Cytoskeleton and Cell Membrane Physiology
Curator: Larry H Bernstein, MD, FCAP

cell-membrane

early evolution of lipid membranes and the three domains of life
Definition and Function
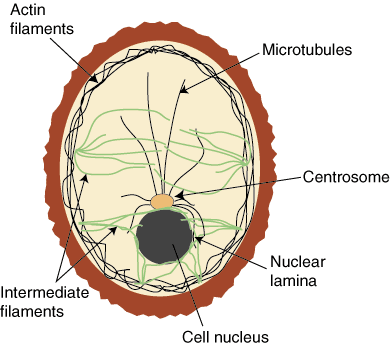
The cytoskeleton is a series of intercellular proteins that help a cell with
- shape,
- support, and
- movement.
Cytoskeleton has three main structural components:
- microfilaments,
- intermediate filaments, and
- movement
The cytoskeleton mediates movement by
- helping the cell move in its environment and
- mediating the movement of the cell’s components.
Thereby it provides an important structural framework for the cell –
- the framework for the movement of organelles, contiguous with the cell membrane, around the cytoplasm. By the activity of
- the network of protein microfilaments, intermediate filaments, and microtubules.
The structural framework supports cell function as follows:
Cell shape. For cells without cell walls, the cytoskeleton determines the shape of the cell. This is one of the functions of the intermediate filaments.
Cell movement. The dynamic collection of microfilaments and microtubles can be continually in the process of assembly and disassembly, resulting in forces that move the cell. There can also be sliding motions of these structures. Audesirk and Audesirk give examples of white blood cells “crawling” and the migration and shape changes of cells during the development of multicellular organisms.
Organelle movement. Microtubules and microfilaments can help move organelles from place to place in the cell. In endocytosis a vesicle formed engulfs a particle abutting the cell. Microfilaments then attach to the vesicle and pull it into the cell. Much of the complex synthesis and distribution function of the endoplasmic reticulum and the Golgi complex makes use of transport vescicles, associated with the cytoskeleton.
Cell division. During cell division, microtubules accomplish the movement of the chromosones to the daughter nucleus. Also, a ring of microfilaments helps divide two developing cells by constricting the central region between the cells (fission).
References:
Hickman, et al. Ch 4 Hickman, Cleveland P., Roberts, Larry S., and Larson, Allan, Integrated Principles of Zoology, 9th. Ed., Wm C. Brown, 1995.
Audesirk & Audesirk Ch 6 Audesirk, Teresa and Audesirk, Gerald, Biology, Life on Earth, 5th Ed., Prentice-Hall, 1999.
http://hyperphysics.phy-astr.gsu.edu/hbase/biology/bioref.html#c1
http://hyperphysics.phy-astr.gsu.edu/hbase/biology/cytoskel.html
Intermediate filaments are 8-12 nanometers in diameter and are twisted together in a cord shape. They are composed of keratin and keratin-like proteins. These filaments are tough and resist tension.
Microtubules are composed of alpha and beta tubulin that form long, hollow cylinders. These are fairly strong proteins and are the largest component of cytoskeleton at 25 nanometers. Tubular monomers can be lengthened or shortened from the positive end.
Microtubules have three different functions.
They make up the cell’s
- centriole
- the flagella and cilia of a cell, and
- they serve as “tracks” for transport vesicles to move along.
http://biology.kenyon.edu/HHMI/Biol113/cytoskeleton.htm
Key Points
Microtubules
- help the cell resist compression,
- provide a track along which vesicles can move throughout the cell, and
- are the components of cilia and flagella.
Cilia and flagella are hair-like structures that
- assist with locomotion in some cells, as well as
- line various structures to trap particles.
The structures of cilia and flagella are a “9+2 array,” meaning that
- a ring of nine microtubules is surrounded by two more microtubules.
Microtubules attach to replicated chromosomes
- during cell division and
- pull them apart to opposite ends of the pole,
- allowing the cell to divide with a complete set of chromosomes in each daughter cell.
Microtubules are the largest element of the cytoskeleton.
The walls of the microtubule are made of
- polymerized dimers of α-tubulin and β-tubulin, two globular proteins.
https://figures.boundless.com/18608/full/figure-04-05-04ab.jpe
With a diameter of about 25 nm, microtubules are the widest components of the cytoskeleton.
https://figures.boundless.com/18608/full/figure-04-05-04ab.jpe
They help the cell
- resist compression,
- provide a track along which vesicles move through the cell, and
- pull replicated chromosomes to opposite ends of a dividing cell.
Like microfilaments, microtubules can dissolve and reform quickly.
Microtubules are also the structural elements of flagella, cilia, and centrioles (the latter are the two perpendicular bodies of the centrosome). In animal cells, the centrosome is the microtubule-organizing center. In eukaryotic cells, flagella and cilia are quite different structurally from their counterparts in prokaryotes.
Intermediate Filaments
Intermediate filaments (IFs) are cytoskeletal components found in animal cells. They are composed of a family of related proteins sharing common structural and sequence features.

epithelial cells
https://figures.boundless.com/22035/full/epithelial-cells.jpe
flagella and cilia share a common structural arrangement of microtubules called a “9 + 2 array.” This is an appropriate name because a single flagellum or cilium is made of a ring of nine microtubule doublets surrounding a single microtubule doublet in the center.
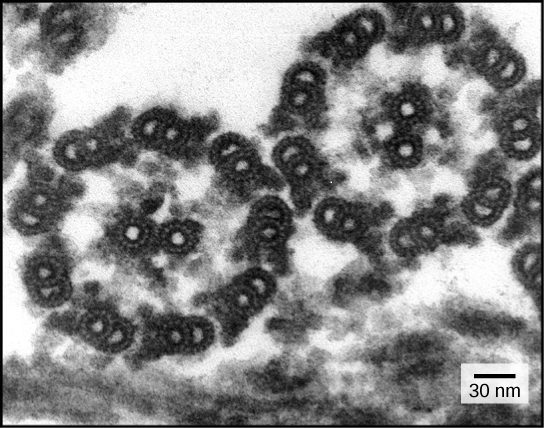
9+2 array
https://figures.boundless.com/18609/full/figure-04-05-05.jpe
http://jcs.biologists.org/content/115/22/4215/F4.large.jpg
{kind=link}
The `Spectraplakins’: cytoskeletal giants with characteristics of both spectrin and plakin families
Katja Röper, Stephen L. Gregory and Nicholas H. Brown
J Cell Sci Nov 15, 2002; 115: 4215-4225
http://dx.doi.org:/10.1242/jcs.00157

cytoskel
http://plantphys.info/plant_physiology/images/cytoskelfcns.gif
{kind=link}
cytoskeleton
http://img.sparknotes.com/figures/D/d479f5da672c08a54f986ae699069d7a/cytoskeleton.gif
{kind=link}
The sequential endosymbiotic origins of eukaryotes: Compared to bacteria and archaea, the typical eukaryotic cell is much more structurally complex.
While the prokaryotes have a rigid cell wall, the ancestral eukaryote appears to have been wall-less (the walls of plant cells appear to represent a adaptation, and are not homologous to prokaryotic cell walls).
In addition to a nucleus (wherein the cell’s DNA is located, and which we will return to in the next section), there are cytoskeletal structures, including distinctive flagella (quite different from those found in prokaryotes), an active (motile) plasma membrane, capable of engulfing other cells, and multiple internal membrane systems. (A more complete description of cell structure is beyond this version of Biofundamentals).
In aerobic bacteria and cyanobacteria, the electron transport chains associated with ATP synthesis (through either photosynthesis or aerobic respiration) located within the plasma membrane (and in the case of cyanobacteria, internal membrane systems as well).
The same processes (aerobic respiration and photosynthesis) occur within eukaryotic cells. Animals have aerobic respiration, while plants have both).
However, these processes do not occur on the plasma membrane, but rather within distinct cytoplasmic organelles: mitochondria for aerobic respiration and chloroplasts for photosynthesis. All eukaryotic cells have mitochondria, plants (which are eukaryotic) have both mitochondria and chloroplasts.
An intriguing evolutionary question was, are these processes related, that is, are the processes of aerobic respiration and photosynthesis found in eukaryotes homologous to the processes found in bacteria and cyanobacteria, or did they originate independently.
The path to understanding that homologous nature of these processes began with studies of cell structure.
http://virtuallaboratory.colorado.edu/Biofundamentals/lectureNotes-Revision/Topic2I_Symbiosis.htm

spectrin protein superfamily.large
http://mmbr.asm.org/content/70/3/605/F4.large.jpg
{kind=link}

The role of secreted factors and extracellular matrix
Focal Adhesions: Transmembrane Junctions Between the Extracellular Matrix and the Cytoskeleton
K Burridge, K Fath, T Kelly, G Nuckolls, and C Turner
Ann Rev Cell Biol Nov 1988; 4: 487-525
http://dx.doi.org:/10.1146/annurev.cb.04.110188.002415
the extracellular matrix (ECM) is a collection of extracellular molecules secreted by cells that
- provides structural and biochemical support to the surrounding cells.[1]
Because multicellularity evolved independently in different multicellular lineages, the composition of ECM varies between multicellular structures; however,
- cell adhesion,
- cell-to-cell communication and
- differentiation
are common functions of the ECM.[2]
The animal extracellular matrix includes
- the interstitial matrix and
- the basement membrane.[3]
Interstitial matrix is present between various animal cells (i.e., in the intercellular spaces).
Gels of polysaccharides and fibrous proteins
- fill the interstitial space and act as
- a compression buffer against the stress placed on the ECM.[4]
Basement membranes are sheet-like depositions of ECM on which various epithelial cells rest.
The Extracellular Matrix (ECM)
http://userpage.chemie.fu-berlin.de/biochemie/aghaucke/lehre/cytoskelet-ECM.pdf
Mechanical support to tissues
{kind=link}
http://www.nature.com/scitable/content/integrin-connects-the-extracellular-matrix-with-the-14707425
Organization of cells into tissues
- Activation of signaling pathways (cell growth, proliferation; development); examples:
- TGF-β, integrins
- specialized roles (tendon, bone; cartilage; cell movement during development; basal lamina in epithelia)
Components
- proteoglycans
- collagen fibers (mechanical strength)
- multiadhesive matrix proteins (linking cell surface receptors to the (ECM)
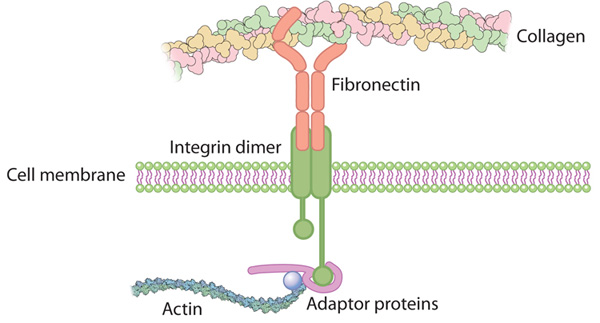
Integrin connects the extracellular matrix with the actin cytoskeleton inside the cell

Fibronectin Integrin
http://www.nature.com/scitable/content/integrin-connects-the-extracellular-matrix-with-the-14707425
Continuous membrane-cytoskeleton adhesion requires continuous accommodation to lipid and cytoskeleton dynamics.
Sheetz MP, Sable JE, Döbereiner HG.
Annu Rev Biophys Struct Biomol. 2006;35:417-34.
The plasma membrane of most animal cells conforms to the cytoskeleton and only occasionally separates to form blebs. Previous studies indicated that
- many weak interactions between cytoskeleton and the lipid bilayer
- kept the surfaces together to counteract the normal outward pressure of cytoplasm.
Either the loss of adhesion strength or the formation of gaps in the cytoskeleton enables the pressure to form blebs. Membrane-associated cytoskeleton proteins, such as spectrin and filamin, can
- control the movement and aggregation of membrane proteins and lipids,
e.g., phosphoinositol phospholipids (PIPs), as well as blebbing.
At the same time, lipids (particularly PIPs) and membrane proteins affect
- cytoskeleton and signaling dynamics.
We consider here the roles of the major phosphatidylinositol-4,5-diphosphate (PIP2) binding protein, MARCKS, and PIP2 levels in controlling cytoskeleton dynamics. Further understanding of dynamics will provide important clues about how membrane-cytoskeleton adhesion rapidly adjusts to cytoskeleton and membrane dynamics. http://www.ncbi.nlm.nih.gov/pubmed/16689643
Interaction of membrane/lipid rafts with the cytoskeleton: impact on signaling and function: membrane/lipid rafts, mediators of cytoskeletal arrangement and cell signaling.
Head BP, Patel HH, Insel PA Epub 2013 Jul 27.
Biochim Biophys Acta. 2014 Feb;1838(2):532-45.
http://dx.doi.org:/10.1016/j.bbamem.2013.07.018
The plasma membrane in eukaryotic cells contains microdomains that are
- enriched in certain glycosphingolipids, gangliosides, and sterols (such as cholesterol) to form membrane/lipid rafts (MLR).
These regions exist as caveolae, morphologically observable flask-like invaginations, or as a less easily detectable planar form. MLR are scaffolds for many molecular entities, including
- signaling receptors and ion channels that
- communicate extracellular stimuli to the intracellular milieu.
Much evidence indicates that this organization and/or the clustering of MLR into more active signaling platforms
- depends upon interactions with and dynamic rearrangement of the cytoskeleton.
Several cytoskeletal components and binding partners, as well as enzymes that regulate the cytoskeleton, localize to MLR and help
- regulate lateral diffusion of membrane proteins and lipids in response to extracellular events
(e.g., receptor activation, shear stress, electrical conductance, and nutrient demand).
MLR regulate
- cellular polarity,
- adherence to the extracellular matrix,
- signaling events (including ones that affect growth and migration), and
- are sites of cellular entry of certain pathogens, toxins and nanoparticles.
The dynamic interaction between MLR and the underlying cytoskeleton thus regulates many facets of the function of eukaryotic cells and their adaptation to changing environments. Here, we review general features of MLR and caveolae and their role in several aspects of cellular function, including
- polarity of endothelial and epithelial cells,
- cell migration,
- mechanotransduction,
- lymphocyte activation,
- neuronal growth and signaling, and
- a variety of disease settings.
This article is part of a Special Issue entitled: Reciprocal influences between cell cytoskeleton and membrane channels, receptors and transporters. Guest Editor: Jean Claude Hervé.
Cell control by membrane–cytoskeleton adhesion
Michael P. Sheetz
Nature Reviews Molecular Cell Biology 2, 392-396 (May 2001) | http://dx.doi.doi:/10.1038/35073095
The rates of mechanochemical processes, such as endocytosis, membrane extension and membrane resealing after cell wounding, are known to be controlled biochemically, through interaction with regulatory proteins. Here, I propose that these rates are also controlled physically, through an apparently continuous adhesion between plasma membrane lipids and cytoskeletal proteins.
Lipid Rafts, Signalling and the Cytoskeleton
http://www.bms.ed.ac.uk/research/others/smaciver/Cyto-Topics/lipid_rafts_and_the_cytoskeleton.htm
Lipid rafts are specialised membrane domains enriched in certain lipids cholesterol and proteins. The existence of lipid rafts was first hypothesised in 1988 (Simons & van Meer, 1988; Simon & Ikonen, 1997), but what we know as “caveolae” were first observed much earlier (Palade, 1953; Yamada, 1955). Caveolae are flask shaped invaginations on the cell surface that are a type of membrane raft, these were named “caveolae intracellulare” (Yamada, 1955). After a long argument (Jacobson & Dietrich, 1999), most now consider that these rafts actually exist, however, there is some confusion surrounding the classification of these rafts. It presently seems that there could be three types; caveolae, glycosphingolipid enriched membranes (GEM), and polyphospho inositol rich rafts. It may also be that there are inside rafts (PIP2 rich and caveolae) and outside rafts (GEM).
The fatty-acid chains of lipids within the rafts tend to be extended and so more tightly packed, creating domains with higher order. It is therefore thought that rafts exist in a separate ordered phase that floats in a sea of poorly ordered lipids. Glycosphingolipids, and other lipids with long, straight acyl chains are preferentially incorporated into the rafts.
Caveolae are similar in composition to GEMs that lack caveolae and in fact cells that lack caveolin-1 do not have morphologically identifiable caveolae but instead have extra GEM. These cells can then be transfected with caveolin-1 cDNA and the caveolae then appear. This suggests that GEM are merely caveolae without caveolin-1. Caveolin-1 is a 21kDa integral membrane protein that binds cholesterol (Maruta et al, 1995). In cells lacking caveolin-1, caveolin-2 is synthesised but remains in the Golgi. Caveolin 1 and 2 colocalise when expressed in the same cells and they may form hetero-dimers (Scherer et al, 1997). Caveolin-3 is expressed in muscle where it forms muscle-type caveolae. Caveolin-3 is involved in certain types of muscular dystrophy (Galbiati et al, ). A slightly confusing finding is that caveolae are the reported site of integrin signalling (). It is difficult to imagine integrins being available in the depths of membrane invaginations for binding extra-cellular ligands.
The function of rafts
Many functions have been attributed to rafts, from cholesterol transport, endocytosis and signal transduction. The later is almost certainly the case. It has been suggested that the primary function of caveolae was in constitutive endocytic trafficking but recent data show that this is not the case, instead caveolae are very stable regions of membranes that are not involved in endocytosis (Thompsen et al, 2002).

lipid raft
Rafts and the Cytoskeleton
Many actin binding proteins are known to bind to polyphosphoinositides and to be regulated by them (see PI and ABPs), by a series of protein domains such as PH, PX and ENTH (see Domains). It is consequently scarcely surprising that some ABPs are suggested to link the actin cytoskeleton and PIP2-enriched rafts. One of these is gelsolin, a Ca2+, pH and polyphosphoinositide regulated actin capping and severing protein (see Gelsolin Family), that partitions into rafts isolated biochemically from brain (Fanatsu et al, 2000).
GEMs too are suggested to link to the actin cytoskeleton through ABPs particularly ERM proteins through EBP50, a protein that binds members of the ERM proteins through the ERM C-terminus (Brdickova et al, 2001).
References:–
Brdickova, N., Brdicka, T., Andrea, L., Spicka, J., Angelisova, P., Milgram, S. L. & Horejsi, V. (2001) Interaction between two adaptor proteins, PAG and EBP50: a possible link between membrane rafts and actin cytoskeleton. FEBS letters. 507, 133-136.
Cary, L. A. & Cooper, J. A. (2000) Molecular switches in lipid rafts. Nature. 404, 945-947.
Czarny, M., Fiucci, G., Lavie, Y., Banno, Y., Nozawa, Y. & Liscovitch, M. (2000) Phospholipase D2: functional interaction with caveolin in low-density membrane microdomains., FEBS letters.
Foger, N., Funatsu, N., Kumanogoh, H., Sokawa, Y. & Maekawa, S. (2000) Identification of gelsolin as an actin regulatory component in a Triton insoluble low density fraction (raft) of newborn bovine brain. Neuroscience Research. 36, 311-317.
Galbiati, F., Engelman, J. A., Volonte, D., Zhang, X. L., Minetti, C., Li, M., Hou jr, H., Kneitz, B., Edelman, W. & Lisanti, M. P. (2001) Caveolin-3 null mice show a loss of caveolae, changes in the microdomain distribution of the dystrophin-glycoprotein complex, and T-tubule abnormalities. J. Biol.Chem. 276, 21425-21433.
… (more)

centralpore-small Gating and Ion Conductivity
Interaction of epithelial ion channels with the actin-based cytoskeleton.
Mazzochi C, Benos DJ, Smith PR.
Am J Physiol Renal Physiol. 2006 Dec;291(6):F1113-22. Epub 2006 Aug 22
The interaction of ion channels with the actin-based cytoskeleton in epithelial cells
- not only maintains the polarized expression of ion channels within specific membrane domains,
- it also functions in the intracellular trafficking and regulation of channel activity.
Initial evidence supporting an interaction between
- epithelial ion channels and the
- actin-based cytoskeleton
came from patch-clamp studies of the effects of cytochalasins on channel activity. Cytochalasins were shown to
- either activate or inactivate epithelial ion channels.
An interaction between
- the actin-based cytoskeleton and epithelial ion channels
was further supported by the fact that the addition of monomeric or filamentous actin to excised patches had an effect on channel activity comparable to that of cytochalasins. Through the recent application of molecular and proteomic approaches, we now know that
- the interactions between epithelial ion channels and actin can either be direct or indirect,
- the latter being mediated through scaffolding or actin-binding proteins
- that serve as links between the channels and the actin-based cytoskeleton.
This review discusses recent advances in our understanding of the interactions between epithelial ion channels and the actin-based cytoskeleton, and the roles these interactions play in regulating the cell surface expression, activity, and intracellular trafficking of epithelial ion channels.

epithelial ion channels
Actin cytoskeleton regulates ion channel activity in retinal neurons.
Maguire G, Connaughton V, Prat AG, Jackson GR Jr, Cantiello HF.
Neuroreport. 1998 Mar 9;9(4):665-70
The actin cytoskeleton is an important contributor to the integrity of cellular shape and responses in neurons. However, the molecular mechanisms associated with functional interactions between the actin cytoskeleton and neuronal ion channels are largely unknown. Whole-cell and single channel recording techniques were thus applied to identified retinal bipolar neurons of the tiger salamander (Ambystoma tigrinum) to assess the role of acute changes in actin-based cytoskeleton dynamics in the regulation of voltage-gated ion channels. Disruption of endogenous actin filaments after brief treatment (20-30 min) with cytochalasin D (CD) activated voltage-gated K+ currents in bipolar cells, which were largely prevented by intracellular perfusion with the actin filament-stabilizer agent, phalloidin. Either CD treatment under cell-attached conditions or direct addition of actin to excised, inside-out patches of bipolar cells activated and/or increased single K+ channels. Thus, acute changes in actin-based cytoskeleton dynamics regulate voltage-gated ion channel activity in bipolar cells.
Cytoskeletal Basis of Ion Channel Function in Cardiac Muscle
Matteo Vatta, Ph.D1,2 and Georgine Faulkner, Ph.D3
The publisher’s final edited version of this article is available at Future Cardiol
The heart is a force-generating organ that responds to self-generated electrical stimuli from specialized cardiomyocytes. This function is modulated by sympathetic and parasympathetic activity.
In order to contract and accommodate the repetitive morphological changes induced by the cardiac cycle,
- cardiomyocytes depend on their highly evolved and specialized cytoskeletal apparatus.
Defects in components of the cytoskeleton, in the long term, affect
- the ability of the cell to compensate at
- both functional and structural levels.
In addition to the structural remodeling, the myocardium becomes
- increasingly susceptible to altered electrical activity leading to arrhythmogenesis.
The development of arrhythmias secondary to structural remodeling defects has been noted, although the detailed molecular mechanisms are still elusive. Here I will review the current knowledge of the molecular and functional relationships between the cytoskeleton and ion channels and, I will discuss the future impact of new data on molecular cardiology research and clinical practice.
Stretch-activated ion channel
Stretch-activated or stretch-gated ion channels are
- ion channels which open their pores in response to
- mechanical deformation of a neuron’s plasma membrane.
[Also see mechanosensitive ion channels and mechanosensitive channels, with which they may be synonymous]. Opening of the ion channels
- depolarizes the afferent neuron producing an action potential with sufficient depolarization.[1]
Channels open in response to two different mechanisms: the prokaryotic model and the mammalian hair cell model.[2][3] Stretch-activated ion channels have been shown to detect vibration, pressure, stretch, touch, sounds, tastes, smell, heat, volume, and vision.[4][5][6] Stretch-activated ion channels have been categorized into
three distinct “superfamilies”:
- the ENaC/DEG family,
- the TRP family, and
- the K1 selective family.
These channels are involved with bodily functions such as blood pressure regulation.[7] They are shown to be associated with many cardiovascular diseases.[3] Stretch-activated channels were first observed in chick skeletal muscles by Falguni Guharay and Frederick Sachs in 1983 and the results were published in 1984.[8] Since then stretch-activated channels have been found in cells from bacteria to humans as well as plants.
Mechanosensitivity of cell membranes. Ion channels, lipid matrix and cytoskeleton.
Petrov AG, Usherwood PN.
Eur Biophys J. 1994;23(1):1-19
Physical and biophysical mechanisms of mechano-sensitivity of cell membranes are reviewed. The possible roles of
- the lipid matrix and of
- the cytoskeleton in membrane mechanoreception
are discussed. Techniques for generation of static strains and dynamic curvatures of membrane patches are considered. A unified model for
- stress-activated and stress-inactivated ion channels
under static strains is described. A review of work on
- stress-sensitive pores in lipid-peptide model membranes
is presented. The possible role of flexoelectricity in mechano-electric transduction, e.g. in auditory receptors is discussed. Studies of
- flexoelectricity in model lipid membranes, lipid-peptide membranes and natural membranes containing ion channels
are reviewed. Finally, possible applications in molecular electronics of mechanosensors employing some of the recognized principles of mechano-electric transduction in natural membranes are discussed.Marhaba, R. & Zoller, M. (2001) Involvement of CD44 in cytoskeleton rearrangement and raft reorganization in T cells. J.Cell Sci. 114, 1169-1178.
FIGURE 2 | The transient pore model.

peroxisomal matrix protein
FROM THE FOLLOWING ARTICLE:
Peroxisomal matrix protein import: the transient pore model
Ralf Erdmann & Wolfgang Schliebs
Nature Reviews Molecular Cell Biology 6, 738-742 (September 2005)
http://dx.doi.org:/10.1038/nrm1710
Peroxisomal matrix protein import: the transient pore model
The transient pore model
The peroxisomal import receptor peroxin-5 (Pex5) recognizes peroxisomal targeting signal-1 (PTS1)-containing cargo proteins in the cytosol. It then moves to the peroxisome where it inserts into the peroxisomal membrane to become an integral part of the protein-import apparatus. Pex14 and/or Pex13, which are associated with Pex17, are proposed to be involved in tethering the receptor to the membrane and in the assembly, stabilization and rearrangement of the translocon. Cargo release into the peroxisomal matrix is thought to be initiated by intraperoxisomal factors — for example, the competitive binding of the intraperoxisomal Pex8, which also has a PTS1. The disassembly and recycling of Pex5 is triggered by a cascade of protein–protein interactions at the peroxisomal membrane that results in the Pex1-, Pex6-driven, ATP-dependent dislocation of Pex5 from the peroxisomal membrane to the cytosol. Pex1 and Pex6 are AAA+ (ATPases associated with a variety of cellular activities) peroxins that are associated with the peroxisome membrane through Pex15 in yeast or its orthologue PEX26 in mammals. Pex4, which is membrane-anchored through Pex22, is a member of the E2 family of ubiquitin-conjugating enzymes, and Pex2, Pex10 and Pex12 contain the RING-finger motif that is a characteristic element of E3 ubiquitin ligases. Mono- or di-ubiquitylation are reversible steps that seem to be required for the efficient recycling of import receptors, whereas polyubiquitylation might signal the proteasome-dependent degradation of receptors when the physiological dislocation of receptors is blocked. Ub, ubiquitin.
Nature Reviews Molecular Cell Biology 6, 738-742 (September 2005) |
http://dx.doi.org:/10.1038/nrm1710
FROM THE FOLLOWING ARTICLE:

peroxisomal protein pore model
Peroxisomal matrix protein import: the transient pore model
Ralf Erdmann & Wolfgang Schliebs
Nature Reviews Molecular Cell Biology 6, 738-742 (September 2005)
http://dx.doi.org:/10.1038/nrm1710
Peroxisomal matrix protein import: the transient pore model
Peroxin-13 (Pex13), Pex14 and Pex17 are constituents of the docking complex for cycling peroxisomal import receptors. Another protein assembly in the peroxisomal membrane comprises the RING-finger-motif-containing peroxins Pex2, Pex10 and Pex12. This motif is a characteristic element of E3 ubiquitin ligases, and this subcomplex is linked to the docking complex by Pex8, which is peripherally attached to the lumenal side of the peroxisomal membrane. Pex4 is a member of the E2 family of ubiquitin-conjugating enzymes and is anchored to the peroxisomal membrane through the cytosolic domain of Pex22. Pex1 and Pex6 are interacting AAA+ proteins (ATPases associated with a variety of cellular activities), which are attached to the membrane through binding to Pex15 in yeast or to its mammalian counterpart PEX26.
Peroxisomal matrix protein import: the transient pore model
Ralf Erdmann & Wolfgang Schliebs
Peroxisomes import folded, even oligomeric, proteins, which distinguishes the peroxisomal translocation machinery from the well-characterized translocons of other organelles. How proteins are transported across the peroxisomal membrane is unclear. Here, we propose a mechanistic model that conceptually divides the import process into three consecutive steps: the formation of a
- translocation pore by the import receptor,
- the ubiquitylation of the import receptors, and
- pore disassembly/receptor recycling.
Phytosphingosine
Masoud Naderi Maralani
Identification of the phytosphingosine metabolic pathway leading to odd-numbered fatty acids
The long-chain base phytosphingosine is a component of sphingolipids and exists in yeast, plants and some mammalian tissues. Phytosphingosine is unique in that it possesses an additional hydroxyl group compared with other long-chain bases. However, its metabolism is unknown. Here we show that phytosphingosine is metabolized to odd-numbered fatty acids and is incorporated into glycerophospholipids both in yeast and mammalian cells. Disruption of the yeast gene encoding long-chain base 1-phosphate lyase, which catalyzes the committed step in the metabolism of phytosphingosine to glycerophospholipids, causes an ~40% reduction in the level of phosphatidylcholines that contain a C15 fatty acid. We also find that 2-hydroxypalmitic acid is an intermediate of the phytosphingosine metabolic pathway. Furthermore, we show that the yeast MPO1 gene, whose product belongs to a large, conserved protein family of unknown function, is involved in phytosphingosine metabolism. Our findings provide insights into fatty acid diversity and identify a pathway by which hydroxyl group-containing lipids are metabolized. nature.com nature.com
About GPCRs
G-protein-coupled receptors (GPCRs) are a class of membrane proteins that allow the transmission of a wide variety of signals over the cell membrane, between different cells and over long distances inside the body. The molecular mechanisms of action of GPCRs were worked in great detail by Brian Kobilka and Robert Lefkowitz for which they were jointly awarded the Nobel Prize in Chemistry for 2012. Read More
Leave a Reply