Metformin, Thyroid-Pituitary Axis, Diabetes Mellitus, and Metabolism
Larry H, Bernstein, MD, FCAP, Author and Curator
and Aviva Lev-Ari, PhD, RN
The following article is a review of the central relationship between the action of
metformin as a diabetic medication and its relationship to AMPK, the important and
essential regulator of glucose and lipid metabolism under normal activity, stress, with
its effects on skeletal muscle, the liver, the action of T3 and more.
We start with a case study and a publication in the J Can Med Assoc. Then we shall look
into key literature on these metabolic relationships.
Part I. Metformin , Diabetes Mellitus, and Thyroid Function
Hypothyroidism, Insulin resistance and Metformin
May 30, 2012 By Janie Bowthorpe
The following was written by a UK hypothyroid patient’s mother –
Sarah Wilson.
My daughter’s epilepsy is triggered by unstable blood sugars. And since taking
Metformin to control her blood sugar, she has significantly reduced the number of
seizures. I have been doing research and read numerous academic medical journals,
which got me thinking about natural thyroid hormone and Hypothyroidism. My hunch
was that when patients develop hypothyroid symptoms, they are actually becoming
insulin resistant (IR). There are many symptoms in common between women with
polycystic ovaries and hypothyroidism–the hair loss, the weight gain, etc.
(http://insulinhub.hubpages.com/hub/PCOS-and-Hypothyroidism).
A hypothyroid person’s body behaves as if it’s going into starvation mode and so, to
preserve resources and prolong life, the metabolism changes. If hypothyroid is prolonged
or pronounced, then perhaps, chemical preservation mode becomes permanent even
with the reintroduction of thyroid hormones. To get back to normal, they need
a “jump-start” reinitiate a higher rate of metabolism. The kick start is initiated through
AMPK, which is known as the “master metabolic regulating enzyme.”
(http://en.wikipedia.org/wiki/AMP-activated protein kinase).
Guess what? This is exactly what happens to Diabetes patients when Metformin is
introduced. http://en.wikipedia.org/wiki/Metformin
Suggested articles: http://www.springerlink.com/content/r81606gl3r603167/ and
http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2265.2011.04029.x/pdf
Note the following comments/partial statements:
“Hypothyroidism is characterized by decreased insulin responsiveness”;
“the pivotal regulatory role of T3 in major metabolic pathways”.
The community knows that T3/NTH (natural thyroid hormone [Armour]) makes
hypothyroid patients feel better – but the medical establishment is averse to T3/NTH
(treating subclinical hypoT (T3/T4 euthyroid) with natural dessicated thyroid (NDT).
The medical establishment might find an alternative view about impaired metabolism
more if shown real proof that the old NDT **was/is** having the right result –i.e., the
T3 is jump-starting the metabolism by re-activating AMPK.
If NDT also can be used for hypothyroidism without the surmised “dangers” of NTH,
then they should consider it. [The reality in the choice is actually recombinant TH
(Synthroid)]. Metformin is cheap, stable and has very few serious side effects. I use the
car engine metaphor, and refer to glucose as our petrol, AMPK as the spark plug and
both T3 and Metformin as the ignition switches. Sometimes if you have flat batteries in
the car, it doesn’t matter how much you turn the ignition switch or pump the petrol
pedal, all it does is flatten the battery and flood the engine.
Dr. Skinner in the UK has been treating “pre-hypothyroidism” the way that some
doctors treat “pre-diabetes”. Those hypothyroid patients who get treated early
might not have had their AMPK pathways altered and the T4-T3 conversion still works.
There seems to be no reason why thyroid hormone replacement therapy shouldn’t
logically be given to ward off a greater problem down the line.
It’s my belief that there is clear and abundant academic evidence that the AMPK/
Metformin research should branch out to also look at thyroid disease.
Point – direct T3 is kicking the closed -down metabolic process back into life,
just like Metformin does for insulin resistance.
http://www.hotthyroidology.com/editorial_79.html
There is serotonin resistance! http://www.ncbi.nlm.nih.gov/pubmed/17250776
Metformin Linked to Risk of Low Levels of Thyroid Hormone
CMAJ (Canadian Medical Association Journal) 09/22/2014
Metformin, the drug commonly for treating type 2 diabetes,
- is linked to an increased risk of low thyroid-stimulating hormone
(TSH) levels - in patients with underactive thyroids (hypothyroidism),
according to a study in CMAJ (Canadian Medical Association Journal).
Metformin is used to lower blood glucose levels
- by reducing glucose production in the liver.
previous studies have raised concerns that
- metformin may lower thyroid-stimulating hormone levels.
Study characteristics:
- Retrospective long-term
- 74 300 patient who received metformin and sulfonylurea
- 25-year study period.
- 5689 had treated hypothyroidism
- 59 937 had normal thyroid function.
Metformin and low levels of thyroid-stimulating hormone in
patients with type 2 diabetes mellitus
Jean-Pascal Fournier, Hui Yin, Oriana Hoi Yun Yu, Laurent Azoulay +
Centre for Clinical Epidemiology (Fournier, Yin, Yu, Azoulay), Lady Davis Institute,
Jewish General Hospital; Department of Epidemiology, Biostatistics and Occupational
Health (Fournier), McGill University; Division of Endocrinology (Yu), Jewish General
Hospital; Department of Oncology (Azoulay), McGill University, Montréal, Que., Cananda
CMAJ Sep 22, 2014, http://dx.doi.org:/10.1503/cmaj.140688
Background:
- metformin may lower thyroid-stimulating hormone (TSH) levels.
Objective:
- determine whether the use of metformin monotherapy, when compared with
sulfonylurea monotherapy, - is associated with an increased risk of low TSH levels(< 0.4 mIU/L)
- in patients with type 2 diabetes mellitus.
Methods:
- Used the Clinical Practice Research Datalink,
- identified patients who began receiving metformin or sulfonylurea monotherapy
between Jan. 1, 1988, and Dec. 31, 2012. - 2 subcohorts of patients with treated hypothyroidism or euthyroidism,
followed them until Mar. 31, 2013.
- Used Cox proportional hazards models to evaluate the association of low TSH
levels with metformin monotherapy, compared with sulfonylurea monotherapy,
in each subcohort.
Results:
- 5689 patients with treated hypothyroidism and 59 937 euthyroid patients were
included in the subcohorts.
For patients with treated hypothyroidism:
- 495 events of low TSH levels were observed (incidence rate 0.1197/person-years).
- 322 events of low TSH levels were observed (incidence rate 0.0045/person-years)
in the euthyroid group.
- metformin monotherapy was associated with a 55% increased risk of low TSH
levels in patients with treated hypothyroidism (incidence rate 0.0795/person-years
vs.0.1252/ person-years, adjusted hazard ratio [HR] 1.55, 95% confidence
interval [CI] 1.09– 1.20), compared with sulfonylurea monotherapy, - the highest risk in the 90–180 days after initiation (adjusted HR 2.30, 95% CI
1.00–5.29). - No association was observed in euthyroid patients (adjusted HR 0.97, 95% CI 0.69–1.36).
Interpretation: The clinical consequences of this needs further investigation.
| Crude and adjusted hazard ratios for suppressed thyroid-stimulating hormone levels (< 0.1 mIU/L) associated with the use metformin monotherapy, compared with sulfonylurea monotherapy, in patients with treated hypothyroidism or euthyroidism and type 2 diabetes |
|||||
| Variable | No. events suppressed TSH levels |
Person-years of exposure |
Incidence rate, per 1000 person-years (95% CI) |
Crude HR |
Adjusted HR*(95% CI) |
| Patients with treated hypothyroidism, n = 5689 | |||||
| Sulfonylure, n = 762 |
18 | 503 | 35.8 (21.2–56.6) |
1.00 | 1.00 (reference) |
| Metformin, n = 4927 |
130 | 3 633 | 35.8 (29.9–42.5) |
1.05 | 0.99 (0.57–1.72) |
| Euthyroid patients, n = 59 937 | |||||
| Sulfonylurea, n = 7980 |
12 | 8 576 | 1.4 (0.7–2.4) |
1.00 | 1.00 (reference) |
| Metformin, n = 51 957 |
75 | 63 047 | 1.2 (0.9–1.5) |
0.85 | 1.03 (0.52–2.03) |
Part II. Metabolic Underpinning
(Source: Wikipedia, AMPK and thyroid)
5′ AMP-activated protein kinase or AMPK or 5′ adenosine monophosphate-activated protein kinase
is an enzyme that plays a role in cellular energy homeostasis.
It consists of three proteins (subunits) that
- together make a functional enzyme, conserved from yeast to humans.
- It is expressed in a number of tissues, including the liver, brain, and skeletal
muscle. - The net effect of AMPK activation is stimulation of
- hepatic fatty acid oxidation and ketogenesis,
- inhibition of cholesterol synthesis,
- lipogenesis, and triglyceride synthesis,
- inhibition of adipocyte lipolysis and lipogenesis,
- stimulation of skeletal muscle fatty acid oxidation and muscle
glucose uptake, and - modulation of insulin secretion by pancreatic beta-cells.
The heterotrimeric protein AMPK is formed by α, β, and γ subunits. Each of these three
subunits takes on a specific role in both the stability and activity of AMPK.
- the γ subunit includes four particular Cystathionine beta synthase (CBS) domains
giving AMPK its ability to sensitively detect shifts in the AMP:ATP ratio. - The four CBS domains create two binding sites for AMP commonly referred to as
Bateman domains. Binding of one AMP to a Bateman domain cooperatively
increases the binding affinity of the second AMP to the other Bateman domain. - As AMP binds both Bateman domains the γ subunit undergoes a conformational
change which exposes the catalytic domain found on the α subunit. - It is in this catalytic domain where AMPK becomes activated when
phosphorylation takes place at threonine-172by an upstream AMPK kinase
(AMPKK). The α, β, and γ subunits can also be found in different isoforms.
AMPK acts as a metabolic master switch regulating several intracellular systems
- the cellular uptake of glucose,
- the β-oxidation of fatty acids and
- the biogenesis of glucose transporter 4 (GLUT4) and
- mitochondria
The energy-sensing capability of AMPK can be attributed to
- its ability to detect and react to fluctuations in the AMP:ATP ratio that take
place during rest and exercise (muscle stimulation).
During muscle stimulation,
- AMP increases while ATP decreases, which changes AMPK into a good substrate
for activation. - AMPK activity increases while the muscle cell experiences metabolic stress
brought about by an extreme cellular demand for ATP. - Upon activation, AMPK increases cellular energy levels by
- inhibiting anabolic energy consuming pathways (fatty acid synthesis,
protein synthesis, etc.) and - stimulating energy producing, catabolic pathways (fatty acid oxidation,
glucose transport, etc.).
- inhibiting anabolic energy consuming pathways (fatty acid synthesis,
A recent JBC paper on mice at Johns Hopkins has shown that when the activity of brain
AMPK was pharmacologically inhibited,
- the mice ate less and lost weight.
When AMPK activity was pharmacologically raised (AICAR see below)
- the mice ate more and gained weight.
Research in Britain has shown that the appetite-stimulating hormone ghrelin also
affects AMPK levels.
The antidiabetic drug metformin (Glucophage) acts by stimulating AMPK, leading to
- reduced glucose production in the liver and
- reduced insulin resistance in the muscle.
(Metformin usually causes weight loss and reduced appetite, not weight gain and
increased appetite, ..opposite of expected from the Johns Hopkins mouse study results.)
Triggering the activation of AMPK can be carried out provided two conditions are met.
First, the γ subunit of AMPK
- must undergo a conformational change so as to
- expose the active site(Thr-172) on the α subunit.
The conformational change of the γ subunit of AMPK can be accomplished
- under increased concentrations of AMP.
Increased concentrations of AMP will
- give rise to the conformational change on the γ subunit of AMPK
- as two AMP bind the two Bateman domains located on that subunit.
- It is this conformational change brought about by increased concentrations
of AMP that exposes the active site (Thr-172) on the α subunit.
This critical role of AMP is further substantiated in experiments that demonstrate
- AMPK activation via an AMP analogue 5-amino-4-imidazolecarboxamide
ribotide (ZMP) which is derived fromthe familiar - 5-amino-4-imidazolecarboxamide riboside (AICAR)
AMPK is a good substrate for activation via an upstream kinase complex, AMPKK
AMPKK is a complex of three proteins,
- STE-related adaptor (STRAD),
- mouse protein 25 (MO25), and
- LKB1 (a serine/threonine kinase).
The second condition that must be met is
- the phosphorylation/activation of AMPK on its activating loop at
Thr-172of the α subunit - brought about by an upstream kinase (AMPKK).
The complex formed between LKB1 (STK 11), mouse protein 25 (MO25), and the
pseudokinase STE-related adaptor protein (STRAD) has been identified as
- the major upstream kinase responsible for phosphorylation of AMPK
on its activating loop at Thr-172
Although AMPK must be phosphorylated by the LKB1/MO25/STRAD complex,
- it can also be regulated by allosteric modulators which
- directly increase general AMPK activity and
- modify AMPK to make it a better substrate for AMPKK
- and a worse substrate for phosphatases.
It has recently been found that 3-phosphoglycerate (glycolysis intermediate)
- acts to further pronounce AMPK activation via AMPKK
Muscle contraction is the main method carried out by the body that can provide
the conditions mentioned above needed for AMPK activation
- As muscles contract, ATP is hydrolyzed, forming ADP.
- ADP then helps to replenish cellular ATP by donating a phosphate group to
another ADP,- forming an ATP and an AMP.
- As more AMP is produced during muscle contraction,
- the AMP:ATP ratio dramatically increases,
- leading to the allosteric activation of AMPK
For over a decade it has been known that calmodulin-dependent protein kinase
kinase-beta (CaMKKbeta) can phosphorylate and thereby activate AMPK,
- but it was not the main AMPKK in liver.
CaMKK inhibitors had no effect on 5-aminoimidazole-4-carboxamide-1-beta-4-
ribofuranoside (AICAR) phosphorylation and activation of AMPK.
- AICAR is taken into the celland converted to ZMP,
- an AMP analogthat has been shown to activate AMPK.
Recent LKB1 knockout studies have shown that without LKB1,
- electrical and AICAR stimulation of muscleresults in very little
phosphorylation of AMPK and of ACC, providing evidence that - LKB1-STRAD-MO25 is the major AMPKK in muscle.
Two particular adipokines, adiponectin and leptin, have even been demonstrated
to regulate AMPK. A main functions of leptin in skeletal muscle is
- the upregulation of fatty acid oxidation.
Leptin works by way of the AMPK signaling pathway, and adiponectin also
stimulates the oxidation of fatty acids via the AMPK pathway, and
- Adiponectin also stimulates the uptake of glucose in skeletal muscle.
An increase in enzymes which specialize in glucose uptake in cells such as GLUT4
and hexokinase II are thought to be mediated in part by AMPK when it is activated.
Increases in AMPK activity are brought about by increases in the AMP:ATP ratio
during single bouts of exercise and long-term training.
One of the key pathways in AMPK’s regulation of fatty acid oxidation is the
- phosphorylation and inactivation of acetyl-CoA carboxylase.
- Acetyl-CoA carboxylase (ACC) converts acetyl-CoA (ACA) to malonyl-CoA
(MCA), an inhibitor of carnitine palmitoyltransferase 1 (CPT-1). - CPT-1 transports fatty acids into the mitochondria for oxidation.
- Inactivation of ACC results in increased fatty acid transport and oxidation.
- the AMPK induced ACC inactivation and reduced conversion to MCA
may occur as a result of malonyl-CoA decarboxylase (MCD) - MCD as an antagonist to ACC, decarboxylatesmalonyl-CoA to acetyl-CoA
(reversal of ACC conversion of ACA to MCA) - This resultsin decreased malonyl-CoA and increased CPT-1 and fatty acid oxidation.
AMPK also plays an important role in lipid metabolism in the liver. It has long been
known that hepatic ACC has been regulated in the liver.
- It phosphorylates and inactivates 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR)
- acetyl-CoA(ACA) is converted to mevalonic acid (MVA) by ACC
with inhibition of CPT-1 - HMGR converts 3-hydroxy-3-methylglutaryl-CoA, which is made from MVA
- which then travels down several more metabolic steps to become cholesterol.
Insulin facilitates the uptake of glucose into cells via increased expression and
translocation of glucose transporter GLUT-4. In addition, glucose is phosphorylated
by hexokinase wheni iot enters the cell. The phosphorylated form keeps glucose from
leaving the cell,
- The decreasedthe concentration of glucose molecules creates a gradient for more
glucose to be transported into the cell.
| AMPK and thyroid hormone regulate some similar processes. Knowing these similarities, Winder and Hardie et al. designed an experiment to see if AMPK was influenced by thyroid hormone. They found that all of the subunits of AMPK were increased in skeletal muscle, especially in the soleus and red quadriceps, with thyroid hormone treatment. There was also an increase in phospho-ACC, a marker of AMPK activity. |
- Winder WW, Hardie DG (July 1999). “AMP-activated protein kinase,
a metabolic master switch: possible roles in type 2 diabetes”. J. Physiol. 277
(1 Pt 1): E1–10. PMID 10409121. - Winder WW, Hardie DG (February 1996). “Inactivation of acetyl-CoA
carboxylase and activation of AMP-activated protein kinase in muscle
during exercise”. J. Physiol. 270 (2 Pt 1): E299–304. PMID 8779952. - Hutber CA, Hardie DG, Winder WW (February 1997). “Electrical stimulation
inactivates muscle acetyl-CoA carboxylase and increases AMP-activated
protein kinase”. Am. J. Physiol. 272 (2 Pt 1): E262–6. PMID 9124333 - Durante PE, Mustard KJ, Park SH, Winder WW, Hardie DG (July 2002).
“Effects of endurance training on activity and expression of AMP-activated
protein kinase isoforms in rat muscles”. Am. J. Physiol. Endocrinol.
Metab. 283 (1): E178–86. doi:10.1152/ajpendo.00404.2001. PMID 12067859 - Corton JM, Gillespie JG, Hardie DG (April 1994). “Role of the AMP-activated
protein kinase in the cellular stress response”. Curr. Biol. 4 (4):
315–24. doi:10.1016/S0960-9822(00)00070-1. PMID 7922340 - Winder WW (September 2001). “Energy-sensing and signaling by
AMP-activated protein kinase in skeletal muscle”. J. Appl. Physiol. 91 (3):
1017–28. PMID 11509493 - Suter M, Riek U, Tuerk R, Schlattner U, Wallimann T, Neumann D (October
2006). “Dissecting the role of 5′-AMP for allosteric stimulation, activation,
and deactivation of AMP-activated protein kinase”. J. Biol. Chem.
281 (43): 32207–6. doi:10.1074/jbc.M606357200. PMID 16943194
Part III. Pituitary-thyroid axis and diabetes mellitus
The Interface Between Thyroid and Diabetes Mellitus
Leonidas H. Duntas, Jacques Orgiazzi, Georg Brabant Clin Endocrinol. 2011;75(1):1-9.
Interaction of Metformin and Thyroid Function
Metformin acts primarily by
- suppressing hepatic gluconeogenesis via activation of AMPK
- It has the opposite effects on hypothalamic AMPK,
- inhibiting activity of the enzyme.
- the metformin effects on hypothalamic AMPK activity will
- counteractT3 effects at the hypothalamic level.
- AMPK therefore represents a direct target for dual regulation
- in the hypothalamic partitioning of energy homeostasis.
- metformin crossesthe blood–brain barrier and
- levels in the pituitary gland are substantially increased.
- It convincinglysuppresses TSH
A recent study recruiting 66 patients with benign thyroid nodules furthermore
demonstrated that metformin significantly decreases nodule size in patients with
insulin resistance.[76] The effect of metformin, which was produced over a
6-month treatment period, parallelled a fall in TSH concentrations and achieved a
shrinkage amounting to 30% of the initial nodule size when metformin was
administered alone and up to 55% when it was added to ongoing LT4 treatment.
These studies reveal a
- suppressive effect of metformin on TSH secretion patterns in
hypothyroid patients, an effect that is apparently - independent of T4 treatment and does not alter the TH profile.
- A rebound of TSH secretion occurs at about 3 months following metformin
withdrawal.
It appears that recommendations for more frequent testing, on an annual to
biannual basis, seems justified in higher risk groups like patients over 50 or 55,
particularly with suggestive symptoms, raised antibody titres or dylipidaemia.
We thus would support the suggestion of an initial TSH and TPO antibody testing
which, as discussed, will help to predict the development of hypothyroidism in
patients with diabetes.
Hypothalamic AMPK and fatty acid metabolism mediate thyroid
regulation of energy balance
M López, L Varela, MJ Vázquez, S Rodríguez-Cuenca, CR González, …, & Vidal-Puig
Nature Medicine 29 Aug 2010; 16: 1001–1008 http://dx.doi.org:/10.1038/nm.2207
Thyroid hormones have widespread cellular effects; however it is unclear whether
their effects on the central nervous system (CNS) contribute to global energy balance.
Here we demonstrate that either
- whole-body hyperthyroidism or central administration of triiodothyronine
(T3) decreases- the activity of hypothalamic AMP-activated protein kinase (AMPK),
- increases sympathetic nervous system (SNS) activity and
- upregulates thermogenic markers in brown adipose tissue (BAT).
Inhibition of the lipogenic pathway in the ventromedial nucleus of the hypothalamus
(VMH) prevents CNS-mediated activation of BAT by thyroid hormone and reverses
the weight loss associated with hyperthyroidism. Similarly, inhibition of thyroid
hormone receptors in the VMH reverses the weight loss associated with hyperthyroidism.
This regulatory mechanism depends on AMPK inactivation, as genetic inhibition of this
enzyme in the VMH of euthyroid rats induces feeding-independent weight loss and
increases expression of thermogenic markers in BAT. These effects are reversed by
pharmacological blockade of the SNS. Thus, thyroid hormone–induced modulation
of AMPK activity and lipid metabolism in the hypothalamus is a major regulator of
whole-body energy homeostasis.
Metabolic Basis for Thyroid Hormone Liver Preconditioning:
Upregulation of AMP-Activated Protein Kinase Signaling
LA Videla,1 V Fernández, P Cornejo, and R Vargas
1Molecular and Clinical Pharmacology Program, Institute of Biomedical Sciences,
Faculty of Medicine, University of Chile, 2Faculty of Medicine, Diego Portales University,
Santiago, Chile
Academic Editors: H. M. Abu-Soud and D. Benke
The Scientific World Journal 2012; 2012, ID 475675, 10 pp
http://dx.doi.org/10.1100/2012/475675
The liver is a major organ responsible for most functions of cellular metabolism and
- a mediator between dietary and endogenous sources of energy for extrahepatic tissues.
- In this context, adenosine-monophosphate- (AMP-) activated protein kinase (AMPK)
constitutes an intrahepatic energy sensor - regulating physiological energy dynamics by limiting anabolism and stimulating
catabolism, thus increasing ATP availability. - This is achieved by mechanisms involving direct allosteric activation and
reversible phosphorylation of AMPK, in response to signals such as- energy status,
- serum insulin/glucagon ratio,
- nutritional stresses,
- pharmacological and natural compounds, and
- oxidative stress status.
Reactive oxygen species (ROS) lead to cellular AMPK activation and
- downstream signaling under several experimental conditions.
Thyroid hormone (L-3,3′,5-triiodothyronine, T3) administration, a condition
that enhances liver ROS generation,
- triggers the redox upregulation of cytoprotective proteins
- affording preconditioning against ischemia-reperfusion (IR) liver injury.
Data discussed in this work suggest that T3-induced liver activation of AMPK
- may be of importance in the promotion of metabolic processes
- favouring energy supply for the induction and operation of preconditioning
mechanisms.
These include
- antioxidant,
- antiapoptotic, and
- anti-inflammatory mechanisms,
- repair or resynthesis of altered biomolecules,
- induction of the homeostatic acute-phase response, and
- stimulation of liver cell proliferation,
which are required to cope with the damaging processes set in by IR.
The liver functions as a mediator between dietary and endogenous sources
of energy and extrahepatic organs that continuously require energy, mainly
the brain and erythrocytes, under cycling conditions between fed and fasted states.
In the fed state, where insulin action predominates, digestion-derived glucose is
converted to pyruvate via glycolysis, which is oxidized to produce energy, whereas
fatty acid oxidation is suppressed. Excess glucose can be either stored as hepatic
glycogen or channelled into de novo lipogenesis.
In the fasted state, considerable liver fuel metabolism changes occur due to decreased
serum insulin/glucagon ratio, with higher glucose production as a consequence of
stimulated glycogenolysis and gluconeogenesis (from alanine, lactate, and glycerol).
Major enhancement in fatty acid oxidation also occurs to provide energy for liver
processes and ketogenesis to supply metabolic fuels for extrahepatic tissues. For these
reasons, the liver is considered as the metabolic processing organ of the body, and
alterations in liver functioning affect whole-body metabolism and energy homeostasis.
In this context, adenosine-monophosphate- (AMP-) activated protein kinase (AMPK)
is the downstream component of a protein kinase cascade acting as an
- intracellular energy sensor regulating physiological energy dynamics by
- limiting anabolic pathways, to prevent excessive adenosine triphosphate (ATP)
utilization, and - by stimulating catabolic processes, to increase ATP production.
Thus, the understanding of the mechanisms by which liver AMPK coordinates hepatic
energy metabolism represents a crucial point of convergence of regulatory signals
monitoring systemic and cellular energy status
Liver AMPK: Structure and Regulation
AMPK, a serine/threonine kinase, is a heterotrimeric complex comprising
- a catalytic subunit α and
- two regulatory subunits β and γ .
The α subunit has a threonine residue (Thr172) within the activation loop of the kinase
domain, with the C-terminal region being required for association with β and γ subunits.
The β subunit associates with α and γ by means of its C-terminal region , whereas
- the γ subunit has four cystathionine β-synthase (CBS) motifs, which
- bind AMP or ATP in a competitive manner.
75675.fig.001 (not shown)
Figure 1: Regulation of AMP-activated protein kinase (AMPK) by
(A) direct allosteric activation and
(B) reversible phosphorylation and downstream responses maintaining
intracellular energy balance.
Regulation of liver AMPK activity involves both direct allosteric activation and
reversible phosphorylation. AMPK is allosterically activated by AMP through
- binding to the regulatory subunit-γ, which induces a conformational change in
the kinase domain of subunit α that protects AMPK from dephosphorylation
of Thr172, probably by protein phosphatase-2C.
Activation of AMPK requires phosphorylation of Thr172 in its α subunit, which can be
attained by either
(i) tumor suppressor LKB1 kinase following enhancement in the AMP/ATP ratio, a
kinase that plays a crucial role in AMPK-dependent control of liver glucose and
lipid metabolism;
(ii) Ca2+-calmodulin-dependent protein kinase kinase-β (CaMKKβ) that
phosphorylates AMPK in an AMP-independent, Ca2+-dependent manner;
(iii) transforming growth-factor-β-activated kinase-1 (TAK1), an important
kinase in hepatic Toll-like receptor 4 signaling in response to lipopolysaccharide.
Among these kinases, the relevance of CaMKKβ and TAK1 in liver AMPK activation
remains to be established in metabolic stress conditions. Both allosteric and
phosphorylation mechanisms are able to elicit
- over 1000-fold increase in AMPK activity, thus allowing
- the liver to respond to small changes in energy status in a highly sensitive fashion.
In addition to rapid AMPK regulation through allosterism and reversible phosphorylation
- long-term effects of AMPK activation induce changes in hepatic gene expression.
This was demonstrated for
(i) the transcription factor carbohydrate-response element-binding protein (ChREBP),
- whose Ser568 phosphorylation by activated AMPK
- blocks its DNA binding capacity and glucose-induced gene transcription
- under hyperlipidemic conditions;(ii) liver sterol regulatory element-binding
protein-1c (SREBP-1c), whose mRNA and protein expression and those of
its target gene for fatty acid synthase (FAS) - are reduced by metformin-induced AMPK activation,
- decreasing lipogenesis and increasing fatty acid oxidation due to
malonyl-CoA depletion;
(iii) transcriptional coactivator transducer of regulated CREB activity-2 (TORC2),
a crucial component of the hepatic gluconeogenic program, was reported
to be phosphorylated by activated AMPK.
This modification leads to subsequent cytoplasmatic sequestration of TORC2 and
inhibition of gluconeogenic gene expression, a mechanism underlying
- the plasma glucose-lowering effects of adiponectin and metformin
- through AMPK activation by upstream LKB1.
Activation of AMPK in the liver is a key regulatory mechanism controlling glucose
and lipid metabolism,
- inhibiting anabolic processes, and
- enhancing catabolic pathways in response to different signals, including
- energy status,
- serum insulin/glucagon ratio,
- nutritional stresses,
- pharmacological and natural compounds, and
- oxidative stress status
Reactive Oxygen Species (ROS) and AMPK Activation
The high energy demands required to cope with all the metabolic functions
of the liver are met by
- fatty acid oxidation under conditions of both normal blood glucose levels and
hypoglycemia, whereas - glucose oxidation is favoured in hyperglycemic states, with consequent
generation of ROS.
Under normal conditions, ROS occur at relatively low levels due to their fast processing
by antioxidant mechanisms, whereas at acute or prolonged high ROS levels, severe
oxidation of biomolecules and dysregulation of signal transduction and gene expression
is achieved, with consequent cell death through necrotic and/or apoptotic-signaling
pathways.
Thyroid Hormone (L-3,3′,5-Triiodothyronine, T3), Metabolic Regulation,
and ROS Production
T3 is important for the normal function of most mammalian tissues, with major actions
on O2 consumption and metabolic rate, thus
- determining enhancement in fuel consumption for oxidation processes
- and ATP repletion.
T3 acts predominantly through nuclear receptors (TR) α and β, forming
- functional complexes with retinoic X receptor that
- bind to thyroid hormone response elements (TRE) to activate gene expression.
T3 calorigenesis is primarily due to the
- induction of enzymes related to mitochondrial electron transport and ATP
synthesis, catabolism, and - some anabolic processes via upregulation of genomic mechanisms.
The net result of T3 action is the enhancement in the rate of O2 consumption of target
tissues such as liver, which may be effected by secondary processes induced by T3
(i) energy expenditure due to higher active cation transport,
(ii) energy loss due to futile cycles coupled to increase in catabolic and anabolic pathways, and
(iii) O2 equivalents used in hepatic ROS generation both in hepatocytes and Kupffer cells
In addition, T3-induced higher rates of mitochondrial oxidative phosphorylation are
likely to induce higher levels of ATP, which are partially balanced by intrinsic uncoupling
afforded by induction of uncoupling proteins by T3. In agreement with this view, the
cytosolic ATP/ADP ratio is decreased in hyperthyroid tissues, due to simultaneous
stimulation of ATP synthesis and consumption.
Regulation of fatty acid oxidation is mainly attained by carnitine palmitoyltransferase Iα (CPT-Iα),
- catalyzing the transport of fatty acids from cytosol to mitochondria for β-oxidation,
and acyl-CoA oxidase (ACO), - catalyzing the first rate-limiting reaction of peroxisomal β-oxidation, enzymes that are
induced by both T3 and peroxisome proliferator-activated receptor α (PPAR-α).
Furthermore, PPAR-α-mediated upregulation of CPT-Iα mRNA is enhanced by PPAR-γ
coactivator 1α (PGC-1α), which in turn
- augments T3 induction of CPT-Iα expression.
Interestingly, PGC-1α is induced by
- T3,
- AMPK activation, and
- ROS,
thus establishing potential links between
- T3 action, ROS generation, and AMPK activation
with the onset of mitochondrial biogenesis and fatty acid β-oxidation.
Liver ROS generation leads to activation of the transcription factors
- nuclear factor-κB (NF-κB),
- activating protein 1 (AP-1), and
- signal transducer and activator of transcription 3 (STAT3)
at the Kupffer cell level, with upregulation of cytokine expression (TNF-α, IL-1, IL-6),
which upon interaction with specific receptors in hepatocytes trigger the expression of
- cytoprotective proteins (Figure 3(A)).
These responses and the promotion of hepatocyte and Kupffer-cell proliferation
represent hormetic effects reestablishing
- redox homeostasis,
- promoting cell survival, and
- protecting the liver against ischemia-reperfusion injury.
T3 liver preconditioning also involves the activation of the
- Nrf2-Keap1 defense pathway
- upregulating antioxidant proteins,
- phase-2 detoxifying enzymes, and
- multidrug resistance proteins, members of the ATP binding cassette (ABC)
superfamily of transporters (Figure 3(B))
In agreement with T3-induced liver preconditioning, T3 or L-thyroxin afford
preconditioning against IR injury in the heart, in association with
- activation of protein kinase C and
- attenuation of p38 and
- c-Jun-N-terminal kinase activation ,
and in the kidney, in association with
- heme oxygenase-1 upregulation.
475675.fig.002
http://www.hindawi.com/journals/tswj/2012/floats/475675/thumbnails/475675.fig.002_th.jpg
Figure 2: Calorigenic response of thyroid hormone (T3) and its relationship with O2
consumption, reactive oxygen species (ROS) generation, and antioxidant depletion in the liver.
Abbreviations: CYP2E1, cytochrome P450 isoform 2E1; GSH, reduced glutathione; QO2, rate
of O2 consumption; SOD, superoxide dismutase.
475675.fig.003

genomic signaling in T3 calorigenesis and ROS production 475675.fig.003
http://www.hindawi.com/journals/tswj/2012/floats/475675/thumbnails/475675.fig.003_th.jpg
| Figure 3: Genomic signaling mechanisms in T3 calorigenesis and liver reactive oxygen species (ROS) production leading to (A) upregulation of cytokine expression in Kupffer cells and hepatocyte activation of genes conferring cytoprotection, (B) Nrf2 activation controling expression of antioxidant and detoxication proteins, and (C) activation of the AMPK cascade regulating metabolic functions.Abbreviations: AP-1, activating protein 1; ARE, antioxidant responsive element; CaMKKβ, Ca2+-calmodulin-dependent kinase kinase-β; CBP, CREB binding protein; CRC, chromatin remodelling complex; EH, epoxide hydrolase; HO-1, hemoxygenase-1; GC-Ligase, glutamate cysteine ligase; GPx, glutathione peroxidase; G-S-T, glutathione-S-transferase; HAT, histone acetyltransferase; HMT, histone arginine methyltransferase; IL1, interleukin 1; iNOS, inducible nitric oxide synthase; LKB1, tumor suppressor LKB1 kinase; MnSOD, manganese superoxide dismutase; MRPs, multidrug resistance proteins; NF-κB, nuclear factor-κB; NQO1, NADPH-quinone oxidoreductase-1; NRF-1, nuclear respiratory factor-1; Nrf2, nuclear receptor-E2-related factor 2; PCAF, p300/CBP-associated factor; RXR, retinoic acid receptor; PGC-1, peroxisome proliferator-activated receptor-γ coactivator-1; QO2, rate of O2 consumption; STAT3, signal transducer and activator of transcription 3; TAK1, transforming-growth-factor-β-activated kinase-1; TNF-α, tumor necrosis factor-α; TR, T 3 receptor; TRAP, T3-receptor-associated protein; TRE, T3 responsive element; UCP, uncoupling proteins; (—), reported mechanisms; (- - - -), proposed mechanisms. |
T3 is a key metabolic regulator coordinating short-term and long-term energy needs,
with major actions on liver metabolism. These include promotion of
(i) gluconeogenesis and hepatic glucose production, and
(ii) fatty acid oxidation coupled to enhanced adipose tissue lipolysis, with
- higher fatty acid flux to the liver and
- consequent ROS production (Figure 2) and
- redox upregulation of cytoprotective proteins
affording liver preconditioning (Figure 3).
Thyroid Hormone and AMPK Activation: Skeletal Muscle and Heart
In skeletal muscle, T3 increases the levels of numerous proteins involved in
- glucose uptake (GLUT4),
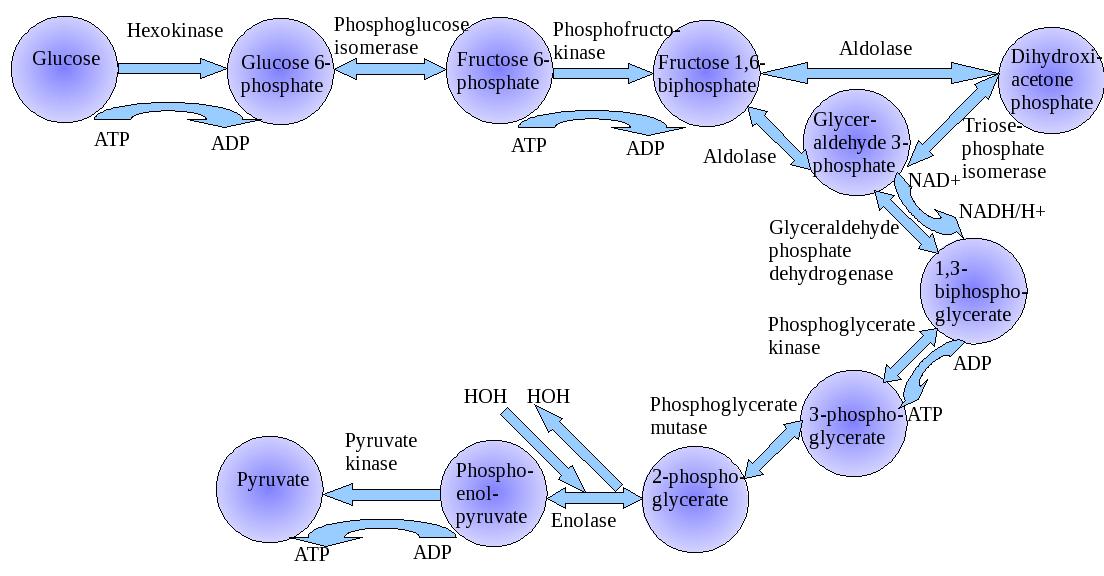
- glycolysis (enolase, pyruvate kinase, triose phosphate isomerase),
- fatty acid oxidation (carnitine palmitoyl transferase-1, mitochondrial thioesterase I),
and uncoupling protein-3,
effects that are achieved through enhanced transcription of TRE-containing genes
Skeletal muscle AMPK activation is characterized by
(i) being a rapid and transient response,
(ii) upstream activation by Ca2+-induced mobilization and CaMKKβ activation,
(iii) upstream upregulation of LKB1 expression, which requires association with STRAD
and MO25 for optimal phosphorylation/activation of AMPK, and
(iv) stimulation of mitochondrial fatty acid β-oxidation.
T3-induced muscle AMPK activation was found to trigger two major downstream
signaling pathways, namely,
(i) peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) mRNA
expression and phosphorylation, a transcriptional regulator for genes related to
- mitochondrial biogenesis,
- fatty acid oxidation, and
- gluconeogenesis and
(ii) cyclic AMP response element binding protein (CREB) phosphorylation, which
- in turn induces PGC-1α expression in liver tissue, thus
- reinforcing mechanism (i).
These data indicate that AMPK phosphorylation of PGC-1α initiates many of the
important gene regulatory functions of AMPK in skeletal muscle.
In heart, hyperthyroidism increased glycolysis and sarcolemmal GLUT4 levels by the
combined effects of AMPK activation and insulin stimulation, with concomitant increase
in fatty acid oxidation proportional to enhanced cardiac mass and contractile function.
Thyroid Hormone, AMPK Activation, and Liver Preconditioning
Recent studies by our group revealed that administration of a single dose of 0.1 mg T3/kg
to rats activates liver AMPK (Figure 4; unpublished work).
- enhancement in phosphorylated AMPK/nonphosphorylated AMPK ratios in T3-
treated rats over control values thatis significant in the time period of 1 to 48
hours after hormone treatment - Administration of a substantially higher dose (0.4 mg T3/kg) resulted in
decreased liver AMPK activation at 4 h to return to control values at 6 h
after treatment
Activation of liver AMPK by T3 may be of relevance in terms of
- promotion of fatty acid oxidation for ATP supply,
- supporting hepatoprotection against IR injury (Figure 3(C)).
This proposal is based on the high energy demands underlying effective liver
preconditioning for full operation of hepatic
- antioxidant, antiapoptotic, and anti-inflammatory mechanisms,
- oxidized biomolecules repair or resynthesis,
- induction of the homeostatic acute-phase response, and
- promotion of hepatocyte and Kupffer cell proliferation,
mechanisms that are needed to cope with the damaging processes set in by IR.
T3 liver preconditioning , in addition to that afforded by
- n-3 long-chain polyunsaturated fatty acids given alone or
- combined with T3 at lower dosages, or
- by iron supplementation,
constitutes protective strategies against hepatic IR injury.
Studies on the molecular mechanisms underlying T3-induced liver AMPK
activation (Figure 4) are currently under assessment in our laboratory.
References
Fernández and L. A. Videla, “Kupffer cell-dependent signaling in thyroid hormone
calorigenesis: possible applications for liver preconditioning,” Current Signal
Transduction Therapy 2009; 4(2): 144–151.
Viollet, B. Guigas, J. Leclerc et al., “AMP-activated protein kinase in the regulation
of hepatic energy metabolism: from physiology to therapeutic perspectives,” Acta
Physiologica 2009; 196(1): 81–98.
Carling, “The AMP-activated protein kinase cascade – A unifying system
for energy control,” Trends in Biochemical Sciences, 2004;. 29(1): 18–24.
E. Kemp, D. Stapleton, D. J. Campbell et al., “AMP-activated protein kinase,
super metabolic regulator,” Biochemical Society Transactions 2003; 31(1):
162–168.
G. Hardie, “AMP-activated protein kinase-an energy sensor that
regulates all ;aspects of cell function,” Genes and Development,
2011; 25(18): 1895–1908.
Woods, P. C. F. Cheung, F. C. Smith et al., “Characterization of AMP-activated
protein kinase βandγ subunits Assembly of the heterotrimeric complex in vitro,”
Journal of Biological Chemistry 1996;271(17): 10282–10290.
Xiao, R. Heath, P. Saiu et al., “Structural basis for AMP binding to mammalian AMP-
activated protein kinase,” Nature 2007; 449(7161): 496–500.
more…
Impact of Metformin and compound C on NIS expression and iodine uptake in vitro and in vivo: a role for CRE in AMPK modulation of thyroid function.
Abdulrahman RM1, Boon MR, Sips HC, Guigas B, Rensen PC, Smit JW, Hovens GC.
Author information
Thyroid. 2014 Jan;24(1):78-87. Epub 2013 Sep 25. PMID: 23819433
http://dx.doi.org:/10.1089/thy.2013.0041.
Although adenosine monophosphate activated protein kinase (AMPK) plays a crucial role
in energy metabolism, a direct effect of AMPK modulation on thyroid function has only
recently been reported, and much of its function in the thyroid is currently unknown.
The aim of this study was
- to investigate the mechanism of AMPK modulation in iodide uptake.
- to investigate the potential of the AMPK inhibitor compound C as an enhancer of
iodide uptake by thyrocytes.
Metformin reduced NIS promoter activity (0.6-fold of control), whereas compound C
stimulated its activity (3.4-fold) after 4 days. This largely coincides with
- CRE activation (0.6- and 3.0-fold).
These experiments show that AMPK exerts its effects on iodide uptake, at least partly,
through the CRE element in the NIS promoter. Furthermore, we have used AMPK-alpha1
knockout mice to determine the long-term effects of AMPK inhibition without chemical compounds.
These mice have a less active thyroid, as shown by reduced colloid volume and reduced
responsiveness to thyrotropin.
NIS expression and iodine uptake in thyrocytes
- can be modulated by metformin and compound C.
These compounds exert their effect by
- modulation of AMPK, which, in turn, regulates
- the activation of the CRE element in the NIS promoter.
Overall, this suggests that AMPK modulating compounds may be useful for the
enhancement of iodide uptake by thyrocytes, which could be useful for the
treatment of thyroid cancer patients with radioactive iodine.
AMPK: Master Metabolic Regulator
© 1996–2013 themedicalbiochemistrypage.org, LLC | info
@ themedicalbiochemistrypage.org

AMPK-activating drugs metformin or phenformin might provide protection against cancer 1741-7007-11-36-5

AMPK and AMPK-related kinase (ARK) family 1741-7007-11-36-4

AMP-activated protein kinase (AMPK) was first discovered as an activity that
- inhibited preparations of acetyl-CoA carboxylase (ACC) and 3-hydroxy-
3-methylglutaryl-CoA reductase (HMG-CoA reductase, HMGR) and was - induced by AMP.
AMPK induces a cascade of events within cells in response to the ever changing energy
charge of the cell. The role of AMPK in regulating cellular energy charge places this
enzyme at a central control point in maintaining energy homeostasis.
More recent evidence has shown that AMPK activity can also be regulated by physiological stimuli, independent of the energy charge of the cell, including hormones and nutrients.
Once activated, AMPK-mediated phosphorylation events
- switch cells from active ATP consumption (e.g. fatty acid and cholesterol
biosynthesis) to active ATP production (e.g. fatty acid and glucose oxidation).
These events are rapidly initiated and are referred to as
- short-term regulatory processes.
The activation of AMPK also exerts
- long-term effects at the level of both gene expression and protein synthesis.
Other important activities attributable to AMPK are
- regulation of insulin synthesis and
- secretion in pancreatic islet β-cells and
- modulation of hypothalamic functions involved in the regulation of satiety.
How these latter two functions impact obesity and diabetes will be discussed below.
Regulation of AMPK
In the presence of AMP the activity of AMPK is increased approximately 5-fold.
However, more importantly is the role of AMP in regulating the level of phosphorylation
of AMPK. An increased AMP to ATP ratio leads to a conformational change in the γ-subunit
leading to increased phosphorylation and decreased dephosphorylation of AMPK.
The phosphorylation of AMPK results in activation by at least 100-fold. AMPK is
phosphorylated by at least three different upstream AMPK kinases (AMPKKs).
Phosphorylation of AMPK occurs in the α subunit at threonine 172 (T172) which
- lies in the activation loop.
One kinase activator of AMPK is
- Ca2+-calmodulin-dependent kinase kinase β (CaMKKβ)
- which phosphorylates and activates AMPK in response to increased calcium.
The distribution of CaMKKβ expression is primarily in the brain with detectable levels
also found in the testes, thymus, and T cells. As described for the Ca2+-mediated
regulation of glycogen metabolism,
- increased release of intracellular stores of Ca2+ create a subsequent demand for
ATP.
Activation of AMPK in response to Ca fluxes
- provides a mechanism for cells to anticipate the increased demand for ATP.
Evidence has also demonstrated that the serine-threonine kinase, LKB1 (also called
serine-threonine kinase 11, STK11) which is encoded by the Peutz-Jeghers syndrome
tumor suppressor gene, is required for activation of AMPK in response to stress.
The active LKB1 kinase is actually a complex of three proteins:
- LKB1,
- Ste20-related adaptor (STRAD) and
- mouse protein 25 (MO25).
Thus, the enzyme complex is often referred to as LKB1-STRAD-MO25. Phosphorylation
of AMPK by LKB1 also occurs on T172. Unlike the limited distribution of CaMKKβ,
- LKB1 is widely expressed, thus making it the primary AMPK-regulating kinase.
Loss of LKB1 activity in adult mouse liver leads to
- near complete loss of AMPK activity and
- is associated with hyperglycemia.
The hyperglycemia is, in part, due to an increase in the transcription of gluconeogenic
genes. Of particular significance is the increased expression of
- the peroxisome proliferator-activated receptor-γ (PPAR-γ) coactivator 1α
(PGC-1α), which drives gluconeogenesis. - Reduction in PGC-1α activity results in normalized blood glucose levels in
LKB1-deficient mice.
The third AMPK phosphorylating kinase is transforming growth factor-β-activated
kinase 1 (TAK1). However, the normal physiological conditions under which TAK1
phosphorylates AMPK are currently unclear.
The effects of AMP are two-fold:
- a direct allosteric activation and making AMPK a poorer substrate for
dephosphorylation.
Because AMP affects both
the rate of AMPK phoshorylation in the positive direction and
dephosphorylation in the negative direction,
the cascade is ultrasensitive. This means that
- a very small rise in AMP levels can induce a dramatic increase in the activity of
AMPK.
The activity of adenylate kinase, catalyzing the reaction shown below, ensures that
- AMPK is highly sensitive to small changes in the intracellular [ATP]/[ADP] ratio.
2 ADP ——> ATP + AMP
Negative allosteric regulation of AMPK also occurs and this effect is exerted by
phosphocreatine. As indicated above, the β subunits of AMPK have a glycogen-binding domain, GBD. In muscle, a high glycogen content
- represses AMPK activity and
- this is likely the result of interaction between the GBD and glycogen,
- the GBD of AMPK allows association of the enzyme with the regulation of glycogen metabolism
- by placing AMPK in close proximity to one of its substrates glycogen synthase.
AMPK has also been shown to be activated by receptors that are coupled to
- phospholipase C-β (PLC-β) and by
- hormones secreted by adipose tissue (termed adipokines) such as leptinand adiponectin (discussed below).
Targets of AMPK
The signaling cascades initiated by the activation of AMPK exert effects on
- glucose and lipid metabolism,
- gene expression and
- protein synthesis.
These effects are most important for regulating metabolic events in the liver, skeletal
muscle, heart, adipose tissue, and pancreas.
Demonstration of the central role of AMPK in the regulation of metabolism in response
to events such as nutrient- or exercise-induced stress. Several of the known physiologic
targets for AMPK are included as well as several pathways whose flux is affected by
AMPK activation. Arrows indicate positive effects of AMPK, whereas, T-lines indicate
the resultant inhibitory effects of AMPK action.
The uptake, by skeletal muscle, accounts for >70% of the glucose removal from the
serum in humans. Therefore, it should be obvious that this event is extremely important
for overall glucose homeostasis, keeping in mind, of course, that glucose uptake by
cardiac muscle and adipocytes cannot be excluded from consideration. An important fact
related to skeletal muscle glucose uptake is that this process is markedly impaired in
individuals with type 2 diabetes.
The uptake of glucose increases dramatically in response to stress (such as ischemia) and
exercise and is stimulated by insulin-induced recruitment of glucose transporters
to the plasma membrane, primarily GLUT4. Insulin-independent recruitment of glucose
transporters also occurs in skeletal muscle in response to contraction (exercise).
The activation of AMPK plays an important, albeit not an exclusive, role in the induction of
GLUT4 recruitment to the plasma membrane. The ability of AMPK to stimulate
GLUT4 translocation to the plasma membrane in skeletal muscle is by a different mechanism
than that stimulated by insulin and insulin and AMPK effects are additive.
Under ischemic/hypoxic conditions in the heart the activation of AMPK leads to the
phosphorylation and activation of the kinase activity of phosphofructokinase-2, PFK-2
(6-phosphofructo-2-kinase). The product of the action of PFK-2 (fructose-2,6-bisphosphate,
F2,6BP) is one of the most potent regulators of the rate of flux through
glycolysis and gluconeogenesis.
In liver the PKA-mediated phosphorylation of PFK-2 results in conversion of the
enzyme from a kinase that generates F2,6BP to a phosphatase that removes the
2-phosphate thus reducing the levels of the potent allosteric activator of the glycolytic
enzyme 6-phosphfructo-1-kinase, PFK-1 and the potent allosteric inhibitor
of the gluconeogenic enzyme fructose-1,6-bisphosphatase (F1,-6BPase).
It is important to note that like many enzymes, there are multiple isoforms of PFK-2
(at least 4) and neither the liver or the skeletal muscle isoforms contain the AMPK
phosphorylation sites found in the cardiac and inducible (iPFK2) isoforms of PFK-2.
Inducible PFK-2 is expressed in the monocyte/macrophage lineage in response to pro-
inflammatory stimuli. The ability to activate the kinase activity by phosphorylation of
PFK-2 in cardiac tissue and macrophages in response to ischemic conditions allows these
cells to continue to have a source of ATP via anaerobic glycolysis. This phenomenon is
recognized as the Pasteur effect: an increased rate of glycolysis in response to hypoxia.
Of pathological significance is the fact that the inducible form of PFK-2 is commonly
expressed in many tumor cells and this may allow AMPK to play an important role in
protecting tumor cells from hypoxic stress. Indeed, techniques for depleting AMPK in
tumor cells have shown that these cells become sensitized to nutritional stress upon loss
of AMPK activity.
Whereas, stress and exercise are powerful inducers of AMPK activity in skeletal muscle,
additional regulators of its activity have been identified.
Insulin-sensitizing drugs of the thiazolidinedione family (activators of PPAR-γ, see
below) as well as the hypoglycemia drug metformin exert a portion of their effects
through regulation of the activity of AMPK.
As indicated above, the activity of the AMPK activating kinase, LKB1, is critical for
regulating gluconeogenic flux and consequent glucose homeostasis. The action of
metformin in reducing blood glucose levels
- requires the activity of LKB1 in the liver for this function.
Also, several adipokines (hormones secreted by adipocytes) either stimulate or inhibit
AMPK activation:
- leptin and adiponectin have been shown to stimulate AMPK activation, whereas,
- resistininhibits AMPK activation.
Cardiac effects exerted by activation of AMPK also include
- phosphorylation of endothelial nitric oxide synthase, eNOSin cardiac endothelium.
AMPK-mediated phosphorylation of eNOS leads to increased activity and consequent
NO production and provides a link between metabolic stresses and cardiac function.
In platelets, insulin action leads to an increase in eNOS activity that is
- due to its phosphorylation by AMPK.
Activation of NO production in platelets leads to
- a decrease in thrombin-induced aggregation, thereby,
- limiting the pro-coagulant effects of platelet activation.
The response of platelets to insulin function clearly indicates why disruption in insulin
action is a major contributing factor in the development of the metabolic syndrome
Activation of AMPK leads to a reduction in the level of SREBP
- a transcription factor ®ulator of the expression of numerous
lipogenic enzymes
Another transcription factor reduced in response to AMPK activation is
- hepatocyte nuclear factor 4α, HNF4α
- a member of the steroid/thyroid hormone superfamily.
- HNF4α is known to regulate the expression of several liver and
pancreatic β-cell genes such as GLUT2, L-PK and preproinsulin.
- Of clinical significance is that mutations in HNF4α are responsible for
- maturity-onset diabetes of the young, MODY-1.
Recent evidence indicates that the gene for the carbohydrate-response-element-
binding protein (ChREBP) is a target for AMPK-mediated transcriptional regulation
in the liver. ChREBP is rapidly being recognized as a master regulator of lipid
metabolism in liver, in particular in response to glucose uptake.
The target of the thiazolidinedione (TZD) class of drugs used to treat type 2 diabetes is
the peroxisome proliferator-activated receptor γ, PPARγ which
- itself may be a target for the action of AMPK.
The transcription co-activator, p300, is phosphorylated by AMPK
- which inhibits interaction of p300 with not only PPARγ but also
- the retinoic acid receptor, retinoid X receptor, and
- thyroid hormone receptor.
PPARγ is primarily expressed in adipose tissue and thus it was difficult to reconcile how
a drug that was apparently acting only in adipose tissue could lead to improved insulin
sensitivity of other tissues. The answer to this question came when it was discovered that the TZDs stimulated the expression and release of the adipocyte hormone (adipokine),
adiponectin. Adiponectin stimulates glucose uptake and fatty acid oxidation in skeletal
muscle. In addition, adiponectin stimulates fatty acid oxidation in liver while inhibiting
expression of gluconeogenic enzymes in this tissue.
These responses to adiponectin are exerted via activation of AMPK. Another
transcription factor target of AMPK is the forkhead protein, FKHR (now referred to as
FoxO1). FoxO1 is involved in the activation of glucose-6-phosphatase expression and,
therefore, loss of FoxO1 activity in response to AMPK activation will lead to reduced
hepatic output of glucose.
This concludes a very complicated perspective that ties together the thyroid hormone
activity, the hypophysis, diabetes mellitus, and AMPK tegulation of metabolism in the
liver, skeletal muscle, adipose tissue, and heart. I also note at this time that there
nongenetic points to be made here:
- The tissue specificity of isoenzymes
- The modulatory role of AMP:ATP ratio in phosphorylation/dephosphorylation
effects on metabolism tied to AMPK - The tie in of stress or ROS with fast reactions to protect harm to tissues
- The relationship of cytokine activation and release to the above metabolic events
- The relationship of effective and commonly used diabetes medications to AMPK
mediated processes - The preceding presentation is notable for the importance of proteomic and
metabolomic invetigations in elucidation common chronic and nongenetic diseases







































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}