Curator: Larry H Bernstein, MD, FACP
The Open Clinical Chemistry Journal, 2011; 4: 34-44
http://occj.com/1874-2416/11 2011/
http://dx.doi.org/11.2011/occl/1874-2416/
Bentham Open Open Access
Introduction: The following document is a seminal article concerning the relationship between hyoerhomocysteinemia and cardiovascular and other diseases. It provides a new insight based on the metabolism of S8 and geographic factors affecting the distribution, the differences of plant and animal sources of dietary intake,
and the great impact on methylation reactions. The result is the finding that hyperhomocysteine is a “signal”, just as CRP is a measure of IL-6, IL-1, TNFa -mediated inflammatory response. A deficiency of S8 due to the unavailability of S8, leads to CVD, and is seen in sulfur deficient regions with inadequate soil content and with veganism. Hyperhomocysteinemia is also an indicator of CVD risk in the well fed populations, and that gives us a good reason to ASK WHY?
I have trimmed the content to make the necessary points that would be sufficient for this content. The article can be viewed at OCCJ online.
The Oxidative Stress of Hyperhomocysteinemia Results from Reduced Bioavailability of Sulfur-Containing Reductants
Yves Ingenbleek*
Laboratory of Nutrition, Faculty of Pharmacy, University Louis Pasteur Strasbourg, France
Abstract
A combination of subclinical malnutrition and S8-deficiency
- maximizes the defective production of Cys, GSH and H2S reductants,
- explaining persistence of unabated oxidative burden.
The clinical entity
- increases the risk of developing cardiovascular diseases (CVD) and stroke
- in underprivileged plant-eating populations
- regardless of Framingham criteria and vitamin-B status.
Although unrecognized up to now,
- the nutritional disorder is one of the commonest worldwide,
- reaching top prevalence in populated regions of Southeastern Asia.
Increased risk of hyperhomocysteinemia and oxidative stress may also affect
- individuals suffering from intestinal malabsorption or
- westernized communities having adopted vegan dietary lifestyles.
Vegetarian subjects
- consuming subnormal amounts of methionine (Met) are characterized by
- subclinical protein malnutrition causing reduction in size of their lean body mass (LBM) best
- identified by the serial measurement of plasma transthyretin (TTR).
As a result, the transsulfuration pathway is depressed at cystathionine-beta-synthase (CbS) level
- triggering the upstream sequestration of homocysteine (Hcy) in biological fluids and
- promoting its conversion to Met.
Maintenance of beneficial Met homeostasis is
- counterpoised by the drop of cysteine (Cys) and glutathione (GSH) values downstream to
- CbS causing in turn declining generation of hydrogen sulfide (H2S) from enzymatic sources.
The biogenesis of H2S via non-enzymatic reduction is further inhibited in areas where
- earth’s crust is depleted in elemental sulfur (S8) and sulfate oxyanions.
Keywords: Vegetarianism, malnutrition, sulfur-deficiency, hyperhomocysteinemia, oxidative stress, hydrogen sulfide, cardiovascular diseases, developing countries, Asia.
Homocysteine (Hcy) Generated by Transmethylation Pathway and Degraded via Transsulfuration Pathway
Homocysteine (Hcy) is a nonproteogenic sulfur containing amino acid (SAA)
- generated by the intrahepatic transmethylation (TM) of dietary Met.
- It may either be recycled to Met following remethylation (RM) pathways or
- catabolized along the transsulfuration (TS) cascade.
Under normal circumstances, the Met-Hcy cycle stands under the regulatory control of three water soluble B-vitamins:
- folates (5-methyl-tetrahydrofolates, B9) are regarded as the main factor working as donor of the CH3 group involved in the remethylation process,
- pyridoxine (pyridoxal-5’-phosphate, PLP, B6) plays the role of co-factor of both
- cystathionase enzymes belonging to the TS pathway and cobalamins (B12) ensure that of methionine-synthase.
Met-Hcy-Met Cycle
The main steps of the Met _ Hcy _Met cycle are summarized in Fig. (1).
FIGURE 1 NR H2S
Fig. (1). Schematic representation of the methionine cycle and homocysteine degradation pathways.
Compounds: ATP, adenosyltriphosphate; THF, tetrahydrofolate; SAM, S- adenosylmethionine; SAH, adenosylhomocysteine; Cysta, cystathionine; Cys, cysteine;
GSH, glutathione; H2S, hydrogen sulfide; Tau, taurine; SO4-2 , sulfate oxyanions.
Enzymes: (1) Met-adenosyltransferase; (2) SAM-methyltransferases; (3) adenosyl-homocysteinase; (4) methylene-THF reductase; (5) Metsynthase; (6) cystathionine
-b-synthase, CbS; (7) cystathionine-b-lyase, CbL; (8) g-glutamyl-synthase; (9) g-glutamyl-transpeptidase; (10)oxidase; (11) reductase; (12) cysteine-dioxygenase, CDO.
Metabolic pathways
Met molecules supplied by dietary proteins are
- submitted to TM processes
- releasing Hcy which may in turn either
- undergo Hcy_Met RM pathways or be
- irreversibly committed into TS decay.
Impairment of CbS activity in protein malnutrition, entails
- supranormal accumulation of Hcy in body fluids,
- stimulation of (5) activity and maintenance of Met homeostasis.
This last beneficial effect is counteracted by
- decreased concentration of most components generated downstream to CbS,
- explaining the depressed CbS- and CbL-mediated enzymatic production of *H2S along the TS cascade.
The restricted dietary intake of elemental S is a limiting factor for
- its non-enzymatic reduction to **H2S which contributes to
- downsizing a common body pool (dotted circle).(Fig 1)
Combined protein- and S-deficiencies work in concert
- to deplete Cys, GSH and H2S from their body reserves,
- impeding these reducing molecules from countering
- the oxidative stress imposed by hyperhomocysteinemia.
Hyperhomocysteinemia
Hyperhomocysteinemia (HHcy) is an acquired metabolic anomaly first identified by McCully [1]
The current consensus is that dietary deficiency in any of
three water soluble vitamins may operate as causal factor of HHcy.
- PLP–deficiency may trigger the upstream accumulation of Hcy in biological fluids [2] whereas
- the shortage of vitamins B9 or B12 is held responsible for its downstream sequestration [3,4].
HHcy is regarded as a major causal determinant of CVD
- working as an independent and graded risk factor
- unrelated to the classical Framingham criteria such as
- hypercholesterolemia,
- dyslipidemia,
- sedentary lifestyle,
- diabetes and
- smoking.
Hcy may invade the intracellular space of many tissues and locally generate [5]
- endothelial dysfunction working as early harbinger of blood vessel injuries and atherosclerosis.
Most investigators contend
- that production of harmful reactive oxygen and nitrogen species (ROS, NOS), notably
- hydrogen peroxide (H2O2), superoxide anion (O2 .-) and peroxinitrite (ONOO.-),
- constitutes a major culprit in the development of HHcy-induced vascular damages [7-10].
Accumulation of ROS
associated with increased risk for
- cardiovascular diseases [11]
- stroke [12],
- arterial hypertension [6],
- kidney dysfunction [13],
- Alzheimer’s disease [14],
- cognitive deterioration [15],
- inflammatory bowel disease [16] and
- bone remodeling [17].
These effects overlook the protective roles played by
- extra- and intracellular reductants such as cysteine (Cys) and glutathione (GSH)
- in the sequence of events leading from HHcy to tissue damage.
Hydrogen Sulfide (H2S)
After the discovery of nitric oxide (NO) and carbon oxide (CO), hydrogen sulfide (H2S) is the
- third gaseous signaling messenger found in mammalian tissues [18].
H2S is a reducing molecule displaying strong scavenging properties
- as the gasotransmitter significantly attenuates [19, 20] or
- even abolishes [21,22] the oxidative injury imposed by HHcy burden.
The endogenous production of the naturally occurring H2S reductant depends on
- Cys bioavailability through
- the mediation of TS enzymes [23,24].
H2S may also be produced in human tissues starting from elemental sulfur,
- by a non-enzymatic reaction requiring the presence of Cys, GSH, and glucose [25,26].
It would be worth disentangling the respective roles played by
- Cys,
- GSH
- H2S
- for the prevention and restoration of HHcy-induced oxidative lesions.
- but the plasma concentration of Cys and GSH is severely depressed in
- subclinically malnourished HHcy patients [27],
- impeding appropriate biogenesis of H2S molecules.
The present paper reviews the biological consequences
- resulting from the complex interplay existing between the 3 reducing molecules,
- to gain insight into the pathophysiologic mechanisms associated with HHcy states.
CLINICAL BACKGROUND
Numerous surveys have conclusively shown that the water soluble vitamin deficiency concept,
- provides only partial causal account of the HHcy metabolic anomaly.
The components of body composition, mainly
- the size of lean body mass (LBM),
- constitutes a critical determinant of HHcy status [28,29].
Because nitrogen (N) and sulfur (S) concentrations
- maintain tightly correlated ratios in tissues, we hypothesize
- defective N intake and accretion rate would cause concomitant and
- proportionate depletion of total body N (TBN) and total body S (TBS) stores [30].
Our clinical investigation undertaken in Central Africa in apparently healthy but
- nevertheless subclinically malnourished vegetarian subjects has
- documented that reduced size of LBM could lead to HHcy states [27].
The field study conducted in the Republic of Chad, populated by the Sara ethnic group [27], is a semi-arid region and
- the staple food consists mainly of cassava, sweet potatoes, beans, millets and groundnuts.
Participants were invited to fill in a detailed dietary questionnaire whose results were compared with values reported in food composition tables [32-34] [27].
The dietary inquiry indicates that participants
- consumed a significantly lower mean SAA intake (10.4 mg.kg-1.d-1)[27]
- than the Recommended Dietary Allowances (RDAs) (13 mg.kg-1.d-1)[33,34].
Blood Analytes
The blood lipid profiles of rural subjects were confined within normal ranges
- ruling out this class of parameters as causal risk factors for CVD disorders.
The normal levels measured for pyridoxine, folates, and cobalamins
- precluded these vitamins from playing any significant role in the rise of Hcy
plasma concentrations [27]. Analysis of plasma SAAs revealed
- unmodified methioninemia, significantly
- elevated Hcy values (18.6 umol/L)
- contrasting with significantly decreased plasma Cys and GSH values [27].
The significant lowering of classical
- anthropometric parameters
- (body weight, BW;
- body mass index, BMI)
- together with that of the main plasma and urinary biomarkers of
- metabolic (visceral) and
- structural (muscular) compartments point to
an estimated 10 % shrinking of LBM [27].
Transthyretin (TTR) and Lean Body Mass (LBM)
We have attached peculiar importance to the measurement of plasma transthyretin (TTR)
- this indicator integrates the evolutionary trends outlined by body protein reserves [35],
- providing from birth until death an overall and balanced estimate of LBM fluctuations [29].
- In the absence of any superimposed inflammatory condition,
- LBM and TTR profiles indeed reveal striking similarities [29].
Scientists belonging to the Foundation for Blood Research (Scarborough, Maine, 04074, USA) have recently published a large number of TTR results recorded
in 68,720 healthy US citizens aged 0-100 yr which constitute a comprehensive reference material to follow the shape of LBM fluctuations in relation with sex and age [29].
- TTR concentrations plotted against Hcy values reveal a strongly negative correlation (r = –0.71) [29,30], confirming that
- the depletion of TBN and TBS stores plays a predominant role in the development of HHcy states.
The body of a reference man weighing 70 kg contains 64 M of N (1,800 g) and 4,400 mM of S (140 g) [36]. Our vegetarian subjects consume diets providing
low fat and high fiber content conferring a large spectrum of well described health benefits notably for the prevention of several chronic disorders such as
cancer and diabetes, together with an effective protection against the risk of hypercholesterolemia-induced CVD [37,38].
Plant-based regimens, however, do not supply appropriate amounts of
- nitrogenous substrates of good biological value which are required to adequately fulfill mammalian tissue needs [30].
- vegetable items contain suboptimal concentrations of both SAAs [33,34,39] below the customary RDA guidelines.
This dietary handicap may be further deteriorated by
- unsuitable food processing [40] and by
- the presence in plant products of naturally occurring anti-nutritional factors
- such as tannins in cereal grains and
- anti-trypsin or anti-chymotrypsin inhibitors in soybeans and kidney beans [41].
LBM loss
LBM shrinking may be the result of either
- dysmaturation of body protein tissues as an effect of protracted dietary SAA deprivation
- or of cytokine-induced depletion of body stores.
Although causally unrelated and evolving along dissimilar adaptive processes,
- both physiopathologic entities lead to comparable LBM downsizing best
- identified by declining plasma TTR ( measured alone or within combined formulas )
- and subsequently rising Hcy values.
All parameters are downregulated with the sole exception of RM flux rates, indicating that
- maintenance of Met homeostasis remains a high metabolic priority in protein-depleted states.
Stressful disorders are characterized by
- TM
- RM
- TS flux rates.
The severity and duration of initial impact determine the magnitude of protein tissue breakdown,
- rendering an account of N : S urinary losses,
- fluctuations of albuminuria and of
- insulin resistance striving to contain LBM integrity.
Both physiopathologic entities are compromized in reducing the oxidative burden imposed by HHcy states owing to
- defective synthesis and/or
- enhanced overconsumption of Cys-GSH-H2S reducing molecules,
- a condition still worsened by its co-existence with elemental S-deficiency.
IMPAIRMENT OF THE TRANSSULFURATION PATHWAY
The hypothesis that subclinical protein malnutrition might be involved in the occurrence of HHcy states via inhibition of cystathionine-b-synthase (CbS) activity
first arose in Senegal in 1986 [42] and was later corroborated in Central Africa [43]. The concept was clearly counterintuitive in that it was unexpected that
- high Hcy plasma values might result from low intake of its precursor Met molecule.
Despite the low SAA intake of our vegetarian patients [27], plasma Met concentrations disclosed noticeable stability permitting
- maintenance of the synthesis and functioning of myriads of Met dependent molecular, structural and metabolic compounds
These clinical investigations have received strong support from recent mouse [45] and rat [46] experiments submitted to Met-restricted regimens.
At the end of the Met-deprivation period, both animal species did manifest meaningful HHcy states (p<0.001) contrasting with
- significantly lower BW (p<0.001) reduced by 33 % [45] and 44 % [46] of control, respectively.
- the uniqueness of Met behavior stands in accordance with balance studies performed on large mammalian species showing
- that the complete withdrawal of Met from otherwise normal diets causes the greatest rate of body loss,
- nearly equal to that generated by protein-free regimens [47,48].
This efficient Met homeostatic mechanism is classically ascribed to a PLP-like inhibition of CbS activity exerted through
- allosteric binding of S-adenosylmethionine (SAM) to the C-terminal regulatory domain of the enzyme [49,50].
The loss of CbS activity may develop via a (post)translational defect
- independently from intrahepatic SAM concentrations [45].
We have postulated the existence of an independent sensor mechanism set in motion by TBS pool shrinkage and
- reduced bioavailability of Met – its main building block – working as an inhibitory feedback loop of CbS activity [30].
Such Met-bodystat, likely to be centrally mediated, is to maintain unaltered Met disposal in conditions of
- decreased dietary provision implies the fulfillment
- of high metabolic priorities of survival value [30,44].
Whereas HHcy may be regarded as the dark side of a beneficial adaptive machinery [43],
- impairment of the TS pathway also depresses the production of compounds situated downstream to the CbS blockade level,
- notably Cys and GSH, keeping in mind that Cys may undergo reversible GSH conversion (Fig. 1).
The plasma concentration of both Cys and GSH reductants is indeed significantly decreased in our vegetarian subjects
- by 33 % and 67 % of control, displaying negative correlations (r = –0.67 and –0.37, respectively) with HHcy values [27].
Reduced dietary intake of the preformed Cys molecule [27] and diminished Cys release from protein breakdown in malnourished states [51]
- may contribute to the lowering effect.
The significantly decreased GSH blood levels may similarly be attributed to dietary composition since the tripeptide is mainly found in meat products
- but is virtually absent from cereals, roots, milk and dairy items [52] and
- because regimens lacking SAAs may lessen the production of blood GSH and its intrahepatic sequestration [53].
BIOGENESIS OF HYDROGEN SULFIDE
The TS degradation pathway schematically proceeds along two main PLP-dependent enzymatic reactions working in succession (Fig. 1).
- The first is catalyzed by CbS (EC 4.2.1.22) governing the replacement of the hydroxyl group of serine with Hcy to generate Cysta plus H2O.
- Cys may however substitute for serine and the replacement of its sulfhydryl group with Hcy releases Cysta and H2S instead of water [54].
- The second is regulated by cystathionine-g-lyase (CgL, EC 4.4.1.1.) hydrolyzing Cysta to release Cys and alpha-ketobutyrate plus ammonia as side-products [55].
- Cys may also undergo nonoxidative desulfuration pathways leading to H2S or sulfanesulfur production [56] under the control of CbS or CgL enzymes.
- Cys may otherwise undergo oxidative conversion regulated by cysteine-dioxygenase (CDO, EC 1.13.11.20) which
- catalyzes the replacement of the SH- group of Cys by SO3 – to yield cysteine-sulfinate [56].
This last compound may be further decarboxylated to hypotaurine that is finally oxidized to Tau (67 %) and SO4 2- oxyanions (33 %) [56]. CbS and CgL, both cytosolic enzymes,
- their relative contribution to the generation of H2S may vary according to
- animal strains,
- tissue specificities and
- nutritional or physiopathological circumstances [23,24].
CbS and CgL are expressed in most organs such as liver, kidneys, brain, heart, large vessels, ileum and pancreas [57,58] potentially
- subjected to HHcy-induced ROS injury while keeping the capacity to desulfurate Cys and to
- locally produce H2S as cytoprotectant signaling agent.
CbS is the principal TS enzyme found in
- cerebral glial cells and astrocytes [59].
CgL predominates in the
- vascular system [60] whereas
- both enzymes are present in the renal proximal tubules [61].
H2S is the third gaseous substrate found in the biosphere [18] after NO and CO. All three gases are characterized by
- severe toxicity when inhaled at high concentrations.
In particular, H2S produced by anaerobic fermentation is
- capable of causing respiratory death by
- inhibition of mitochondrial cytochrome C oxidase [62].
NO, CO and H2S are synthesized from arginine, glycine and Cys, respectively, exerting at low concentrations major biological functions in living organisms.
Most of our knowledge on these atypical signal messengers [63] are derived from animal experiments and tissue cultures. These transmitter molecules may
- share some properties in common such as penetration of cellular membranes independently from specific receptors [64].
They are also manifesting dissimilar activities: whereas NO and CO activate guanylyl cyclase to generate biological responses via cGMP-dependent kinases,
- H2S induces Ca2+-dependent effects through ATP-sensitive K+ channels [65].
Some of these potentialities may work in concert while others operate antagonistically. For instance,
- NO and H2S express vasorelaxant tone on endogenous smooth muscle [66]
- but reveal different effects on large artery vessels [67].
These gaseous substances maintain whole body homeostasis through complex interactions and multifaceted crosstalks between signaling pathways.
Elemental S (32.064 as atomic mass) is a primordial constituent of lava flows in areas of volcanic or sedimental origin usually presenting as crown-shaped
stable octamolecules – hence its S8 symbolic denomination – which may conglomerate to form brimstone rocks. The vegetable kingdom is
- unable to assimilate S8 and requires as prior step its natural or bacterial oxidation to SO4 2- derivatives before launching
- the synthesis of SAA molecules along narrowly regulated metabolic pathways [30,44].
Distinct anabolic processes are identified in mammalian tissues which lack the enzymatic equipment required to organize sulfate oxyanions
- but possess the capacity of direct S8 conversion into H2S.
S8 is poorly soluble in tap waters [68] may be taken up and transported to mammalian tissues loosely fastened to serum albumin (SA) [69].
S may also be covalently bound to intracellular S-atoms taking the form of sulfane-sulfur compounds [70] either
- firmly attached to cytosolic organelles or in
- untied form to mitochondria [57,58,71,72] to undergo
- later release in response to specific endogenous requirements [71].
Sulfane-sulfur compounds are somewhat unstable and may decompose in the presence of reducing agents allowing the restitution of S [70,71].
S may either endorse the role of stimulatory factor of several mammalian apoenzyme activities as shown for
- succinic dehydrogenase [73] and NADH dehydrogenase [74] or
- operate as inhibitory agent of other mammalian apoenzymes such as
- adenylate kinase [75] and liver tyrosine aminotransferase [76].
Elemental S resulting from dietary supply or from sulfane-sulfur decay may be subjected to
non-enzymatic reduction in the presence of Cys and GSH [25,26] and/or reducing equivalents obtained from
- glucose oxidation [25], hence yielding at physiological pH additional provision of H2S.
The gaseous mediator is a weakly acidic molecule endowed with strong lipophilic affinities. In experimental models, the blockade of the TS cascade
- at CbS or CgL levels significantly depresses or even
- abolishes the vitally required production of Cys
- operating at the crossroad of multiple converting processes (Fig. 1).
Addition of Cys to the incubation milieu
- resumes the generation of H2S [19] in a Cys concentration-dependent manner [77].
The compounds situated downstream both cystathionases in the context of SAA deprivation
- keep their functional potentialities
- but are unable to express their converting Cys – H2S capacities
- in the absence of precursor substrate.
Summing up
inhibition of CbS activity contributes to
- promote efficient RM processes and
- maintenance of Met homeostasis
but entails as side-effects
- upstream sequestration of Hcy molecules in biological fluids
- while decreasing the bioavailability of Cys and GSH
- working as limiting factors for H2S production.
These last adverse effects thus constitute the Achilles heel of a remarkable adaptive machinery.
ROLES PLAYED BY HYDROGEN SULFIDE
The first demonstration that human tissues may reduce S to H2S was incidentally provided in 1924 when a man given colloid sulfur
- for the treatment of polyarthritis did rapidly exhale the typical rotten egg malodor [78].
- H2S may be produced by the intestinal flora [79] and serves as a metabolic fuel for colonocytes [80].
- Prevention of endogenous poisoning by excessive enteral production is insured by the detoxifying activities of mucosal cells [81],
- hindering any systemic effect of the gaseous substrate.
The normal H2S concentration measured in mammalian plasmas usually ranges from 10 to 100 μM with a mean average turning around 40-50 μM [19,21,82,83].
This H2S plasma level, appearing as the net product of organs possessing CbS and CgL enzymes and supplemented by the non-enzymatic conversion of S,
- flows transiently into the vasculature and freely penetrates into all body cells.
- Supposing that the gaseous reductant is evenly distributed in total body water (45 L in a 70 kg reference man) allows an estimate of
- bioavailable H2S pool turning around 2 mM which represents, in terms of S participation, largely less than 1 / 1,000 of TBS.
The peculiar adaptive physiology of vegetarian subjects renders very unlikely that their TBS pool might be solicited to release
- S-substrates prone to undergo conversion to nascent H2S molecules since
- they adapt to declining energy and nutrient intakes
- by switching overall body economy toward downregulated steady state activities.
The release from TBS of substantial amounts of S-compounds occurs
- only during the onset of hypercatabolic states as documented in trauma patients [31]
- and in infectious diseases [84], exacting as preliminary step
- cytokine-induced breakdown of tissue proteins, a selective hallmark of stressful disorders [85].
H2S in fulfilling ROS Scavenger Tasks
The limited disposal of H2S endogenously produced might be readily exhausted in fulfilling ROS scavenging tasks at the site of oxidative lesions.
All body organs generating H2S from TS enzymes are
- simultaneously producers and consumers of the gaseous substrate whose actual concentration
- reflects the balance between synthetic and catabolic rates [86].
Clinical investigations show that H2S concentrations found in cerebral homogenates from Alzheimer’s disease (AD) patients are
- very much lower than expected from values measured in healthy brains [87], suggesting that
- the gaseous messenger is locally submitted to enhanced consumption rates reflecting disease severity.
The concept is strongly supported by studies pointing to the
- negative correlation linking the severity of AD to H2S plasma values [88].
- in pediatric [89] and elderly [90] hypertensive patients as well
- more severe HHcy-dependent oxidative burden is
- associated with more intense H2S uptake rates.
- These H2S cleansing properties are mainly exerted by mitochondrial organelles
- known to be centrally involved in oxidative disorders [20,91].
Malnourished subjects deprived of Cys and GSH disposal thus incur the risk of H2S-deficiency
- rendering them unable to properly overcome HHcy-imposed oxidative lesions.
The rapid exhaustion of H2S stores have detrimental consequences as shown disclosing
- the beneficial effects of exogenous administration of commonly used sulfide salt donors (Na2S and NaHS)
- generating H2S gas once in solution.
Such supply significantly augments
- H2S plasma concentrations allowing to counteract ROS damages.
H2S was primarily recognized as a physiological substrate working as
- neuromodulator [92] and soon later as
- vasorelaxant factor [65].
H2S is now regarded as endowed with a broader spectrum of biological properties [18],
- operating as a general protective mediator
- against most degenerative organ injuries,
- being capable of neutralizing or
- abolishing most ROS harmful effects.
Table 1 collects findings displaying that H2S may promote the synthesis and activity of several
- anti-oxidative enzymes (catalases, Cu- and Mn-superoxide dismutases, GSH-peroxidases) and
- stimulate the production of anti-inflammatory reactants (interleukin-10) or
- conversely downregulate
- pro-oxidative enzymes (collagenases, elastases),
- pro-inflammatory cytokines (interleukine-1b, tumor-necrosis factor a) and
- immune reactions (hyperleukocytosis, diapedesis, phagocytosis).
It has been calculated that 81.5% of H2S undergoes catabolic disintegration in the form of hydrosulfide anion (HS-) or sulfide anion (S2-) [117].
Since S is the main element in the diprotonated H2S molecule (34.08 as molecular mass), it may be considered that
- partial or complete repair of HHcy-induced lesions constitutes the therapeutic proof that
- S-deficiency is causally involved in the development of ROS damages.
The concept is sustained by the observation that all synthetic drugs (diclofenac, indomethacine, sildenafil) utilized as surrogate providers of H2S [64,118] are
- characterized by a large diversity of molecular conformations but
- share in common the presence of Satom(s) mimicking, once released,
- H2S-like pharmacological properties.
It remains to be clarified whether the beneficial effects of S-fortification to S-deficient subjects are mediated, among other possible mechanisms, via
- stimulation [73,74] of anti-oxidative enzymes or inhibition [75,76] of pro-oxidative enzymes.
It is only very recently that the essentiality of S has been recognized, causing Hcy elevation in deficient individuals [119]. It is worth reminding that the
- gaseous NO substrate may work in concert or antagonistically [66,83] to fine-tuning the helpful properties exerted by H2S on body tissues.
Preliminary studies suggest for instance that NO operates, in combination with H2S, as a potential modulator of endothelial remodeling since
- NO-synthase isoforms contribute to the activation of metalloproteinases involved in the regulation of the collagen/elastin balance defining vascular elastance [83,120].
SUBCLINICAL MALNUTRITION AS WORLDWIDE SCOURGE
A growing body of data collected along the last decades indicates that
large proportions of mankind still suffer varying degrees of protein and energy deficiency that is associated with
- increased morbidity and mortality rates.
The determinants of malnutrition are complex and interrelated, comprising
- socioeconomic and political conditions,
- insufficient dietary intakes,
- inadequate caring practices and
- superimposed inflammatory burden.
Children living in developing countries are paying a heavy toll to chronic malnutrition [121,122] whereas adult populations are handicapped by
- feeble physical and working capacities,
- increased vulnerability to infectious complications and
- reduced life expectancy [123,124].
Cross-sectional studies collected in the eighties indicate that chronic malnutrition remains a worldwide scourge with
- top prevalence recorded in Asia, whereas
- sub-Saharan Africa endures medium nutritional distress and
- Latin America appears as the least affected [125,126].
Along the last decades, significant progresses have been achieved in some countries such as Vietnam [127] and Bangladesh [128]
- owing to appropriate education programs and improved economic development.
Inequalities however persist between middle class population groups mainly located in affluent urban areas and
- underprivileged rural communities remaining stagnant on the sidelines of household income growth.
Representative models of these socio-economic disparities in global nutrition and health are illustrated in the two most populated countries in the world, China and India.
Large surveys undertaken in 105 counties of China and recently published have concluded that the rural communities haven’t yet reached the stage of overall welfare [129].
In India, similar investigations have documented that extreme poverty still prevails in the northern mountainous states of the subcontinent [130]. Taken together, southern
Asian countries fail to overcome malnutrition burden [131]. In some African countries, there exists even upward trends suggesting nutritional
deterioration over the years [132] still aggravated by a severe drought. The assessment of malnutrition in children usually rely on anthropometric criteria such as height-for-age, weight for-height, mid upper arm circumference and skinfold thickness allowing to draw the degree of stunting and wasting from these estimates. In adult subjects, BW and BMI are currently selected parameters to which some biochemical measurements are frequently added, notably SA, classical marker of protein nutritional status, and creatininuria (u-Cr), held as indicator of sarcopenia. The former biometric approaches are very useful in that they correctly provide a static picture of the declared stages of malnutrition but fail to recognize the dynamic mechanisms occurring during the preceding months and the adaptive alterations running behind.
Table 1. Reversal of HHcy-Induced Oxidative Damages by Administration of Exogenous H2S
BRAIN EFFECTS
H2S is overproduced in response to neuronal excitation [93], and
- increases the sensitivity of N-methyl-D-aspartate (NMDA) reactions to glutamate in hippocampal neurons [23,94].
- improves long-term potentiation, a synaptic model of memory [92,93]
- stimulates the inhibitory effects of catalase and superoxide dismutase (SOD) in oxidative stress of endothelial cells [95].
- regulates Ca 2+ homeostasis in microglial cells [96]and it inhibits TNFa expression in microglial cultures [97].
- protects brain cells from neurotoxicity by preventing the rise of ROS in mitochondria [98].
CARDIOVASCULAR EFFECTS
- H2S releases vascular smooth muscle,
- inhibits platelet aggregation and
- reduces the force output of the left ventricule of the heart [18].
- maintains vascular smooth muscle tone [66] and
- insures protection against arterial hypertension [99].
- modifies leucocyte-vascular epithelium interactions in vivo by
- modulating leucocyte adhesion and
- diapedesis at the site of inflammation [100].
- attenuates myocardial ischemia-reperfusion injury by
- depressing IL-1b and mitochondrial function [20].
- upregulates the expression of depressed anti-oxidative enzymes in heart infarction and
- inhibits myocardial injury [21].
- alleviates smooth muscle pain by
- stimulating K+ ATP channels [101].
- prevents apoptosis of human neutrophil cells
- by inhibiting p38 MAP kinase and caspase 3 [102].
- potentiates angiogenesis and wound healing [103].
RENAL EFFECTS
- H2S downregulates the increased activity of metalloproteinases 2 and 9 involved in extracellular matrix degradation (elastases, collagenases) [19].
- Prevents apoptotic cell death in renal cortical tissues [19].
- Improves the expression of desmin (marker of podocyte injury) and
- restores the drop of nephrin (component of normal slit diaphragm) in the cortical tissues
- resulting in reduced proteinuria [19].
- Induces hypometabolism revealing protective effects on renal function and survival [104].
- Normalizes GSH status and production of ROS in renal diseases [19].
- Controls renal ischemia-reperfusion injury and dysfunction [105].
- Depresses the expression of inflammatory molecules involved in glomerulosclerosis [106].
- Increases renal blood flow, glomerular filtration and urinary Na+ excretion [77].
OTHER ORGAN EFFECTS
Gastrointestinal
- H2S insures protection against ROS stress in gastric mucosal epithelia [22].
- Accelerates gastric ulcer healing [107].
- Reduces gastric injury caused by nonsteroidal anti-inflammatory drugs [108].
- Relaxes ileal smooth muscle tone and increases colonic secretions [79].
- Attenuates intestinal ischemia-reperfusion injury by increasing SOD and GSH peroxidase status [109].
- Stimulates insulin secretion [110] and controls inflammatory events associated with acute pancreatitis [111].
- Alleviates hepatic ischemia-reperfusion injury [112].
Pulmonary
- Prevents lung oxidative stress in hypoxic pulmonary hypertension caused by low GSH content [113].
- Promotes SOD and catalase activities and reduces the production of malondialdehyde in oxidative lung injury [114].
- Reduces lung inflammation and remodeling in asthmatic animals [115] and in pulmonary hypertension [116]. ..(see OCCJ 2011;4:34-44)
Assessing Protein-Depleted States
- SA is an insensitive marker of protein-depleted states compared to TTR [134]
- SA is an indicator of population than of individual protein status in subclinical PEM.
- u-Cr is likewise a meagerly informative tool as 10 % loss of muscle mass is required before it reaches significantly decreased urinary concentrations [135].
The data imply that the magnitude of subclinical malnutrition is largely
- underscored when classical biometric and laboratory investigations are performed.
Moreover, ruling out the protein component involved in HHcy epidemiology and confining solely attention to the B-vitamin triad led to unachieved conclusions.
- surveys undertaken in Taiwan [136] and in India [137] established HHcy variance turning around 30 %, indicating that
- a sizeable percentage of subjects do not come within the vitamin shortage concept.
- only one recent review recommending the use of TTR in vegetarian subjects [138].
The main reason for making the choice of TTR is grounded on the striking similar plasma profile disclosed by this marker with both LBM and Hcy [29].
Under healthy conditions, the 3 parameters –
- TTR,
- LBM,
- Hcy –
- indeed show low concentrations at birth,
- linear increase without sexual difference in preadolescent children,
- gender dimorphism in teenagers with higher values recorded in adolescent male subjects
- thereafter maintenance of distinct plateau levels during adulthood [29,139,140].
Under morbid circumstances, the plasma concentrations of
- Hcy manifest gradual elevation
- negatively correlated with LBM downsizing and
- TTR decline.
In vegetarian subjects and subclinically malnourished patients,
- rising Hcy and
- diminished TTR plasma concentrations look as mirror image of each other,
- revealing divergent distortion from normal and
- allowing early detection of preclinical steps
- at the very same time both SA and u-Cr markers still remain silent.
Any disease process characterized by quantitative or qualitative dietary protein restriction or intestinal malabsorption
- may cause LBM shrinking,
- downregulation of TTR concentrations and
- subsequent HHcy upsurge.
These conditions are documented in frank kwashiorkor [141], subclinical protein restriction [27,43] and anorexia nervosa [142].
In patients submitted to weight-reducing programs,
- LBM was found the sole independent variable
- negatively correlated with rising Hcy values [143].
Morbid obesity may be alleviated by medical treatment [143] or surgical gastroplasty [144,145],
- conditions frequently associated with secondary malabsorptive syndromes and malnutrition [146],
How does this account play out in the typical patient with excessive body fat, lipoprotein disoreder, and perhaps diabetes and disordered sleep – an account of acquired HHcy?
Have the studies been done? Would you expect to see a clear benefit from reduced HHcy_emia based on a 30 min daily walk, and
- eating of well fat trimmed meats, fruits and vegetables, and fish, flax seed, or krill oil?
In westernized countries, subclinical protein-depleted states are illustrated in immigrants originating from
- developing regions but keeping alive their traditional feeding practices [147] or
- by communities having adopted, for socio-cultural reasons, strict vegan dietary lifestyles [148].
THE ADDITIONAL BURDEN OF S-DEFICIENCY
After N, K and P, elemental S is recognized as the fourth most important macronutrient required for plant development. The essentiality of S in the vegetable kingdom
arose from observations made many decades ago by pedologists and agronomists [149,150] revealing that the withdrawal of sulfate salts from nutrient sources produces
rapid growth retardation,
- depressed chlorophyllous synthesis,
- yellowing of leaves and
- reduction in fertility and crop yields.
A large number of field studies, mainly initiated for economical reasons, has provided continuing gain in fundamental and applied knowledge and led to the overall consensus
- that SO4 2- -deficiency is a major wordwide problem [151,152].
Field investigations have shown that the concentration of SO4 2- oxyanions in soils and drinking waters
- may reveal considerable variations ranging from less than 2 mg/L to more than 1 g/L,
- meaning a ratio exceeding 1 / 500 under extreme circumstances [30].
The main causal factors responsible for unequal distribution of SO4 2- oxyanions are geographical distance from eruptive sites and
- intensity of soil weathering in rainy countries.
SO4 2- -dependent nutritional deficiencies entail detrimental effects to most African and Latin American crops [151]
- reaching nevertheless top incidence in southeastern Asia [151,153].
- and the Indo-Gangetic plain extending from Pakistan to Bangladesh and covering the North of India and Nepal [154].
- Intensive agricultural production,
- lack of animal manure and
- use of fertilizers providing N, K and P substrates
- but devoid of sulfate salts may further aggravate that imbalanced situation.
As global population increases steadily and the production of staple plants predicted to escalate considerably,
- SO4 2- deficient disorders are expected to become more pregnant along the coming years [155] with significant harmful impact for mankind.
Nevertheless, effective preventive efforts are developed in some countries aiming at fortification of soils mainly
- by ammonium sulfate or calcium sulfate (gypsum) salts,
- resulting in meaningful improvements in crop yield,
- SAAs content and biological value and
- opening more optimistic perspectives for livestock and human consumption [152,155-158].
Contrasting with the tremendously high amounts of data accumulated over decades by pedologists and agronomists on sulfate requirements and metabolism,
the available knowledge on elemental sulfur in human nutrition looks like a black hole. Despite the fact that S8 follows H, C, O, N, Ca and P as the seven most
abundant element in mammalian tissues, it appears as a forgotten item. Not the slightest attention is dedicated to S8 in the authoritative “Present Knowledge
in Nutrition” series of monographs even though they go over most oligo- and trace-elements in minute detail.
The geographical distribution of S8 throughout the earth’s crust is not well-known
- as extreme paucity of measurements in soils and tap waters prevents reaching a comprehensive overview.
Nevertheless, and because S8 is the obligatory precursor substrate for the oxidative production of sulfate salts,
a decremental dispersion pattern paralleling those of SO4 2- oxyanions is likely to occur with
- highest values recorded in the vicinity of volcano sources
- and lowest values found in remote and washed-out areas.
Obviously, a great deal of research on elemental S remains to be completed by clinical biochemists before rejoining the status of plant agronomy.
Taken together, these data imply that subclinically malnourished subjects living in areas recognized as
- SO4 2- -deficient for the vegetable kingdom also
- incur increased risks to become S8-depleted.
This clinical entity most probably prevails in all regions, notably Northern India, where protein malnutrition [130] and sulfur-deficiency [154] coexist.
Combination of both nutritional deprivations explains why the bulk of local dwellers, including young subjects [159,160], may develop HHcy states and CVD disorders
- characterized by strong refractoriness to vitamin-B supplementation [160] or
- high incidence of stroke [161] unrelated to the classical Framingham criteria.
The current consensus is that “the problem of CVD in South Asia is different in etiology and magnitude from other parts of the world” [162]. These disquieting findings are
confirmed in several Asian countries [163] and have prompted local cardiologists to exhort their governments to focus more attention on CVD epidemiology [164].
CONCLUDING REMARKS
- vegetarian subjects are not protected against the risk of CVD and stroke which should no longer be regarded as solely affecting populations living in westernized societies
- whose morbidity and mortality risks are stratified by classical Framingham criteria.
- Likewise hypercholesterolemia, hyperhomocysteinemia should be incriminated as
- emblematic risk factor for a panoply of CVD and related disorders.
- Whereas the causality of cholesterol and lipid fractions largely prevails in affluent societies consuming high amounts of animal-based items,
- that of homocysteine predominates in population groups whose dietary lifestyle gives more importance to plant products.
|
MAIN PHYSICO-CHEMICAL AND METABOLIC CHARACTERISTICS* OF 3 CARRIER-PROTEINS INVOLVED IN THE STRESS RESPONSE
|
|
CBG
|
TTR
|
RBP
|
| Molecular mass (Da.) |
42,650
|
54,980
|
21,200
|
| Conformation |
monomeric
|
tetrameric
|
monomeric
|
| Amino acid sequence |
383
|
4 x 127
|
182
|
| Carbohydrate load |
18 % glycosylated
|
unglycosylated
|
unglycosylated
|
| Hormonal binding sites |
one for cortisol
|
two for TH
|
one for retinol
|
| Association constant (M-1) |
3 x 107
|
7 x 107 (T4)
|
1.9 x 107
|
| Normal plasma concentration |
30 mg/L.
|
300 mg/L.
|
50 mg/L.
|
| Biological half-life |
5 days
|
2 days
|
14 hrs
|
| Bound ligand concentration |
120 µg/L.
|
80 µg TT4/L.
|
500 µg/L.
|
| Free ligand concentration |
5 µg/L.
|
20 ng FT4/L.
|
1 µg/L.
|
| Ratio free : bound ligands |
4 %
|
0.034 %
|
0.14 %
|
| Distribution volume of free moieties |
18 L.
|
12 L.
|
18 L.
|
|
STIMULATORY AND INHIBITORY EFFECTS MODULATED
BY GLUCOCORTICOIDS
|
|
TARGET SYSTEMS
|
|
INDUCED EFFECTS
|
REF.
|
| Thymidine kinase |
_
|
transcription of induced DNA into RNA |
112
|
| Alkaline phosphodiesterase I |
_
|
cleavage of phosphodiester bonds |
113
|
| Tyrosine transaminase |
_
|
transfer of tyrosine amino group |
114
|
| Tryptophane oxygenase |
_
|
formylkynurenine and Trp catabolites |
115
|
| Alkaline phosphatase |
_
|
release of P from phosphoric esters |
116
|
| Phosphoenolpyruvate carboxykinase (liver) |
_
|
glycolysis from pyruvate and ATP production |
117
|
| Mannolsyltransferases |
_
|
dolichol-linked glycosylation of APRs |
118
|
| Haptoglobin |
_
|
APR combining with hemoglobin |
119
|
| α1-Anti (chymo) trypsin (α1 AT, α1 ACT) |
_
|
serpin molecules allowing N-sparing effects |
120
|
| α1-Acid glycoprotein (AGP) |
_
|
glycosylated APR with antibody-like actions |
121
|
| Serum amyloid protein (SAA) |
_
|
defense systems against oxidative burst |
122
|
| γ-Fibrinogen |
_
|
clotting processes and tissue repair |
123
|
| C-Reactive Protein (CRP) |
_
|
complement processes and opsonization |
124
|
| Corticosteroid-binding globulin (CBG) |
_
|
CBG levels, favoring free hypercortisolemia |
100
|
| Phosphoenolpyruvate carboxykinase (adipocytes) |
_
|
ATP turnover and glycolysis |
113
|
THE DUAL MORBID ENTITIES CAUSING LBM DOWNSIZING AND SUBSEQUENT Hcy UPSURGE
Primary causal factor
- Reduced dietary intake of methionine (39,151,152)
- Cytokine-induced tissue breakdown (164,165)
Main clinical conditions
- Protein malnutrition,
- veganism,
- intestinal malabsorption (139,155,156,158-160,281)
- Trauma,
- sepsis,
- burns,
- Inflammatory & neoplastic disorders (163,166,170,176,179,180)
Physiopathologic mechanisms
- Unachieved LBM replenishment (30,33)
- Excessive LBM losses (33,167,179)
Overall protein metabolic status
- Downregulated
- Upregulated
Plasma biomarker(s) of protein status
- Transthyretin (TTR) (144,145)
- TTR coupled with CRP or other inflammatory indices (31,177,178,284,285)
Insulin resistance status
- Normal or low (286)
- Increased in proportion of tissue breakdown (177,178,181-183)
status of Cys-GSH-H2S reducing molecules
- Decreased enzymatic and non-enzymatic production (39,161,162,287)
- Increased production cancelled out by tissue overconsumption (78,171)
Urinary SO42- and S-compounds
- Decreased kidney output (76,78,79)
- Variable depending on exogenous SAA supply and
- extent of tissue breakdown (78,163,168,173)
Transmethylation pathway
- Depressed (48,93)
- Overstimulated (169)
Remethylation pathway
- Stimulated (76,83,153)
- Overstimulated (169)
Transsulfuration pathway
- Inhibited (49,76,83)
- Overstimulated (170,173)
REFERENCES
[1] McCully, K.S. Vascular pathology of homocysteinemia: implications
for the pathogenesis of arteriosclerosis. Am. J. Pathol., 1969,
56, 111-128.
[2] Ubbink, J.B.; van der Merwe, A.; Delport, R.; Allen, R.H.; Stabler,
S.P.; Riezler, R.; Vermaak, W.J. The effect of subnormal vitamin
B-6 status on homocysteine metabolism. J. Clin. Invest., 1996, 98,
177-184.
[4] Stabler, S.P.; Allen, R.H.; Savage, D.G.; Lindenbaum, J. Clinical
spectrum and diagnosis of cobalamin deficiency. Blood, 1990, 76,
871-881.
[6] Cheng, Z.; Yang, X.; Wang, H. Hyperhomocysteinemia and endothelial
dysfunction. Curr. Hypertens. Rev., 2009, 5,158-165.
[7] Loscalzo, J. The oxidant stress of hyperhomocyst(e)inemia. J. Clin.
Invest., 1996, 98, 5-7.
[8] Jacobsen, D.W. Hyperhomocysteinemia and oxidative stress: Time
for a reality check ? Arterioscler. Thromb. Vasc. Biol., 2000, 20,
1182-1184.
[10] McCully, K.S. Chemical pathology of homocysteine: IV. Excitotoxicity,
oxidative stress, endothelial dysfunction, and inflammation.
Annals Clin. Lab. Sci., 2009, 39, 219-232.
[11] Bautista, L.E.; Arenas, I.A.; Penuela, A.; Martinez, I.X. Total
plasma homocysteine level and risk of cardiovascular disease: a
meta-analysis of prospective cohort studies. J. Clin. Epidemiol.,
2002, 55, 882-887.
[12] Furie, K.L.; Kelly, P. J. Homocyst(e)ine and stroke. Semin. Neurol.,
2006, 26, 24-32.
[13] Van Guldener, C. Homocysteine and the kidney. Curr. Drug Metab.,
2005, 6, 23-26.
[14] McCaddon, A.; Davies, G.; Hudson, P.; Tandy, S.; Cattell, H. Total
serum homocysteine in senile dementia of Alzheimer type. Int. J.
Geriatr. Psychiatry 1998, 13, 235-239.
[15] Troen, A.M.; Rosenberg, I.H. Homocysteine and cognitive function.
Semin. Vasc. Med., 2005, 5, 209-214.
[16] Danese, S.; Sgambato, A.; Papa, A.; Scaldaferri, F.; Pola, R.; Sans,
M.; Lovecchio M. Homocysteine triggers mucosal microvascular
activation in inflammatory bowel disease. Am. J. Gastroenterol.,
2005, 100, 886-895.
[20] Elrod, J.W.; Calvert, J.W.; Morrison, J.; Doeller, J.E.; Kraus, D.W.;
Tao, L.; Jiao, X.; Scalia, R.; Kiss, L.; Szabó, C.; Kimura, H.;
Chow, C.W.; Lefer, D.J. Hydrogen sulfide attenuates myocardial
ischemia-reperfusion injury by preservation of mitochondrial function.
Proc. Natl. Acad. Sci. USA, 2007, 104, 15560-15565.
[21] Chang, L.; Geng, B.; Yu, F.; Zhao, J .; Jiang, H.; Du, J.; Tang C.
Hydrogen sulfide inhibits myocardial injury induced by homocysteine
in rats. Amino Acids, 2008, 34, 573-585.
[22] Yonezawa, D.; Sekiguchi, F.; Miyamoto, M.; Taniguchi, E.; Honjo,
M.; Masuko, T.; Nishikawa, H.; Kawabata, A. A protective role of
hydrogen sulfide against oxidative stress in rat gastric mucosal epithelium.
Toxicology, 2007, 241, 11-18.
[23] Dominy, J.E.; Stipanuk, M.H. New roles for cysteine and transulfuration
enzymes: production of H2S, a neuromodulator and smooth
muscle relaxant. Nutr. Rev. 2004, 62, 348-353.
[27] Ingenbleek, Y.; McCully, K. Vegetarianism produces subclinical
malnutrition, hyperhomocysteinemia and atherogenesis. Nutrition.
Doi:10.1016/j.nut.2011.04.009: on line 27th August 2011.
[28] Battezzatti, A.; Bertoli, S.; San Romerio, A.; Testolin, G. Body
composition: an important determinant of homocysteine and methionine
concentrations in healthy individuals. Nutr. Metab. Cardiovasc.
Dis., 2007, 17, 525-534.
[29] Ingenbleek, Y. Plasma Transthyretin Reflects the Fluctuations of
Lean Body Mass in Health and Disease. In: Recent Advances in
Transthyretin Evolution, Structure and Biological Functions;
Richardson, S.J. and Cody, V., Eds.; Springer Verlag : Berlin,
2009, pp. 329-357.
[30] Ingenbleek, Y. The nutritional relationship linking sulfur to nitrogen
in living organisms. J. Nutr., 2006, 136, S1641-S1651.
[31] Cuthbertson, D.P. The distribution of nitrogen and sulphur in the
urine during conditions of increased catabolism. Biochem. J., 1931,
25, 236-244.
[34] Souci, S.W.; Fachman, W.; Kraut, H. Food Composition and Nutrition
Tables. 5th ed., CRC Press: Boca Raton, 1994.
[35] Ingenbleek, Y.; Young, V.R. Significance of transthyretin in protein
metabolism. Clin. Chem. Lab. Med., 2002, 40, 1281-1291.
[36] Forbes, G.B. Body Composition. In: Present Knowledge in Nutrition;
7th ed.; Ziegler, E.E. and Filer L.J, Eds.; ILSI Press: Washington
D.C. 1996; pp. 7-12.
42 The Open Clinical Chemistry Journal, 2011, Volume 4 Yves Ingenbleek
[37] Walter, P. Effects of the vegetarian diets on aging and longevity.
Nutr. Rev., 1997, 55, S61-S65.
[38] Key, T.J.; Appleby, P.N.; Rosell, M.S. Health effects of vegetarian
and vegan diets. Proc. Nutr. Soc., 2006, 65, 35-41.
[39] Young, V.R.; Pellet, P.L. Plant proteins in relation to human protein
and amino acid nutrition. Am. J. Clin. Nutr., 1994, 59, S1203-
S1212.
[42] Ingenbleek, Y.; Barclay, D.; Dirren, H. Nutritional significance of
alterations in serum amino acid patterns in goitrous patients. Am. J.
Clin. Nutr., 1986, 43, 310-319.
[43] Ingenbleek, Y.; Hardillier, E.; Jung, L. Subclinical protein malnutrition
is a determinant of hyperhomocysteinemia. Nutrition, 2002,
18, 40-46.
[44] Ingenbleek, Y.; Young, V.R. The essentiality of sulfur is closely
related to nitrogen metabolism: a clue to hyperhomocysteinemia.
Nutr. Res. Rev., 2004, 17, 135-153.
[45] Tang, B.; Mustafa, A.; Gupta, S.; Melnyk, S.; James S.J.; Kruger,
W.D. Methionine-deficient diet induces post-transcriptional downregulation
of cystathionine-g-synthase. Nutrition, 2010, 26, 1170-
1175.
[46] Elshorbagy, A.K.; Valdivia-Garcia, M.; Refsum, H.; Smith, A.D.;
Mattocks, D.A.; Perrone, C.E. Sulfur amino acids in methionine restricted
rats: Hyperhomocysteinemia. Nutrition, 2010, 26, 1201-
1204.
[47] Owens, F.N.; Bergen, W.G. Nitrogen metabolism in ruminant
animals: historical perspective, current understanding and future
implications. J. Anim. Sci., 1983, 57 (Suppl. 2), 498-518.
[55] Stipanuk, M.H. Sulfur amino acid metabolism : pathways for production
and removal of homocysteine and cysteine. Annu. Rev.
Nutr., 2004, 24, 539-577.
[56] Stipanuk, M.H.; Ueki, I. Dealing with methionine/homocysteine
sulfur : cysteine metabolism to taurine and inorganic sulfur. J. Inherit.
Metab. Dis., 2011, 34, 17-32.
[57] Kamoun, P. Endogenous production of hydrogen sulfide in mammals.
Amino Acids, 2004, 26, 243-254.
[58] Kimura, H. Hydrogen sulfide : its production, release and functions.
Amino Acids, 2011, 41, 113-121.
[59] Enokido, Y.; Suzuki, E.; Iwasawa, K.; Namekata, K.; Okazawa, H.;
Kimura, H. Cystathionine -synthase, a key enzyme for homocysteine
metabolism, is preferentially expressed in the radial
glia/astrocyte lineage of developing mouse CNS. FASEB J., 2005,
19, 1854-1856.
[60] Zhao, W.; Ndisang, J.F.; Wang, R. Modulation of endogenous
production of H2S in rat tissues. Can. J. Physiol. Pharmacol., 2003,
81, 848-853.
[61] House, J.D.; Brosnan, M.E.; Brosnan, J.T. Characterization of
homocysteine metabolism in the rat kidney. Biochem. J., 1997,
328, 287-292.
[62] Dorman, D.C.; Moulin, F.J.; McManus, B.E.; Mahle, K.C.; James,
R.A.; Struve, M.F. Cytochrome oxidase inhibition induced by acute
hydrogen sulfide inhalation : correlation with tissue sulfide concentrations
in the rat brain, liver, lung, and nasal epithelium. Toxicol.
Sci., 2002, 65, 18-25.
[64] Mancardi, D.; Penna, C.; Merlino, A.; Del Soldato, P.; Wink, D.A.;
Pagliaro, P. Physiological and pharmacological features of the
novel gasotransmitter : Hydrogen sulfide. Biochim. Biophys. Acta,
2009, 1787, 864-872.
[65] Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of
H(2)S as a novel endogenous gaseous K(ATP) channel opener.
EMBO J., 2001, 20, 6008-6016.
[66] Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen
sulfide as an endogenous smooth muscle relaxant in synergy with
nitric oxide. Biochem. Biophys. Res. Commun., 1997, 237, 527-
531.
[75] Russell, P.J.; Conner, J.; Sisson, S. Sulfur specifically inhibits
adenylate kinase in assays for creatine kinase. Clin. Chem., 1984,
30, 1555-1557.
[76] Hargrove, J.L.; Wichman, R.D. A cystine-dependent inactivator of
tyrosine aminotransferase co-purifies with -cystathionase (cysteine
desulfurase). J. Biol. Chem., 1987, 262, 7351-7357.
[77] Xia, M.; Chen, L.; Muh, R.W.; Li, P.L.; Li, N. Production and
actions of hydrogen sulfide, a novel gaseous bioactive substance, in
the kidneys. J. Pharmacol. Exp. Ther., 2009, 329, 1056-1062.
[78] Monaghan, W.J.; Garai, F. Treatment of acute and chronic polyarthritis,
arthritis deformans, and septicemias, with activated colloid
sulphur. Med. J. Record, 1924, 120, 24-26.
[79] Fiorucci, S.; Distrutti, E.; Cirino, G.; Wallace, J.L. The emerging
role of hydrogen sulfide in the gastrointestinal tract and liver. Gastroenterology,
2006, 131, 259-271.
[80] Goubern, M.; Andriamihaja, M.; Nubel, T.; Blachier, F.; Bouillaud,
F. Sulfide, the first inorganic substrate for human cells. FASEB J.,
2007, 21, 1699-1706.
[81] Furne, J.; Springfield, J.; Koenig, T.; DeMaster, E.; Levitt, MD.
Oxidation of hydrogen sulfide and methanethiol to thiosulfate by
rat tissues: a specialized function of the colonic mucosa. Biochem.
Pharmacol., 2001, 62, 255-259.
[82] Chen, YH.; Yao, W.Z.; Geng, B.; Ding, Y.L.; Lu, M.; Zhao, M.W.;
Tang, C.S. Endogenous hydrogen sulfide in patients with COPD.
Chest, 2005, 128, 3205-3211.
[83] Whiteman, M.; Moore, P.K. Hydrogen sulfide and the vasculature:
a novel vasculoprotective entity and regulator of nitric oxide
bioavailability? J. Cell. Mol. Med., 2009, 13, 488-507.
[85] Ingenbleek, Y.; Bernstein, L. The stressful condition as a nutritionally
dependent adaptive dichotomy. Nutrition, 1999, 15, 305-320.
Malnutrition, Sulfur-Deficiency and Hyperhomocysteinemia The Open Clinical Chemistry Journal, 2011, Volume 4 43
[89] Chen, L.; Ingrid, S.; Ding, Y.G.; Liu, Y.; Qi, J.G.; Tang, J.B. Imbalance
of endogenous homocysteine and hydrogen sulfide metabolic
pathway in essential hypertensive children. Chin. Med. J.,
2007, 120, 389-393.
[90] Sun, N.L.; Xi, Y.; Yang, S.N.; Ma, Z.; Tang, C.S. Plasma hydrogen
sulfide and homocysteine levels in hypertensive patients with different
blood pressure levels and complications. Zonghua Xin Xue
Guan Bing Za Zhi, 2007, 35, 1145-1148.
[91] Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative
stress in neurodegenerative diseases. Nature, 2006, 443, 787-795.
[97] Hu, L.F.; Wong, P.T.; Moore, P.K.; Bian, J.S. Hydrogen sulfide
attenuates lipopolysaccharide-induced inflammation by inhibition
of p38 mitogen-activated protein kinase in microglia. J.
Neurochem., 2007, 100, 1121-1128.
[98] Tang, X.Q.; Shen, X.T.; Huang, Y.E.; Ren, Y.K.; Chen, R.Q.; Hu,
B.; He, J.Q.; Yin, W.L.; Xu, J.H.; Jiang, Z.S. Hydrogen sulfide antagonizes
homocysteine-induced neurotoxicity in PC12 cells. Neurosci.
Res., 2010, 68, 241-249.
[99] Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.;
Mustafa, A.K.; Mu, W.; Zhang, S. H2S as a physiologic vasorelaxant
: hypertension in mice with deletion of cystathionine gammalyase.
Science, 2008, 322, 587-590.
[100] P.; Vitale, M. Hydrogen sulfide prevents apoptosis of human PMN
via inhibition of p38 and caspase 3. Lab. Invest., 2006, 86, 391-
397.
[105] Tripatara, P.; Patel, N.S.; Collino, M.; Gallichio, M.; Kieswich, J.;
Castiglia, S.; Benetti, E.; Stewart, K.N.; Brown, P.A.; Yaqoob,
M.M.; Fantozzi, R.; Thiemermann, C. Generation of endogenous
hydrogen sulfide by cystathionine gamma-lyase limits renal ischemia/
reperfusion injury and dysfunction. Lab. Invest., 2008, 88,
1038-1048.
[106] Sen, U.; Munjal, C.; Qipshidze, N.; Abe, O.; Gargoum, R.; Tyagi,
S.C. Hydrogen sulfide regulates homocysteine-mediated glomerulosclerosis.
Am. J. Nephrol., 2010, 31, 442-455.
[114] Fu, Z.; Liu, X.; Geng, B.; Fang, L.; Tang, C. Hydrogen sulfide
protects rat lung from ischemia-reperfusion injury. Life Sci., 2008,
82, 1196-1202.
[115] Chen, Y.H.; Wu, R.; Geng, B.; Qi, Y.F.; Wang, P.P.; Yao, W.Z.;
Tang, C.S. Endogenous hydrogen sulfide reduces airway inflammation
and remodeling in a rat model of asthma. Cytokine 2009, 45,
117-123.
[116] Li, X.; Jin, H.; Bin, G.; Wang, L.; Tang, C.; Du, J. Endogenous
hydrogen sulfide regulates pulmonary artery collagen remodeling
in rats with high pulmonary blood flow. Exp. Biol. Med.
(Maywood), 2009, 234, 504-512.
[117] Dombkowski, R.A.; Russell, M.J.; Olson, K.R. Hydrogen sulfide as
an endogenous regulator of vascular smooth muscle tone in trout.
Am. J. Physiol. Regul. Integr. Comp. Physiol., 2004, 286, R678-
R685.
[118] Wallace, J.L. Hydrogen sulfide-releasing anti-inflammatory drugs.
Trends Pharmacol. Sci., 2007, 28, 501-505.
[119] Ingenbleek, Y. Hyperhomocysteinemia is a biomarker of sulfurdeficiency
in human morbidities. Open Clin. Chem. J., 2009, 2, 49-
60.
[120] Steed, M.M.; Tyagi, S.C. Mechanisms of cardiovascular remodeling
in hyperhomocysteinemia. Antioxid. Redox. Signal., 2011, 15,
1927-1943.
[121] Black, R.E.; Morris, S.S.; Bryce, J. Where and why are 10 millions
children dying every year? Lancet, 2003, 361, 2226-2234.
[122] El-Ghannam, A.R. The global problems of child malnutrition and
mortality in different world regions. J. Health Soc. Policy, 2003,
16, 1-26.
[128] Faruque, A.S.; Shamsir Ahmed, A.M.; Tahmeed Ahmed.; Munirul
M.; Iqbal Hossain, M.; Roy, S.K.; Nurul Alam; Iqbal Kabir; Sack,
D.A. Nutrition : Basis for healthy children and mothers in Bangladesh.
J. Health Popul. Nutr., 2008, 26, 325-339.
[129] Wang, X.; Wang, Y.; Kang, C. Feeding practices in 105 countries
of rural China. Child Care Health Dev., 2005, 31, 417-423.
44 The Open Clinical Chemistry Journal, 2011, Volume 4 Yves Ingenbleek
[130] Antony, G.M.; Laxmaiah, A. Human development, poverty, health
& nutrition situation in India. Indian J. Med. Res., 2008, 128, 198-
205.
[134] Ingenbleek, Y.; De Visscher, M ; De Nayer, P. Measurement of
prealbumin as index of protein-calorie malnutrition. Lancet, 1972,
ii, 106-109.
[135] Shenkin, A.; Cederblad, G.; Elia, M.; Isaksson, B. International
Federation of Clinical Chemistry. Laboratory assessment of protein-
energy status. Clin. Chim. Acta, 1996, 253, S5-S59.
[136] Hung, C.J.; Huang, P.C.; Lu, S.C.; Li, Y.H.; Huang, H.B.; Lin,
B.F.; Chang, S.J.; Chou, H.F. Plasma homocysteine levels in Taiwanese
vegetarians are higher than those of omnivores. J. Nutr.,
2002, 132, 152-158.
[138] Benzie, I.F.; Wachtel-Galor, S. Biomarkers in long-term vegetarian
diets. Adv. Clin. Chem., 2009, 47, 171-222.
[139] Must, A.; Jacques, P.F.; Rogers, G.; Rosenberg, I.H.; Selhub, J.
Serum total homocysteine concentrations in children and adolescents
: results from the third National Health and Nutrition Examination
Survey (NHANES III). J. Nutr., 2003, 133, 2643-2649.
[141] Abdel, G.A.; Abdullah, S.H.; Kordofani, A.Y. Plasma homocysteine
levels in cardiovascular disease, malaria and protein-energy
malnutrition in Sudan. East Mediterr. Health J., 2009, 15, 1432-
1439.
[142] Moyano, D.; Vilaseca, M.A.; Artuch, R.; Valls, C.; Lambruschini,
N. Plasma total-homocysteinemia in anorexia nervosa. Eur. J. Clin.
Nutr., 1998, 52, 172-175.
[143] Gallistl, S.; Sudi, K.M.; Erwa, W.; Aigner, R.; Borkenstein, M.
Determinants of homocysteine during weight reduction in obese
children and adolescents. Metabolism, 2001, 50, 1220-1223.
[144] Borzon-Chazot, F.; Harthe, C.; Teboul, F.; Labrousse, F.; Gaume,
C.; Guadagnino, L.; Claustrat, B.; Berthezène, F.; Moulin, P. Occurrence
of hyperhomocysteinemia 1 year after gastroplasty for severe
obesity. J. Clin. Endocrinol. Metab., 1999, 84, 541-545.
[145] Sheu, W.H.; Wu, H.S.; Wang, C.W.; Wan, C.J.; Lee, W.J. Elevated
plasma homocysteine concentrations after gastroplasty in morbidly
obese subjects. Intern. Med., 2001, 40, 584-588.
[147] Chandalia, M.; Abate, N.; Cabo-Chan, A.V. Jr.; Devaraj, S.; Jialal,
I.; Grundy, S.M. Hyperhomocysteinemia in Asian Indians living in
the United States. J. Clin. Endocrinol. Metab., 2003, 88, 1089-
1095.
[148] Koebnick, C.; Garcia, A.L.; Dagnelie, P.C.; Strassner, C.; Lindemans,
J.; Katz, N.; Leitzmann, C.; Hoffmann, I. Long-term consumption
of a raw food diet is associated with favorable serum
LDL cholesterol and triglycerides but also with elevated plasma
homocysteine and low HDL cholesterol in humans. J. Nutr., 2005,
135, 2372-2375.
[149] Coleman, R. The importance of sulfur as a plant nutrient in world
crop production. Soil Sci., 1966, 101, 230-239.
[150] Dijkshoorn, W.; Van Wijk, A.L. The sulphur requirements of
plants as evidenced by the sulphur-nitrogen ratio in the organic
matter: a review of published data. Plant Soil, 1967, 26, 129-157.
[152] Jez, J.M. Sulfur: A Missing Link between Soils, Crops, and Nutrition;
American Society of Agronomy, Crop Science Society of
America, Soil Science Society of America: Madison, 2008.
[154] Khurana, M.P.S.; Sadana, U.S.; Bijay-Singh. Sulfur-Nutrition of
Crops in the Indo-Gangetic Plains of South Asia. In: Sulfur: A
Missing Link between Soils, Crops, and Nutrition; Jez, J.M., Ed.;
American Society of Agronomy, Crop Science Society of America,
Soil Science Society of America: Madison, 2008; Vol. 50, pp. 11-
24.
[157] Galili, G.; Amir, R.; Hoefgen, R.; Hesse, H. Improving the levels
of essential amino acids and sulfur metabolites in plants. Biol.
Chem., 2005, 386, 817-831.
[159] Kumar, Y.; Das, R.; Garewal, G.; Bali, H.K. High prevalence of
hyperhomocysteinemia in young population of North India – a potential
risk factor for coronary artery disease? Thromb. Res., 2009,
123, 800-802.
[160] Ghosh, K.; Khare, A.; Shetty, S. Fasting plasma homocysteine
levels are increased in young patients with acute myocardial infarction
in Western India. Indian Heart J., 2007, 59, 242-245.
[161] Christopher, R.; Nagaraja, D.; Shankar, S.K. Homocysteine and
cerebral stroke in developing countries. Curr. Med. Chem., 2007,
14, 2393-2401.
Received: September 30, 2011 Revised: October 12, 2011 Accepted: October 12, 2011
© Yves Ingenbleek; Licensee Bentham Open.
This is an open access article licensed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/
by-nc/3.0/) which permits unrestricted, non-commercial use, distribution and reproduction in any medium, provided the work is properly cited.
Related articles

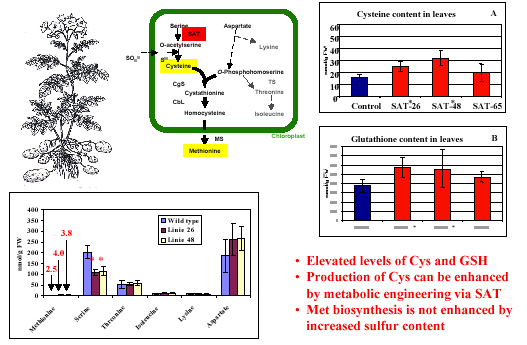
English: Biosynthesis of cysteine from homocysteine and serine via cystathione intermediate (Photo credit: Wikipedia)
Read Full Post »
















 and Protein C Pat...")





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}