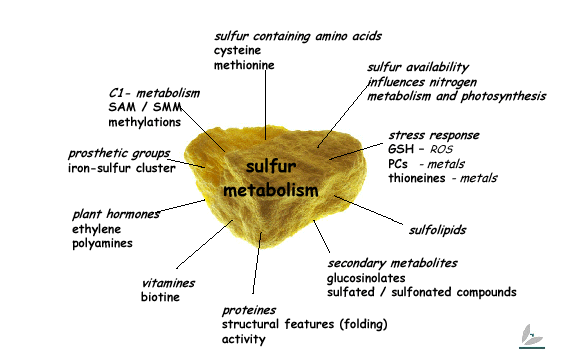
The biochemistry of S amino acids
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Amino Acid and Sulfur Metabolism
Dr. Rainer Höfgen
http://www.mpimp-golm.mpg.de/5892/2hoefgen
Sulfur is together with nitrogen, phosphorous and potassium a plant macronutrient and a crucial element affecting plant growth, plant performance and yield. The group of Dr. Rainer Hoefgen focuses on characterising the regulation of cysteine and methionine as a result of sulfate uptake and assimilation in the model plant Arabidopsis thaliana.
Cysteine and methionine are two essential amino acids which contain sulfur. We are also looking at interconnections between sulfur metabolism and other plant nutrients. Further, we are investigating means of improving the nutritional quality of crops, with a current focus on rice (Oryza sativa) with respect to a balanced amino acid composition.
In our studies of plant sulfur metabolism, we use two mutually supporting approaches as the basis of our research portfolio. The first is a targeted, pathway-oriented approach aimed at understanding pathway architecture and coordination, and the regulation of the sulfur-containing metabolites as such. The second is a non-biased approach in which functional genomics is used to work out how sulfur metabolism is embedded and controlled within the whole plant system.

sulfur uptake and assimilation
{kind=link}
Sulfur is a required macronutrient, sulfur uptake and assimilation are crucial determinants in how quickly plants grow and cope with various stresses, and therefore, in how well crops yield.
Inorganic sulfate is taken up through plant roots and, via cysteine biosynthesis, incorporated as organic sulfur. Our investigations focus on fundamental questions about cysteine (cys) and methionine (met) biosynthesis and on the possibility of engineering crop plants enriched in these sulfur-containing amino acids. Methionine is essential for non-ruminant mammals (including man) and uptake of cysteine reduces the methionine requirement. We have used transgenic strategies to generate many plant lines affected in cysteine and methionine biosynthesis, and subjected them to detailed molecular and biochemical analyses. Recently, we embarked on a course to study sulfur metabolism in a holistic way, rather than focusing on single pathways as such. By applying functional genomic tools like transcript, metabolite, and protein profiling in our analysis of transgenic potato (Solanum tuberosum) and of the model plant Arabidopsis thaliana, we are heading for a better understanding of the sulfur metabolism network in plants.
To learn about the control mechanisms involved in sulfur-containing amino acid biosynthesis, we are isolating and studying the involved genes and their promoters. The model plant systems of our investigations are potato and Arabidopsis, although a limited amount of work is also dedicated to rice (Oryza sativa), cucumber (Cucumis sativus), and tomato (Lycopersicon esculentum). Various transgenic plants exhibiting reduced or increased expression of relevant genes in the pathway have been produced and analysed. Fundamental knowledge of pathway regulation has been obtained as well as an improvement of the nutritional quality of a crop plant: Nutritional quality is largely determined by methionine, which is often the most limited of the essential amino acids.
The main thrust of our research recently shifted to analysing sulfur metabolism networks. In a systems biology approach, we investigate the response of Arabidopsis to different periods or degrees of sulfur starvation by applying non-biased, multiparallel tools including transcript, protein, and metabolite profiling. Our results are integrated to form working models for further detailed investigations with a focus on regulatory aspects of metabolism. This work entails the detailed analysis of Arabidopsis mutants and pulls many of our earlier results together into biological context (eg. the increased thiol levels seen during SAT over-expression, glutathione involvement in stress response mechanisms towards active oxygen species, etc.). Our long-term goal is to imbed sulfur metabolism in a broader context such as carbohydrate and nitrogen metabolic networks, which will occur through close collaborations with external and in house research groups.

metabolite profiling
http://www.mpimp-golm.mpg.de/12388/teaser_image_horizontal.jpg
{kind=link}
Plants are sessile organisms; if they are to survive and reproduce, they must adapt to the growth conditions in which they find themselves. We use variations in sulfur levels as a stimulus and analyse the complex response using diverse multiparallel techniques, particularly transcript and metabolite profiling, trying to piece together the total system response. The plant of choice here is, obviously, Arabidopsis thaliana, although results obtained in this model system are likely to be transferable to other plant species and crop plants. The goal is to provide a consistent and holistic description of plant sulfur metabolism and its regulation.
H Hesse and R Höfgen (2001) Application of Genomics in Agriculture. In: Molecular analysis of plant adapatation to the environment. Eds: Malcolm J. Hawkesford, Peter Buchner. Kluwer AP, Dordrecht, The Netherlands, 61-79
V Nikiforova, J Freitag, S Kempa, M Adamik, H Hesse, R Hoefgen (2003) Transcriptome analysis of sulfur depletion in Arabidopsis thaliana: Interlacing of biosynthetic pathways provides response specificity. The Plant Journal, 33, 633-650.
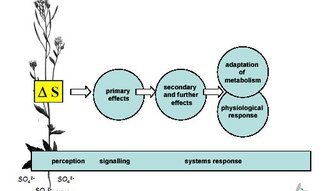
Regulation
Plants adapt to available sulfur soil levels by regulating gene expression and protein activity to maintain homeostasis. Sulfur availability in the environment is not static, nor is the plant’s dependence on sulfur at various developmental stages. Thus, one can assume not only that the activities of regulatory proteins are dynamic, but also that changes in the expression of transcription factors involved in triggering downstream gene expression change temporally. Sulfur deprivation triggers a slow adaptive process that resets the level of sulfur homeostasis. Using transcript profiling, we have been able to identify a number of transcription factors involved in this process, which are now the target of further investigations.
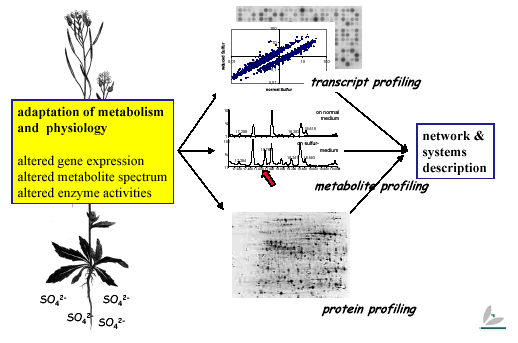
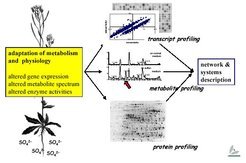
Metabolome analysis and bioinformatics

system response
{kind=link}
http://www.mpimp-golm.mpg.de/12342/Figure_2_Sulfur_Metabolism1.jpg
{kind=link}
Gene expression, metabolite spectrum and enzyme activities change under sulfur-limiting conditions.
The response of steady state transcription levels to the sulfur stimulus is but the first chapter of the story. To understand the system response, we have to turn the page and look at protein profiles – levels and activities – and before closing the book, at metabolite profiles, which adjust rapidly in response to changes in protein expression. We are now focusing on metabolome analysis: The same samples used for transcriptome analysis are examined using element analysis (ICP-AES) and metabolite analyses (HPLC, CE, GC/MS, GC/TOF, LC/MS), either in house or in collaboration with outside research groups.
Malcolm J. Hawkesford, Rothamsted Research, UK
As these analyses are refined and data accumulates, it will become more and more important to overlay and compare transcript and metabolite profiles in order to try to generate an in silico representation of the plant sulfur regulatory complement. Various approaches are and will be followed here: bioinformatic tools have to be developed and/or adapted to fully mine the data. Otherwise, it will not be possible to fully describe the system: by looking only at the most highly expressed genes in isolation, we would simply be scratching at the surface.
Transcriptome Analysis

gene expression
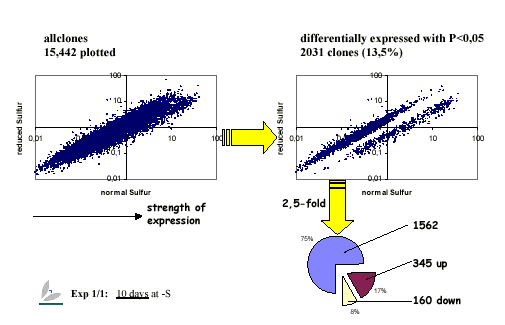{kind=link}
Scatterplots of gene expression of the ratio -/+ S
http://www.mpimp-golm.mpg.de/12424/Figure_4_Scatterplot1.jpg
{kind=link}
Plants and some photoautotrophic bacteria assimilate inorganic sulfur from sulfates into cysteine, the first sulfur-containing organic compound and, effectively, the sole molecular doorway for reduced sulfur in all living beings. This essential process has been as finely tuned through millennia of evolution as photosynthesis. Cysteine is subsequently converted to methionine, and then into a variety of other sulfur-containing organic compounds. Sulfur assimilation is even more spendy in terms of reduction equivalents than nitrogen assimilation. Obviously, such a costly enterprise is highly controlled in juxtaposition with the rest of metabolism.
To elucidate this network of interactions, we stimulate Arabidopsis with sulfur (i.e. sulfate) at its rhizosphere with various concentrations and at different developmental stages to institute periods of starvation and replenishment. The plant tissue samples (roots, shoots) are then subjected to array hybridisation/transcript profiling after RNA extraction using either macro-arrays of 7,200 non-redundant genes on nylon filters and now full genome chips. The expression profiles are processed to select differentially expressed genes. Depending on the duration of treatment, anything between a handful and thousands of genes exhibit altered expression mirroring the gradual response of the system to conditions of altered sulfur availability. Among these responsive genes we expect to find sulfur-regulated genes; genes involved in perception, signalling, and immediate responses; and genes further down the line involved in more pleiotropic mechanisms like general stress responses. Since they arise in response to sulfur stimulation, the latter are still regarded as sulfur-responsive genes.
Sulfur-responsive genes are grouped by functional category or biosynthetic pathway. As expected, genes of the sulfur assimilation pathway are altered in expression. Furthermore, genes involved in the flavonoid, auxin, and jasmonate biosynthesis pathways are up regulated when sulfur is limiting. We focus most of our attention, however, on the regulatory elements, transcription factors.
V Nikiforova, J Freitag, S Kempa, M Adamik, H Hesse, R Hoefgen (2003) Transcriptome analysis of sulfur depletion in Arabidopsis thaliana: Interlacing of biosynthetic pathways provides response specificity. The Plant Journal, 33, 633-650
Further reading
MY Hirai, T Fujiwara, M Awazuhara, T Kimura, M Noji, K Saito (2003) Global expression profiling of sulfur-starved Arabidopsis by DNA macroarray reveals the role of O-acetyl-L-serine as a general regulator of gene expression in response to sulfur nutrition. Plant Journal. 33(4)651-663
A Maruyama-Nakashita, E Inoue, A Watanabe-Takahashi, T Yarnaya, and H Takahashi (2003) Transcriptome profiling of sulfur-responsive genes in Arabidopsis reveals global effects of sulfur nutrition on multiple metabolic pathways. Plant Physiology. 132(2)597-605
Sulfur and Other Plant Nutrients
The plant sulfur assimilation pathway is intricately interconnected with various other pathways and regulatory circuits.
Systems Analysis of Plant Sulfur Metabolism
Every organism is a complex, multi-elemental, multi-functional system living in an ever-changing environment. The viability of the system is provided by, and likewise dependent upon, flexible, effective control circuits of multiple informational fluxes, which interconnect in a dense network of metabolic physiological responses.

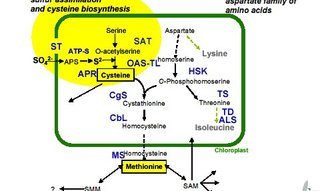
L-cysteine L-Met

Methionine is synthesised from cysteine and phosphohomoserine
http://www.mpimp-golm.mpg.de/12530/teaser_image_horizontal.jpg
{kind=link}
Pathway Analysis of Sulfur Containing Amino Acids
To learn about the control mechanisms involved in the biosynthesis of sulfur-containing amino acids, we are isolating and studying genes involved and their promoters. Methionine is synthesised from cysteine and phosphohomoserine via the enzymes cystathionine gamma-synthase (CgS), cystathionine beta-lyase (CbL), and methionine synthase (MS); we have cloned and characterised these three genes in potato.

Biosynthesis of Sulfur-Containing Amino Acids
http://www.mpimp-golm.mpg.de/12502/Figure_51.jpg
{kind=link}
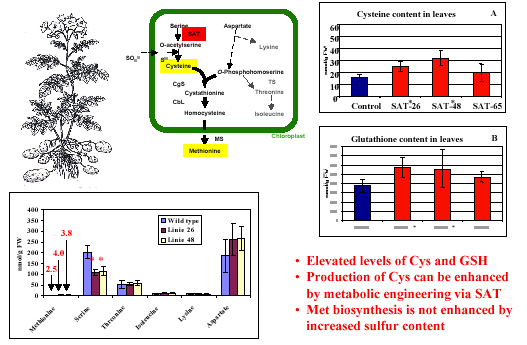
Genes from Arabidopsis and potato and, when appropriate, E. coli involved in cysteine and methionine biosynthesis have also been cloned, including various isoforms of O-acetylserine (thiol)-lyase, the enzyme that converts O-acetylserine to cys; ATP-sulfurylase, the enzyme activating the inert sulfate through binding to ATP; and serine acetyltransferase (SAT), the enzyme catalysing the activation of serine to O-acetylserine. We manipulated the expression of these genes in an attempt to create conditions in which flux to either cysteine or methionine is increased.
For example, the over-expression of SAT using an E. coli gene targeted to plastids resulted in cysteine and glutathione (a tripeptide containing glutamic acid, cysteine, and glycine) levels almost twice as high as usual. By blocking the competing pathway to threonine using the partial antisense inhibition of threonine synthase in Arabidopsis and potato, we were able to increase leaf and tuber methionine levels significantly. Moreover, analysis of these transformants made it clear that there are species-specific differences in the regulation of methionine biosynthesis.
Our results in Nicotiana plumbaginifolia and potato have established the essential, but not rate-limiting, role of CbL in plant methionine biosynthesis. Furthermore, we found that regulation at the level of CgS differs between the plant species Arabidopsis and potato. Our objective now is to deepen our understanding of the regulation of methionine biosynthesis and to exploit what we learn in order to improve the nutritional quality of crop plants, which is largely determined by methionine content.
Cysteine Biosynthesis
Cysteine biosynthesis represents the essential step in the incorporation of inorganic sulfide to organic sulfur in plants. In order to gain insight into the control mechanisms involved in cysteine biosynthesis, we are isolating and studying the involved genes and their promoters, including genes coding for O-acetylserine(thiol)-lyase (OAS-TL), the enzyme which converts O-acetylserine to cysteine, and serine acetyltransferase (SAT), the enzyme catalysing the activation of serine to O-acetylserine.

Serine acetyltransferase
{kind=link}
Serine acetyltransferase
http://www.mpimp-golm.mpg.de/12602/Figure_6_Serine_Acetyltransferase1.jpg
{kind=link}
In addition, spatial and developmental aspects of regulation are investigated with respect to gene expression and enzyme activity. We are manipulating the expression of various genes in transgenic potato plants in an attempt to create conditions in which flux to either cysteine or methionine is increased. For example, the heterologous over-expression of an E. coli SAT gene targeted to plastids resulted in a doubling of both cysteine and glutathione (a tripeptide containing glutamic acid, cysteine, and glycine that is involved in stress tolerance) levels. However, these alterations had no effect on outward plant appearance or on the expression and enzymatic activity of OAS-TL. This example demonstrates the importance of SAT in plant cysteine biosynthesis and shows that the accumulation of cysteine and related sulfur-containing compounds is limited by the supply of activated carbon backbones derived from serine. We are currently investigating this and other transgenic plants affected in cysteine and methionine biosynthesis in respect to sulfur assimilation and glutathione-mediated stress tolerance.
Despite the increase of reduced organic sulfur in our potato SAT over-producers, we did not observe an increase in methionine, although other groups reported methionine increases when using a similar approach in maize (Tsakraklides et al., 2002). Again, species specific differences, probably as a result of adaptation to specific environmental or physiological conditions, have to be taken into account, especially when generalising and transferring these data to plant breeding.
V Nikiforova, S Kempa, M Zeh, S Maimann, O Kreft, A P Casazza, K Riedel, E Tauberger, R Hoefgen, H Hesse. (2002) Engineering of cysteine and methionine biosynthesis in potato. Amino Acids 22(259-278).
K Harms, P von Ballmoos, C Brunold, R Höfgen, and H Hesse (2000) Expression of a bacterial serine acetyltransferase in transgenic potato plants leads to increased levels of cysteine and glutathione. Plant J. 22, 335-343
Further reading
MJ Hawkesford (2003) Transporter gene families in plants: the sulphate transporter gene family – redundancy or specialization? Physiologia Plantarum, 117,155-163
G Tsakraklides, M Martin, R Chalam,, MC Tarczynski, A Schmidt, and T Leustek (2002) Sulfate reduction is increased in transgenic Arabidopsis thaliana expressing 5′-adenylylsulfate reductase from Pseudomonas aeruginosa. Plant J. 32, 879
Met metabolism occurs primarily by activation of Met to AdoMet and further metabolism of AdoMet by either the transmethylation-transsulfuration pathway or the polyamine biosynthetic pathway. The catabolism of the methyl group and sulfur atom of Met ultimately appears to be dependent upon the transmethylation-transsulfuration pathway because the MTA formed as the co-product of polyamine synthesis is efficiently recycled to Met. On the other hand, the fate of the four-carbon chain of Met appears to depend upon the initial fate of the Met molecule. During transsulfuration, the carbon chain is released as alpha-ketobutyrate, which is further metabolized to CO2. In the polyamine pathway, the carboxyl carbon of Met is lost in the formation of dAdoMet, whereas the other three carbons are ultimately excreted as polyamine derivatives and degradation products. The role of the transamination pathway of Met metabolism is not firmly established. Cys (which may be formed from the sulfur of Met and the carbons of serine via the transsulfuration pathway) appears to be converted to taurine and CO2 primarily by the cysteinesulfinate pathway, and to sulfate and pyruvate primarily by desulfuration pathways in which a reduced form of sulfur with a relatively long biological half-life appears to be an intermediate. With the exception of the nitrogen of Met that is incorporated into polyamines, the nitrogen of Met or Cys is incorporated into urea after it is released as ammonium [in the reactions catalyzed by cystathionase with either cystathionine (from Met) or cystine (from Cys) as substrate] or it is transferred to a keto acid (in Cys or Met transamination). Many areas of sulfur-containing amino acid metabolism need further study. The magnitude of AdoMet flux through the polyamine pathway in the intact animal as well as details about the reactions involved in this pathway remain to be determined. Both the pathways and the possible physiological role of alternate (AdoMet-independent) Met metabolism, including the transamination pathway, must be elucidated. Despite the growing interest in taurine, investigation of Cys metabolism has been a relatively inactive area during the past two decades. Apparent discrepancies in the reported data on Cys metabolism need to be resolved. Future work should consider the role of extrahepatic tissues in amino acid metabolism as well as species differences in the relative roles of various pathways in the metabolism of Met and Cys.
The Sulfur-Containing Amino Acids: An Overview1,2
John T. Brosnan3 and Margaret E. Brosnan
J. Nutr. June 2006; 136(6): 1636S-1640S
http://jn.nutrition.org/content/136/6/1636S.full
Methionine and cysteine may be considered to be the principal sulfur-containing amino acids because they are 2 of the canonical 20 amino acids that are incorporated into proteins. However, homocysteine and taurine also play important physiological roles (Fig. 1). Why does nature employ sulfur in her repertoire of amino acids? The other canonical amino acids are comprised only of carbon, hydrogen, oxygen, and nitrogen atoms. Because both sulfur and oxygen belong to the same group (Group 6) of the Periodic Table and, therefore, are capable of making similar covalent linkages, the question may be restated: why would methionine and cysteine analogs, in which the sulfur atom is replaced by oxygen, not serve the same functions? One of the critical differences between oxygen and sulfur is sulfur’s lower electronegativity. Indeed, oxygen is the second most electronegative element in the periodic table. This accounts for the use of sulfur in methionine; replacement of the sulfur with oxygen would result in a much less hydrophobic amino acid. Cysteine readily forms disulfide linkages because of the ease with which it dissociates to form a thiolate anion. Serine, on the other hand, which differs from cysteine only in the substitution of an oxygen for the sulfur, does not readily make dioxide linkages. The difference results from the fact that thiols are much stronger acids than are alcohols, so that the alcohol group in serine does not dissociate at physiological pH. Substitution of oxygen for sulfur inS-adenosylmethionine would produce so powerful a methylating agent that it would promiscuously methylate cellular nucleophiles without the need for an enzyme.

FIGURE 1
Structures of the sulfur-containing amino acids.
Methionine and cysteine in proteins.
Although both methionine and cysteine play critical roles in cell metabolism, we suggest that, in general, the 20 canonical amino acids were selected for the roles they play in proteins, not their roles in metabolism. It is important, therefore, to review the role played by these amino acids in proteins. Methionine is among the most hydrophobic of the amino acids. This means that most of the methionine residues in globular proteins are found in the interior hydrophobic core; in membrane-spanning protein domains, methionine is often found to interact with the lipid bilayer. In some proteins a fraction of the methionine residues are somewhat surface exposed. These are susceptible to oxidation to methionine sulfoxide residues. Levine et al. (1) regard these methionine residues as endogenous antioxidants in proteins. In E. coli glutamine synthetase, they tend to be arrayed around the active site and may guard access to this site by reactive oxygen species. Oxidation of these methionine residues has little effect on the catalytic activity of the enzyme. These residues may be reduced to methionine by means of the enzyme methionine sulfoxide reductase (2). Thus, an oxidation–reduction cycle occurs in which exposed methionine residues are oxidized (e.g., by H2O2) to methionine sulfoxide residues, which are subsequently reduced:

It is considered that the impaired activity of methionine sulfoxide reductase and the subsequent accumulation of methionine sulfoxide residues are associated with age-related diseases, neurodegeneration, and shorter lifespan (2).
Methionine is the initiating amino acid in the synthesis of eukaryotic proteins; N-formyl methionine serves the same function in prokaryotes. Because most of these methionine residues are subsequently removed, it is apparent that their role lies in the initiation of translation, not in protein structure. In eukaryotes, translation initiation involves the association of the initiator tRNA (met-tRNAimet) with eIF-2 and the 40S ribosomal subunit together with a molecule of mRNA. Drabkin and Rajbhandary (3) suggest that the hydrophobic nature of methionine is key to the binding of the initiator tRNA to eIF-2. Using appropriate double mutations (in codon and anticodon), they were able to show that the hydrophobic valine could be used for initiation in mammalian cells but that the polar glutamine was very poor.
Cysteine plays a critical role in protein structure by virtue of its ability to form inter- and intrachain disulfide bonds with other cysteine residues. Most disulfide linkages are found in proteins destined for export or residence on the plasma membrane. These disulfide bonds can be formed nonenzymatically; protein disulfide isomerase, an endoplasmic reticulum protein, can reshuffle any mismatched disulfides to ensure the correct protein folding (4).
S-Adenosylmethionine.
S-Adenosylmethionine (SAM)4 is a key intermediate in methionine metabolism. Discovered in 1953 by Cantoni (5) as the “active methionine” required for the methylation of guanidioacetate to creatine, it is now evident that SAM is a coenzyme of remarkable versatility (Fig. 2). In addition to its role as a methyl donor, SAM serves as a source of methylene groups (for the synthesis of cyclopropyl fatty acids), amino groups (in biotin synthesis), aminoisopropyl groups (in the synthesis of polyamines and, also, in the synthesis of ethylene, used by plants to promote plant ripening), and 5′-deoxyadenosyl radicals. SAM also serves as a source of sulfur atoms in the synthesis of biotin and lipoic acid (6). In mammals, however, the great bulk of SAM is used in methyltransferase reactions. The key to SAM’s utility as a methyl donor lies in the sulfonium ion and in the electrophilic nature of the carbon atoms that are adjacent to the sulfur atom. The essence of these methyltransferase reactions is that the positively charged sulfonium renders the adjoining methyl group electron-poor, which facilitates its attack on electron-rich acceptors (nucleophiles).

Metabolic versatility of S-adenosylmethionine.
Metabolic versatility of S-adenosylmethionine.
SAM can donate its methyl group to a wide variety of acceptors, including amino acid residues in proteins, DNA, RNA, small molecules, and even to a metal, the methylation of arsenite (7,8). At present, about 60 methyltransferases have been identified in mammals. However, the number is almost certainly much larger. A bioinformatic analysis of a number of genomes, including the human genome, by Katz et al. (9) has suggested that Class-1 SAM-dependent methyltransferases account for 0.6–1.6% of open reading frames in these genomes. This would indicate about 300 Class 1 methyltransferases in humans, in addition to a smaller number of Class 2 and 3 enzymes. In humans, it appears that guanidinoacetate N-methyltransferase (responsible for creatine synthesis) and phosphatidylethanolamine N-methyltransferase (synthesis of phosphatidylcholine) are the major users of SAM (10). In addition, there is substantial flux through the glycine N-methyltransferase (GNMT) when methionine intakes are high (11). An important property of all known SAM-dependent methyltransferases is that they are inhibited by their product, S-adenosylhomocysteine (SAH).
Methionine metabolism.
Methionine metabolism begins with its activation to SAM (Fig. 3) by methionine adenosyltransferase (MAT). The reaction is unusual in that all 3 phosphates are removed from ATP, an indication of the “high-energy” nature of this sulfonium ion. SAM then donates its methyl group to an acceptor to produce SAH. SAH is hydrolyzed to homocysteine and adenosine by SAH hydrolase. This sequence of reactions is referred to as transmethylation and is ubiquitously present in cells. Homocysteine may be methylated back to methionine by the ubiquitously distributed methionine synthase (MS) and, also, in the liver as well as the kidney of some species, by betaine:homocysteine methyltransferase (BHMT). MS employs 5-methyl-THF as its methyl donor, whereas BHMT employs betaine, which is produced during choline oxidation as well as being provided by the diet (10). Both MS and BHMT effect remethylation, and the combination of transmethylation andremethylation comprise the methionine cycle, which occurs in most, if not all, cells.

Major pathways of sulfur-containing amino acid metabolism.
Major pathways of sulfur-containing amino acid metabolism.
The methionine cycle does not result in the catabolism of methionine. This is brought about by the transsulfuration pathway, which converts homocysteine to cysteine by the combined actions of cystathionine β-synthase (CBS) and cystathionine γ-lyase (CGL). The transsulfuration pathway has a very limited tissue distribution; it is restricted to the liver, kidney, intestine, and pancreas. The conversion of methionine to cysteine is an irreversible process, which accounts for the well-known nutritional principle that cysteine is not a dietary essential amino acid provided that adequate methionine is available, but methionine is a dietary essential amino acid, regardless of cysteine availability. This pathway for methionine catabolism suggests a paradox: is methionine catabolism constrained by the need for methylation reactions? If this were so, the methionine in a methionine-rich diet might exceed the methylation demand so that full catabolism could not occur via this pathway. GNMT methylates glycine to sarcosine, which may, in turn, be metabolized by sarcosine dehydrogenase to regenerate the glycine and oxidize the methyl group to 5,10-methylene-THF.
Application of sophisticated stable isotope tracer methodology to methionine metabolism in humans has yielded estimates of transmethylation, remethylation, and transsulfuration. Such studies reveal important points of regulation. For example, the sparing effect of cysteine on methionine requirements is evident as an increase in the fraction of the homocysteine pool that is remethylated and a decrease in the fraction that undergoes transsulfuration (12). In young adults ingesting a diet containing 1–1.5 g protein·kg−1·d−1, about 43% of the homocysteine pool was remethylated, and 57% was metabolized through the transsulfuration pathway (transmethylation = 9.7, transulfuration = 5.4, remethylation = 4.4 μmol·kg−1·h−1) (13).
Methionine metabolism affords a remarkable example of the role of vitamins in cell chemistry. MS utilizes methylcobalamin as a prosthetic group, 1 of only 2 mammalian enzymes that are known to require Vitamin B-12. The methyl group utilized by MS is provided from the folic acid 1-carbon pool. Methylenetetrahydrofolate reductase (MTHFR), which reduces 5,10-methylene-THF to 5-methyl-THF, contains FAD as a prosthetic group. Both of the enzymes in the transsulfuration pathway (CBS and CGL) contain pyridoxal phosphate. It is hardly surprising, therefore, that deficiencies of each of these vitamins (Vitamin B-12, folic acid, riboflavin, and pyridoxine) are associated with elevated plasma homocysteine levels. The oxidative decarboxylation of the α-ketobutyrate produced by CGL is brought about by pyruvate dehydrogenase so that niacin (NAD), thiamine (thiamine pyrophosphate), and pantothenic acid (coenzyme A) may also be regarded as being required for methionine metabolism.
Not only are vitamins required for methionine metabolism, but methionine metabolism plays a crucial role in the cellular assimilation of folate. MS has 2 principal functions. In addition to its role in methionine conservation, MS converts 5-methyl-THF to THF, thereby making it available to support DNA synthesis and other functions. Because 5-methyl-THF is the dominant circulating form that is taken into cells, MS is essential for cellular folate assimilation. Impaired MS activity (e.g., brought about by cobalamin deficiency) results in the accumulation of the folate coenzymes as 5-methyl-THF, the so-called methyl trap (14). This hypothesis explains the fact that Vitamin B-12 deficiency causes a functional cellular folate deficiency.
The combined transmethylation and transsulfuration pathways are responsible for the catabolism of the great bulk of methionine. However, there is also evidence for the occurrence of a SAM-independent catabolic pathway that begins with a transamination reaction (15). This is a very minor pathway under normal circumstances, but it becomes more significant at very high methionine concentrations. Because it produces powerful toxins such as methane thiol, it has been considered to be responsible for methionine toxicity. The identity of the initiating transaminase is uncertain; the glutamine transaminase can act on methionine, but it is thought to be unlikely to do so under physiological conditions (15). In view of the likelihood that this pathway plays a role in methionine toxicity, more work is warranted on its components, tissue distribution, and physiological function.
Regulation of methionine metabolism.
The major means by which methionine metabolism is regulated are 1) allosteric regulation by SAM and 2) regulation of the expression of key enzymes. In the liver, SAM exerts powerful effects at a variety of loci. The liver-specific MAT has a highKm for methionine and, therefore, is well fitted to remove excess dietary methionine. It exhibits the unusual property of feedback activation; it is activated by its product, SAM (16). This property has been incorporated into a computer model of hepatic methionine metabolism, and it is clear that it renders methionine disposal exquisitely sensitive to the methionine concentration (17). SAM is also an allosteric activator of CBS and an allosteric inhibitor of MTHFR (18). Therefore, elevated SAM promotes transsulfuration (methionine oxidation) and inhibits remethylation (methionine conservation). Many of the enzymes involved in methionine catabolism (MAT 1, GNMT, CBS) are increased in activity on ingestion of a high-protein diet (18).
In addition to its function in methionine catabolism, the transsulfuration pathway also provides cysteine for glutathione synthesis. Cysteine availability is often limiting for glutathione synthesis, and it appears that in a number of cells (e.g., hepatocytes), at least half of the cysteine required is provided by transsulfuration, even in the presence of physiological concentrations of cysteine (19). Transsulfuration is sensitive to the balance of prooxidants and antioxidants; peroxides increase the transsulfuration flux, whereas antioxidants decrease it (20). It is thought that redox regulation of the transsulfuration pathway occurs at the level of CBS, which contains a heme that may serve as a sensor of the oxidative environment (21).
Taurine.
Taurine is remarkable, both for its high concentrations in animal tissues and because of the variety of functions that have been ascribed to it. Taurine is the most abundant free amino acid in animal tissues. Table 1 shows that, although taurine accounts for only 3% of the free amino acid pool in plasma, it accounts for 25%, 50%, 53%, and 19%, respectively, of this pool in liver, kidney, muscle, and brain. The magnitude of the intracellular taurine pool deserves comment. For example, skeletal muscle contains 15.6 μmol of taurine per gram of tissue, which amounts to an intracellular concentration of about 25 mM. In addition to its role in the synthesis of the bile salt taurocholate, taurine has been proposed, inter alia, to act as an antioxidant, an intracellular osmolyte, a membrane stabilizer, and a neurotransmitter. It is an essential nutrient for cats; kittens born to mothers fed taurine-deficient diets exhibit retinal degeneration (24). Taurine is found in mother’s milk, may be conditionally essential for human infants, and is routinely added to most infant formulas. Recent work has begun to reveal taurine’s action in the retina. It appears that taurine, via an effect on a glycine receptor, promotes the generation of rod photoreceptor cells from retinal progenitor cells (25).
View this table:
Perspective.
The sulfur-containing amino acids present a fascinating subject to the protein chemist, the nutritionist, and the metabolic scientist, alike. They play critical roles in protein synthesis, structure, and function. Their metabolism is vital for many critical functions. SAM, a remarkably versatile molecule, is said to be second, only to ATP, in the number of enzymes that require it. Vitamins play a crucial role in the metabolism of these amino acids, which, in turn, play a role in folic acid assimilation. Despite the great advances in our knowledge of the sulfur-containing amino acids, there are important areas where further work is required. These include methionine transamination and the molecular basis for the many functions of taurine.
Disorders of Sulfur Amino Acid Metabolism
- Generoso Andria, Brian Fowler, Gianfranco Sebastio
Chapter Inborn Metabolic Diseases pp 224-231
Editors
http://link.springer.com/chapter/10.1007%2F978-3-662-04285-4_18
http://dx.doi.org:/10.1007/978-3-662-04285-4_18
Several defects can exist in the conversion of the sulfur-containing amino acid methionine to cysteine and the ultimate oxidation of cysteine to inorganic sulfate (Fig. 18.1). Cystathionine-β-synthase (CBS) deficiency is the most important. It is associated with severe abnormalities of four organs or organ systems: the eye (dislocation of the lens), the skeleton (dolichostenomelia and arachnodactyly), the vascular system (thromboembolism), and the central nervous system (mental retardation, cerebrovascular accidents). A low-methionine, highcystine diet, pyridoxine, folate, and betaine in various combinations, and antithrombotic treatment may halt the otherwise unfavorable course of the disease. Methionine adenosyltransferase deficiency and γ-cystathionase deficiency usually do not require treatment. Isolated sulfite oxidase deficiency leads (in its severe form) to refractory convulsions, lens dislocation, and early death. No effective treatment exists.
-
1.
Rubba P, Faccenda F, Pauciullo P, Carbone L, Mancini M, Strisciuglio P, Carrozzo R, Sartorio R, Del Giudice E, Andria G (1990) Early signs of vascular disease in homocystinuria: a noninvasive study by ultrasound methods in eight families with cystathionine ß-synthase deficiency. Metabolism 39: 1191–1195 PubMedCrossRef
-
2.
Kang S-S, Wong PWK, Malinow MR (1992) Hyperhomocyst(e)inemia as a risk factor for occlusive vascular disease. Annu Rev Nutr 12: 279–288 PubMedCrossRef
-
3.
Boushey CJ, Beresford SA, Omenn GS, Motulsky AG (1995) A quantitative assessment of plasma homocysteine as a risk factor for vascular disease. Probable benefits of increasing folic acid intakes. JAMA 274: 1049–1057
-
4.
Mudd SH, Skovby F, Levy HL, Pettigrew KD, Wilcken B, Pyeritz RE, Andria G, Boers GHJ, Bromberg IL, Cerone R, Fowler B, Grobe H, Schmidt H, Schweitzer L (1985) The natural history of homocystinuria due to cystathionine (3-synthase deficiency. Am J Hum Genet 37: 1–31 PubMed
-
5.
de Franchis R, Sperandeo MP, Sebastio G, Andria G. The Italian Collaborative Study Group on Homocystinuria (1998) Clinical aspects of cystathionine ß-synthase deficiency: how wide is the spectrum? Eur J Pediatr 157: S67–7o
-
6.
Kraus JP (1994) Molecular basis of phenotype expression in homocystinuria. J Inherited Metab Dis 17: 383–390 PubMedCrossRef
- more…
Leave a Reply