Growth Factors, Suppressors and Receptors in Tumorigenesis
Writer and Curator: Larry H Bernstein, MD, FCAP
7.1 Growth Factors, Suppressors and Receptors in Tumorigenesis
7.1.1 Friend or Foe: Endoplasmic reticulum protein 29 (ERp29) in epithelial cancer
7.1.2 Putting together structures of epidermal growth factor receptors
7.1.3 Complex Relationship between Ligand Binding and Dimerization in the Epidermal Growth Factor Receptor
7.1.4 IGFBP-2.PTEN- A critical interaction for tumors and for general physiology
7.1.5 Emerging-roles-for-the-Ph-sensing-G-protein-coupled-receptor
7.1.6 Protein amino-terminal modifications and proteomic approaches for N-terminal profiling
7.1.7 Protein homeostasis networks in physiology and disease
7.1.8 Proteome sequencing goes deep
7.1.1 Friend or Foe: Endoplasmic reticulum protein 29 (ERp29) in epithelial cancer
Chen S1, Zhang D2
FEBS Open Bio. 2015 Jan 30; 5:91-8
http://dx.doi.org:/10.1016/j.fob.2015.01.004
The endoplasmic reticulum (ER) protein 29 (ERp29) is a molecular chaperone that plays a critical role in protein secretion from the ER in eukaryotic cells. Recent studies have also shown that ERp29 plays a role in cancer. It has been demonstrated that ERp29 is inversely associated with primary tumor development and functions as a tumor suppressor by inducing cell growth arrest in breast cancer. However, ERp29 has also been reported to promote epithelial cell morphogenesis, cell survival against genotoxic stress and distant metastasis. In this review, we summarize the current understanding on the biological and pathological functions of ERp29 in cancer and discuss the pivotal aspects of ERp29 as “friend or foe” in epithelial cancer.
The endoplasmic reticulum (ER) is found in all eukaryotic cells and is complex membrane system constituting of an extensively interlinked network of membranous tubules, sacs and cisternae. It is the main subcellular organelle that transports different molecules to their subcellular destinations or to the cell surface [10,85].
The ER contains a number of molecular chaperones involved in protein synthesis and maturation. Of the ER chaperones, protein disulfide isomerase (PDI)-like proteins are characterized by the presence of a thioredoxin domain and function as oxido-reductases, isomerases and chaperones [33]. ERp29 lacks the active-site double-cysteine (CxxC) motif and does not belong to the redox-active PDIs [5,47]. ERp29 is recognized as a characterized resident of the cellular ER, and it is expressed ubiquitously and abundantly in mammalian tissues [50]. Protein structural analysis showed that ERp29 consists of N-terminal and C-terminal domains [5]: N-terminal domain involves dimerization whereas the C-terminal domain is essential for substrate binding and secretion [78]. The biological function of ERp29 in protein secretion has been well established in cells [8,63,67].
ERp9 is proposed to be involved in the unfolded protein response (UPR) as a factor facilitating transport of synthesized secretory proteins from the ER to Golgi [83]. The expression of ERp29 was demonstrated to be increased in cells exposed to radiation [108], sperm cells undergoing maturation [42,107], and in certain cell types both under the pharmacologically induced UPR and under the physiological conditions (e.g., lactation, differentiation of thyroid cells) [66,82]. Under ER stress, ERp29 translocates the precursor protein p90ATF6 from the ER to Golgi where it is cleaved to be a mature and active form p50ATF by protease (S1P and S2P) [48]. In most cases, ERp29 interacts with BiP/GRP78 to exert its function under ER stress [65].
ERp29 is considered to be a key player in both viral unfolding and secretion [63,67,77,78] Recent studies have also demonstrated that ERp29 is involved in intercellular communication by stabilizing the monomeric gap junction protein connexin43 [27] and trafficking of cystic fibrosis transmembrane conductance regulator to the plasma membrane in cystic fibrosis and non-cystic fibrosis epithelial cells [90]. It was recently reported that ERp29 directs epithelial Na(+) channel (ENaC) toward the Golgi, where it undergoes cleavage during its biogenesis and trafficking to the apical membrane [40]. ERp29 expression protects axotomized neurons from apoptosis and promotes neuronal regeneration [111]. These studies indicate a broad biological function of ERp29 in cells.
Recent studies demonstrated a tumor suppressive function of ERp29 in cancer. It was found that ERp29 expression inhibited tumor formation in mice [4,87] and the level of ERp29 in primary tumors is inversely associated with tumor development in breast, lung and gallbladder cancer [4,29].
However, its expression is also responsible for cancer cell survival against genotoxic stress induced by doxorubicin and radiation [34,76,109]. The most recent studies demonstrate other important roles of ERp29 in cancer cells such as the induction of mesenchymal–epithelial transition (MET) and epithelial morphogenesis [3,4]. MET is considered as an important process of transdifferentiation and restoration of epithelial phenotype during distant metastasis [23,52]. These findings implicate ERp29 in promoting the survival of cancer cells and also metastasis. Hence, the current review focuses on the novel functions of ERp29 and discusses its pathological importance as a “friend or foe” in epithelial cancer.
ERp29 regulates mesenchymal–epithelial transition
Epithelial–mesenchymal transition (EMT) and MET
The EMT is an essential process during embryogenesis [6] and tumor development [43,96]. The pathological conditions such as inflammation, organ fibrosis and cancer progression facilitate EMT [16]. The epithelial cells after undergoing EMT show typical features characterized as: (1) loss of adherens junctions (AJs) and tight junctions (TJs) and apical–basal polarity; (2) cytoskeletal reorganization and distribution; and (3) gain of aggressive phenotype of migration and invasion [98]. Therefore, EMT has been considered to be an important process in cancer progression and its pathological activation during tumor development induces primary tumor cells to metastasize [95]. However, recent studies showed that the EMT status was not unanimously correlated with poorer survival in cancer patients examined [92].
In addition to EMT in epithelial cells, mesenchymal-like cells have capability to regain a fully differentiated epithelial phenotype via the MET [6,35]. The key feature of MET is defined as a process of transdifferentiation of mesenchymal-like cells to polarized epithelial-like cells [23,52] and mediates the establishment of distant metastatic tumors at secondary sites [22]. Recent studies demonstrated that distant metastases in breast cancer expressed an equal or stronger E-cadherin signal than the respective primary tumors and the re-expression of E-cadherin was independent of the E-cadherin status of the primary tumors [58]. Similarly, it was found that E-cadherin is re-expressed in bone metastasis or distant metastatic tumors arising from E-cadherin-negative poorly differentiated primary breast carcinoma [81], or from E-cadherin-low primary tumors [25]. In prostate and bladder cancer cells, the nonmetastatic mesenchymal-like cells were interacted with metastatic epithelial-like cells to accelerate their metastatic colonization [20]. It is, therefore, suggested that the EMT/MET work co-operatively in driving metastasis.
Molecular regulation of EMT/MET
E-cadherin is considered to be a key molecule that provides the physical structure for both cell–cell attachment and recruitment of signaling complexes [75]. Loss of E-cadherin is a hallmark of EMT [53]. Therefore, characterizing transcriptional regulators of E-cadherin expression during EMT/MET has provided important insights into the molecular mechanisms underlying the loss of cell–cell adhesion and the acquisition of migratory properties during carcinoma progression [73].
Several known signaling pathways, such as those involving transforming growth factor-β (TGF-β), Notch, fibroblast growth factor and Wnt signaling pathways, have been shown to trigger epithelial dedifferentiation and EMT [28,97,110]. These signals repress transcription of epithelial genes, such as those encoding E-cadherin and cytokeratins, or activate transcription programs that facilitate fibroblast-like motility and invasion [73,97].
The involvement of microRNAs (miRNAs) in controlling EMT has been emphasized [11,12,18]. MiRNAs are small non-coding RNAs (∼23 nt) that silence gene expression by pairing to the 3′UTR of target mRNAs to cause their posttranscriptional repression [7]. MiRNAs can be characterized as “mesenchymal miRNA” and “epithelial miRNA” [68]. The “mesenchymal miRNA” plays an oncogenic role by promoting EMT in cancer cells. For instance, the well-known miR-21, miR-103/107 are EMT inducer by repressing Dicer and PTEN [44].
The miR-200 family has been shown to be major “epithelial miRNA” that regulate MET through silencing the EMT-transcriptional inducers ZEB1 and ZEB2 [13,17]. MiRNAs from this family are considered to be predisposing factors for cancer cell metastasis. For instance, the elevated levels of the epithelial miR-200 family in primary breast tumors associate with poorer outcomes and metastasis [57]. These findings support a potential role of “epithelial miRNAs” in MET to promote metastatic colonization [15].
ERp29 promotes MET in breast cancer
The role of ERp29 in regulating MET has been established in basal-like MDA-MB-231 breast cancer cells. It is known that myosin light chain (MLC) phosphorylation initiates to myosin-driven contraction, leading to reorganization of the actin cytoskeleton and formation of stress fibers [55,56]. ERp29 expression in this type of cells markedly reduced the level of phosphorylated MLC [3]. These results indicate that ERp29 regulates cortical actin formation through a mechanism involved in MLC phosphorylation (Fig. 1). In addition to the phenotypic change, ERp29 expression leads to: expression and membranous localization of epithelial cell marker E-cadherin; expression of epithelial differentiation marker cytokeratin 19; and loss of the mesenchymal cell marker vimentin and fibronectin [3] (Fig. 1). In contrast, knockdown of ERp29 in epithelial MCF-7 cells promotes acquisition of EMT traits including fibroblast-like phenotype, enhanced cell spreading, decreased expression of E-cadherin and increased expression of vimentin [3,4]. These findings further substantiate a role of ERp29 in modulating MET in breast cancer cells.
Fig. 1 ERp29 triggers mesenchymal–epithelial transition. Exogenous expression of ERp29 in mesenchymal MDA-MB-231 breast cancer cells inhibits stress fiber formation by suppressing MLC phosphorylation. In addition, the overexpressed ERp29 decreases the …
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4329646/bin/gr1.gif
ERp29 targets E-cadherin transcription repressors
The transcription repressors such as Snai1, Slug, ZEB1/2 and Twist have been considered to be the main regulators for E-cadherin expression [19,26,32]. Mechanistic studies revealed that ERp29 expression significantly down-regulated transcription of these repressors, leading to their reduced nuclear expression in MDA-MB-231 cells [3,4] (Fig. 2). Consistent with this, the extracellular signal-regulated kinase (ERK) pathway which is an important up-stream regulator of Slug and Ets1 was highly inhibited [4]. Apparently, ERp29 up-regulates the expressions of E-cadherin transcription repressors through repressing ERK pathway. Interestingly, ERp29 over-expression in basal-like BT549 cells resulted in incomplete MET and did not significantly affect the mRNA or protein expression of Snai1, ZEB2 and Twist, but increased the protein expression of Slug [3]. The differential regulation of these transcriptional repressors of E-cadherin by ERp29 in these two cell-types may occur in a cell-context-dependent manner.
Fig. 2 ERp29 decreases the expression of EMT inducers to promote MET. Exogenous expression of ERp29 in mesenchymal MDA-MB-231 breast cancer cells suppresses transcription and protein expression of E-cadherin transcription repressors (e.g., ZEB2, SNAI1 and Twist), ..
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4329646/bin/gr2.gif
ERp29 antagonizes Wnt/ β-catenin signaling
Wnt proteins are a family of highly conserved secreted cysteine-rich glycoproteins. The Wnt pathway is activated via a binding of a family member to a frizzled receptor (Fzd) and the LDL-Receptor-related protein co-receptor (LRP5/6). There are three different cascades that are activated by Wnt proteins: namely canonical/β-catenin-dependent pathway and two non-canonical/β-catenin-independent pathways that include Wnt/Ca2+ and planar cell polarity [84]. Of note, the Wnt/β-catenin pathway has been extensively studied, due to its important role in cancer initiation and progression [79]. The presence of Wnt promotes formation of a Wnt–Fzd–LRP complex, recruitment of the cytoplasmic protein Disheveled (Dvl) to Fzd and the LRP phosphorylation-dependent recruitment of Axin to the membrane, thereby leading to release of β-catenin from membrane and accumulation in cytoplasm and nuclei. Nuclear β-catenin replaces TLE/Groucho co-repressors and recruits co-activators to activate expression of Wnt target genes. The most important genes regulated are those related to proliferation, such as Cyclin D1 and c-Myc [46,94], which are over-expressed in most β-catenin-dependent tumors. When β-catenin is absent in nucleus, the transcription factors T-cell factor/lymphoid enhancer factors (TCF/LEF) recruits co-repressors of the TLE/Groucho family and function as transcriptional repressors.
β-catenin is highly expressed in the nucleus of mesenchymal MDA-MB-231 cells. ERp29 over-expression in this type of cells led to translocation of nuclear β-catenin to membrane where it forms complex with E-cadherin [3] (Fig. 3). This causes a disruption of β-catenin/TCF/LEF complex and abolishes its transcription activity. Indeed, ERp29 significantly decreased the expression of cyclin D1/D2 [36], one of the downstream targets of activated Wnt/β-catenin signaling [94], indicating an inhibitory effect of ERp29 on this pathway. Meanwhile, expression of ERp29 in this cell type increased the nuclear expression of TCF3, a transcription factor regulating cancer cell differentiation while inhibiting self-renewal of cancer stem cells [102,106]. Hence, ERp29 may play dual functions in mesenchymal MDA-MB-231 breast cancer cells by: (1) suppressing activated Wnt/β-catenin signaling via β-catenin translocation; and (2) promoting cell differentiation via activating TCF3 (Fig. 3). Because β-catenin serves as a signaling hub for the Wnt pathway, it is particularly important to focus on β-catenin as the target of choice in Wnt-driven cancers. Though the mechanism by which ERp29 expression promotes the disassociation of β-catenin/TCF/LEF complex in MDA-MB-231 cells remains elusive, activating ERp29 expression may exert an inhibitory effect on the poorly differentiated, Wnt-driven tumors.
Fig. 3 ERp29 over-expression “turns-off” activated Wnt/β-catenin signaling. In mesenchymal MDA-MB-231 cells, high expression of nuclear β-catenin activates its downstream signaling involved in cell cycles and cancer stem cell …
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4329646/bin/gr3.gif
ERp29 regulates epithelial cell integrity
Cell adherens and tight junctions
Adherens junctions (AJs) and tight junctions (TJs) are composed of transmembrane proteins that adhere to similar proteins in the adjacent cell [69]. The transmembrane region of the TJs is composed mainly of claudins, tetraspan proteins with two extracellular loops [1]. AJs are mediated by Ca2+-dependent homophilic interactions of cadherins [71] which interact with cytoplasmic catenins that link the cadherin/catenin complex to the actin cytoskeleton [74].
The cytoplasmic domain of claudins in TJs interacts with occludin and several zona occludens proteins (ZO1-3) to form the plaque that associates with the cytoskeleton [99]. The AJs form and maintain intercellular adhesion, whereas the TJs serve as a diffusion barrier for solutes and define the boundary between apical and basolateral membrane domains [21]. The AJs and TJs are required for integrity of the epithelial phenotype, as well as for epithelial cells to function as a tissue [75].
The TJs are closely linked to the proper polarization of cells for the establishment of epithelial architecture[86]. During cancer development, epithelial cells lose the capability to form TJs and correct apico–basal polarity [59]. This subsequently causes the loss of contact inhibition of cell growth [91]. In addition, reduction of ZO-1 and occludin were found to be correlated with poorly defined differentiation, higher metastatic frequency and lower survival rates [49,64]. Hence, TJs proteins have a tumor suppressive function in cancer formation and progression.
Apical–basal cell polarity
The apical–basal polarity of epithelial cells in an epithelium is characterized by the presence of two specialized plasma membrane domains: namely, the apical surface and basolateral surface [30]. In general, the epithelial cell polarity is determined by three core complexes. These protein complexes include: (1) the partitioning-defective (PAR) complex; (2) the Crumbs (CRB) complex; and (3) the Scribble complex[2,30,45,51]. PAR complex is composed of two scaffold proteins (PAR6 and PAR3) and an atypical protein kinase C (aPKC) and is localized to the apical junction domain for the assembly of TJs [31,39]. The Crumbs complex is formed by the transmembrane protein Crumbs and the cytoplasmic scaffolding proteins such as the homologue of Drosophila Stardust (Pals1) and Pals-associated tight junction protein (Patj) and localizes to the apical [38]. The Scribble complex is comprised of three proteins, Scribble, Disc large (Dlg) and Lethal giant larvae (Lgl) and is localized in the basolateral domain of epithelial cells [100].
Fig. 4 ERp29 regulates epithelial cell morphogenesis. Over-expression of ERp29 in breast cancer cells induces the transition from a mesenchymal-like to epithelial-like phenotype and the restoration of tight junctions and cell polarity. Up-regulation and membrane …
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4329646/bin/gr4.gif
The current data from breast cancer cells supports the idea that ERp29 can function as a tumor suppressive protein, in terms of suppression of cell growth and primary tumor formation and inhibition of signaling pathways that facilitate EMT. Nevertheless, the significant role of ERp29 in cell survival against drugs, induction of cell differentiation and potential promotion of MET-related metastasis may lead us to re-assess its function in cancer progression, particularly in distant metastasis. Hence, it is important to explore in detail the ERp29’s role in cancer as a “friend or foe” and to elucidate its clinical significance in breast cancer and other epithelial cancers. Targeting ERp29 and/or its downstream molecules might be an alternative molecular therapeutic approach for chemo/radio-resistant metastatic cancer treatment
7.1.2 Putting together structures of epidermal growth factor receptors
Bessman NJ1, Freed DM2, Lemmon MA3
Curr Opin Struct Biol. 2014 Dec; 29:95-101
http://dx.doi.org:/10.1016/j.sbi.2014.10.002
Highlights
- Several studies suggest flexible linkage between extracellular and intracellular regions.
- Others imply more rigid connections, required for allosteric regulation of dimers.
- Interactions with membrane lipids play important roles in EGFR regulation.
- Cellular studies suggest half-of-the-sites negative cooperativity for human EGFR.
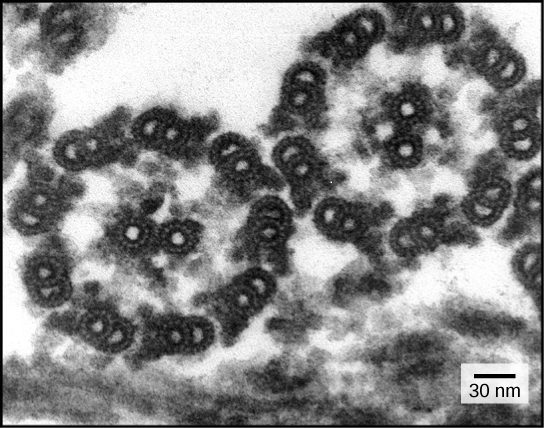
Numerous crystal structures have been reported for the isolated extracellular region and tyrosine kinase domain of the epidermal growth factor receptor (EGFR) and its relatives, in different states of activation and bound to a variety of inhibitors used in cancer therapy. The next challenge is to put these structures together accurately in functional models of the intact receptor in its membrane environment. The intact EGFR has been studied using electron microscopy, chemical biology methods, biochemically, and computationally. The distinct approaches yield different impressions about the structural modes of communication between extracellular and intracellular regions. They highlight possible differences between ligands, and also underline the need to understand how the receptor interacts with the membrane itself.
http://ars.els-cdn.com/content/image/1-s2.0-S0959440X14001304-gr1.sml
http://ars.els-cdn.com/content/image/1-s2.0-S0959440X14001304-gr2.sml
7.1.3 Complex Relationship between Ligand Binding and Dimerization in the Epidermal Growth Factor Receptor
Bessman NJ1, Bagchi A2, Ferguson KM2, Lemmon MA3.
Cell Rep. 2014 Nov 20; 9(4):1306-17.
http://dx.doi.org/10.1016/j.celrep.2014.10.010
Highlights
- Preformed extracellular dimers of human EGFR are structurally heterogeneous
- EGFR dimerization does not stabilize ligand binding
- Extracellular mutations found in glioblastoma do not stabilize EGFR dimerization
- Glioblastoma mutations in EGFR increase ligand-binding affinity
Summary
The epidermal growth factor receptor (EGFR) plays pivotal roles in development and is mutated or overexpressed in several cancers. Despite recent advances, the complex allosteric regulation of EGFR remains incompletely understood. Through efforts to understand why the negative cooperativity observed for intact EGFR is lost in studies of its isolated extracellular region (ECR), we uncovered unexpected relationships between ligand binding and receptor dimerization. The two processes appear to compete. Surprisingly, dimerization does not enhance ligand binding (although ligand binding promotes dimerization). We further show that simply forcing EGFR ECRs into preformed dimers without ligand yields ill-defined, heterogeneous structures. Finally, we demonstrate that extracellular EGFR-activating mutations in glioblastoma enhance ligand-binding affinity without directly promoting EGFR dimerization, suggesting that these oncogenic mutations alter the allosteric linkage between dimerization and ligand binding. Our findings have important implications for understanding how EGFR and its relatives are activated by specific ligands and pathological mutations.
http://www.cell.com/cms/attachment/2020816777/2040986303/fx1.jpg
X-ray crystal structures from 2002 and 2003 (Burgess et al., 2003) yielded the scheme for ligand-induced epidermal growth factor receptor (EGFR) dimerization shown in Figure 1. Binding of a single ligand to domains I and III within the same extracellular region (ECR) stabilizes an “extended” conformation and exposes a dimerization interface in domain II, promoting self-association with a KD in the micromolar range (Burgess et al., 2003, Dawson et al., 2005, Dawson et al., 2007). Although this model satisfyingly explains ligand-induced EGFR dimerization, it fails to capture the complex ligand-binding characteristics seen for cell-surface EGFR, with concave-up Scatchard plots indicating either negative cooperativity (De Meyts, 2008, Macdonald and Pike, 2008) or distinct affinity classes of EGF-binding site with high-affinity sites responsible for EGFR signaling (Defize et al., 1989). This cooperativity or heterogeneity is lost when the ECR from EGFR is studied in isolation, as also described for the insulin receptor (De Meyts, 2008).

ligand-induced-dimerization-of-the-hegfr-ecr
Figure 1
Structural View of Ligand-Induced Dimerization of the hEGFR ECR
(A) Surface representation of tethered, unliganded, sEGFR from Protein Data Bank entry 1NQL (Ferguson et al., 2003). Ligand-binding domains I and III are green and cysteine-rich domains II and IV are cyan. The intramolecular domain II/IV tether is circled in red.
(B) Hypothetical model for an extended EGF-bound sEGFR monomer based on SAXS studies of an EGF-bound dimerization-defective sEGFR variant (Dawson et al., 2007) from PDB entry 3NJP (Lu et al., 2012). EGF is blue, and the red boundary represents the primary dimerization interface.
(C) 2:2 (EGF/sEGFR) dimer, from PDB entry 3NJP (Lu et al., 2012), colored as in (B). Dimerization arm contacts are circled in red.
http://www.cell.com/cms/attachment/2020816777/2040986313/gr1.sml
Here, we describe studies of an artificially dimerized ECR from hEGFR that yield useful insight into the heterogeneous nature of preformed ECR dimers and into the origins of negative cooperativity. Our data also argue that extracellular structures induced by ligand binding are not “optimized” for dimerization and conversely that dimerization does not optimize the ligand-binding sites. We also analyzed the effects of oncogenic mutations found in glioblastoma patients (Lee et al., 2006), revealing that they affect allosteric linkage between ligand binding and dimerization rather than simply promoting EGFR dimerization. These studies have important implications for understanding extracellular activating mutations found in EGFR/ErbB family receptors in glioblastoma and other cancers and also for understanding specificity of ligand-induced ErbB receptor heterodimerization
Predimerizing the EGFR ECR Has Modest Effects on EGF Binding
To access preformed dimers of the hEGFR ECR (sEGFR) experimentally, we C-terminally fused (to residue 621 of the mature protein) either a dimerizing Fc domain (creating sEGFR-Fc) or the dimeric leucine zipper from S. cerevisiae GCN4 (creating sEGFR-Zip). Size exclusion chromatography (SEC) and/or sedimentation equilibrium analytical ultracentrifugation (AUC) confirmed that the resulting purified sEGFR fusion proteins are dimeric (Figure S1). To measure KD values for ligand binding to sEGFR-Fc and sEGFR-Zip, we labeled EGF with Alexa-488 and monitored binding in fluorescence anisotropy (FA) assays. As shown in Figure 2A, EGF binds approximately 10-fold more tightly to the dimeric sEGFR-Fc or sEGFR-Zip proteins than to monomeric sEGFR (Table 1). The curves obtained for EGF binding to sEGFR-Fc and sEGFR-Zip showed no signs of negative cooperativity, with sEGFR-Zip actually requiring a Hill coefficient (nH) greater than 1 for a good fit (nH = 1 for both sEGFRWT and sEGFR-Fc). Thus, our initial studies argued that simply dimerizing human sEGFR fails to restore the negatively cooperative ligand binding seen for the intact receptor in cells.
One surprise from these data was that forced sEGFR dimerization has only a modest (≤10-fold) effect on EGF-binding affinity. Under the conditions of the FA experiments, isolated sEGFR (without zipper or Fc fusion) remains monomeric; the FA assay contains just 60 nM EGF, so the maximum concentration of EGF-bound sEGFR is also limited to 60 nM, which is over 20-fold lower than the KD for dimerization of the EGF/sEGFR complex (Dawson et al., 2005, Lemmon et al., 1997). This ≤10-fold difference in affinity for dimeric and monomeric sEGFR seems small in light of the strict dependence of sEGFR dimerization on ligand binding (Dawson et al., 2005,Lax et al., 1991, Lemmon et al., 1997). Unliganded sEGFR does not dimerize detectably even at millimolar concentrations, whereas liganded sEGFR dimerizes with KD ∼1 μM, suggesting that ligand enhances dimerization by at least 104– to 106-fold. Straightforward linkage of dimerization and binding equilibria should stabilize EGF binding to dimeric sEGFR similarly (by 5.5–8.0 kcal/mol). The modest difference in EGF-binding affinity for dimeric and monomeric sEGFR is also significantly smaller than the 40- to 100-fold difference typically reported between high-affinity and low-affinity EGF binding on the cell surface when data are fit to two affinity classes of binding site (Burgess et al., 2003, Magun et al., 1980).
Mutations that Prevent sEGFR Dimerization Do Not Significantly Reduce Ligand-Binding Affinity
The fact that predimerizing sEGFR only modestly increased ligand-binding affinity led us to question the extent to which domain II-mediated sEGFR dimerization is linked to ligand binding. It is typically assumed that the domain II conformation stabilized upon forming the sEGFR dimer in Figure 1C optimizes the domain I and III positions for EGF binding. To test this hypothesis, we introduced a well-characterized pair of domain II mutations into sEGFRs that block dimerization: one at the tip of the dimerization arm (Y251A) and one at its “docking site” on the adjacent molecule in a dimer (R285S). The resulting (Y251A/R285S) mutation abolishes sEGFR dimerization and EGFR signaling (Dawson et al., 2005, Ogiso et al., 2002). Importantly, we chose isothermal titration calorimetry (ITC) for these studies, where all interacting components are free in solution. Previous surface plasmon resonance (SPR) studies have indicated that dimerization-defective sEGFR variants bind immobilized EGF with reduced affinity (Dawson et al., 2005), and we were concerned that this reflects avidity artifacts, where dimeric sEGFR binds more avidly than monomeric sEGFR to sensor chip-immobilized EGF.
Surprisingly, our ITC studies showed that the Y251A/R285S mutation has no significant effect on ligand-binding affinity for sEGFR in solution (Table 1). These experiments employed sEGFR (with no Fc fusion) at 10 μM—ten times higher than KD for dimerization of ligand-saturated WT sEGFR (sEGFRWT) (KD ∼1 μM). Dimerization of sEGFRWT should therefore be complete under these conditions, whereas the Y251A/R285S-mutated variant (sEGFRY251A/R285S) does not dimerize at all (Dawson et al., 2005). The KD value for EGF binding to dimeric sEGFRWT was essentially the same (within 2-fold) as that for sEGFRY251A/R285S (Figures 2B and 2C; Table 1), arguing that the favorable Gibbs free energy (ΔG) of liganded sEGFR dimerization (−5.5 to −8 kcal/mol) does not contribute significantly (<0.4 kcal/mol) to enhanced ligand binding. …
Thermodynamics of EGF Binding to sEGFR-Fc
If there is no discernible positive linkage between sEGFR dimerization and EGF binding, why do sEGFR-Fc and sEGFR-Zip bind EGF ∼10-fold more strongly than wild-type sEGFR? To investigate this, we used ITC to compare EGF binding to sEGFR-Fc and sEGFR-Zip (Figures 3A and 3B ) with binding to isolated (nonfusion) sEGFRWT. As shown in Table 1, the positive (unfavorable) ΔH for EGF binding is further elevated in predimerized sEGFR compared with sEGFRWT, suggesting that enforced dimerization may actually impair ligand/receptor interactions such as hydrogen bonds and salt bridges. The increased ΔH is more than compensated for, however, by a favorable increase in TΔS. This favorable entropic effect may reflect an “ordering” imposed on unliganded sEGFR when it is predimerized, such that it exhibits fewer degrees of freedom compared with monomeric sEGFR. In particular, since EGF binding does induce sEGFR dimerization, it is clear that predimerization will reduce the entropic cost of bringing two sEGFR molecules into a dimer upon ligand binding, possibly underlying this effect.
Possible Heterogeneity of Binding Sites in sEGFR-Fc
Close inspection of EGF/sEGFR-Fc titrations such as that in Figure 3A suggested some heterogeneity of sites, as evidenced by the slope in the early part of the experiment. To investigate this possibility further, we repeated titrations over a range of temperatures. We reasoned that if there are two different types of EGF-binding sites in an sEGFR-Fc dimer, they might have different values for heat capacity change (ΔCp), with differences that might become more evident at higher (or lower) temperatures. Indeed, ΔCp values correlate with the nonpolar surface area buried upon binding (Livingstone et al., 1991), and we know that this differs for the two Spitz-binding sites in the asymmetric Drosophila EGFR dimer (Alvarado et al., 2010). As shown in Figure 3C, the heterogeneity was indeed clearer at higher temperatures for sEGFR-Fc—especially at 25°C and 30°C—suggesting the possible presence of distinct classes of binding sites in the sEGFR-Fc dimer. We were not able to fit the two KD values (or ΔH values) uniquely with any precision because the experiment has insufficient information for unique fitting to a model with four variables. Whereas binding to sEGFRWT could be fit confidently with a single-site binding model throughout the temperature range, enforced sEGFR dimerization (by Fc fusion) creates apparent heterogeneity in binding sites, which may reflect negative cooperativity of the sort seen with dEGFR. …
Ligand Binding Is Required for Well-Defined Dimerization of the EGFR ECR
To investigate the structural nature of the preformed sEGFR-Fc dimer, we used negative stain electron microscopy (EM). We hypothesized that enforced dimerization might cause the unliganded ECR to form the same type of loose domain II-mediated dimer seen in crystals of unliganded Drosophila sEGFR (Alvarado et al., 2009). When bound to ligand (Figure 4A), the Fc-fused ECR clearly formed the characteristic heart-shape dimer seen by crystallography and EM (Lu et al., 2010, Mi et al., 2011). Figure 4B presents a structural model of an Fc-fused liganded sEGFR dimer, and Figure 4C shows a calculated 12 Å resolution projection of this model. The class averages for sEGFR-Fc plus EGF (Figure 4A) closely resemble this model, yielding clear densities for all four receptor domains, arranged as expected for the EGF-induced domain II-mediated back-to-back extracellular dimer shown in Figure 1 (Garrett et al., 2002, Lu et al., 2010). In a subset of classes, the Fc domain also appeared well resolved, indicating that these particular arrangements of the Fc domain relative to the ECR represent highly populated states, with the Fc domains occupying similar positions to those of the kinase domain in detergent-solubilized intact receptors (Mi et al., 2011). …
Our results and those of Lu et al. (2012)) argue that preformed extracellular dimers of hEGFR do not contain a well-defined domain II-mediated interface. Rather, the ECRs in these dimers likely sample a broad range of positions (and possibly conformations). This conclusion argues against recent suggestions that stable unliganded extracellular dimers “disfavor activation in preformed dimers by assuming conformations inconsistent with” productive dimerization of the rest of the receptor (Arkhipov et al., 2013). The ligand-free inactive dimeric ECR species modeled by Arkhipov et al. (2013) in their computational studies of the intact receptor do not appear to be stable. The isolated ECR from EGFR has a very low propensity for self-association without ligand, with KD in the millimolar range (or higher). Moreover, sEGFR does not form a defined structure even when forced to dimerize by Fc fusion. It is therefore difficult to envision how it might assume any particular autoinhibitory dimeric conformation in preformed dimers. …
Extracellular Oncogenic Mutations Observed in Glioblastoma May Alter Linkage between Ligand Binding and sEGFR Dimerization
Missense mutations in the hEGFR ECR were discovered in several human glioblastoma multiforme samples or cell lines and occur in 10%–15% of glioblastoma cases (Brennan et al., 2013, Lee et al., 2006). Several elevate basal receptor phosphorylation and cause EGFR to transform NIH 3T3 cells in the absence of EGF (Lee et al., 2006). Thus, these are constitutively activating oncogenic mutations, although the mutated receptors can be activated further by ligand (Lee et al., 2006, Vivanco et al., 2012). Two of the most commonly mutated sites in glioblastoma, R84 and A265 (R108 and A289 in pro-EGFR), are in domains I and II of the ECR, respectively, and contribute directly in inactive sEGFR to intramolecular interactions between these domains that are thought to be autoinhibitory (Figure 5). Domains I and II become separated from one another in this region upon ligand binding to EGFR (Alvarado et al., 2009), as illustrated in the lower part of Figure 5. Interestingly, analogous mutations in the EGFR relative ErbB3 were also found in colon and gastric cancers (Jaiswal et al., 2013).
We hypothesized that domain I/II interface mutations might activate EGFR by disrupting autoinhibitory interactions between these two domains, possibly promoting a domain II conformation that drives dimerization even in the absence of ligand. In contrast, however, sedimentation equilibrium AUC showed that sEGFR variants harboring R84K, A265D, or A265V mutations all remained completely monomeric in the absence of ligand (Figure 6A) at a concentration of 10 μM, which is similar to that experienced at the cell surface (Lemmon et al., 1997). As with WT sEGFR, however, addition of ligand promoted dimerization of each mutated sEGFR variant, with KD values that were indistinguishable from those of WT. Thus, extracellular EGFR mutations seen in glioblastoma do not simply promote ligand-independent ECR dimerization, consistent with our finding that even dimerized sEGFR-Fc requires ligand binding in order to form the characteristic heart-shaped dimer. …
We suggest that domain I is normally restrained by domain I/II interactions so that its orientation with respect to the ligand is compromised. When the domain I/II interface is weakened with mutations, this effect is mitigated. If this results simply in increased ligand-binding affinity of the monomeric receptor, the biological consequence might be to sensitize cells to lower concentrations of EGF or TGF-α (or other agonists). However, cellular studies of EGFR with glioblastoma-derived mutations (Lee et al., 2006, Vivanco et al., 2012) clearly show ligand-independent activation, arguing that this is not the key mechanism. The domain I/II interface mutations may also reduce restraints on domain II so as to permit dimerization of a small proportion of intact receptor, driven by the documented interactions that promote self-association of the transmembrane, juxtamembrane, and intracellular regions of EGFR (Endres et al., 2013, Lemmon et al., 2014, Red Brewer et al., 2009).
Setting out to test the hypothesis that simply dimerizing the EGFR ECR is sufficient to recover the negative cooperativity lost when it is removed from the intact receptor, we were led to revisit several central assumptions about this receptor. Our findings suggest three main conclusions. First, we find that enforcing dimerization of the hEGFR ECR does not drive formation of a well-defined domain II-mediated dimer that resembles ligand-bound ECRs or the unliganded ECR from Drosophila EGFR. Our EM and SAXS data show that ligand binding is necessary for formation of well-defined heart-shaped domain II-mediated dimers. This result argues that the unliganded extracellular dimers modeled by Arkhipov et al. (2013)) are not stable and that it is improbable that stable conformations of preformed extracellular dimers disfavor receptor activation by assuming conformations that counter activating dimerization of the rest of the receptor. Recent work from the Springer laboratory employing kinase inhibitors to drive dimerization of hEGFR (Lu et al., 2012) also showed that EGF binding is required to form heart-shaped ECR dimers. These findings leave open the question of the nature of the ECR in preformed EGFR dimers but certainly argue that it is unlikely to resemble the crystallographic dimer seen for unligandedDrosophila EGFR (Alvarado et al., 2009) or that suggested by computational studies (Arkhipov et al., 2013).
This result argues that ligand binding is required to permit dimerization but that domain II-mediated dimerization may compromise, rather than enhance, ligand binding. Assuming flexibility in domain II, we suggest that this domain serves to link dimerization and ligand binding allosterically. Optimal ligand binding may stabilize one conformation of domain II in the scheme shown in Figure 1 that is then distorted upon dimerization of the ECR, in turn reducing the strength of interactions with the ligand. Such a mechanism would give the appearance of a lack of positive linkage between ligand binding and ECR dimerization, and a good test of this model would be to determine the high-resolution structure of a liganded sEGFR monomer (which we expect to differ from a half dimer). This model also suggests a mechanism for selective heterodimerization over homodimerization of certain ErbB receptors. If a ligand-bound EGFR monomer has a domain II conformation that heterodimerizes with ErbB2 in preference to forming EGFR homodimers, this could explain several important observations. It could explain reports that ErbB2 is a preferred heterodimerization partner of EGFR (Graus-Porta et al., 1997) and might also explain why EGF binds more tightly to EGFR in cells where it can form heterodimers with ErbB2 than in cells lacking ErbB2, where only EGFR homodimers can form (Li et al., 2012).
7.1.4 IGFBP-2.PTEN- A critical interaction for tumors and for general physiology
Li Zeng, Claire M. Perks, Jeff M.P. Holly
Growth Hormone & IGF Research online 7 February 2015
http://dx.doi.org/10.1016/j.ghir.2015.01.003
Highlights
- IGFBP-2 is the second most abundant of the IGFBPs in the circulation.
- IGFBP2 levels are increased in a variety of tumors and associated with progression and poor prognosis.
- PTEN is a phosphatase that returns the PI3K/AKT/mTOR pathway to its inactivated state.
- PTEN is the second most commonly mutated gene in a variety of common cancers.
- Recent evidence indicates that IGFBP-2 regulates PTEN in a variety of normal and malignant cell types.
- This review summarizes the evidence that these extracellular and intracellular modulators of the IGF-system are linked.
Abstract
IGFBP-2 is an important modulator of IGF availability and activity. It is the second most abundant of the IGFBPs in the circulation and its levels are increased in a variety of tumors and associated with progression and poor prognosis. PTEN is a phosphatase that returns the PI3K/AKT/mTOR pathway to its inactivated state and is therefore a critical modulator of one of the main intracellular signaling pathways activated by the IGFs. Recent evidence has indicated that IGFBP-2 regulates PTEN in a variety of normal and malignant cell types. This review summarizes the recent evidence that these extracellular and intracellular modulators are linked to provide a synchronous system for cell regulation with coordinated control of both the ‘accelerator’ and the ‘brake’.

IGFBP-2.PTEN
http://ars.els-cdn.com/content/image/1-s2.0-S1096637415000167-gr1.sml
7.1.5 Emerging-roles-for-the-Ph-sensing-G-protein-coupled-receptor
Sanderlin EJ, Justus CR, Krewson EA, Yang LV
CHC March 2015 Volume 2015:7 Pages 99—109
Protons (hydrogen ions) are the simplest form of ions universally produced by cellular metabolism including aerobic respiration and glycolysis. Export of protons out of cells by a number of acid transporters is essential to maintain a stable intracellular pH that is critical for normal cell function. Acid products in the tissue interstitium are removed by blood perfusion and excreted from the body through the respiratory and renal systems. However, the pH homeostasis in tissues is frequently disrupted in many pathophysiologic conditions such as in ischemic tissues and tumors where protons are overproduced and blood perfusion is compromised. Consequently, accumulation of protons causes acidosis in the affected tissue. Although acidosis has profound effects on cell function and disease progression, little is known about the molecular mechanisms by which cells sense and respond to acidotic stress. Recently a family of pH-sensing G protein-coupled receptors (GPCRs), including GPR4, GPR65 (TDAG8), and GPR68 (OGR1), has been identified and characterized. These GPCRs can be activated by extracellular acidic pH through the protonation of histidine residues of the receptors. Upon activation by acidosis the pH-sensing GPCRs can transduce several downstream G protein pathways such as the Gs, Gq/11, and G12/13 pathways to regulate cell behavior. Studies have revealed the biological roles of the pH-sensing GPCRs in the immune, cardiovascular, respiratory, renal, skeletal, endocrine, and nervous systems, as well as the involvement of these receptors in a variety of pathological conditions such as cancer, inflammation, pain, and cardiovascular disease. As GPCRs are important drug targets, small molecule modulators of the pH-sensing GPCRs are being developed and evaluated for potential therapeutic applications in disease treatment.
Cellular metabolism produces acid as a byproduct. Metabolism of each glucose molecule by glycolysis generates two pyruvate molecules. Under anaerobic conditions the metabolism of pyruvate results in the production of the glycolytic end product lactic acid, which has a pKa of 3.9. Lactic acid is deprotonated at the carboxyl group and results in one lactate ion and one proton at the physiological pH. Under aerobic conditions pyruvate is converted into acetyl-CoA and CO2 in the mitochondria. CO2in water forms a chemical equilibrium of carbonic acid and bicarbonate, an important physiological pH buffering system. The body must maintain suitable pH for proper physiological functions. Some regulatory mechanisms to control systemic pH are respiration, renal excretion, bone buffering, and metabolism.1–4 The respiratory system can buffer the blood by excreting carbonic acid as CO2 while the kidney responds to decreased circulatory pH by excreting protons and electrolytes to stabilize the physiological pH. Bone buffering helps maintain systemic pH by Ca2+ reabsorption and mineral dissolution. Collectively, it is clear that several biological systems require tight regulation to maintain pH for normal physiological functions. Cells utilize vast varieties of acid-base transporters for proper pH homeostasis within each biological context.5–8 Some such transporters are H+-ATPase, Na+/H+exchanger, Na+-dependent HCO3–/C1– exchanger, Na+-independent anion exchanger, and monocarboxylate transporters. Cells can also maintain short-term pH homeostasis of the intracellular pH by rapid H+ consuming mechanisms. Some such mechanisms utilize metabolic conversions that move acids from the cytosol into organelles. Despite these cellular mechanisms that tightly maintain proper pH homeostasis, there are many diseases whereby pH homeostasis is disrupted. These pathological conditions are characterized by either local or systemic acidosis. Systemic acidosis can occur from respiratory, renal, and metabolic diseases and septic shock.1–4,9 Additionally, local acidosis is characterized in ischemic tissues, tumors, and chronically inflamed conditions such as in asthma and arthritis caused by deregulated metabolism and hypoxia.10–15
Acidosis is a stress for the cell. The ability of the cell to sense and modulate activity for adaptation to the stressful environment is critical. There are several mechanisms whereby cells sense acidosis and modulate cellular functions to facilitate adaptation. Cells can detect extracellular pH changes by acid sensing ion channels (ASICs) and transient receptor potential (TRP) channels.16 Apart from ASIC and TRP channels, extracellular acidic pH was shown to stimulate inositol polyphosphate formation and calcium efflux.17,18 This suggested the presence of an unknown cell surface receptor that may be activated by a certain functional group, namely the imidazole of a histidine residue. The identity of the acid-activated receptor was later unmasked by Ludwig et al as a family of proton-sensing G protein-coupled receptors (GPCRs). This group identified human ovarian cancer GPCR 1 (OGR1) which upon activation will produce inositol phosphate and calcium efflux through the Gq pathway.19 These pH-sensing GPCR family members, including GPR4, GPR65 (TDAG8), and GPR68 (OGR1), will be discussed in this review (Figure 1). The proton-sensing GPCRs sense extracellular pH by protonation of several histidine residues on their extracellular domain. The activation of these proton-sensing GPCRs facilitates the downstream signaling through the Gq/11, Gs, and G12/13 pathways. Their expression varies in different cell types and play critical roles in sensing extracellular acidity and modulating cellular functions in several biological systems.
Figure 1 Biological roles and G protein coupling of the pH-sensing GPCRs

Biological roles and G protein coupling of the pH-sensing GPCRs
http://www.dovepress.com/cr_data/article_fulltext/s60000/60508/img/fig1small.jpg
Cells encounter acidotic stress in many pathophysiologic conditions such as inflammation, cancer, and ischemia. Intricate molecular mechanisms, including a large array of acid/base transporters and acid sensors, have evolved for cells to sense and respond to acidotic stress. Emerging evidence has demonstrated that a family of the pH-sensing GPCRs can be activated by extracellular acidotic stress and regulate the function of multiple physiological systems (Table 1). The pH-sensing GPCRs also play important roles in various pathological disorders. Agonists, antagonists and other modulators of the pH-sensing GPCRs are being actively developed and evaluated as potential novel treatment for acidosis-related diseases.
| Table 1 The main biological functions of the pH-sensing GPCRs |

Table1 The main biological functions of the pH-sensing GPCRs
http://www.dovepress.com/cr_data/article_fulltext/s60000/60508/img/Table1small.jpg
7.1.6 Protein amino-terminal modifications and proteomic approaches for N-terminal profiling
Lai ZW1, Petrera A2, Schilling O3.
Curr Opin Chem Biol. 2015 Feb; 24:71-9
http://dx.doi.org/10.1016/j.cbpa.2014.10.026
Highlights
- N-terminal acetylation, pyroglutamate formation, N-degrons and proteolysis are reviewed.
- N-terminomics provide comprehensive profiling of modification at protein N-termini in a proteome-wide manner.
- We outline a number of established methodologies for the enrichment of protein N-termini through positive and negative selection strategies.
- Peptidomics-based approach is beneficial for the study of post-translational processing of protein N-termini.
Amino-/N-terminal processing is a crucial post-translational modification affecting almost all proteins. In addition to altering the chemical properties of the N-terminus, these modifications affect protein activation, conversion, and degradation, which subsequently lead to diversified biological functions. The study of N-terminal modifications is of increasing interest; especially since modifications such as proteolytic truncation or pyroglutamate formation have been linked to disease processes. During the past decade, mass spectrometry has played an important role in facilitating the investigation of N-terminal modifications. Continuous progress is being made in the development and application of robust methods for the dedicated analysis of native and modified protein N-termini in a proteome-wide manner. Here we highlight recent progress in our understanding of protein N-terminal biology as well as outlining present enrichment strategies for mass spectrometry-based studies of protein N-termini
7.1.7 Protein homeostasis networks in physiology and disease
Claudio Hetz1,2,3,* and Laurie H. Glimcher3,4,*
Curr Opin Cell Biol. 2011 Apr; 23(2): 123–125.
http://dx.doi.org/10.1016%2Fj.ceb.2011.01.004
Although most text books of biochemistry describe the process of protein folding to a three dimensional native state as an intrinsic property of the primary sequence, it is becoming increasingly clear that this process can go wrong in an almost infinite number of ways. In fact, many different diseases are caused by the misfolding and aggregation of certain proteins without genetic mutations in the primary sequence. An integrative view of the mechanisms that maintain protein folding homeostasis is emerging, which could be thought as a balanced and dynamic network of interconnected processes tightly regulated by a series of quality control mechanisms. This protein homeostasis network involves families of folding catalysts, co-factors under specific environmental and metabolic conditions. Maintaining protein homeostasis is particularly challenging in specialized secretory cells where the high demand for protein synthesis generates a constant source of stress that could lead to proteotoxicity.
Protein folding is assisted and monitored by diverse interconnected processes that follow a sequential pattern over time. The calnexin/calreticulin cycle ensures the proper folding of glycosylated proteins through the secretory pathway, which establishes the final pattern of disulfide bond formation through interactions with the disulfide isomerase ERp57. Coupled to this cycle is the ER-associated degradation (ERAD) pathway, which translocates terminally misfolded proteins to the cytosol for degradation by proteasomes. In addition, macroautophagy is becoming a relevant mechanism for the clearance of damaged proteins and abnormal protein aggregates through lysosomal hydrolysis, a process also referred to as ERAD-II. The folding status at the ER is constantly monitored by the Unfolded Protein Response (UPR), a specialized signaling pathway initiated by the activation of three types of stress sensors. The process underlying the surveillance of protein folding stress by the UPR is not fully understood, but it may require coupling to key folding mediators such as BiP or the direct recognition of the misfolded peptides by stress sensors. The UPR regulates genes and processs related to almost every folding step in the secretory pathway to reduce the load of misfolded proteins, including protein translation into the ER, translocation, folding, quality control, ERAD, the redox status, and many other related functions. Protein folding stress is observed in many disease conditions such as cancer, diabetes, and neurodegeneration. For example, abnormal protein aggregation and the accumulation of protein inclusions is associated with Parkinson’s and Alzheimer’s Disease, and amyotrophic lateral sclerosis. In those diseases and many others, neuronal dysfunction and disease progression correlates with the presence of a strong ER stress response; however, the direct in vivo role of the UPR in the disease process has been experimentally defined in only a few cases. Therapeutic strategies are currently being developed to increase protein folding and clearance of misfolded proteins, with the goal of alleviating ER stress.
In this issue of Current Opinion in Cell Biology we present a series of focused reviews from recognized experts in the field, that provide an overview of mechanisms underlying protein folding and quality control, and how balance of protein homeostasis is maintained in physiology and deregulated in diseases. Daniela Roth and William Balch integrate the concept of protein homeostasis networks into an interesting model termed FoldFx, showing how the interconnection between different pathways in the context of the cellular proteome determines the energetic barrier required to generate a functional folded peptide. The authors have previously proposed the term Proteostasis to refer to the set of interacting activities that maintain the health of the proteome and the organism (protein homeostasis). The ER is a central subcellular compartment for protein synthesis and quality control in the secretory pathway. Yukio Kimata and Kenji Kohno give an overview of the signaling pathways that control adaptation to ER stress and maintenance of protein folding homeostasis. The authors summarize the models proposed so far for the activation of UPR stress sensors, and discuss how this directly or indirectly relates to the accumulation of unfolded proteins in the ER lumen. Chronic or irreversible ER stress triggers cell death by apoptosis. Gordon Shore, Feroz Papa, and Scott Oakes summarize the complex signaling pathways initiating apoptosis by ER stress, where cross talk between the ER and the mitochondria play a central role. The authors focus on addressing the role of the BCL-2 protein family on the activation of intrinsic mitochondrial apoptosis pathways, highlighting different cytosolic and transcriptional events that determine the transition between adaptive responses to apoptosis programmed by the UPR to eliminate irreversibly injured cells.
Although diverse families of chaperones, foldases and co-factors are expressed at the ER, only a few protein folding networks have been well defined. However, molecular explanations for specific substrate recognition and quality control mechanisms are poorly defined. Here we present a series of reviews covering different aspects of protein maturation. Amy Lee summarizes what is known about the biology of the key ER folding chaperone BiP/Grp78, and its emerging role in diverse pathological conditions including cancer. In two reviews, David B. Williams and Linda M. Hendershot describe the best characterized mechanism of protein quality control at the ER, the calnexin cycle. In addition, they give an overview of the function of a family of ER foldases, the protein disulfide isomerases (PDIs), in folding, quality control and degradation of abnormally folded proteins. PDIs are also becoming key factors in establishing the redox tone of the ER. Riccardo Bernasconi and Maurizio Molinari overview the ERAD process and how this pathway affects the efficiency of the protein folding process at the ER and its relation to pathological conditions.
Lysosomal-mediated degradation is becoming a fundamental process for the control of the haft-life of proteins and the degradation of misfolded, aggregate prone proteins. Ana Maria Cuervo reviews the relevance of Chaperone-mediated autophagy in the selective degradation of soluble cytosolic proteins in lysosomes, and also points out a key role for Chaperone-mediated autophagy in the cellular defense against proteotoxicity. David Rubinsztein and Guido Kroemer present two reviews highlighting the emerging relevance of macroautophagy in maintaining the homeostasis of the nervous system. They also discuss the actual impact of macroautophagy in the clearance of protein aggregates related to neurodegenerative diseases, including Parkinson’s disease, amyotrophic lateral sclerosis, Huntington’s disease among others. In addition, recent evidence suggesting an actual impairment of macroautophagy as a causative factor in aging-related disorders is also discussed.
Strategies to increase the efficiency of quality control mechanisms, to reduce protein aggregation and to enhance folding are suggested to be beneficial in the setting of diseases associated with the disruption of protein homeostasis. Jeffery Kelly reviews recent chemical and biological therapeutic strategies to restore protein homeostasis, which could be achieved by enhancing the biological capacity of the proteostasis network or through small molecule to stabilize misfolding-prone proteins. In summary, this volume of Current Opinion in Cell Biology compiles the most recent advances in understanding the impact of protein folding stress in physiology and disease, and integrates a variety of complex mechanisms that evolved to maintain protein homeostasis in a dynamic way in the context of a changing environment. The biomedical applications of developing strategies to cope with protein folding stress have profound implications for the treatment of the most prevalent diseases in the human population.
7.1.8 Proteome sequencing goes deep
Richards AL1, Merrill AE2, Coon JJ3.
Curr Opin Chem Biol. 2015 Feb; 24:11-7
http://dx.doi.org/10.1016/j.cbpa.2014.10.017
Highlights
- Recent MS advances have transformed the depth of coverage of the human proteome.
- Expression of half the estimated human protein coding genes can be verified by MS.
- MS sample preparation, instrumentation, and data analysis techniques are highlighted.
Advances in mass spectrometry (MS) have transformed the scope and impact of protein characterization efforts. Identifying hundreds of proteins from rather simple biological matrices, such as yeast, was a daunting task just a few decades ago. Now, expression of more than half of the estimated ∼20 000 human protein coding genes can be confirmed in record time and from minute sample quantities. Access to proteomic information at such unprecedented depths has been fueled by strides in every stage of the shotgun proteomics workflow — from sample processing to data analysis — and promises to revolutionize our understanding of the causes and consequences of proteome variation.
- Advances in proteomic sample preparation
- Advances in peptide separation and MS instrumentation
- Advances in computational proteomics
- Conclusions and outlook
Mg²+ is critical for maintaining the positional integrity of closely clustered phosphate groups. These clusters appear in numerous and distinct parts of the cell nucleus and cytoplasm. The Mg²+ ion maintains the integrity of nucleic acids, ribosomes and proteins. In addition, this ion acts as an oligo-element with role in energy catalysis. [6] Biological cell membranes and cell walls exhibit poly-anionic charges on the surface. This finding has important implications for the transport of ions, particularly because different membranes preferentially bind different ions. Both Mg²+ and Ca²+ regularly stabilize membranes by cross-linking the carboxylated and phosphorylated head groups of lipids.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}