus of α-actinin-2,
- and a conserved sequence called the ZASP like motif (ZM)
- found in the alternatively spliced exons 4 and 6.
It has also been reported
- to bind to the FATZ (calsarcin) family of Z-disc proteins (Figure 1).
The complex protein interactions in the sarcomere entwine telethonin to other Z-line components through the family of the telethonin-binding proteins of the
- Z-disc,
- FATZ, also known as calsarcin and
- myozenin
FATZ binds to calcineurin,
- γ-filamin as well as the
- spectrin-like repeats (R3–R4) of α-actinin-2, the major component of the Z-line and a pivotal F-actin cross-linker (Figure 1).

sarcomere structure
Figure 1. Sarcomere structure
The diagram illustrates the sarcomeric structure. The Z-line determines the boundaries of the contractile unit, while Titin connects the Z-line to the M-line and acts as a functional spring during contraction/relaxation cycles.
Sarcomeric Proteins and Ion Channels
In addition to systolic dysfunction characteristic of dilated cardiomyopathy (DCM) and diastolic dysfunction featuring hypertrophic cardiomyopathy (HCM), the clinical phenotype of patients with severe cardiomyopathy is very often associated with a high incidence of cardiac arrhythmias. Therefore, besides fiber stretch associated with mechanical and hemodynamic impairment, cytoskeletal alterations due to primary genetic defects or indirectly to alterations in response to cellular injury can potentially
- affect ion channel anchoring, and trafficking, as well as
- functional regulation by second messenger pathways,
- causing an imbalance in cardiac ionic homeostasis that will trigger arrhythmogenesis.
Intense investigation of
- the sarcomeric actin network,
- the Z-line structure, and
- chaperone molecules docking in the plasma membrane,
has shed new light on the molecular basis of
- cytoskeletal interactions in regulating ion channels.
In 1991, Cantiello et al., demonstrated that
- although the epithelial sodium channel and F-actin are in close proximity,
- they do not co-localize.
Actin disruption using cytochalasin D, an agent that interferes with actin polymerization, increased Na+ channel activity in 90% of excised patches tested within 2 min, which indicated that
- the integrity of the filamentous actin (F-actin) network was essential
- for the maintenance of normal Na+ channel function.
Later, the group of Dr. Jonathan Makielski demonstrated that
- actin disruption induced a dramatic reduction in Na+ peak current and
- slowed current decay without affecting steady-state voltage-dependent availability or recovery from inactivation.
These data were the first to support a role for the cytoskeleton in cardiac arrhythmias.
F-actin is intertwined in a multi-protein complex that includes
- the composite Z-line structure.
Further, there is a direct binding between
- the major protein of the Z-line, α-actinin-2 and
- the voltage-gated K+ channel 1.5 (Kv1.5), (Figure 2).
The latter is expressed in human cardiomyocytes and localizes to
- the intercalated disk of the cardiomyocyte
- in association with connexin and N-cadherin.
Maruoka et al. treated HEK293 cells stably expressing Kv1.5 with cytochalasin D, which led to
- a massive increase in ionic and gating IK+ currents.
This was prevented by pre-incubation with phalloidin, an F-actin stabilizing agent. In addition, the Z-line protein telethonin binds to the cytoplasmic domain of minK, the beta subunit of the potassium channel KCNQ1 (Figure 2).

Molecular interactions between the cytoskeleton and ion channels
Figure 2. Molecular interactions between the cytoskeleton and ion channels
The figure illustrates the interactions between the ion channels on the sarcolemma, and the sarcomere in cardiac myocytes. Note that the Z-line is connected to the cardiac T-tubules. The diagram illustrates the complex protein-protein interactions that occur between structural components of the cytoskeleton and ion channels. The cytoskeleton is involved in regulating the metabolism of ion channels, modifying their expression, localization, and electrical properties. The cardiac sodium channel Nav1.5 associates with the DGC, while potassium channels such as Kv1.5, associate with the Z-line.
Ion Channel Subunits and Trafficking
Correct localization is essential for ion channel function and this is dependent upon the ability of auxiliary proteins to
- shuttle ion channels from the cytoplasm to their final destination such as
- the plasma membrane or other sub-cellular compartments.
In this regard, Kvβ-subunits are
- cytoplasmic components known to assemble with the α-subunits of voltage-dependent K+ (Kv) channels
- at their N-terminus to form stable Kvα/β hetero-oligomeric channels.
When Kvβ is co-expressed with Kv1.4 or Kv1.5, it enhances Kv1.x channel trafficking to the cell membrane without changing the overall protein channel content. The regulatory Kvβ subunits, which are also expressed in cardiomyocytes, directly decrease K+ current by
- accelerating Kv1.x channel inactivation.
Therefore, altered expression or mutations in Kvβ subunits could cause abnormal ion channel transport to the cell surface, thereby increasing the risk of cardiac arrhythmias.
Ion Channel Protein Motifs and Trafficking
Cell membrane trafficking in the Kv1.x family may occur in a Kvβ subunit-independent manner through specific motifs in their C-terminus. Mutagenesis of the final asparagine (N) in the Kv1.2 motif restores the leucine (L) of the Kv1.4 motif
- re-establishing high expression levels at the plasma membrane in a Kvβ-independent manner
Cytoskeletal Proteins and Ion Channel Trafficking
Until recently, primary arrhythmias such as LQTS have been almost exclusively regarded as ion channelopathies. Other mutations have been identified with regard to channelopathies. However, the conviction that primary mutations in ion channels were solely responsible for
- the electrical defects associated with arrhythmias
has been shaken by the identification of mutations in the
- ANK2 gene encoding the cytoskeletal protein ankyrin-B
that is associated with LQTS in animal models and humans.
Ankyrin-B acts as a chaperone protein, which shuttles the cardiac sodium channel from the cytoplasm to the membrane. Immunohistochemical analysis has localized ankyrin-B to the Zlines/T-tubules on the plasma membrane in the myocardium. Mutations in ankyrin-B associated with LQTS
- alter sodium channel trafficking due to loss of ankyrin-B localization at the Z-line/transverse (T)-tubules.
Reduced levels of ankyrin-B at cardiac Z-lines/T-tubules were associated with the deficiency of ankyrin-B-associated proteins such as Na/K-ATPase, Na/Ca exchanger (NCX) and inositol-1, 4, 5-trisphosphate receptors (InsP3R).
Dystrophin component of the Dystrophin Glycoprotien Complex (DGC)
Synchronized contraction is essential for cardiomyocytes, which are connected to each other via the extracellular matrix (ECM) through the DGC. The N-terminus domain of dystrophin
- binds F-actin, and connects it to the sarcomere, while
- the cysteine-rich (CR) C-terminus domain ensures its connection to the sarcolemma (Figure 2).
The central portion of dystrophin, the rod domain, is composed of
- rigid spectrin-like repeats and four hinge portions (H1–H4) that determine the flexibility of the protein.
Dystrophin possesses another F-actin binding domain in the Rod domain region, between the basic repeats 11- 17 (DysN-R17).
Dystrophin, originally identified as the gene responsible for Duchenne and Becker muscular dystrophies (DMD/BMD), and later for the X-linked form of dilated cardiomyopathy (XLCM), exerts a major function in physical force transmission in striated muscle. In addition to its structural significance, dystrophin and other DGC proteins such as syntrophins are required for the
- correct localization,
- clustering and
- regulation of ion channel function.
Syntrophins have been implicated in ion channel regulation. Syntrophins contain two pleckstrin homology (PH) domains, a PDZ domain, and a syntrophin-unique (SU) C-terminal region. The interaction between syntrophins and dystrophin occurs at the PH domain distal to the syntrophin N-terminus and through the highly conserved SU domain. Conversely, the PH domain proximal to the N-terminal portion of the protein and the PDZ domain interact with other membrane components such as
- phosphatidyl inositol-4, 5-bisphosphate,
- neuronal NOS (nNOS),
- aquaporin-4,
- stress-activated protein kinase-3, and
- 5,
thereby linking all these molecules to the dystrophin complex (Figure 2).
Among the five known isoforms of syntrophin, the 59 KDa α1-syntrophin isoform is the most highly represented in human heart, whereas in skeletal muscle it is only present on the
- sarcolemma of fast type II fibers.
In addition, the skeletal muscle γ2-syntrophin was found at high levels only at the
- postsynaptic membrane of the neuromuscular junctions.
In addition to syntrophin, other scaffolding proteins such as caveolin-3 (CAV3), which is present in the caveolae, flask-shaped plasma membrane microdomains, are involved
- in signal transduction and vesicle trafficking in myocytes,
- modulating cardiac remodeling during heart failure.
CAV3 and α1-syntrophin, localizes at the T-tubule and are part of the DGC. In addition, α1-syntrophin binds Nav1.5, while
- caveolin-3 binds the Na+/Ca2+ exchanger, Nav1.5 and the L-type Ca2+ channel as well as nNOS and the DGC (Figure 2).
Although ankyrin-B is the only protein found mutated in patients with primary arrhythmias, other proteins such as caveolin-3 and the syntrophins if mutated may alter ion channel function.
Conclusions
It is important to be aware of the enormous variety of clinical presentations that derive from distinct variants in the same pool of genetic factors. Knowledge of these variants could facilitate tailoring the therapy of choice for each patient. In particular, the recent findings of structural and functional links between
- the cytoskeleton and ion channels
could expand the therapeutic interventions in
- arrhythmia management in structurally abnormal myocardium, where aberrant binding
- between cytoskeletal proteins can directly or indirectly alter ion channel function.
Executive Summary
Arrhythmogenesis and myocardial structure
- Rhythm alterations can develop as a secondary consequence of myocardial structural abnormalities or as a result of a primary defect in the cardiac electric machinery.
- Until recently, no molecular mechanism has been able to fully explain the occurrence of arrhythmogenesis in heart failure, however genetic defects that are found almost exclusively in ion channel genes account for the majority of primary arrhythmias such as long QT syndromes and Brugada syndrome. The contractile apparatus is linked to ion channels
- The sarcomere, which represents the contractile unit of the myocardium not only generates the mechanical force necessary to exert the pump function, but also provides localization and anchorage to ion channels.
- Alpha-actinin-2, and telethonin, two members of the Z-line scaffolding protein complex in the striated muscle associate with the potassium voltage-gated channel alpha subunit Kv1.5 and the beta subunit KCNE1 respectively.
- Mutations in KCNE1 have previously been associated with the development of arrhythmias in LQTS subjects.
- Mutations in both alpha-actinin-2, and telethonin were identified in individuals with cardiomyopathy. The primary defect is structural leading to ventricular dysfunction, but the secondary consequence is arrhythmia.
Ion channel trafficking and sub-cellular compartments
- Ion channel trafficking from the endoplasmic reticulum (ER) to the Golgi complex is an important check-point for regulating the functional channel molecules on the plasma membrane. Several molecules acting as chaperones bind to and shuttle the channel proteins to their final localization on the cell surface
- Ion channel subunits such as Kvβ enhance Kv1.x ion channel presentation on the sarcolemma. The α subunits of the Kv1.x potassium channels can be shuttled in a Kvβ-independent manner through specific sequence motif at Kv1.x protein level.
- In addition, cytoskeletal proteins such as ankyrin-G bind Nav1.5 and are involved in the sodium channel trafficking. Another member of the ankyrin family, ankyrin-B was found mutated in patients with LQTS but the pathological mechanism of ankyrin-B mutations is still obscure, although the sodium current intensity is dramatically reduced.
The sarcolemma and ion channels
- The sarcolemma contains a wide range of ion channels, which are responsible for the electrical propagating force in the myocardium.
- The DGC is a protein complex, which forms a scaffold for cytoskeletal components and ion channels.
- Dystrophin is the major component of the DGC and mutations in dystrophin and DGC cause muscular dystrophies and X-linked cardiomyopathies (XLCM) in humans. Cardiomyopathies are associated with arrhythmias
- Caveolin-3 and syntrophins associate with Nav1.5, and are part of the DGC. Syntrophins can directly modulate Nav1.5 channel function.
Conclusions
- The role of the cytoskeleton in ion channel function has been hypothesized in the past, but only recently the mechanism underlying the development of arrhythmias in structurally impaired myocardium has become clearer.
- The recently acknowledged role of the cytoskeleton in ion channel function suggests that genes encoding cytoskeletal proteins should be regarded as potential candidates for variants involved in the susceptibility to arrhythmias, as well as the primary target of genetic mutations in patients with arrhythmogenic syndromes such as LQTS and Brugada syndrome.
- Studies of genotype-phenotype correlation and and patient risk stratification for mutations in cytoskeletal proteins will help to tailor the therapy and management of patients with arrhythmias.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}