Healthcare analytics, AI solutions for biological big data, providing an AI platform for the biotech, life sciences, medical and pharmaceutical industries, as well as for related technological approaches, i.e., curation and text analysis with machine learning and other activities related to AI applications to these industries.
Research investigators at Wayne State University in collaboration with La Jolla Institute for Allergy and Immunology (LJAI) are developing a T-cell peptide-based vaccine for cardiovascular disease, specifically, to reduce immune-based inflammatory plaques in arteries. The scientists published their findings in the December 2013 issue of Frontiers in Immunology, titled “Atheroprotective vaccination with MCH-II restricted peptides from Apo B-100.” These experiments show proof of concept for the development of an autoantigen-specific vaccine for reducing the amount of atherosclerotic plaques in mice.
The published work was done in the laboratory of Klaus Ley, M.D., a prominent vascular biolist of LIAI based on the discovery by Harley Tse, Ph.D., Professor of immunology and microbiology at Wayne Stae University School of Medicine, and Wayne State’s Cardiovascular Research Institute with Michael Shae, Ph.D., adjunct assistant professor of immunology and microbiology.Shaw and Tse are the first to demonstrate that two T-cell epitopes of the autoantigen apoB100 are deeply involved in the development of the disease. The discovery is reported in J Immunol Clin Res Apr-Jun, 2014; 2: “Identification of two immunogenic T cell epitopes of ApoB100 and their Autoimmune Implications.”
Atheroprotective Vaccination with MHC-II Restricted Peptides from ApoB-100.
BACKGROUND: Subsets of CD4(+) T-cells have been proposed to serve differential roles in the development of atherosclerosis. Some T-cell types are atherogenic (T-helper type 1), while others are thought to be protective (regulatory T-cells). Lineage commitment toward one type of helper T-cell versus another is strongly influenced by the inflammatory context in which antigens are recognized. Immunization of atherosclerosis-prone mice with low-density lipoprotein (LDL) or its oxidized derivative (ox-LDL) is known to be atheroprotective. However, the antigen specificity of the T-cells induced by vaccination and the mechanism of protection are not known.
METHODS: Identification of two peptide fragments (ApoB3501-3516 and ApoB978-993) from murine ApoB-100 was facilitated using I-Ab prediction models, and their binding to I-Ab determined. Utilizing a vaccination scheme based on complete and incomplete Freund’s adjuvant (CFA and IFA) [1 × CFA + 4 × IFA], we immunized Apoe(-/-)mice with ApoB3501-3516 or ApoB978-993 emulsified in CFA once and subsequently boosted in IFA four times over 15 weeks. Spleens, lymph nodes, and aortas were harvested and evaluated by flow cytometry and real time RT-PCR. Total atherosclerotic plaque burden was determined by aortic pinning and by aortic root histology.
RESULTS: Mice immunized with ApoB3501-3516 or ApoB978-993 demonstrated 40% reduction in overall plaque burden when compared to adjuvant-only control mice. Aortic root frozen sections from ApoB3501-3516 immunized mice showed a >60% reduction in aortic sinus plaque development. Aortas from both ApoB3501-3516 and ApoB978-993 immunized mice contained significantly more mRNA for IL-10. Both antigen-specific IgG1 and IgG2c titers were elevated in ApoB3501-3516 or ApoB978-993 immunized mice, suggesting helper T-cell immune activity after immunization.
CONCLUSION: Our data show that MHC Class II restricted ApoB-100 peptides can be atheroprotective, potentially through a mechanism involving elevated IL-10.
Atherosclerosis is decreased in ApoB3501–3516 and ApoB978–993-treated mice compared to controls. (A) Vaccination schedule: 8-week-old female Apoe−/− mice were immunized once with either PBS or peptide in CFA, then boosted four more times with PBS or peptide in IFA. WD was maintained for 13 weeks. Mice were sacrificed and organs harvested at 23 weeks of age. (B,C) Results of aortic pinning analysis after Sudan IV staining are shown with representative photographs. N = 12–15 in each group, *p < 0.05 when compared to 1× CFA + 4× IFA group. (D) Representative aortic root staining sections after ORO staining, counter-stained with hematoxylin. (E) Plaque area from aortic roots stained from each group. Lesion sizes from 30 to 40 μm distal to start of the aortic valve were averaged per group. N = 5 in each group, *p < 0.05 when compared to 1× CFA + 1× IFA control group.
Inhibition of T cell response to native low density lipoprotein reduces atherosclerosis
Atherosclerosis is a chronic inflammatory disease in which lipoproteins accumulate, eliciting an inflammatory response in the arterial wall. Adaptive immune responses that engage clonally expanded T cell populations contribute to this process, as do innate immune responses that are mounted by macrophages and other cells. Several studies have suggested that components of low-density lipoprotein (LDL) particles trigger vascular inflammation (Tabas et al., 2007; Hartvigsen et al., 2009).
As a consequence of oxidation, the double bonds of fatty acid residues in phospholipids, cholesteryl esters, and triglycerides are cleaved, thus generating reactive aldehydes and truncated lipids (Esterbauer et al., 1990). Among the latter, modified phospholipids, such as lysophosphatidylcholine and oxidized 1-palmitoyl-2-arachidonyl-sn-glycero-3-phosphocholine (ox-PAPC), induce endothelial cells, macrophages, and B1-type B cells to initiate innate immune responses, effecting adhesion molecule expression, chemokine production, and secretion of natural antibodies containing germline IgM sequences (Leitinger et al., 1997; Binder et al., 2004; Gharavi et al., 2007).
Immune responses to oxidized low-density lipoprotein (oxLDL) are proposed to be important in atherosclerosis. To identify the mechanisms of recognition that govern T cell responses to LDL particles, we generated T cell hybridomas from human ApoB100 transgenic (huB100tg) mice that were immunized with human oxLDL. Surprisingly, none of the hybridomas responded to oxidized LDL, only to native LDL and the purified LDL apolipoprotein ApoB100.
However, sera from immunized mice contained IgG antibodies to oxLDL, suggesting that T cell responses to native ApoB100 help B cells making antibodies to oxLDL. ApoB100 responding CD4+ T cell hybridomas were MHC class II–restricted and expressed a single T cell receptor (TCR) variable (V) chain, TRBV31, with different V chains. Immunization of huB100tgxLdlr/ mice with a TRBV31-derived peptide induced anti-TRBV31 antibodies that blocked T cell recognition of ApoB100. This treatment significantly reduced atherosclerosis by 65%, with a concomitant reduction of macrophage infiltration and MHC class II expression in lesions. In conclusion, CD4+ T cells recognize epitopes on native ApoB100 protein, this response is associated with a limited set of clonotypic TCRs, and blocking TCR-dependent antigen recognition by these T cells protects against atherosclerosis.
Impact of multiple antigenic epitopes from ApoB100, hHSP60 and Chlamydophila pneumoniae on atherosclerotic lesion development in Apobtm2SgyLdlrtm1HerJ mice
► We produced 5 constructs using dendroaspin as a scaffold for immunization study. ► All constructs have the effect on lesion reduction. ► Modulation in atherosclerosis-related autoimmunity appears by Tregs.
Atherosclerosis is increasingly recognized as a complex chronic inflammatory disease of the arterial walls [1], [2] and [3], as evidenced by the presence of inflammatory cells, activated immune cells and cytokines in lesions, all of which indicate involvement of the immune system. Atherosclerotic plaques are known to contain macrophage-derived foam cells in which macrophages interact with T-cells to produce a wide array of cytokines that can exert both pro- and anti-inflammatory effects.
Antibodies against aldehyde-modified ApoB100, a major constituent of low-density lipoprotein, reduce atherosclerosis in mice expressing human ApoB100, suggesting an immunogenic role of ApoB100. Antibodies against epitopes of the human heat shock protein 60 (hHSP60) molecule (hHSP60153–163: AELKKQSKPVT and hHSP60303-312: PGFGDNRKNQ) are present in atherosclerotic patients and share considerable homology with human cytomegalovirus (HCMV)-derived protein (immediate early protein UL122) and Porphyromonas gingivalis microbial HSP60. Sequence homology between microbial HSP60 and hHSP60 has been suggested to result in immunological cross-reactivity, which may play a role in atherogenesis. Titers of Cpn antibodies are not always positively associated with the Cpn organism in atheroma; however, these antibodies might exert cross-reactivity to non-Cpn antigens.
Immunization of mice with a single construct containing multiple epitopes derived from ApoB100, hHSP60 and Cpn was more effective in reducing early atherosclerotic lesions through the induction of a specific Treg-cell response than was the construct containing either mono- or bi-epitopes. This approach offers attractive opportunities for the design of protein-based, multivalent vaccines against atherosclerosis.
Immunization with a combination of ApoB and HSP60 epitopes significantly reduces early atherosclerotic lesion in Apobtm2SgyLdlrtm1Her/J mice
Objective: HSP60 is emerging as an immune-dominant target of autoantibodies in atherosclerosis and recent studies have revealed oxLDL as a key antigen in the development of atherosclerosis. In this study, we assay whether immunizing Apobtm2SgyLdlrtm1Her/J mice with a combination of ApoB and human HSP60 peptides has an additive effect on athero-protection compared to ApoB or HSP60 peptides applied alone by following atherosclerotic lesion development. Methods and results: In this study, 2 weeks after the first immunization, Apobtm2SgyLdlrtm1Her/J mice were placed on a high-fat diet for 8 weeks followed by 2 weeks on a normal diet allowing the mice to adapt to the environment before sacrifice. High levels of ApoB and HSP60 antibodies were detectable in week 2 and week 12 following the first immunization with KLH-conjugated ApoB and HSP60 peptides either individually or in combination. Histological analyses demonstrated that mice immunized with both, ApoB and HSP60 peptides, showed the most significant reduction in atherosclerotic lesions (41.3%; p < 0.001) compared to a reduction of 14.7% (p < 0.05) and 21.1% (p < 0.01) in mice immunized with ApoB or HSP60 peptides, respectively; control mice were immunized with either PBS or adjuvant alone. These results
were further supported by significant differences in the cellular and humoral immune responses between test animals. Conclusions: Immunization with a combination of ApoB and HSP60 peptide antigens significantly reduced early atherosclerotic lesions in the Apobtm2SgyLdlrtm1Her/J mouse model of atherosclerosis. This approach offers promise as a novel strategy for developing anti-atherosclerotic agents.
Chlamydophila (Chlamydia) pneumoniae infection promotes vascular smooth muscle cell adhesion and migration through IQ domain GTPase-activating protein 1
► C. pneumoniae infection increases the adhesion of vascular smooth muscle cells. ► C. pneumoniae infection promotes the migration of vascular smooth muscle cells. ► IQGAP1 expression was increased in the infected vascular smooth muscle cells. ► Depletion of IQGAP1 inhibits the infection-induced cell adhesion and migration.
The mechanisms for Chlamydophila (Chlamydia) pneumoniae (C. pneumoniae) infection-induced atherosclerosis are still unclear. Cell adhesion has important roles in vascular smooth muscle cell (VSMC) migration required in the development of atherosclerosis. However, it is still unknown whether IQ domain GTPase-activating protein 1 (IQGAP1) plays pivotal roles in C. pneumoniae infection-induced the adhesion and migration of rat primary VSMCs. Accordingly, in this study, we demonstrated that rat primary VSMC adhesion (P < 0.001) and migration (P < 0.01) measured by cell adhesion assay and Transwell assay, respectively, were significantly enhanced after C. pneumoniae infection. Reverse transcription-polymerase chain reaction analysis revealed that the mRNA expression levels of IQGAP1 in the infected rat primary VSMCs were found to increase gradually to reach a peak and then decrease gradually to a level similar to the control. We further showed that the increases in rat primary VSMC adhesion to Matrigel (P < 0.001) and migration (P < 0.01) caused by C. pneumoniae infection were markedly inhibited after IQGAP1 knockdown by a pool of four short hairpin RNAs. Taken together, our results suggest that C. pneumoniae infection may promote the adhesion and migration of VSMCs possibly by upregulating the IQGAP1 expression.
Rosiglitazone negatively regulates c-Jun N-terminal kinase and toll-like receptor 4 proinflammatory signalling during initiation of experimental aortic aneurysms
► Rosiglitazone has a marked effect on both aneurysm rupture and development. ► Rosiglitazone modulates inflammation by blocking TLR4/JNK signalling. ► Specific antagonists of JNK and TLR4 may be therapeutic for aneurysms.
Development and rupture of aortic aneurysms (AA) is a complex process involving inflammation, cell death, tissue and matrix remodelling. The thiazolidinediones (TZDs) including Rosiglitazone (RGZ) are a family of drugs which act as agonists of the nuclear peroxisome proliferator-activated receptors and have a broad spectrum of effects on a number of biological processes in the cardiovascular system. In our previous study we have demonstrated that RGZ has a marked effect on both aneurysm rupture and development, however, the precise mechanism of this is unknown.
Methods and results In the present study, we examined possible targets of RGZ action in the early stages of Angiotensin II-induced AA in apolipoprotein E-deficient mice. For this purpose we employed immunoblotting, ELISA and antibody array approaches. We found that RGZ significantly inhibited c-Jun N-terminal kinase (JNK) phosphorylation and down-regulated toll-like receptor 4 (TLR4) expression at the site of lesion formation in response to Angiotensin II infusion in the initiation stage (6–72 h) of experimental AA development. Importantly, this effect was also associated with a decrease of CD4 antigen and reduction in production of TLR4/JNK-dependant proinflammatory chemokines MCP-1 and MIP-1α. Conclusion These data suggest that RGZ can modulate inflammatory processes by blocking TLR4/JNK signalling in initiation stages of AA development.
Atheroprotective immunization with malondialdehyde-modified LDL is hapten specific and dependent on advanced MDA adducts: implications for development of an atheroprotective vaccine.
Gonen A, Hansen LF, Turner WW, Montano EN, Que X,…, Hartvigsen K.
J Lipid Res. 2014 Oct;55(10):2137-55. http://dx.doi.org:/10.1194/jlr.M053256. Epub 2014 Aug 20.
Immunization with homologous malondialdehyde (MDA)-modified LDL (MDA-LDL) leads to atheroprotection in experimental models supporting the concept that a vaccine to oxidation-specific epitopes (OSEs) of oxidized LDL could limit atherogenesis. However, modification of human LDL with OSE to use as an immunogen would be impractical for generalized use. Furthermore, when MDA is used to modify LDL, a wide variety of related MDA adducts are formed, both simple and more complex. To define the relevant epitopes that would reproduce the atheroprotective effects of immunization with MDA-LDL, we sought to determine the responsible immunodominant and atheroprotective adducts. We now demonstrate that fluorescent adducts of MDA involving the condensation of two or more MDA molecules with lysine to form malondialdehyde-acetaldehyde (MAA)-type adducts generate immunodominant epitopes that lead to atheroprotective responses. We further demonstrate that a T helper (Th) 2-biased hapten-specific humoral and cellular response is sufficient, and thus, MAA-modified homologous albumin is an equally effective immunogen. We further show that such Th2-biased humoral responses per se are not atheroprotective if they do not target relevant antigens. These data demonstrate the feasibility of development of a small-molecule immunogen that could stimulate MAA-specific immune responses, which could be used to develop a vaccine approach to retard or prevent atherogenesis.
Low density lipoprotein oxidation and atherogenesis: from experimental models to clinical studies.
Napoli C
G Ital Cardiol. 1997 Dec; 27(12):1302-14.
Oxidative modifications of low-density lipoproteins (LDL) (“oxidation hypothesis”) appears to be the pathophysiologic mechanism implicated in early atherogenesis. Oxidized LDL (ox-LDL) may also induce several pro-atherogenic mechanisms, such as the regulation of vascular tone, by interfering with nitric oxide, the stimulation of cytokines and chemotactic factors (MCP-1, M-CSF, VCAM-1, etc.) and transcription factors (AP1 and NFk beta). These phenomena complicate the spectrum of direct and indirect actions of ox-LDL. The immunogenicity of ox-LDL was used to generate monoclonal antibodies against many epitopes of ox-LDL, such as malondialdehyde-lysine (MDA-2) or 4-hydroxynonenal-lysine (NA59). These antibodies showed the occurrence of ox-LDL in vivo. Another issue is the role of the humoral and cellular immune system in atherogenesis, in particular whether the immune response to ox-LDL enhances or reduces early atherogenesis. Moreover, the induction of autoantibodies against ox-LDL and the recognition by “natural” antibodies, and the use of the antigens to screen human sera may serve as a marker of atherosclerosis. In this review, we have stressed the importance of methodologic approach in the assessment of LDL-oxidation and the fact that lipoprotein (a) may also undergo oxidative modifications. Several clinical conditions are associated with increased rate of LDL-oxidation. Recently, we have observed the presence of LDL oxidation-specific epitopes in human fetal aortas. Antioxidants studies in primary prevention of atherosclerosis have produced contradictory results. This may be explained in part by the selection of patients who had advanced lesions and were often smokers. New trails suggest that antioxidants be administered early in children. Lastly, antioxidant studies in the secondary prevention of coronary heart disease (CHAOS, WACS, and HOPE) show clear evidence of the benefits of antioxidants in reducing new cardiovascular events.
Atherosclerosis is a chronic inflammatory disease and immunological mechanisms are of central importance. It is known that oxidized LDL and its oxidized moieties were a major class of immunodominant epitopes within the atherosclerotic plaque. Oxidation of LDL leads to the generation of a variety of oxidized lipids and oxidized lipid-apo-B adducts.
Technology Description
UC San Deigo researchers proposed that an immunization strategy could be used to inhibit the progression of atherosclerosis by showing that immunization of rabbits and/or mice (and ultimately humans) with MDA-LDL could inhibit atherosclerosis. To develop a safe vaccine for human use would require the identification of the specific immunogenic oxidation-specific epitope(s) that provides the atheroprotective immunity. Until now, the mechanism of the protection, that is, the immunodominant epitope(s) has not yet been determined.
UC San Diego researchers have been able to identify a small group of MDA-derived adducts which are immunodominant and atheroprotective in mice following immunization. The invention described here has the potential to provide an antigen to formulate a wholly synthetic vaccine to inhibit the development of atherosclerosis in man. Furthermore, in vivo levels of the adducts, and the autoantibodies recognizing them, may be used as diagnostic tools in patients with cardiovascular and other inflammatory diseases.
State Of Development
Mice have been immunized with the adducts resulting in atheroprotection. Techniques are currently being developed for a totally synthetic immunogen suitable for human clinical studies. Assays are also being developed.
Intellectual Property Info
A patent application has been filed on this technology.
Introduction – The Evolution of Cancer Therapy and Cancer Research: How We Got Here?
Author and Curator: Larry H Bernstein, MD, FCAP
The evolution of progress we have achieved in cancer research, diagnosis, and therapeutics has originated from an emergence of scientific disciplines and the focus on cancer has been recent. We can imagine this from a historical perspective with respect to two observations. The first is that the oldest concepts of medicine lie with the anatomic dissection of animals and the repeated recurrence of war, pestilence, and plague throughout the middle ages, and including the renaissance. In the awakening, architecture, arts, music, math, architecture and science that accompanied the invention of printing blossomed, a unique collaboration of individuals working in disparate disciplines occurred, and those who were privileged received an education, which led to exploration, and with it, colonialism. This also led to the need to increasingly, if not without reprisal, questioning long-held church doctrines.
It was in Vienna that Rokitansky developed the discipline of pathology, and his student Semelweis identified an association between then unknown infection and childbirth fever. The extraordinary accomplishments of John Hunter in anatomy and surgery came during the twelve years war, and his student, Edward Jenner, observed the association between cowpox and smallpox resistance. The development of a nursing profession is associated with the work of Florence Nightengale during the Crimean War (at the same time as Leo Tolstoy). These events preceded the work of Pasteur, Metchnikoff, and Koch in developing a germ theory, although Semelweis had committed suicide by infecting himself with syphilis. The first decade of the Nobel Prize was dominated by discoveries in infectious disease and public health (Ronald Ross, Walter Reed) and we know that the Civil War in America saw an epidemic of Yellow Fever, and the Armed Services Medical Museum was endowed with a large repository of osteomyelitis specimens. We also recall that the Russian physician and playwriter, Anton Checkov, wrote about the conditions in prison camps.
But the pharmacopeia was about to open with the discoveries of insulin, antibiotics, vitamins, thyroid action (Mayo brothers pioneered thyroid surgery in the thyroid iodine-deficient midwest), and pitutitary and sex hormones (isolatation, crystal structure, and synthesis years later), and Karl Landsteiner’s discovery of red cell antigenic groups (but he also pioneered in discoveries in meningitis and poliomyelitis, and conceived of the term hapten) with the introduction of transfusion therapy that would lead to transplantation medicine. The next phase would be heralded by the discovery of cancer, which was highlighted by the identification of leukemia by Rudolph Virchow, who cautioned about the limitations of microscopy. This period is highlighted by the classic work – “Microbe Hunters”.
John Hunter
Walter Reed
Robert Koch
goldberger 1916 Pellagra
Louis Pasteur
A multidisciplinary approach has led us to a unique multidisciplinary or systems view of cancer, with different fields of study offering their unique expertise, contributions, and viewpoints on the etiology of cancer. Diverse fields in immunology, biology, biochemistry, toxicology, molecular biology, virology, mathematics, social activism and policy, and engineering have made such important contributions to our understanding of cancer, that without cooperation among these diverse fields our knowledge of cancer would never had evolved as it has. In a series of posts “Heroes in Medical Research:” the work of researchers are highlighted as examples of how disparate scientific disciplines converged to produce seminal discoveries which propelled the cancer field, although, at the time, they seemed like serendipitous findings. In the post Heroes in Medical Research: Barnett Rosenberg and the Discovery of Cisplatin (Translating Basic Research to the Clinic) discusses the seminal yet serendipitous discoveries by bacteriologist Dr. Barnett Rosenberg, which eventually led to the development of cisplatin, a staple of many chemotherapeutic regimens. Molecular biologist Dr. Robert Ting, working with soon-to-be Nobel Laureate virologist Dr. James Gallo on AIDS research and the associated Karposi’s sarcoma identified one of the first retroviral oncogenes, revolutionizing previous held misconceptions of the origins of cancer (described in Heroes in Medical Research: Dr. Robert Ting, Ph.D. and Retrovirus in AIDS and Cancer). Located here will be a MONTAGE of PHOTOS of PEOPLE who made seminal discoveries and contributions in every field to cancer Each of these paths of discovery in cancer research have led to the unique strategies of cancer therapeutics and detection for the purpose of reducing the burden of human cancer. However, we must recall that this work has come at great cost, while it is indeed cause for celebration. The current failure rate of clinical trials at over 70 percent, has been a cause for disappointment, and has led to serious reconsideration of how we can proceed with greater success. The result of the evolution of the cancer field is evident in the many parts and chapters of this ebook. Volume 4 contains chapters that are in a predetermined order:
The concepts of neoplasm, malignancy, carcinogenesis, and metastatic potential, which encompass:
(a) How cancer cells bathed in an oxygen rich environment rely on anaerobic glycolysis for energy, and the secondary consequences of cachexia and sarcopenia associated with progression
invasion
ARTS protein and cancer
Glycolysis
Krebs cycle
Metabolic control analysis of respiration in human cancer tissue
akip1-expression-modulates-mitochondrial-function
(b) How advances in genetic analysis, molecular and cellular biology, metabolomics have expanded our basic knowledge of the mechanisms which are involved in cellular transformation to the cancerous state.
nucleotides
Methylation of adenine
ampk-and-ampk-related-kinase-ark-family-
ubiquitylation
(c) How molecular techniques continue to advance our understanding of how genetics, epigenetics, and alterations in cellular metabolism contribute to cancer and afford new pathways for therapeutic intervention.
genomic effects
LKB1AMPK pathway
mutation-frequencies-across-12-cancer-types
AMPK-activating drugs metformin or phenformin might provide protection against cancer
2. The distinct features of cancers of specific tissue sites of origin
3. The diagnosis of cancer by
(a) Clinical presentation
(b) Age of onset and stage of life
(c) Biomarker features
hairy cell leukemia
lymphoma leukemia
(d) Radiological and ultrasound imaging
Treatments
Prognostic differences within and between cancer types
We have introduced the emergence of a disease of great complexity that has been clouded in more questions than answers until the emergence of molecular biology in the mid 20th century, and then had to await further discoveries going into the 21st century. What gave the research impetus was the revelation of
1 the mechanism of transcription of the DNA into amino acid sequences
Proteins in Disease
2 the identification of stresses imposed on cellular function
NO beneficial effects
3 the elucidation of the substructure of the cell – cell membrane, mitochondria, ribosomes, lysosomes – and their functions, respectively
AKIP1 Expression Modulates Mitochondrial Function
4 the elucidation of oligonucleotide sequences
nucleotides
dna-replication-unwinding
dna-replication-ligation
dna-replication-primer-removal
dna-replication-leading-strand
dna-replication-lagging-strand
dna-replication-primer-synthesis
dna-replication-termination
5 the further elucidation of functionally relevant noncoding lncDNA
6 the technology to synthesis mRNA and siRNA sequences
Figure. RNAi and gene silencing
7 the repeated discovery of isoforms of critical enzymes and their pleiotropic properties
8. the regulatory pathways involved in signaling
Figure. Signaling Pathways Map
This is a brief outline of the modern progression of advances in our understanding of cancer. Let us go back to the beginning and check out a sequence of Nobel Prizes awarded and related discoveries that have a historical relationship to what we know. The first discovery was the finding by Louis Pasteur that fungi that grew in an oxygen poor environment did not put down filaments. They did not utilize oxygen and they produced used energy by fermentation. This was the basis for Otto Warburg sixty years later to make the comparison to cancer cells that grew in the presence of oxygen, but relied on anaerobic glycolysis. He used a manometer to measure respiration in tissue one cell layer thick to measure CO2 production in an adiabatic system.
Lavoisier Antoine-Laurent and Laplace Pierre-Simon (1783) Memoir on heat. Mémoirs de l’Académie des sciences. Translated by Guerlac H, Neale Watson Academic Publications, New York, 1982.
The Warburg apparatus is a manometric respirometer which was used for decades in biochemistry for measuring oxygen consumption of tissue homogenates or tissue slices.
The aqueous phase is vigorously shaken to equilibrate with a gas phase, from which oxygen is consumed while the evolved carbon dioxide is trapped, such that the pressure in the constant-volume gas phase drops proportional to oxygen consumption. The Warburg apparatus was introduced to study cell respiration, i.e. the uptake of molecular oxygen and the production of carbon dioxide by cells or tissues. Its applications were extended to the study of fermentation, when gas exchange takes place in the absence of oxygen. Thus the Warburg apparatus became established as an instrument for both aerobic and anaerobic biochemical studies [2, 3].
The respiration chamber was a detachable glass flask (F) equipped with one or more sidearms (S) for additions of chemicals and an open connection to a manometer (M; pressure gauge). A constant temperature was provided by immersion of the Warburg chamber in a constant temperature water bath. At thermal mass transfer equilibrium, an initial reading is obtained on the manometer, and the volume of gas produced or absorbed is determined at specific time intervals. A limited number of ‘titrations’ can be performed by adding the liquid contained in a side arm into the main reaction chamber. A Warburg apparatus may be equipped with more than 10 respiration chambers shaking in a common water bath. Since temperature has to be controlled very precisely in a manometric approach, the early studies on mammalian tissue respiration were generally carried out at a physiological temperature of 37 °C.
The Warburg apparatus has been replaced by polarographic instruments introduced by Britton Chance in the 1950s. Since Chance and Williams (1955) measured respiration of isolated mitochondria simultaneously with the spectrophotometric determination of cytochrome redox states, a water chacket could not be used, and measurements were carried out at room temperature (or 25 °C). Thus most later studies on isolated mitochondria were shifted to the artifical temperature of 25 °C.
Today, the importance of investigating mitochondrial performance at in vivo temperatures is recognized again in mitochondrial physiology. Incubation times of 1 hour were typical in experiments with the Warburg apparatus, but were reduced to a few or up to 20 min, following Chance and Williams, due to rapid oxygen depletion in closed, aqueous phase oxygraphs with high sample concentrations. Today, incubation times of 1 hour are typical again in high-resolution respirometry, with low sample concentrations and the option of reoxygenations.
Oesper P (1964) The history of the Warburg apparatus: Some reminiscences on its use. J Chem Educ 41: 294.
Koppenol WH, Bounds PL, Dang CV (2011) Otto Warburg’s contributions to current concepts of cancer metabolism. Nature Reviews Cancer 11: 325-337.
Gnaiger E, Kemp RB (1990) Anaerobic metabolism in aerobic mammalian cells: information from the ratio of calorimetric heat flux and respirometric oxygen flux. Biochim Biophys Acta 1016: 328-332. – “At high fructose concentrations, respiration is inhibited while glycolytic end products accumulate, a phenomenon known as the Crabtree effect. It is commonly believed that this effect is restricted to microbial and tumour cells with uniquely high glycolytic capacities (Sussman et al, 1980). However, inhibition of respiration and increase of lactate production are observed under aerobic conditions in beating rat heart cell cultures (Frelin et al, 1974) and in isolated rat lung cells (Ayuso-Parrilla et al, 1978). Thus, the same general mechanisms responsible for the integration of respiration and glycolysis in tumour cells (Sussman et al, 1980) appear to be operating to some extent in several isolated mammalian cells.”
Mitochondria are sometimes described as “cellular power plants” because they generate most of the cell’s supply of adenosine triphosphate (ATP), used as a source of chemical energy.[2] In addition to supplying cellular energy, mitochondria are involved in other tasks such as signaling, cellular differentiation, cell death, as well as the control of the cell cycle and cell growth.[3] The organelle is composed of compartments that carry out specialized functions. These compartments or regions include the outer membrane, the intermembrane space, the inner membrane, and the cristae and matrix. Mitochondrial proteins vary depending on the tissue and the species. In humans, 615 distinct types of proteins have been identified from cardiac mitochondria,[9Leonor Michaelis discovered that Janus green can be used as a supravital stain for mitochondria in 1900. Benjamin F. Kingsbury, in 1912, first related them with cell respiration, but almost exclusively based on morphological observations.[13] In 1913 particles from extracts of guinea-pig liver were linked to respiration by Otto Heinrich Warburg, which he called “grana”. Warburg and Heinrich Otto Wieland, who had also postulated a similar particle mechanism, disagreed on the chemical nature of the respiration. It was not until 1925 when David Keilin discovered cytochromes that the respiratory chain was described.[13]
The Clark Oxygen Sensor
Dr. Leland Clark – inventor of the “Clark Oxygen Sensor” (1954); the Clark type polarographic oxygen sensor remains the gold standard for measuring dissolved oxygen in biomedical, environmental and industrial applications . ‘The convenience and simplicity of the polarographic ‘oxygen electrode’ technique for measuring rapid changes in the rate of oxygen utilization by cellular and subcellular systems is now leading to its more general application in many laboratories. The types and design of oxygen electrodes vary, depending on the investigator’s ingenuity and specific requirements of the system under investigation.’Estabrook R (1967) Mitochondrial respiratory control and the polarographic measurement of ADP:O ratios. Methods Enzymol. 10: 41-47. “one approach that is underutilized in whole-cell bioenergetics, and that is accessible as long as cells can be obtained in suspension, is the oxygen electrode, which can obtain more precise information on the bioenergetic status of the in situ mitochondria than more ‘high-tech’ approaches such as fluorescent monitoring ofΔψm.” Nicholls DG, Ferguson S (2002) Bioenergetics 3. Academic Press, London.
Great Figures in Cancer
Dr. Elizabeth Blackburn,
j_michael_bishop onogene
Harold Varmus
Potts and Habener (PTH mRNA, Harvard MIT) JCI
JCI Fuller Albright and hPTH AA sequence
Dr. E. Donnall Thomas Bone Marrow Transplants
Dr Haraldzur Hausen EBV HPV
Dr. Craig Mello
Lee Hartwell – Hutchinson Cancer Res Center
Judah Folkman, MD
Gertrude B. Elien (1918-1999)
The Nobel Prize in Physiology or Medicine 1922
Archibald V. Hill, Otto Meyerhof
AV Hill –
“the production of heat in the muscle” Hill started his research work in 1909. It was due to J.N. Langley, Head of the Department of Physiology at that time that Hill took up the study on the nature of muscular contraction. Langley drew his attention to the important (later to become classic) work carried out by Fletcher and Hopkins on the problem of lactic acid in muscle, particularly in relation to the effect of oxygen upon its removal in recovery. In 1919 he took up again his study of the physiology of muscle, and came into close contact with Meyerhof of Kiel who, approaching the problem differently, arrived at results closely analogous to his study. In 1919 Hill’s friend W. Hartree, mathematician and engineer, joined in the myothermic investigations – a cooperation which had rewarding results.
Otto Meyerhof –
otto-fritz-meyerhof
lactic acid production in muscle contraction Under the influence of Otto Warburg, then at Heidelberg, Meyerhof became more and more interested in cell physiology. In 1923 he was offered a Professorship of Biochemistry in the United States, but Germany was unwilling to lose him. In 1929 he was he was placed in charge of the newly founded Kaiser Wilhelm Institute for Medical Research at Heidelberg. From 1938 to 1940 he was Director of Research at the Institut de Biologie physico-chimique at Paris, but in 1940 he moved to the United States, where the post of Research Professor of Physiological Chemistry had been created for him by the University of Pennsylvania and the Rockefeller Foundation. Meyerhof’s own account states that he was occupied chiefly with oxidation mechanisms in cells and with extending methods of gas analysis through the calorimetric measurement of heat production, and especially the respiratory processes of nitrifying bacteria. The physico-chemical analogy between oxygen respiration and alcoholic fermentation caused him to study both these processes in the same subject, namely, yeast extract. By this work he discovered a co-enzyme of respiration, which could be found in all the cells and tissues up till then investigated. At the same time he also found a co-enzyme of alcoholic fermentation. He also discovered the capacity of the SH-group to transfer oxygen; after Hopkins had isolated from cells the SH bodies concerned, Meyerhof showed that the unsaturated fatty acids in the cell are oxidized with the help of the sulfhydryl group. After studying closer the respiration of muscle, Meyerhof investigated the energy changes in muscle. Considerable progress had been achieved by the English scientists Fletcher and Hopkins by their recognition of the fact that lactic acid formation in the muscle is closely connected with the contraction process. These investigations were the first to throw light upon the highly paradoxical fact, already established by the physiologist Hermann, that the muscle can perform a considerable part of its external function in the complete absence of oxygen.
But it was indisputable that in the last resort the energy for muscle activity comes from oxidation, so the connection between activity and combustion must be an indirect one, and observed that in the absence of oxygen in the muscle, lactic acid appears, slowly in the relaxed state and rapidly in the active state, disappearing in the presence of oxygen. Obviously, then, oxygen is involved when muscle is in the relaxed state. http://upload.wikimedia.org/wikipedia/commons/e/e1/Glycolysis.jpg
The Nobel Prize committee had been receiving nominations intermittently for the previous 14 years (for Eijkman, Funk, Goldberger, Grijns, Hopkins and Suzuki but, strangely, not for McCollum in this period). Tthe Committee for the 1929 awards apparently agreed that it was high time to honor the discoverer(s) of vitamins; but who were they? There was a clear case for Grijns, but he had not been re-nominated for that particular year, and it could be said that he was just taking the relatively obvious next steps along the new trail that had been laid down by Eijkman, who was also now an old man in poor health, but there was no doubt that he had taken the first steps in the use of an animal model to investigate the nutritional basis of a clinical disorder affecting millions. Goldberger had been another important contributor, but his recent death put him out of consideration. The clearest evidence for lack of an unknown “something” in a mammalian diet was presented by Gowland Hopkins in 1912. This Cambridge biochemist was already well known for having isolated the amino acid tryptophan from a protein and demonstrated its essential nature. He fed young rats on an experimental diet, half of them receiving a daily milk supplement, and only those receiving milk grew well, Hopkins suggested that this was analogous to human diseases related to diet, as he had suggested already in a lecture published in 1906. Hopkins, the leader of the “dynamic biochemistry” school in Britain and an influential advocate for the importance of vitamins, was awarded the prize jointly with Eijkman. A door was opened. Recognition of work on the fat-soluble vitamins begun by McCollum. The next award related to vitamins was given in 1934 to George Whipple, George Minot and William Murphy “for their discoveries concerning liver therapy in cases of [then incurable pernicious] anemia,” The essential liver factor (cobalamin, or vitamin B12) was isolated in 1948, and Vitamin B12 was absent from plant foods. But William Castle in 1928 had demonstrated that the stomachs of pernicious anemia patients were abnormal in failing to secrete an “intrinsic factor”.
Szent-Györgyi was a Hungarian biochemist who had worked with Otto Warburg and had a special interest in oxidation-reduction mechanisms. He was invited to Cambridge in England in 1927 after detecting an antioxidant compound in the adrenal cortex, and there, he isolated a compound that he named hexuronic acid. Charles Glen King of the University of Pittsburgh reported success In isolating the anti-scorbutic factor in 1932, and added that his crystals had all the properties reported by Szent-Györgyi for hexuronic acid. But his work on oxidation reactions was also important. Fumarate is an intermediate in the citric acid cycle used by cells to produce energy in the form of adenosine triphosphate (ATP) from food. It is formed by the oxidation of succinate by the enzyme succinate dehydrogenase. Fumarate is then converted by the enzyme fumarase to malate. An enzyme adds water to the fumarate molecule to form malate. The malate is created by adding one hydrogen atom to a carbon atom and then adding a hydroxyl group to a carbon next to a terminal carbonyl group.
In the same year, Norman Haworth from the University of Birmingham in England received a Nobel prize from the Chemistry Committee for having advanced carbohydrate chemistry and, specifically, for having worked out the structure of Szent-Györgyi’s crystals, and then been able to synthesize the vitamin. This was a considerable achievement. The Nobel Prize in Chemistry was shared with the Swiss organic chemist Paul Karrer, cited for his work on the structures of riboflavin and vitamins A and E as well as other biologically interesting compounds. This was followed in 1938 by a further Chemistry award to the German biochemist Richard Kuhn, who had also worked on carotenoids and B-vitamins, including riboflavin and pyridoxine. But Karrer was not permitted to leave Germany at that time by the Nazi regime. However, the American work with radioisotopes at Lawrence Livermore Laboratory, UC Berkeley, was already ushering in a new era of biochemistry that would enrich our studies of metabolic pathways. The importance of work involving vitamins was acknowledged in at least ten awards in the 20th century.
1. Carpenter, K.J., Beriberi, White Rice and Vitamin B, University of California Press, Berkeley (2000).
2. Weatherall, M.W. and Kamminga, H., The making of a biochemist: the construction of Frederick Gowland Hopkins’ reputation. Medical History vol.40, pp. 415-436 (1996).
3. Becker, S.L., Will milk make them grow? An episode in the discovery of the vitamins. In Chemistry and Modern Society (J. Parascandela, editor) pp. 61-83, American Chemical Society,
Washington, D.C. (1983).
4. Carpenter, K.J., The History of Scurvy and Vitamin C, Cambridge University Press, New York (1986).
Transport and metabolism of exogenous fumarate and 3-phosphoglycerate in vascular smooth muscle.
The keto (linear) form of exogenous fructose 1,6-bisphosphate, a highly charged glycolytic intermediate, may utilize a dicarboxylate transporter to cross the cell membrane, support glycolysis, and produce ATP anaerobically. We tested the hypothesis that fumarate, a dicarboxylate, and 3-phosphoglycerate (3-PG), an intermediate structurally similar to a dicarboxylate, can support contraction in vascular smooth muscle during hypoxia. 3-PG improved maintenance of force (p < 0.05) during the 30-80 min period of hypoxia. Fumarate decreased peak isometric force development by 9.5% (p = 0.008) but modestly improved maintenance of force (p < 0.05) throughout the first 80 min of hypoxia. 13C-NMR on tissue extracts and superfusates revealed 1,2,3,4-(13)C-fumarate (5 mM) metabolism to 1,2,3,4-(13)C-malate under oxygenated and hypoxic conditions suggesting uptake and metabolism of fumarate. In conclusion, exogenous fumarate and 3-PG readily enter vascular smooth muscle cells, presumably by a dicarboxylate transporter, and support energetically important pathways.
Comparison of endogenous and exogenous sources of ATP in fueling Ca2+ uptake in smooth muscle plasma membrane vesicles.
A smooth muscle plasma membrane vesicular fraction (PMV) purified for the (Ca2+/Mg2+)-ATPase has endogenous glycolytic enzyme activity. In the presence of glycolytic substrate (fructose 1,6-diphosphate) and cofactors, PMV produced ATP and lactate and supported calcium uptake. The endogenous glycolytic cascade supports calcium uptake independent of bath [ATP]. A 10-fold dilution of PMV, with the resultant 10-fold dilution of glycolytically produced bath [ATP] did not change glycolytically fueled calcium uptake (nanomoles per milligram protein). Furthermore, the calcium uptake fueled by the endogenous glycolytic cascade persisted in the presence of a hexokinase-based ATP trap which eliminated calcium uptake fueled by exogenously added ATP. Thus, it appears that the endogenous glycolytic cascade fuels calcium uptake in PMV via a membrane-associated pool of ATP and not via an exchange of ATP with the bulk solution. To determine whether ATP produced endogenously was utilized preferentially by the calcium pump, the ATP production rates of the endogenous creatine kinase and pyruvate kinase were matched to that of glycolysis and the calcium uptake fueled by the endogenous sources was compared with that fueled by exogenous ATP added at the same rate. The rate of calcium uptake fueled by endogenous sources of ATP was approximately twice that supported by exogenously added ATP, indicating that the calcium pump preferentially utilizes ATP produced by membrane-bound enzymes.
Evidence for succinate production by reduction of fumarate during hypoxia in isolated adult rat heart cells.
Archives of Biochemistry and Biophysics (Impact Factor: 3.37). 01/1988; 259(2):527-35. http://dx.doi.org:/10.1016/0003-9861(87)90519-4 It has been demonstrated that perfusion of myocardium with glutamic acid or tricarboxylic acid cycle intermediates during hypoxia or ischemia, improves cardiac function, increases ATP levels, and stimulates succinate production. In this study isolated adult rat heart cells were used to investigate the mechanism of anaerobic succinate formation and examine beneficial effects attributed to ATP generated by this pathway. Myocytes incubated for 60 min under hypoxic conditions showed a slight loss of ATP from an initial value of 21 +/- 1 nmol/mg protein, a decline of CP from 42 to 17 nmol/mg protein and a fourfold increase in lactic acid production to 1.8 +/- 0.2 mumol/mg protein/h. These metabolite contents were not altered by the addition of malate and 2-oxoglutarate to the incubation medium nor were differences in cell viability observed; however, succinate release was substantially accelerated to 241 +/- 53 nmol/mg protein. Incubation of cells with [U-14C]malate or [2-U-14C]oxoglutarate indicates that succinate is formed directly from malate but not from 2-oxoglutarate. Moreover, anaerobic succinate formation was rotenone sensitive.
We conclude that malate reduction to succinate occurs via the reverse action of succinate dehydrogenase in a coupled reaction where NADH is oxidized (and FAD reduced) and ADP is phosphorylated.Furthermore, by transaminating with aspartate to produce oxaloacetate, 2-oxoglutarate stimulates cytosolic malic dehydrogenase activity, whereby malate is formed and NADH is oxidized.
In the form of malate, reducing equivalents and substrate are transported into the mitochondria where they are utilized for succinate synthesis.
1953 Hans Adolf Krebs –
” discovery of the citric acid cycle” and In the course of the 1920’s and 1930’s great progress was made in the study of the intermediary reactions by which sugar is anaerobically fermented to lactic acid or to ethanol and carbon dioxide. The success was mainly due to the joint efforts of the schools of Meyerhof, Embden, Parnas, von Euler, Warburg and the Coris, who built on the pioneer work of Harden and of Neuberg. This work brought to light the main intermediary steps of anaerobic fermentations.
In contrast, very little was known in the earlier 1930’s about the intermediary stages through which sugar is oxidized in living cells. When, in 1930, I left the laboratory of Otto Warburg (under whose guidance I had worked since 1926 and from whom I have learnt more than from any other single teacher), I was confronted with the question of selecting a major field of study and I felt greatly attracted by the problem of the intermediary pathway of oxidations.
These reactions represent the main energy source in higher organisms, and in view of the importance of energy production to living organisms (whose activities all depend on a continuous supply of energy) the problem seemed well worthwhile studying. http://www.johnkyrk.com/krebs.html
Interactive Krebs cycle
There are different points where metabolites enter the Krebs’ cycle. Most of the products of protein, carbohydrates and fat metabolism are reduced to the molecule acetyl coenzyme A that enters the Krebs’ cycle. Glucose, the primary fuel in the body, is first metabolized into pyruvic acid and then into acetyl coenzyme A. The breakdown of the glucose molecule forms two molecules of ATP for energy in the Embden Meyerhof pathway process of glycolysis.
On the other hand, amino acids and some chained fatty acids can be metabolized into Krebs intermediates and enter the cycle at several points. When oxygen is unavailable or the Krebs’ cycle is inhibited, the body shifts its energy production from the Krebs’ cycle to the Embden Meyerhof pathway of glycolysis, a very inefficient way of making energy.
Fritz Albert Lipmann –
“discovery of co-enzyme A and its importance for intermediary metabolism”.
In my development, the recognition of facts and the rationalization of these facts into a unified picture, have interplayed continuously. After my apprenticeship with Otto Meyerhof, a first interest on my own became the phenomenon we call the Pasteur effect, this peculiar depression of the wasteful fermentation in the respiring cell. By looking for a chemical explanation of this economy measure on the cellular level, I was prompted into a study of the mechanism of pyruvic acid oxidation, since it is at the pyruvic stage where respiration branches off from fermentation.
For this study I chose as a promising system a relatively simple looking pyruvic acid oxidation enzyme in a certain strain of Lactobacillus delbrueckii1. In 1939, experiments using minced muscle cells demonstrated that one oxygen atom can form two adenosine triphosphate molecules, and, in 1941, the concept of phosphate bonds being a form of energy in cellular metabolism was developed by Fritz Albert Lipmann.
In the following years, the mechanism behind cellular respiration was further elaborated, although its link to the mitochondria was not known.[13]The introduction of tissue fractionation by Albert Claude allowed mitochondria to be isolated from other cell fractions and biochemical analysis to be conducted on them alone. In 1946, he concluded that cytochrome oxidase and other enzymes responsible for the respiratory chain were isolated to the mitchondria. Over time, the fractionation method was tweaked, improving the quality of the mitochondria isolated, and other elements of cell respiration were determined to occur in the mitochondria.[13]
The most important event during this whole period, I now feel, was the accidental observation that in the L. delbrueckii system, pyruvic acid oxidation was completely dependent on the presence of inorganic phosphate. This observation was made in the course of attempts to replace oxygen by methylene blue. To measure the methylene blue reduction manometrically, I had to switch to a bicarbonate buffer instead of the otherwise routinely used phosphate. In bicarbonate, pyruvate oxidation was very slow, but the addition of a little phosphate caused a remarkable increase in rate. The phosphate effect was removed by washing with a phosphate free acetate buffer. Then it appeared that the reaction was really fully dependent on phosphate.
A coupling of this pyruvate oxidation with adenylic acid phosphorylation was attempted. Addition of adenylic acid to the pyruvic oxidation system brought out a net disappearance of inorganic phosphate, accounted for as adenosine triphosphate. The acetic acid subunit of acetyl CoA is combined with oxaloacetate to form a molecule of citrate. Acetyl coenzyme A acts only as a transporter of acetic acid from one enzyme to another. After Step 1, the coenzyme is released by hydrolysis to combine with another acetic acid molecule and begin the Krebs’ Cycle again.
Hugo Theorell –
“the nature and effects of oxidation enzymes”
From 1933 until 1935 Theorell held a Rockefeller Fellowship and worked with Otto Warburg at Berlin-Dahlem, and here he became interested in oxidation enzymes. At Berlin-Dahlem he produced, for the first time, the oxidation enzyme called «the yellow ferment» and he succeeded in splitting it reversibly into a coenzyme part, which was found to be flavin mononucleotide, and a colourless protein part. On return to Sweden, he was appointed Head of the newly established Biochemical Department of the Nobel Medical Institute, which was opened in 1937.
Succinate is oxidized by a molecule of FAD (Flavin Adenine Dinucleotide). The FAD removes two hydrogen atoms from the succinate and forms a double bond between the two carbon atoms to create fumarate.
They followed the path that became clear from intense collaborative work in California by George Beadle, by Avery and McCarthy, Max Delbruck, TH Morgan, Max Delbruck and by Chargaff that indicated that genetics would be important.
1965
François Jacob, André Lwoff and Jacques Monod –
” genetic control of enzyme and virus synthesis”.
In 1958 the remarkable analogy revealed by genetic analysis of lysogeny and that of the induced biosynthesis of ß-galactosidase led François Jacob, with Jacques Monod, to study the mechanisms responsible for the transfer of genetic information as well as the regulatory pathways which, in the bacterial cell, adjust the activity and synthesis of macromolecules. Following this analysis, Jacob and Monod proposed a series of new concepts, those of messenger RNA, regulator genes, operons and allosteric proteins.
Francois Jacob
Having determined the constants of growth in the presence of different carbohydrates, it occurred to me that it would be interesting to determine the same constants in paired mixtures of carbohydrates. From the first experiment on, I noticed that, whereas the growth was kinetically normal in the presence of certain mixtures (that is, it exhibited a single exponential phase), two complete growth cycles could be observed in other carbohydrate mixtures, these cycles consisting of two exponential phases separated by a-complete cessation of growth.
Lwoff, after considering this strange result for a moment, said to me, “That could have something to do with enzyme adaptation.”
“Enzyme adaptation? Never heard of it!” I said.
Lwoff’s only reply was to give me a copy of the then recent work of Marjorie Stephenson, in which a chapter summarized with great insight the still few studies concerning this phenomenon, which had been discovered by Duclaux at the end of the last century. Studied by Dienert and by Went as early as 1901 and then by Euler and Josephson, it was more or less rediscovered by Karström, who should be credited with giving it a name and attracting attention to its existence.
Lwoff’s intuition was correct. The phenomenon of “diauxy” that I had discovered was indeed closely related to enzyme adaptation, as my experiments, included in the second part of my doctoral dissertation, soon convinced me. It was actually a case of the “glucose effect” discovered by Dienert as early as 1900. That agents that uncouple oxidative phosphorylation, such as 2,4-dinitrophenol, completely inhibit adaptation to lactose or other carbohydrates suggested that “adaptation” implied an expenditure of chemical potential and therefore probably involved the true synthesis of an enzyme.
With Alice Audureau, I sought to discover the still quite obscure relations between this phenomenon and the one Massini, Lewis, and others had discovered: the appearance and selection of “spontaneous” mutants. We showed that an apparently spontaneous mutation was allowing these originally “lactose-negative” bacteria to become “lactose-positive”. However, we proved that the original strain (Lac-) and the mutant strain (Lac+) did not differ from each other by the presence of a specific enzyme system, but rather by the ability to produce this system in the presence of lactose. This mutation involved the selective control of an enzyme by a gene, and the conditions necessary for its expression seemed directly linked to the chemical activity of the system.
1974
Albert Claude, Christian de Duve and George E. Palade –
” the structural and functional organization of the cell”.
I returned to Louvain in March 1947 after a period of working with Theorell in Sweden, the Cori’s, and E Southerland in St. Louis, fortunate in the choice of my mentors, all sticklers for technical excellence and intellectual rigor, those prerequisites of good scientific work. Insulin, together with glucagon which I had helped rediscover, was still my main focus of interest, and our first investigations were accordingly directed on certain enzymatic aspects of carbohydrate metabolism in liver, which were expected to throw light on the broader problem of insulin action. But I became distracted by an accidental finding related to acid phosphatase, drawing most of my collaborators along with me. The studies led to the discovery of the lysosome, and later of the peroxisome.
In 1962, I was appointed a professor at the Rockefeller Institute in New York, now the Rockefeller University, the institution where Albert Claude had made his pioneering studies between 1929 and 1949, and where George Palade had been working since 1946. In New York, I was able to develop a second flourishing group, which follows the same general lines of research as the Belgian group, but with a program of its own.
1968
Robert W. Holley, Har Gobind Khorana and Marshall W. Nirenberg –
“interpretation of the genetic code and its function in protein synthesis”.
1969
Max Delbrück, Alfred D. Hershey and Salvador E. Luria –
” the replication mechanism and the genetic structure of viruses”.
1975 David Baltimore, Renato Dulbecco and Howard Martin Temin –
” the interaction between tumor viruses and the genetic material of the cell”.
1976
Baruch S. Blumberg and D. Carleton Gajdusek –
” new mechanisms for the origin and dissemination of infectious diseases” The editors of the Nobelprize.org website of the Nobel Foundation have asked me to provide a supplement to the autobiography that I wrote in 1976 and to recount the events that happened after the award. Much of what I will have to say relates to the scientific developments since the last essay. These are described in greater detail in a scientific memoir first published in 2002 (Blumberg, B. S., Hepatitis B. The Hunt for a Killer Virus, Princeton University Press, 2002, 2004).
1980
Baruj Benacerraf, Jean Dausset and George D. Snell
” genetically determined structures on the cell surface that regulate immunological reactions”.
1992
Edmond H. Fischer and Edwin G. Krebs
“for their discoveries concerning reversible protein phosphorylation as a biological regulatory mechanism”
1994
Alfred G. Gilman and Martin Rodbell –
“G-proteins and the role of these proteins in signal transduction in cells”
2011
Bruce A. Beutler and Jules A. Hoffmann –
” the activation of innate immunity“and the other half to Ralph M. Steinman – “the dendritic cell and its role in adaptive immunity”.
Renato L. Baserga, M.D.
Kimmel Cancer Center and Keck School of Medicine
Dr. Baserga’s research focuses on the multiple roles of the type 1 insulin-like growth factor receptor (IGF-IR) in the proliferation of mammalian cells. The IGF-IR activated by its ligands is mitogenic, is required for the establishment and the maintenance of the transformed phenotype, and protects tumor cells from apoptosis. It, therefore, serves as an excellent target for therapeutic interventions aimed at inhibiting abnormal growth. In basic investigations, this group is presently studying the effects that the number of IGF-IRs and specific mutations in the receptor itself have on its ability to protect cells from apoptosis.
This investigation is strictly correlated with IGF-IR signaling, and part of this work tries to elucidate the pathways originating from the IGF-IR that are important for its functional effects. Baserga’s group has recently discovered a new signaling pathway used by the IGF-IR to protect cells from apoptosis, a unique pathway that is not used by other growth factor receptors. This pathway depends on the integrity of serines 1280-1283 in the C-terminus of the receptor, which bind 14.3.3 and cause the mitochondrial translocation of Raf-1.
Another recent discovery of this group has been the identification of a mechanism by which the IGF-IR can actually induce differentiation in certain types of cells. When cells have IRS-1 (a major substrate of the IGF-IR), the IGF-IR sends a proliferative signal; in the absence of IRS-1, the receptor induces cell differentiation. The extinction of IRS-1 expression is usually achieved by DNA methylation.
Janardan Reddy, MD
Northwestern University
The central effort of our research has been on a detailed analysis at the cellular and molecular levels of the pleiotropic responses in liver induced by structurally diverse classes of chemicals that include fibrate class of hypolipidemic drugs, and phthalate ester plasticizers, which we designated hepatic peroxisome proliferators. Our work has been central to the establishment of several principles, namely that hepatic peroxisome proliferation is associated with increases in fatty acid oxidation systems in liver, and that peroxisome proliferators, as a class, are novel nongenotoxic hepatocarcinogens.
We introduced the concept that sustained generation of reactive oxygen species leads to oxidative stress and serves as the basis for peroxisome proliferator-induced liver cancer development. Furthermore, based on the tissue/cell specificity of pleiotropic responses and the coordinated transcriptional regulation of fatty acid oxidation system genes, we postulated that peroxisome proliferators exert their action by a receptor-mediated mechanism. This receptor concept laid the foundation for the discovery of
a three member peroxisome proliferator-activated receptor (PPARalpha-, ß-, and gamma) subfamily of nuclear receptors.
PPARalpha is responsible for peroxisome proliferator-induced pleiotropic responses, including
hepatocarcinogenesis and energy combustion as it serves as a fatty acid sensor and regulates fatty acid oxidation.
Our current work focuses on the molecular mechanisms responsible for PPAR action and generation of fatty acid oxidation deficient mouse knockout models. Transcription of specific genes by nuclear receptors is a complex process involving the participation of multiprotein complexes composed of transcription coactivators.
Jose Delgado de Salles Roselino, Ph.D.
Leloir Institute, Brazil
Warburg effect, in reality “Pasteur-effect” was the first example of metabolic regulation described. A decrease in the carbon flux originated at the sugar molecule towards the end metabolic products, ethanol and carbon dioxide that was observed when yeast cells were transferred from anaerobic environmental condition to an aerobic one. In Pasteur´s works, sugar metabolism was measured mainly by the decrease of sugar concentration in the yeast growth media observed after a measured period of time. The decrease of the sugar concentration in the media occurs at great speed in yeast grown in anaerobiosis condition and its speed was greatly reduced by the transfer of the yeast culture to an aerobic condition. This finding was very important for the wine industry of France in Pasteur time, since most of the undesirable outcomes in the industrial use of yeast were perceived when yeasts cells took very long time to create a rather selective anaerobic condition. This selective culture media was led by the carbon dioxide higher levels produced by fast growing yeast cells and by a great alcohol content in the yeast culture media. This finding was required to understand Lavoisier’s results indicating that chemical and biological oxidation of sugars produced the same calorimetric results. This observation requires a control mechanism (metabolic regulation) to avoid burning living cells by fast heat released by the sugar biological oxidative processes (metabolism). In addition, Lavoisier´s results were the first indications that both processes happened inside similar thermodynamics limits.
In much resumed form, these observations indicates the major reasons that led Warburg to test failure in control mechanisms in cancer cells in comparison with the ones observed in normal cells. Biology inside classical thermo dynamics poses some challenges to scientists. For instance, all classical thermodynamics must be measured in reversible thermodynamic conditions. In an isolated system, increase in P (pressure) leads to decrease in V (volume) all this in a condition in which infinitesimal changes in one affects in the same way the other, a continuum response. Not even a quantic amount of energy will stand beyond those parameters. In a reversible system, a decrease in V, under same condition, will led to an increase in P.
In biochemistry, reversible usually indicates a reaction that easily goes from A to B or B to A. This observation confirms the important contribution of E Schrodinger in his What´s Life: “This little book arose from a course of public lectures, delivered by a theoretical physicist to an audience of about four hundred which did not substantially dwindle, though warned at the outset that the subject-matter was a difficult one and that the lectures could not be termed popular, even though the physicist’s most dreaded weapon, mathematical deduction, would hardly be utilized. The reason for this was not that the subject was simple enough to be explained without mathematics, but rather that it was much too involved to be fully accessible to mathematics.”
Hans Krebs describes the cyclic nature of the citrate metabolism. Two major research lines search to understand the mechanism of energy transfer that explains how ADP is converted into ATP. One followed the organic chemistry line of reasoning and therefore, searched how the breakdown of carbon-carbon link could have its energy transferred to ATP synthesis. A major leader of this research line was B. Chance who tried to account for two carbon atoms of acetyl released as carbon dioxide in the series of Krebs cycle reactions. The intermediary could store in a phosphorylated amino acid the energy of carbon-carbon bond breakdown. This activated amino acid could transfer its phosphate group to ADP producing ATP. Alternatively, under the possible influence of the excellent results of Hodgkin and Huxley a second line of research appears.
The work of Hodgkin & Huxley indicated the storage of electrical potential energy in transmembrane ionic asymmetries and presented the explanation for the change from resting to action potential in excitable cells. This second line of research, under the leadership of P Mitchell postulated a mechanism for the transfer of oxide/reductive power of organic molecules oxidation through electron transfer as the key for energetic transfer mechanism required for ATP synthesis. Paul Boyer could present how the energy was transduced by a molecular machine that changes in conformation in a series of 3 steps while rotating in one direction in order to produce ATP and in opposite direction in order to produce ADP plus Pi from ATP (reversibility). Nonetheless, a victorious Peter Mitchell obtained the correct result in the conceptual dispute, over the B. Chance point of view, after he used E. Coli mutants to show H gradients in membrane and its use as energy source.
However, this should not detract from the important work of Chance. B. Chance got the simple and rapid polarographic assay method of oxidative phosphorylation and the idea of control of energy metabolism that bring us back to Pasteur. This second result seems to have been neglected in searching for a single molecular mechanism required for the understanding of the buildup of chemical reserve in our body. In respiring mitochondria the rate of electron transport, and thus the rate of ATP production, is determined primarily by the relative concentrations of ADP, ATP and phosphate in the external media (cytosol) and not by the concentration of respiratory substrate as pyruvate. Therefore, when the yield of ATP is high as is in aerobiosis and the cellular use of ATP is not changed, the oxidation of pyruvate and therefore of glycolysis is quickly (without change in gene expression), throttled down to the resting state. The dependence of respiratory rate on ADP concentration is also seen in intact cells. A muscle at rest and using no ATP has very low respiratory rate.
I have had an ongoing discussion with Jose Eduardo de Salles Roselino, inBrazil. He has made important points that need to be noted.
The constancy of composition which animals maintain in the environment surrounding their cells is one of the dominant features of their physiology. Although this phenomenon, homeostasis, has held the interest of biologists over a long period of time, the elucidation of the molecular basis for complex processes such as temperature control and the maintenance of various substances at constant levels in the blood has not yet been achieved. By comparison, metabolic regulation in microorganisms is much better understood, in part because the microbial physiologist has focused his attention on enzyme-catalyzed reactions and their control, as these are among the few activities of microorganisms amenable to quantitative study. Furthermore, bacteria are characterized by their ability to make rapid and efficient adjustments to extensive variations in most parameters of their environment; therefore, they exhibit a surprising lack of rigid requirements for their environment, and appears to influence it only as an incidental result of their metabolism. Animal cells on the other hand have only a limited capacity for adjustment and therefore require a constant milieu. Maintenance of such constancy appears to be a major goal in their physiology (Regulation of Biosynthetic Pathways H.S. Moyed and H EUmbarger Phys Rev,42 444 (1962)).
A living cell consists in a large part of a concentrated mixture of hundreds of different enzymes, each a highly effective catalyst for one or more chemical reactions involving other components of the cell. The paradox of intense and highly diverse chemical activity on the one hand and strongly poised chemical stability (biological homeostasis) on the other is one of the most challenging problems of biology (Biological feedback Control at the molecular Level D.E. Atkinson Science vol. 150: 851, 1965). Almost nothing is known concerning the actual molecular basis for modulation of an enzyme`s kinetic behavior by interaction with a small molecule. (Biological feedback Control at the molecular Level D.E. Atkinson Science vol. 150: 851, 1965). In the same article, since the core of Atkinson´s thinking seems to be strongly linked with Adenylates as regulatory effectors, the previous phrases seems to indicate a first step towards the conversion of homeostasis to an intracellular phenomenon and therefore, one that contrary to Umbarger´s consideration could be also studied in microorganisms.
Most biochemical studies using bacteria, were made before the end of the third upper part of log growth phase. Therefore, they could be considered as time-independent as S Luria presented biochemistry in Life an Unfinished Experiment. The sole ingredient on the missing side of the events that led us into the molecular biology construction was to consider that proteins, a macromolecule, would never be affected by small molecules translational kinetic energy. This, despite the fact that in a catalytic environment and its biological implications S Grisolia incorporated A K Balls observation indicating that the word proteins could be related to Proteus an old sea god that changed its form whenever he was subjected to inquiry (Phys Rev v 4,657 (1964).
In D.E. Atkinson´s work (Science vol 150 p 851, 1965), changes in protein synthesis acting together with factors that interfere with enzyme activity will lead to “fine-tuned” regulation better than enzymatic activity regulation alone. Comparison of glycemic regulation in granivorous and carnivorous birds indicate that when no important nutritional source of glucose is available, glycemic levels can be kept constant in fasted and fed birds. The same was found in rats and cats fed on high protein diets. Gluconeogenesis is controlled by pyruvate kinase inhibition. Therefore, the fact that it can discriminate between fasting alone and fasting plus exercise (carbachol) requirement of gluconeogenic activity (correspondent level of pyruvate kinase inhibition) the control of enzyme activity can be made fast and efficient without need for changes in genetic expression (20 minute after stimulus) ( Migliorini,R.H. et al Am J. Physiol.257 (Endocrinol. Met. 20): E486, 1989). Regrettably, this was not discussed in the quoted work. So, when the control is not affected by the absorption of nutritional glucose it can be very fast, less energy intensive and very sensitive mechanism of control despite its action being made in the extracellular medium (homeostasis).
Biochemical role: AA serves a basic biochemical role of accelerating hydroxylation in several biochemical reactions. It provides electrons to metal ions, the reduced forms of which are required for the full enzymatic activity of some enzymes. Most emphasized role of AA is as a cofactor for the enzyme required for the biosynthesis of collagen.
Molecular structure and the oxidized form of AA, dihydroascorbic acid, bear similarity to that of glucose.
Biological role: AA is an essential vitamin for humans and its deficiency leads to disease called Scurvy characterized by initial symptoms of malaise and lethargy, followed by formation of spots on the skin, spongy gums, and bleeding from the mucous membranes. As scurvy advances, there can be open, suppurating wounds, loss of teeth, jaundice, fever, neuropathy and death. AA is water soluble and found in high concentrations in several tissues including eye lens, WBCs, adrenal glad and pituitary gland. Some of the roles of ascorbate include:
Possibly an ergogenic aid (Ergogenic aids are substances, devices, or practices that enhance an individual’s energy use, production, or recovery.)
Vitamin C and Cancer
As early as in 1949, vitamin C was implicated in cancer therapy. Since then, several research articles have been published exploring the role of ascorbate in cancer therapy. Among the plethora of literature discussing the relationship between vitamin C and cancer, one of the very significant and comprehensive reviews was published in 1979 in Cancer Research (2).
Mechanisms of action of AA (1) with respect to cancer have been divided and subdivided into the following:
Ascorbate acts as a cancer preventive agent by virtue of its strong antioxidant activities. Being one of the strongest reductants and radical scavenger, it absorbs unstable oxygen, nitrogen, and sulphur-centered radicals. AA can prevent biomembranes from peroxidative damage from peroxyl radicals. Ascorbate can trap peroxyl radicals and lead to their peroxidation in the aqueous phase before they reach the lipid rich biomembranes and cause damage. Ascorbate has been speculated to have a biomembrane protective action by its synergistic antioxidant activity with vitamin E (tocopherol). Vitamin E is lipid-soluble and tocopheroxyl radical is generated in the cell membranes as a result of its antioxidant activity. Ascorbate reacts with the tocopheroxyl radical and regenerates tocopherol transferring the oxidative challenge to the aqueous phase. At this point, the less active ascorbate radical might be reduced to AA by an NADP-dependent system. The probably mechanism might explain the reduction of nitrates via ascorbate to prevent the formation of carcinogenic nitrosamines.
Anticancer mechanisms
1. Primary anticancer mechanisms
i. Oxidative, oxidant and pro-oxidant properties: Ascorbate has been reported to be cytotoxic at high concentrations, which has been demonstrated in a number of malignant cell lines. Transcription factor NFkB is potentially activated via ascorbate and its radicals leading to the inhibition of cell growth. Also, ascorbate inhibits certain prostaglandins leading to decrease in cell proliferation.
ii. Hydrogen peroxide: On oxidation with oxygen, ascorbate produces a hydrogen peroxide, a reactive oxygen species. Hydrogen peroxide can generate several other reactive species and can have several damaging effects on cells including decrease in cell viability by damaging cell membranes of malignant cells. The amount of these reactive species produced via oxidation is limited in healthy cells unlike that in malignant cells where they exist in large amounts. The amount of hydrogen peroxide generated has been correlated to the amount of ascorbate in the cells. The reactive species can lead to multiple negative effects on cells including DNA strand breaks, lipid peroxidation leading to membrane function disruption, cellular ATP depletion.
Authors state that “the failure to maintain high ATP production may be a consequence of oxidative inactivation of key enzymes especially those related to the Krebs cycle and the electron transport chain.” This might result in alteration of transmembrane potential and distortion of mitochondrial function, suggestive of the important role of mitochondria in the process of carcinogenesis. In this paper, vitamin C has been correlated with cancer with the involvement of altered mitochondrial function. In addition, ascorbate has been detected in mitochondria where it is also regenerated. Different aspects of mitochondrial involvement in cancer have been discussed in several posts published earlier (3-8).
iii. Other oxidation products of AA: Other oxidation products of AA include 2,3-diketoglutonic acid, and 5-methyl 1-3, 4-dehydrotetrone and other degradation products, have demonstrated antitumor activity. Additionally, some degradation and oxidation products of AA, gamma-cronolactone and 3-hydroxyl-2-pyrone, have been found to inhibit tumor growth. The mechanism of their antitumor actions is complex and might involve multitude of steps, including generation of reactive oxygen species, lipid peroxidation, inducing structural changes in important cellular proteins, inhibition of mitosis and so on.
iv. Intracellular transport of ascorbate and its tumor specificity: Oxidized ascorbate, dihydroascorbic acid, is transported intracellularly where it is reduced back to ascorbate. Owing to its structural similarity with glucose, dihydroascorbic transport is facilitated via glucose transporters (GLUTs). Ascrobate in its reduced form is transported through a sodium-dependent cotransporter in some cells. Tumor cells require large amounts of glucose, which leads to an increase in the number of GLUTs, hence, resulting in an increase in ascorbate concentration within cancer cells. Because of this selective increased uptake of ascorbate and its cytotoxic effects in cancer cells (generation of hydrogen peroxide, DNA damage, other cytotoxic effects), AA has become a selective, nontoxic chemotherapeutic agent. The difference in the levels of catalase enzyme has been found to lead to intracellular tumor selectivity in cancer cells.
Ascorbate induced cytotoxicity in cancer cells involves its final electron acceptor, oxygen, which interferes with the anaerobic respiration within malignant cells. This gives an important clue for the involvement of mitochondria in malignant cells.
v. Intravenous AA: High concentrations of AA in plasma (>200mg/dL) have been found to be cytotoxic to cancer cells. Clinically high plasma concentrations of AA can be achieved by its intravenous administration. It was observed that 60g infusion of AA given to cancer patients for 60 minutes followed by 20g given over the next 60 minutes resulted in a 240 minutes high plasma AA concentration of >400mg/dL, that is known to be cytotoxic.
Lipoic acid when administered with AA, is able to reduce the high-dose requirement of AA for its cytotoxic activity reducing it from 700mg/dL to 120mg/dL. Lipoic acid can recycle vitamin C, mediate the reduction of dihydroascorbic acid and improves mitochondrial function. Thus, energy intermediates such as coenzyme Q, vitamin K3, B-complex vitamins, alpha-ketoglutarate aspartate, magnesium might aid in cancer therapy by intercting with ascorbate, directly or indirectly, thereby stimuating/interacting/correcting aerobic mitochondrial respiration.
Hence, the pro-oxidant activity of vitamin C is being referred to as the primary mechanism of anticancer action.
2. Secondary anticancer mechanisms
i. AA and intracellular matrix: Collagen is an important constituent of the matrix and its concentration determines the strength of the tissue along with its resistance to the infiltration of malignant cancer cells. In Scurvy, a disease resulting from a chronic deficiency of vitamin C, there is generalized tissue disintegration, dissolution of intercellular ground substance and the disruption of collagen bundles. This disintegration leads to ulceration; bacterial colonization and general undifferentiated cellular proliferation with specialized cells reverting back to their primitive form, very much like cancer. Lack of ascorbate causes a reduction in the hydroxylation of prolyl and lysyl residues into hydroxyproline and hydroxylysine, leading to instability of the collagen triple helix, a common feature in scurvy and also in cancer. Thus, a secondary mechanism of ascorbic acid anticancer mechanism would be to repair these sites, which is emphasized by its role in wound healing, including surgical recovery and other traumatic injuries.
ii. Ascorbate and immunocompetence: Ascorbate plays several roles for the efficient functioning of immune system in ways that are invoved in both humoral and cell-mediated. Ascorbate provides humoral immunocompetence as it is essential for immunoglobulin synthesis. In addition, lymphocytes, seminal cells involved in cell-mediated immunity have been found to contain high concentrations of ascorbate. Other immune system roles include, aid in active phagocytosis and enhancing of interferon production.
Classical vitamin C and Cancer controversy-A possible explanation
Conflicting results were obtained from the studies performed by Pauling (Pauling Institute) and Cameron (Mayo Clinic) with vitamin C and its effect on cancer, the issue was debated a few decades ago. Both the studies, however, used oral doses of ascorbate (10g). Gonzalez et al, authors of the review on which the post is based, analyzed and expressed their views on the controversy. They state that the plasma concentration cannot be replicated when the dose is given orally as opposed to when the dose is given intravenously. According to their research, when AA is administered intravenously, higher plasma levels of ascorbate are achieved that could be retained for longer time periods. Also, the authors advocate the use of substantially higher doses (25-200g) to be given intravenously for selective toxicity towards cancer cells.
Modern vitamin C and Cancer controversy-Chemotherapy and radiation
A recent concern regarding the antioxidants like vitamin C is that they might reduce the effectiveness of chemotherapy and radiation by reducing the potency of free radicals necessary for killing cells. A publication by Agus et al (13) has a major role to play in this misconception. The authors describe how cancer cells acquire and concentrate vitamin C providing malignant cells with metabolic advantage. However, details or explanations regarding the theory are missing. Some studies, on the other hand, explain that high concentrations of AA in cancer cells is cytotoxic and is achieved because of similarity in structure between AA and glucose. Cancer cells uptake AA derivative, dehydroascorbic acid via glucose transporters (GLUTs).
In a case report published in PNAS in 1985 (12), two patients with ovarian cancer stage IIIC were found to respond positively to chemotherapy along with high-dose of antioxidants. Antioxidant, AA was administered intravenously to maintain a high plasma dose of 200 mg/dL. The two patients didn’t show disease recurrence after three years of chemotherapy and vitamin C administration. Vast literature exists on the topic indicating that antioxidants, including ascorbate, provide beneficial effects in several cancers without reducing the efficacy of chemotherapy or radiation during treatment of these cancers. Some data, in fact, suggests increase in effectiveness of chemotherapy when supplemented with antioxidants along with an increase in adverse effects. The topic has been summarized and discussed in a series of articles by Lawson and Brignall (9-11).
REFERENCES
The post is primarily based on the following two review articles:
Mitochondrial Damage and Repair under Oxidative Stress
Curator: Larry H Bernstein, MD, FCAP
Keywords: Mitochondria, mitochondrial dysfunction, electron transport chain, mtDNA, oxidative stress, oxidation-reduction, NO, DNA repair, lipid peroxidation, thiols, ROS, RNS, sulfur,base excision repair, ferredoxin. Summary: The mitochondrion is the energy source for aerobic activity of the cell, but it also has regulatory functions that will be discussed. The mitochondrion has been discussed in other posts at this site. It has origins from organisms that emerged from an anaerobic environment, such as the bogs and marshes, and may be related to the chloroplast. The aerobic cell was an advance in evolutionary development, but despite the energetic advantage of using oxygen, the associated toxicity of oxygen abundance required adaptive changes. Most bacteria that reduce nitrate (producing nitrite, nitrous oxide or nitrogen) are called facultative anaerobes use electron acceptors such as ferric ions, sulfate or carbon dioxide which become reduced to ferrous ions, hydrogen sulfide and methane, respectively, during the oxidation of NADH (reduced nicotinamide adenine dinucleotide is a major electron carrier in the oxidation of fuel molecules).
The underlying problem we are left with is oxidation-reduction reactions that are necessary for catabolic and synthetic reactions, and that cumulatively damage the organism associated with cancer, cardiovascular disease, neurodegerative disease, and inflammatory overload. Aerobic organisms tolerate have evolved mechanisms to repair or remove damaged molecules or to prevent or deactivate the formationof toxic species that lead to oxidative stress and disease. However, the normal balance between production of pro-oxidant species and destruction by the antioxidant defenses is upset in favor of overproduction of the toxic species, which leads to oxidative stress and disease. How this all comes together is the topic of choice.
The transformation of energy is central to mitochondrial function. The system of energetics includes:
the enzymes of the Kreb’s citric acid or TCA cycle,
some of the enzymes involved in fatty acid catabolism (β-oxidation), and
the proteins needed to help regulate these systems,
central to mitochondrial physiology through the production of reducing equivalents. Reducing equivalents are also used for anabolic reactions. Electron Transport Chain
It also houses the protein complexes involved in the electron transport component of oxidative phosphorylation and proteins involved in substrate and ion transport. The chemical energy contained in both fats and amino acids can also be converted into NADH and FADH2 through mitochondrial pathways. The major mechanism for harvesting energy from fats is β-oxidation; the major mechanism for harvesting energy from amino acids and pyruvate is the TCA cycle. Once the chemical energy has been transformed into NADH and FADH2, these compounds are fed into the mitochondrial respiratory chain.
Under physiological conditions, electrons generally enter either through complex I (NADH-mediated, examined in vitro using substrates such as glutamate/malate) or complex II (FADH2-mediated, examined in vitro using succinate).
Electrons are then sequentially passed through a series of electron carriers.
The progressive transfer of electrons (and resultant proton pumping) converts the chemical energy stored in carbohydrates, lipids, and amino acids into potential energy in the form of the proton gradient. The potential energy stored in this gradient is used to phosphorylate ADP forming ATP. Redox-Cycling
In redox cycling the reductant is continuously regenerated, thereby providing substrate for the “auto-oxidation” reaction.
When partially oxidized compounds are enzymatically reduced, the auto-oxidative generation of superoxide and other ROS to start again. Several enzymes
NADPH-cytochrome P450 reductase,
NADPH-cytochrome b5 reductase [EC 1.6.2.2]
NADPH-ubiquinone oxidoreductase [EC 1.6.5.3], and
xanthine oxidase [EC 1.2.3.2]),
can reduce quinones into semiquinones in a single electron process.
The semiquinone can then reduce dioxygen to superoxide during its oxidation to a quinone.
Redox cycling is thought to play a role in carcinogenesis. The naturally occurring estrogen metabolites (the catecholestrogens) have been implicated in hormone-induced cancer, possibly as a result of their redox cycling and production of ROS. It is thought that diethylstilbestrol causes the production of the mutagenic lesion 8-hydroxy-2’deoxyguanosine. It can also cause DNA strand breakage.
Another oxidative reaction that is associated with H2O2 is a significant problem for living organisms as a consequence of the reaction between hydrogen peroxide and oxidizable metals, the Fenton reaction [originally described in the oxidation of an α-hydroxy acid to an α-keto acid in the presence of hydrogen peroxide (or hypochlorite) and low levels of iron salts (Fenton (1876, 1894)). Chemical Reactions and Biological Significance
The hydroxyl free radical is so aggressive that it will react within 5 (or so) molecular diameters from its site of production. The damage caused by it, therefore, is very site specific. Biological defenses have evolved that reduce the chance that the hydroxyl free radical will be produced to repair damage. An antioxidant would have to occur at the site of hydroxyl free radical production and be at sufficient concentration to be effective.
Some endogenous markers have been proposed as a useful measures of total “oxidative stress” e.g., 8-hydroxy-2’deoxyguanosine in urine. The ideal scavenger
must be non-toxic,
have limited or no biological activity,
readily reach the site of hydroxyl free radical production,
react rapidly with the free radical, be specific for this radical, and
neither the scavenger nor its product(s) should undergo further metabolism.
Unlike oxygen, nitrogen does not possess unpaired electrons and is therefore considered diamagnetic. Nitrogen does not possess available d orbitals so it is limited to a valency of 3. In the presence of oxygen, nitrogen can produce Nitric oxide which occurs physiologically with the immune system which, when activated, can produce large quantities of nitric oxide.
Nitric oxide is produced by stepwise oxidation of L-arginine catalyzed by nitric oxide synthase (NOS). Nitric oxide is formed from the guanidino nitrogen of the L-arginine in a reaction that
consumes five electrons and
requires flavin adenine dinucleotide (FAD),
flavin mononucleotide (FMN) tetrahydrobiopterin (BH4), and
iron protoporphyrin IX as cofactors.
The primary product of NOS activity may be the nitroxyl anion that is then converted to nitric oxide by electron acceptors.
NOS cDNAs show homology with the cytochrome P450 reductase family. Based on molecular genetics there appears to be at least three distinct forms of NOS:
A Ca2+/calmodulin-requiring constitutive enzyme (c-NOS; ncNOS or type I)
A calcium-independent inducible enzyme (i-NOS; type II), which is primarily involved in the mediation of the cellular immune response; and
A second Ca2+/calmodulin-requiring constitutive enzyme found in aortic and umbilical endothelia (ec-NOS or type III)
This has been discussed extensively in this series of posts. Recently, a mitochondrial form of the enzyme, which appears to be similar to the endothelial form, has been found in brain and liver tissue. Although the exact role of nitric oxide in the mitochondrion remains elusive, it may play a role in the regulation of cytochrome oxidase. Nitric Oxide
Nitric oxide appears to regulate its own production through a negative feedback loop. The binding of nitric oxide to the heme prosthetic group of NOS inhibits this enzyme, and c-NOS and ec-NOS are much more sensitive to this regulation than i-NOS. It appears that in the brain, NO can regulate its own synthesis and therefore the neurotransmission process.
On the one hand, inhibition of ec-NOS will prevent the cytotoxicity associated with excessive nitric oxide production.
On the other, the insensitivity of i-NOS to nitric oxide will enable high levels of nitric oxide to be produced for cytotoxic effects.
Endogenous inhibitors of NOS (guanidino-substituted derivatives of arginine) occur in vivo as a result of post-translational modification of protein contained arginine residues by S-adenosylmethionine. The dimethylarginines (NG,NG-dimethyl-L-arginine and NG,N’G-dimethyl-L-arginine) occurs in tissue proteins, plasma, and urine of humans and they are thought to act as both regulators of NOS activity and reservoirs of arginine for the synthesis of nitric oxide.
It has been calculated that even though membrane makes up about 3% of the total tissue volume, 90% of the reaction of nitric oxide with oxygen occurs within this compartment. Thus the membrane is an important site for nitric oxide chemistry.
There are two major aspects to nitric oxide chemistry.
It can undergo single electron oxidation and reduction reactions producing nitrosonium and nitroxyl
Having a single unpaired electron in its π*2p molecular orbital it will react readily with other molecules that also have unpaired electrons, such as free radicals and transition metals.
Examples of the reaction of nitric oxide with radical species include:
Nitric oxide will react with oxygen to form the peroxynitrite (nitrosyldioxyl) radical (ONO2)
and with superoxide to form the powerful oxidizing and nitrating agent, peroxynitrite anion (ONO2-). Peroxynitrite causes damage to many important biomolecules
Importance:
nitrosothiols that are important in the regulation of blood pressure terminates lipid peroxidation
3-nitrosotyrosine and/or 4-O-nitrosotyrosine can affect the activity of enzymes that utilize tyrosyl radicals
rapidly reacts with oxyhemoglobin, the primary route of its destruction in vivo
the reaction between nitric oxide and transition metal complexes
During the last reaction a “ligand” bond is formed (the unpaired electron of nitric oxide is partially transferred to the metal cation),
resulting in a nitrosated (nitrosylated) complex.
For example, such complexes can be formed with free iron ions,
iron bound to heme or iron located in iron-sulfur clusters.
Ligand formation allows nitric oxide to act as a signal, activating some enzymes while inhibiting others. Thus, the binding of nitric oxide to the Fe (II)-heme of guanylate (guanalyl) cyclase [GTP-pyrophosphate lyase: cyclizing] is the signal transduction mechanism. Guanylate cyclase exists as cytosolic and membrane-bound isozymes. Thiol-Didulfide Redox Couple
The thiol-disulfide redox couple is very important to oxidative metabolism. For example, GSH is a reducing cofactor for glutathione peroxidase, an antioxidant enzyme responsible for the destruction of hydrogen peroxide.
The importance of the antioxidant role of the thiol-disulfide redox couple:
Thiols and disulfides can readily undergo exchange reactions, forming mixed disulfides. Thiol-disulfide exchange is biologically very important. For example,
GSH can react with protein cystine groups and influence the correct folding of proteins.
GSH may also play a direct role in cellular signaling through thiol-disulfide exchange reactions with membrane bound receptor proteins
the insulin receptor complex)
transcription factors (e.g., nuclear factor κB)
and regulatory proteins in cells
Conditions that alter the redox status of the cell can have important consequences on cellular function.
The generation of ROS by redox cycling is only one possible explanation for the action of many drugs. Rifamycin not only owes its activity to ROS generation but also to its ability to block bacterial RNA synthesis as well. Quinones (and/or semiquinones) can also form adducts with nucleophiles, especially thiols. These adducts may act as toxins directly or indirectly through the inhibition of key enzymes (e.g., by reacting with essential cysteinyl residues) or the depletion of GSH. DNA Adduct Formation
By far the most intense research in this field has been directed towards the chemistry and biology of DNA adduct formation. Attack of the free bases and nucleosides by pro-oxidants can yield a wide variety of adducts and DNA-protein cross-links. Such attack usually occurs
at the C-4 and C-8 position of purines and
C-5 and C-6 of pyrimidines.
Hydroxyl free radical-induced damage to purine bases and nucleosides can proceed through a C-8-hydroxy N-7 radical intermediate, and then either undergo oxidation with the production of an 8-hydroxy purine, or reduction, probably by cellular thiols, followed by ring opening and the formation of FAPy (formamido-pyrimidine) metabolites (hydroxyl free radical-induced damage to guanosine). Although most research has focused on 8-hydroxy-purine adducts a growing number of publications are attempting to measure the FAPy derivative.
Nitrosation of the Amines of the Nucleic Acid Bases.
Primary aromatic amines produce deaminated products, while secondary amines form N-nitroso compounds. Formation of Peroxynitrite from Nitric Oxide.
Peroxynitrite shows complex reactivity
with DNA initiating DNA strand breakage, oxidation (e.g., formation of 8-hydroxyguanine, 8-OH2’dG, (5-hydroxymethyl)-uracil, and FAPyGua),
nitration (e.g., 8-nitroguanine), and
deamination of bases.
Peroxynitrite can also promote the production of lipid peroxidation related active carbonyls and cause the activation of NAD+ ADP-ribosyltransferase.
Modification of Guanine
Although all DNA bases can be oxidatively damaged, it is the modification of guanine that is the most frequent. 8OH2’dG is the most abundant DNA adduct. This can affect its hydrogen bonding between base-pairs. These base-pair substitutions are usually found clustered into areas called “hot spots”. Guanine normally binds to cytosine.
8OH2’dG, however, can form hydrogen bonds with adenine. The formation of 8OH2’dG in DNA can therefore result in a G→T transversion.
8-Hydroxyguanine was also shown to induce codon 12 activation of c-Ha-ras and K-ras in mammalian systems. G→T transversions are also the most frequent hot spot mutations found in the p53 supressor gene which is associated with human tumors.
Other mechanisms by which ROS/RNS can lead to mutations have been
proposed. Direct mechanisms include:
conformational changes in the DNA template that reduces the accuracy of replication by DNA polymerases
altered methylation of cytosine that affects gene control
Indirect mechanisms include:
Oxidative damage to proteins, including DNA polymerases and repair enzymes.
Damage to lipids causes the production of mutagenic carbonyl compounds
Misalignment mutagenesis (“slippery DNA”)
DNA Mismatch Repair 5 (Photo credit: Allen Gathman)
Repair of ROS/RNS-induced DNA Damage
The repair of damaged DNA is an ongoing and continuous process involving a
number of repair enzymes. Damaged DNA appears to be mended by two major mechanisms:
base excision repair (BER) and
nucleotide excision repair (NER)
Isolated DNA is found to contain low levels of damaged bases, so it appears that these repair processes are not completely effective. Base Excision Repair
BER is first started by DNA glycosylases which recognize specific base
modifications (e.g., 8OH2’dG). For example,
Formamido-pyrimidine-DNA glycosylase (Fpg protein) recognizes damaged purines such as 8-oxoguanine and FAPyGua.
Damaged pyrimidines are recognized by endonuclease III, which acts as both a glycosylase and AP endonuclease.
Glycosylases cleave the N-glycosylic bond between the damaged base and the sugar
Following the glycosylase step, AP endonucleases then remove the 3′-deoxyribose moiety by cleavage of the phosphodiester bonds thereby generating a 3’-hydroxyl group that can then be extended by DNA polymerase.
The final step in mending damaged DNA is the rejoining of the free ends of DNA by a DNA ligase. It also appears that the presence of 8-oxoguanine modified bases in DNA is not only a result of ROS attack on this macromolecule. Oxidized nucleosides and nucleotides from free cellular pools can also be incorporated into DNA by polymerases and cause AT to CG base substitution mutations.
Mitochondrial DNA Repair
The mitochondrion genome encodes the various complexes of the electron transport chain, but contains no genetic information for DNA repair enzymes. These enzymes must be obtained from the nucleus. As mitochondria are continuously producing DNA damaging pro-oxidant species, effective DNA repair mechanisms must exist within the mitochondrial matrix in order for these organelles to function. Mitochondria have a short existence, and excessively damaged mitochondria will be quickly removed. Mitochondria contain many BER enzymes and are proficient at repair, but they do not appear to repair damaged DNA by NER mechanisms.
Single Strand DNA Damage and PARP Activation
Single strand DNA breakage activates NAD+ ADP-ribosyltransferase (PARP). PARP is a protein-modifying, nucleotide-polymerizing enzyme and is found at high levels in the nucleus. Activated PARP
cleaves NAD+ into ADP-ribose and nicotinamide
then attaches the ADP-ribose units to a variety of nuclear proteins (including histones and its own automodification domain).
then polymerizes the initial ADP-ribose modification with other ADP-ribose units to form the nucleic acid-like polymer, poly (ADP) ribose.
PARP only appears to be involved with BER and not NER. In BER PARP does not appear to play a direct role but rather it probably helps by keeping the chromatin in a conformation that enables other repair enzymes to be effective. It may also provide temporary protection to DNA molecules while it is being repaired. Conflicting evidence suggests that PARP may not be an important DNA repair enzyme as cells from a PARP knockout mouse model have normal repair characteristics.
Activation of PARP can be dangerous to the cell. For each mole of ADP-ribose transferred, one mole of NAD+ is consumed, and through the regeneration of NAD+ four ATP molecules are wasted. Thus the activation of PARP can rapidly deplete a cell’s energy store and even lead to cell death. Some researchers suggest that this may be one mechanism whereby cells with excessive DNA damage are effectively removed. However, a variety of diseases may involve PARP overactivation including
circulatory shock,
CNS injury,
diabetes,
drug-induced cytotoxicity, and
inflammation.
The Indirect Pathway.
This (mutation) pathway does not involve oxidative damage to the protein per se. This process involves oxidative damage to the DNA molecule encoding the protein. Thus pro-oxidants can cause changes in the base sequence of the DNA molecule. If such base modification is in a coding region of DNA (exon) and not corrected, the DNA molecule may be transcribed incorrectly. Translation of the mutant mRNA can result in a mutant protein containing a wrong amino acid in its primary sequence. If this modified amino acid occurs in an essential part of the protein (e.g., the active site of an enzyme or a portion that alters folding), the function of that protein may be impaired. Fortunately, unlike modified DNA
that can pass from cell to cell during mitosis thereby continuing the production of mutant protein, damage to a protein is non-replicating and stops with its destruction.
The Direct Pathway
This (post-translational) pathway involves the action of a pro-oxidant on a protein resulting in
modification of amino acid residues,
the formation of carbonyl adducts,
cross-linking and
polypeptide chain fragmentation.
Such changes often result in altered protein conformation and/or activity. Proteins will produce a variety of carbonyl products when exposed to metal-based systems (metal/ascorbate and metal/hydrogen peroxide) in vitro. For example, histidine yields aspartate, asparagine and 2-oxoimidazoline, while proline produces glutamate, pyroglutamate, 4-hydroxyproline isomers, 2-pyrrolidone and γ-aminobutyric acid. Metal-based systems and other pro-oxidant conditions can oxidize methionine to its sulfoxide.
This portion of the presentation is endebted to THE HANDBOOK OF REDOX
BIOCHEMISTRY, Ian N. Acworth, August 2003, esa. (inacworth@esainc.com).
We shall now identify more recent work related to this presentation.
Oxygen and Oxidative Stress
The reduction of oxygen to water proceeds via one electron at a time. In the mitochondrial respiratory chain, Complex IV (cytochrome oxidase) retains all partially reduced intermediates until full reduction is achieved. Other redox centres in the electron transport chain, however, may leak electrons to oxygen, partially reducing this molecule to superoxide anion (O2_•). Even though O2_• is not a strong oxidant, it is a precursor of most other reactive oxygen species, and it also becomes involved in the propagation of oxidative chain reactions. Despite the presence of various antioxidant defences, the mitochondrion appears to be the main intracellular source of these oxidants. This review describes the main mitochondrial sources of reactive species and the antioxidant defences that evolved to prevent oxidative damage in all the mitochondrial compartments.
Reactive oxygen species (ROS) is a phrase used to describe a variety of molecules and free radicals (chemical species with one unpaired electron) derived from molecular oxygen. Molecular oxygen in the ground state is a bi-radical, containing two unpaired electrons in the outer shell (also known as a triplet state).
Since the two single electrons have the same spin, oxygen can only react with one electron at a time and therefore it is not very reactive with the two electrons in a chemical bond.
On the other hand, if one of the two unpaired electrons is excited and changes its spin, the resulting species (known as singlet oxygen) becomes a powerful oxidant as the two electrons with opposing spins can quickly react with other pairs of electrons, especially double bonds.
The formation of OH• is catalysed by reduced transition metals, which in turn may be re-reduced by O2 -•, propagating this process. In addition, O2-• may react with other radicals including nitric oxide (NO•) in a reaction controlled by the rate of diffusion of both radicals. The product, peroxynitrite, is also a very powerful oxidant. The oxidants derived from NO• have been recently called reactive nitrogen species (RNS).
‘Oxidative stress’ is an expression used to describe various deleterious processes resulting from an imbalance between the excessive formation of ROS and/or RNS and limited antioxidant defences.
Whilst small fluctuations in the steady-state concentration of these oxidants may actually play a role in intracellular signalling,
uncontrolled increases in the steady-state concentrations of these oxidants lead to free radical mediated chain reactions
which indiscriminately target
proteins,
lipids,
polysaccharides.
In vivo, O2-• is produced both enzymatically and nonenzymatically.
Enzymatic sources include
NADPH oxidases located on the cell membrane of
polymorphonuclear cells,
macrophages and
endothelial cells and
cytochrome P450-dependent oxygenases.
The proteolytic conversion of xanthine dehydrogenase to xanthine oxidase provides another enzymatic source of both O2 -• and H2O2 (and therefore constitutes a source of OH•) and has been proposed to mediate deleterious processes in vivo.
Given the highly reducing intramitochondrial environment, various respiratory components, including flavoproteins, iron–sulfur clusters and ubisemiquinone, are thermodynamically capable of transferring one electron to oxygen. Moreover, most steps in the respiratory chain involve single-electron reactions, further favouring the monovalent reduction of oxygen. On the other hand, the mitochondrion possesses various antioxidant defences designed to eliminate both O2- • and H2O2.
The rate of O2 -• formation by the respiratory chain is controlled primarily by mass action, increasing both when electron flow slows down (increasing the concentration of electron donors, R•) and when the concentration of oxygen increases (eqn (1); Turrens et al. 1982).
d[O2]/dt = k [O2] [R•].
The energy released as electrons flow through the respiratory chain is converted into a H+ gradient through the inner mitochondrial membrane (Mitchell, 1977). This gradient, in turn, dissipates through the ATP synthase complex (Complex V) and is responsible for the turning of a rotor-like protein complex required for ATP synthesis. In the absence of ADP,
the movement of H+ through ATP synthase ceases and
the H+ gradient builds up
causing electron flow to slow down and
the respiratory chain to become more reduced (State IV respiration).
Mitochondrial Antioxidant Defences
The deleterious effects resulting from the formation of ROS in the mitochondrion are, to a large extent, prevented by various antioxidant systems. Superoxide is enzymatically converted to H2O2 by a family of metalloenzymes called superoxide dismutases (SOD). Since O2-• may either reduce transition metals, which in turn can react with H2O2 producing OH• or spontaneously react with NO• to produce peroxynitrite, it is important to maintain the steady-state concentration of O2-• at the lowest possible level. Thus, although the dismutation of O2-• to H2O2 and O2 can also occur spontaneously, the role of SODs is to increase the rate of the reaction to that of a diffusion-controlled process.
The mitochondrial matrix contains a specific form of SOD, with manganese in the active site, which eliminates the O2 -• formed in the matrix or on the inner side of the inner membrane. The expression of MnSOD is further induced by agents that cause oxidative stress, including radiation and hyperoxia, in a process mediated by the oxidative activation of the nuclear transcription factor NFkB .
Reactive oxygen species (ROS) are products of normal metabolism and xenobiotic exposure, and depending on their concentration, ROS can be beneficial or harmful to cells and tissues.
At physiological low levels, ROS function as “redox messengers” in intracellular signaling and regulation, whereas
excess ROS induce oxidative modification of cellular macromolecules, inhibit protein function, and promote cell death.
Additionally, various redox systems, such as
the glutathione,
thioredoxin, and
pyridine nucleotide redox couples,
NADPH and antioxidant defense
NAD+ and the function of sirtuin proteins
participate in cell signaling and modulation of cell function, including apoptotic cell death. Cell apoptosis is initiated by extracellular and intracellular signals via two main pathways,
the death receptor and
the mitochondria-mediated pathways.
ROS and JNK-mediated apoptotic signaling
GSH redox status and apoptotic signaling
Various pathologies can result from oxidative stress-induced apoptotic signaling that is consequent to
ROS increases and/or antioxidant decreases,
disruption of intracellular redox homeostasis, and
irreversible oxidative modifications of lipid, protein, or DNA.
We focus on several key aspects of ROS and redox mechanisms in apoptotic signaling and highlight the gaps in knowledge and potential avenues for further investigation. A full understanding of the redox control of apoptotic initiation and execution could underpin the development of therapeutic interventions targeted at oxidative stress-associated disorders.
Iron-sulfur (FeyS) cluster-containing proteins catalyze a number of electron transfer and metabolic reactions. The components and molecular mechanisms involved in the assembly of the FeyS clusters have been identified only partially. In eukaryotes, mitochondria have been proposed to execute a crucial task in the generation of intramitochondrial and extramitochondrial FeyS proteins. Herein, we identify the essential ferredoxin Yah1p of Saccharomyces cerevisiae mitochondria as a central component of the FeyS protein biosynthesis machinery. Depletion of Yah1p by regulated gene expression resulted in a
30-fold accumulation of iron within mitochondria,
similar to what has been reported for other components involved in FeyS protein biogenesis. Yah1p was shown to be required for the assembly of FeyS proteins both inside mitochondria and in the cytosol. Apparently, at least one of the steps of FeyS cluster biogenesis within mitochondria requires reduction by ferredoxin. Our findings lend support to the idea of a primary function of mitochondria in the biosynthesis of FeyS proteins outside the organelle. To our knowledge, Yah1p is the first member of the ferredoxin family for which a function in FeyS cluster formation has been established. A similar role may be predicted for the bacterial homologs that are encoded within iron-sulfur cluster assembly (isc) operons of prokaryotes.
H Lange, A Kaut, G Kispal, and R Lill. A mitochondrial ferredoxin is essential for biogenesis of cellular iron-sulfur proteins. PNAS 2000; 97(3): 1050–1055.
DNA Charge Transport
Damaged bases in DNA are known to lead to errors in replication and transcription, compromising the integrity of the genome. The authors proposed a model where repair proteins containing redoxactive [4Fe-4S] clusters utilize DNA charge transport (CT) as a first step in finding lesions. In this model, the population of sites to search is reduced by a localization of protein in the vicinity of lesions. Here, we examine this model using single-molecule atomic force microscopy (AFM). XPD, a 5′-3′ helicase involved in nucleotide
excision repair, contains a [4Fe-4S] cluster and exhibits a DNA bound redox potential that is physiologically relevant.
In AFM studies, they observe the redistribution of XPD onto kilobase DNA strands containing a single base mismatch, which is not a specific substrate for XPD but, like a lesion, inhibits CT. They also provide evidence for DNA-mediated signaling between XPD and Endonuclease III (EndoIII), a base excision repair glycosylase that also contains a [4Fe-4S] cluster.
When XPD and EndoIII are mixed together, they coordinate in relocalizing onto the mismatched strand.
However, when a CT-deficient mutant of either repair protein is combined with the CT-proficient repair partner, no relocalization occurs.
The data presented here indicate that XPD, an archaeal protein from the NER pathway, may cooperate with other proteins that are proficient at DNA CT to localize in the vicinity of damage. XPD, a superfamily 2 DNA helicase with 5′-3′ polarity, is a component of TFIIH that is essential for repair of bulky lesions generated by exogenous sources such as UV light and chemical carcinogens. XPD contains a conserved [4Fe-4S] cluster suggested to be conformationally controlled by ATP binding and hydrolysis.
Mutations in the iron-sulfur domain of XPD can lead to diseases including TTD and XP, yet the function of the [4Fe-4S] cluster appears to be unknown.
Electrochemical studies have shown that when BER proteins MutY and EndoIII bind to DNA, their [4Fe-4S] clusters are activated toward one electron oxidation. XPD exhibits a DNA-bound midpoint potential similar to that of EndoIII and MutY when bound to DNA (approximately 80 mV vs. NHE), indicative of a possible role for the [4Fe-4S] cluster in DNA-mediated CT.
For EndoIII we have also already determined a direct correlation between the ability of proteins to redistribute in the vicinity of mismatches as measured by AFM, and the CT proficiency of the proteins measured electrochemically. Thus, we may utilize single-molecule AFM as a tool to probe the redistribution of proteins in the vicinity of base lesions and in so doing, the proficiency of the protein to carry out DNA CT.
Here we show that, like the BER protein EndoIII, XPD, involved both in transcription and NER, redistributes in the vicinity of a lesion. Importantly, this ability to relocalize is associated with the ability of XPD to carry out DNA CT. The mutant L325V is defective in its ability to carry out DNA CTand this XPD mutant also does not redistribute effectively onto the mismatched strand.
These data not only indicate a general link between the ability of a repair protein to carry out DNA CT and its ability to redistribute onto DNA strands near lesions but also provide evidence for coordinated DNA CT between different repair proteins in their search for damage in the genome. These data also provide evidence that two different repair proteins, each containing a [4Fe-4S] cluster at similar DNA bound potential, can communicate with one another through DNA-mediated CT.
The signaling function of mitochondria is considered with a special emphasis on their role in the regulation of redox status of the cell, possibly determining a number of pathologies including cancer and aging. The review summarizes the transport role of mitochondria in energy supply to all cellular compartments (mitochondria as an electric cable in the cell), the role of mitochondria in plastic metabolism of the cell including synthesis of
heme,
steroids,
iron-sulfur clusters, and
reactive oxygen and nitrogen species.
Mitochondria also play an important role in the Ca2+-signaling and the regulation of apoptotic cell death. Knowledge of mechanisms responsible for apoptotic cell death is important for the strategy for prevention of unwanted degradation of postmitotic cells such as cardiomyocytes and neurons.
In accordance with P. Mitchell’s chemiosmotic concept, vectorial transmembrane transfer of electrons and protons is accompanied by generation of electrochemical difference of proton electrochemical potential on the inner mitochondrial membrane; its utilization by ATP synthase induces conformational rearrangements resulting in ATP synthesis from ADP and inorganic phosphate. Details of the mechanism responsible for ATP synthesis are given elsewhere.
Membrane potential (DY) generated across the inner mitochondrial membrane is the component of the transmembrane electrochemical potential of H+ ions (DμH+), which provides ATP synthesis together with the concentration component (DpH). Maintenance of constant membrane potential is a vitally important precondition for functioning of mitochondria and the cell. Under conditions of limited supply of the cell with oxygen (hypoxia) and inability to carry out aerobic ATP synthesis, mitochondria become ATP consumers (rather than generators) and ATP is hydrolyzed by mitochondrial ATPase, and this is accompanied by generation of membrane potential.
Redox homeostasis, i.e. the sum of redox components (including proteins, low molecular weight redox components such as NAD/NADH, flavins, coenzymes Q, oxidized and reduced substrates, etc.) is one of important preconditions for normal cell functioning.
Single-strand and double-strand DNA damage (Photo credit: Wikipedia)
Mitochondria generate such potent regulators of redox potential as
superoxide anion,
hydrogen peroxide,
nitric oxide,
peroxynitrite, etc.
They are actively involved in regulation of cell redox potential and consequently
control proteolysis,
activation of transcription,
changes in mitochondrial DNA (mDNA),
cell metabolism, and
cell differentiation.
Zorov DB, Isaev NK, Plotnikov EY, Zorova LD, et al. The Mitochondrion as Janus Bifrons. Biochemistry (Moscow) 2007; 72(10): 1115-1126. ISSN 0006-2979.
DOI: 10.1134/S0006297907100094
Structure of the human mitochondrial genome. (Photo credit: Wikipedia)
Gene Expression Associated with Oxidoreduction and Mitochondria
The naked mole-rat (Heterocephalus glaber) is a long-lived, cancer resistant rodent and there is a great interest in identifying the adaptations responsible for these and other of its unique traits. We employed RNA sequencing to compare liver gene expression profiles between naked mole-rats and wild-derived mice. Our results indicate that genes associated with oxidoreduction and mitochondria were expressed at higher relative levels in naked mole-rats. The largest effect is nearly
300-fold higher expression of epithelial cell adhesion molecule (Epcam), a tumour-associated protein.
Also of interest are the
protease inhibitor, alpha2-macroglobulin (A2m), and the
mitochondrial complex II subunit Sdhc,
both ageing-related genes found strongly over-expressed in the naked mole-rat.
These results hint at possible candidates for specifying species differences in ageing and cancer, and in particular suggest complex alterations in mitochondrial and oxidation reduction pathways in the naked mole-rat. Our differential gene expression analysis obviated the need for a reference naked mole-rat genome by employing a combination of Illumina/Solexa and 454 platforms for transcriptome sequencing and assembling transcriptome contigs of the non-sequenced species. Overall, our work provides new research foci and methods for studying the naked mole-rat’s fascinating characteristics.
The complete set of viable deletion strains in Saccharomyces cerevisiae was screened for sensitivity of mutants to five oxidants to identify cell functions involved in resistance to oxidative stress. This screen identified a unique set of mainly constitutive functions providing the first line of defense against a particular oxidant; these functions are very dependent on the nature of the oxidant. Most of these functions are distinct from those involved in repair and recovery from damage, which are generally induced in response to stress, because there was little correlation between mutant sensitivity and the reported transcriptional response to oxidants of the relevant gene. The screen identified 456 mutants sensitive to at least one of five different types of oxidant, and these were ranked in order of sensitivity. Many genes identified were not previously known to have a role in resistance to reactive oxygen species. These encode functions including
protein sorting,
ergosterol metabolism,
autophagy, and
vacuolar acidification.
two mutants were sensitive to all oxidants examined, 12 were sensitive to at least four,
Different oxidants had very different spectra of deletants that were sensitive. These findings highlight the specificity of cellular responses to different oxidants:
No single oxidant is representative of general oxidative stress.
Mitochondrial respiratory functions were overrepresented in mutants sensitive to H2O2, and
vacuolar protein-sorting mutants were enriched in mutants sensitive to diamide.
Core functions required for a broad range of oxidative-stress resistance include
Isolated complex I deficiency is the most common enzymatic defect of the oxidative phosphorylation (OXPHOS) system, causing a wide range of clinical phenotypes. Th authers reported before that the rates at which reactive oxygen species (ROS)-sensitive dyes are converted into their fluorescent oxidation products are markedly increased in cultured skin fibroblasts of patients with nuclear-inherited isolated complex I deficiency.
Using videoimaging microscopy we show here that these cells also display a marked increase in NAD(P)H autofluorescence. Linear regression analysis revealed a negative correlation with the residual complex I activity and a positive correlation with the oxidation rates of the ROS sensitive dyes (5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein and hydroethidine for a large cohort of 10 patient cell lines.
On the other hand, video-imaging microscopy of cells selectively expressing reduction-oxidation sensitive GFP1 in either the mitochondrial matrix or cytosol showed the absence of any detectable change in thiol redox state. In agreement with this result, neither the glutathione nor the glutathione disulfide content differed significantly between patient and healthy fibroblasts.
Finally, video-rate confocal microscopy of cells loaded with C11-BODIPY581/591 demonstrated that the extent of lipid peroxidation, which is regarded as a measure of oxidative damage, was not altered in patient fibroblasts. Our results indicate that fibroblasts of patients with isolated complex I deficiency maintain their thiol redox state despite marked increases in ROS production.
Mitochondrial research has recently been driven by the
identification of mitochondria-associated diseases and the role of mitochondria in apoptosis.
Moreover, mitochondria have been implicated in the process of carcinogenesis because of their vital role in
energy production,
nuclear-cytoplasmic signal integration and
control of metabolic pathways.
At some point during neoplastic transformation, there is an increase in reactive oxygen species (ROS), which damage the mitochondrial genome. This accelerates the somatic mutation rate of mitochondrial DNA.
Mitochondrial characteristics
There are several biological characteristics which cast mitochondria and, in particular, the mitochondrial genome, as a biological tool for early detection and monitoring of neoplasia and its potential progression. These vital characteristics are important in cancer research, as not all neoplasias become malignant. Mitochondria are archived in the cytoplasm of the ovum and as such do not recombine.
This genome has an accelerated mutation rate, by comparison with the nucleus, and accrues somatic mutations in tumour tissue. Moreover, mitochondrial DNA (mtDNA) has a high copy number in comparison with the nuclear archive of DNA. There are potentially thousands of mitochondrial genomes per cell, which enables detection of important biomarkers, even at low levels. In addition, mtDNA can be heteroplasmic, which means that disease-associated mutations occur in a subset of the genomes.
The presence of heteroplasmy is an indication of disease and is found in many human tumours. Identification of low levels of heteroplasmy may allow unprecedented early identification and monitoring of neoplastic progression to malignancy.
Coding for just 13 enzyme complex subunits, 22 transfer RNAs and two ribosomal RNAs, the mitochondrial genome is packaged in a compact 16,569 base pair (bp) circular molecule. These products participate in the critical electron transport process of ATP production. Collectively, mitochondria generate 80 per cent of the chemical fuel which fires cellular metabolism.
As a result, nuclear investment in the mitochondria is high — that is, several thousand nuclear genes control this organelle in order to accomplish the complex interactions required to maintain a network of pathways, which coordinate energy demand and supply.
It has been proposed that these mutations may serve as an early indication of potential cancer development and may represent a means for tracking tumour progression.
Does this provide a potential utility in that these mutations may be used for the identification and monitoring of neoplasia and malignant transformation where appropriate body fluids or non-invasive tissue access is available for mtDNA recovery? Specifically discussed are:
prostate,
breast,
colorectal,
skin and
lung cancers
There are many important questions yet to be addressed: such as
the relationship between mtDNA and the actual disease;
are mutations causative or merely a reflection of nuclear instability?
And, are these processes independent events?
Alterations in the non-coding D-loop suggest genome instability;
however, as studies focus more on the coding regions of the
mitochondrial genome,
Particularly in the case of nonsynonymous mutations in the genes
contributing products to the electron transport process, metabolic
implications are evident. Moreover, mutations in mitochondrial transfer RNAs indicate the possibility of a global mitochondrial translational shut down.
RL Parr, GD Dakubo, RE Thayer, K McKenney, MA Birch-Machin. Mitochondrial DNA as a potential tool for early cancer detection. HUMAN GENOMICS 2006; 2(4). 252–257.
Mitochondrial DNA (mtDNA) is particularly prone to oxidation due to the lack of histones and a deficient mismatch repair system. This explains an increased mutation rate of mtDNA that results in heteroplasmy, e.g., the coexistence of the mutant and wild-type mtDNA molecules within the same mitochondrion. Hyperglycemia is a key risk factor not only for diabetes-related disease, but also for cardiovascular and all-cause mortality. One can assume an increase in the risk of cardiovascular disease by 18% for each unit (%) glycated hemoglobin HbA1c. In the Glucose Tolerance in Acute Myocardial Infarction study of patients with acute coronary syndrome, abnormal glucose tolerance was the strongest independent predictor of subsequent cardiovascular complications and death. In the Asian Pacific Study, fasting plasma glucose was shown to be an independent predictor of cardiovascular events up to a level of 5.2 mmol/L.
Glucose level fluctuations and hyperglycemia are triggers for inflammatory responses via increased mitochondrial superoxide production and endoplasmic reticulum stress. Inflammation leads to insulin resistance and β-cell dysfunction, which further aggravates hyperglycemia. The molecular pathways that integrate hyperglycemia, oxidative stress, and diabetic vascular complications have been most clearly described in the pathogenesis of endothelial dysfunction, which is considered as the first step in atherogenesis according to the response to injury hypothesis.
In diabetes mellitus,
glycotoxicity,
advanced oxidative stress,
collagen cross-linking, and
accumulation of lipid peroxides
in foam macrophage cells and arterial wall cells may significantly
decrease the mutation threshold,
endothelial dysfunction,
promoting atherosclerosis.
Alterations in mitochondrial DNA (mtDNA), known as homoplasmic and heteroplasmic mutations, may influence mitochondrial OXPHOS capacity, and in turn contribute to the magnitude of oxidative stress in micro- and macrovascular networks in diabetic patients.
The authors critically consider the impact of mtDNA mutations on the pathogenesis of cardiovascular diabetic complications.
Mutation Threshhold
Although cells may harbor mutant mtDNA, the expression of disease is dependent on the percent of alleles bearing mutations. Modeling confirms that an upper threshold level might exist for mutations beyond which the mitochondrial population collapses, with a subsequent decrease in ATP. This decrease in ATP results in the phenotypic expression of disease. It is estimated that in many patients with clinical manifestations of mitochondrial disorders, the proportion of mutant DNA exceeds 50%.
For the MELAS (mitochondrial encephalopathy, lactic acidosis and stroke-like syndrome)-causing mutation m.3243 A>G in the mitochondrial gene encoding tRNALeu, which is also associated with diabetes plus deafness, a strong correlation between the level of mutational heteroplasmy and documented disease has been found. Increased percentages of mutant mtDNA in muscle cells (up to 71%) can lead to mitochondrial myopathy. Levels of heteroplasmy of over 80% may lead to recurrent stroke and mutation levels of 95% have been associated with MELAS.
Regardless of the type of mutation or the level of heteroplasmy in affected mitochondria, unrepaired damage leads to a decrease in ATP, which in turn causes the phenotypic manifestation of disease. The manifestation of disease not only depends on the ATP level but also on the tissue affected. Various tissues have differing levels of demand on OXPHOS capacity. To evaluate a tissue threshold, Leber’s hereditary optic neuropathy can be used as a model for mitochondrial neurodegenerative disease. For neural and skeletal muscle tissues, the tissue threshold should be as high as or higher than 90% of
damaged (mutated) mtDNA. To induce mitochondrial malfunctions, the tissue threshold of the cardiac muscle is estimated to be significantly lower (approximately 64%-67%). In chronic vascular disease such as atherosclerosis, a mutation threshold in the affected vessel wall (e.g., in the postmortem aortic atherosclerotic plaques) was observed to be significantly lower. For example, for mutations m.3256 C>T, m.12315 G>A, m.15059 G>A, and m.15315 G>A, the heteroplasmy range of 18%-66% in the atherosclerotic lesions was 2-3.5-fold that in normal vascular tissue.
Mitochondrial stress and insulin resistance
Mitochondrial damage precedes the development of atherosclerosis and tracks the extent of the lesion in apoE-null mice, and
mitochondrial dysfunction caused by heterozygous deficiency of a superoxide dismutase increases atherosclerosis and vascular mitochondrial damage in the same model.
Blood vessels destined to develop atherosclerosis may be characterized by inefficient ATP production due to the uncoupling of respiration and OXPHOS. Blood vessels have regions of hypoxia, which lower the ratio of state 3 (phosphorylating) to state 4 (nonphosphorylating) respiration. Human atherosclerotic lesions have been known for decades to be deficient in essential fatty acids, a condition that causes respiratory uncoupling and atherosclerosis.
The finding by Kokaze et al. helps to explain, at least in part, the anti-atherogenic effect of the allele m. 5178A due to its relation with the favorable lipid profile. The nucleotide change causes leucine-to-methionine substitution at codon 237 (Leu-237Met) of the NADH dehydrogenase subunit 2 located in the loop between 7th and 8th transmembrane domains of the mitochondrial protein. Given that this methionine residue is exposed at the surface of respiratory Complex I, this residue may be available as an efficient oxidant scavenger. Complex I
accepts electrons from NADH,
transfers them to ubiquinone, and
uses the energy released to pump protons across the mitochondrial inner membrane.
Thus, the Leu237Met replacement in the ND2 subunit might have a protective effect against oxidative damage to mitochondria.
Most fatty acid oxidation, which is promoted by peroxisome proliferator-activated receptor α (PPARα) activation, occurs in the mitochondria. Mitochondrial effects could explain why PPARα- deficient mice are protected from diet-induced insulin resistance and atherosclerosis as well as glucocorticoid induced insulin resistance and hypertension. Caloric restriction,

























































{kind=link}
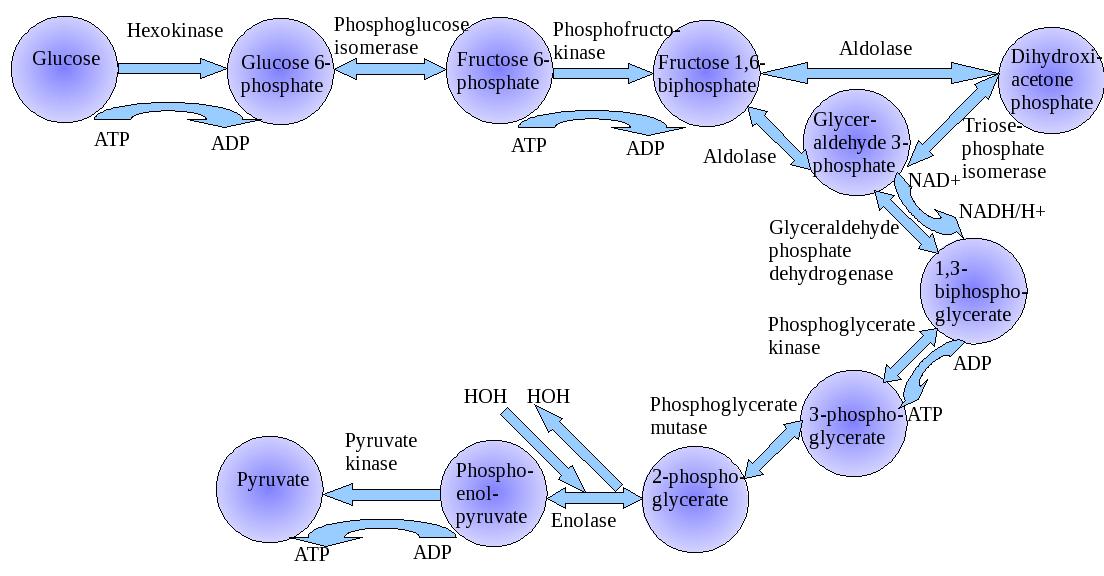{kind=link}