|
Posts Tagged ‘genomics’




Cancer Initiatives
Posted in Big Data, Big Data & Analytics, CANCER BIOLOGY & Innovations in Cancer Therapy, Cancer Genomics, Cancer Informatics, Clinical Genomics, Data Science, tagged Cancer Genomics, Cancer research, Childhood cancer, data sharing, genomics, Joe Biden, Moonshot, pediatric cancer on April 23, 2016| Leave a Comment »
Cancer initiatives
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Updated 4/12/2019
AACR 2016: Biden Calls for Overhauling Cancer Research Incentives
The first priority cited by the vice president was data sharing. Biden defended the concept as essential to advancing the process of cancer research and countered a January 21 New England Journal of Medicine editorial in which editor-in-chief Jeffrey Drazen, M.D., contended that data sharing could breed data “parasites.”
Four days later, Dr. Drazen clarified NEJM’s position by adding that with “appropriate systems” in place, “we will require a commitment from authors to make available the data that underlie the reported results of their work within 6 months after we publish them.”
Other priorities Biden said should serve as the basis of new incentives:
- Involve patients in clinical trial design—Raising awareness of trials, and allowing patients to participate in how they are designed and conducted, could help address the difficulty of recruiting patients for studies. Only 4% of cancer patients are involved in a trial, he said.
- “Let scientists do science”—Biden contrasted unfavorably NIH’s roughly 1-year process for decisions on grants to that of the Prostate Cancer Foundation, which limits grant applications to 10 pages and decides on those funding requests within 30 days: “Why is it that it takes multiple submissions and more than a year to get an answer from us?” Biden said.
- Encourage grants from younger researchers—Biden decried the current professional system under which younger researchers are sidetracked for years doing administrative work in labs before they can pursue their own research grants: “It’s like asking Derek Jeter to take several years off to sell bonds to build Yankee Stadium,” the VP quipped.
- Measure progress by outcomes—Rather than the quantity of research papers generated by grants, Biden said, “what you propose and how it affects patients, it seems to me, should be the basis of whether you continue to get the grant.”
- Promote open-access publication of results—Biden criticized academic publishing’s reliance on paid-subscription journals that block content behind paywalls and which own data for up to a year. He contrasted that system with the Bill and Melinda Gates Foundation’s stipulation that the research it funds be published in an open-access journal and be freely available once published.
- Reward verification—Research that verifies results through replication should be encouraged, Biden said, which acknowledging that few people now get such funding.
Biden recalled how following Beau’s diagnosis with cancer, he and his wife Jill Biden, Ed.D., who introduced the VP at the AACR event, “had access to the best doctors in the world.”
“The more we talked to them, the more we understood that we are on the cusp of a real inflection point in the fight against cancer.”
Updated 4/12/2019
Pediatric Cancer Initiatives
Data Sharing for Pediatric Cancers: President Trump Announces Pledge to Fight Childhood Cancer Will Involve Genomic Data Sharing Effort
In the journal Science, Drs. Olena Morozova Vaske ( and David Haussler University of California, Santa Cruz) recently wrote an editorial entitled “Data Sharing for Pediatric Cancers“, in which they discuss the implications of President Trump’s intentions to increase funding for pediatric cancers with a corresponding effort for genomic data sharing. Also discussed is the current efforts on pediatric genomic data sharing as well as some opinions on coordinating these efforts on a world-wide scale to benefit the patients, researchers, and clinicians.
The article is found below as it is a very good read on the state of data sharing in the pediatric cancer field and offers some very good insights in designing such a worldwide system to handle this data sharing, including allowing patients governance over their own data.
Last month, in a conference call held by the U.S. Department of Health and Human Services and National Institutes of Health (NIH), it was revealed that a large focus of President Trump’s pledge to fund childhood cancer research will be genomic data sharing. Although the United States has only 5% of the world’s pediatric cancer cases, it has disproportionately more resources and access to genomic information compared to low-income countries. We hope that the spotlight on genomic data sharing in the United States will galvanize the world’s pediatric cancer community to elevate genomic data sharing to a level where its full potential can finally be realized.
Pediatric cancers are rare, affecting 50 to 200 children per million a year worldwide. Thus, with 16 different major types and many subtypes, no cancer center encounters large cohorts of patients with the same diagnosis. To advance their understanding of particular cancer subtypes, pediatric oncologists must have access to data from similar cases at other centers. Because subtypes of pediatric cancer are rare, assembling large cohorts is a limiting factor in clinical trials as well. Here, too, data sharing is the first critical step.
Typically, pediatric cancers don’t have the number of mutations that make immunotherapies effective, and only a few subtypes have recurrent mutations that can be used to develop gene-targeted therapies. However, the abnormal expression level of genes gives a vivid picture of genetic misregulation, and just sharing this information would be a huge step forward. Using gene expression and mutation data, analysis of genetic misregulation in different pediatric cancer subtypes could point the way to new treatments.
A major challenge in genomic data sharing is the patient’s young age, which frequently precludes an opportunity for informed consent. Compounding this, the rarity of subtypes requires the aggregation of patients from multiple jurisdictions, raising barriers to assembling large representative data sets. A greater percentage of children than adults with cancer participate in research studies, and children often participate in multiple studies. However, this means that data collected on individual children may be found at multiple institutions, creating difficulties if there are no standards for data sharing.
To enable effective sharing of genomic and clinical data, the Global Alliance for Genomics and Health has developed the Key Implications for Data Sharing (KIDS) framework for pediatric genomics. The recommendations include involving children in the data-sharing decision-making process and imposing an ethical obligation on data generators to provide children and parents with the opportunity to share genomic and clinical information with researchers. Although KIDS guidelines are not legally binding, they could inform policy development worldwide.
To advance the sharing culture, along with the NIH, pediatric cancer foundations such as the St. Baldrick’s Foundation and Alex’s Lemonade Stand Foundation have incorporated genomic data-sharing requirements into their grants processes. Researchers and clinicians around the world have created dozens of pediatric cancer genomic databases and portals, but pulling these together into a larger network is problematic, especially for patients with data at more than one institution, as patient identifiers are stripped from shared data. However, initiatives like the Children’s Oncology Group’s Project Every Child and the European Network for Cancer Research in Children and Adolescents’ Unified Patient Identity may resolve this issue.
We urge the creators of pediatric cancer genomic resources to collaborate and build a real-time federated data-sharing system, and hope that the new U.S. initiative will inspire other countries to link databases rather than just create new siloed regional resources. The great advances in information technology and life sciences in the last decades have given us a new opportunity to save our children from the scourge of cancer. We must resolve to use them.
Source: Data sharing for pediatric cancers.
NIH-NCI Initiative: International collaboration to create new cancer models to accelerate research
LIVE 1:45 pm – 3:10 pm 4/25/2016 Forum Opening, A War or Moonshot: Where Do We Stand? Creating a Disruptive Cancer Pipeline @2016 World Medical Innovation Forum: CANCER, April 25-27, 2016, Westin Hotel, Boston
Will President Obama’ s Cancer Immunotherapy Colloquium (dubbed Moonshot) mean Government is Fully Behind the War on Cancer or have we heard this before?
Exome Aggregation Consortium (ExAC), generated the largest catalogue so far of variation in human protein-coding regions: Sequence data of 60,000 people, NOW is a publicly accessible database
Healthcare conglomeration to access Big Data and lower costs
CRISPR/Cas9, Familial Amyloid Polyneuropathy ( FAP) and Neurodegenerative Disease
Posted in Alzheimer's Disease, Biological Networks, Gene Regulation and Evolution, Cell Biology, Signaling & Cell Circuits, Cerebrovascular and Neurodegenerative Diseases, Chemical Biology and its relations to Metabolic Disease, Chemical Genetics, Clinical & Translational, Computational Biology/Systems and Bioinformatics, CRISPR/Cas9 & Gene Editing, Curation, Disease Biology, Small Molecules in Development of Therapeutic Drugs, Drug Toxicity, Epigenetics and Environmental Factors, Experimental validation, FDA Regulatory Affairs, Gene Regulation and Evolution, Genetics & Innovations in Treatment, Genetics & Pharmaceutical, Genome Biology, Genomic Testing: Methodology for Diagnosis, Heart Failure (HF), Human aging, Innovations, Innovations in Neurophysiology & Neuropsychology, ISO 10993 for Product Registration: FDA & CE Mark for Development of Medical Devices and Diagnostics, Metabolism, Metabolomics, Microbiology, Molecular Genetics & Pharmaceutical, Mutant Gene Expression, Neurological Diseases, Neuroscience, Pharmaceutical Analytics, Pharmaceutical Drug Discovery, Pharmacologic toxicities, Pharmacotherapy and Cell Activity, Population genetics, Population Health Management, Genetics & Pharmaceutical, Proteins, Proteomics, RNA Biology, Signaling & Cell Circuits, Small Molecules in Development of Therapeutic Drugs, Technology Advance Assessment of, tagged amyloid in peripheral nerves and the heart, amyloid transthyretin polyneuropathy, amyloidogenic variants of the transthyretin protein, CRISPR-based gene editing, familial amyloidosis, familial amyloidotic cardiomyopathy, gene mutations, genetics, genomics, human chromosome 18q12.1-11.2, Molecular Biology, Portuguese and Swedish PAF, Protein folding, RNAi, Tafamidis, translational therapy, Transthyretin amyloidosis on April 17, 2016| Leave a Comment »

CRISPR/Cas9, Familial Amyloid Polyneuropathy ( FAP) and Neurodegenerative Disease
Curator: Larry H. Bernstein, MD, FCAP
CRISPR/Cas9 and Targeted Genome Editing: A New Era in Molecular Biology
The development of efficient and reliable ways to make precise, targeted changes to the genome of living cells is a long-standing goal for biomedical researchers. Recently, a new tool based on a bacterial CRISPR-associated protein-9 nuclease (Cas9) from Streptococcus pyogenes has generated considerable excitement (1). This follows several attempts over the years to manipulate gene function, including homologous recombination (2) and RNA interference (RNAi) (3). RNAi, in particular, became a laboratory staple enabling inexpensive and high-throughput interrogation of gene function (4, 5), but it is hampered by providing only temporary inhibition of gene function and unpredictable off-target effects (6). Other recent approaches to targeted genome modification – zinc-finger nucleases [ZFNs, (7)] and transcription-activator like effector nucleases [TALENs (8)]– enable researchers to generate permanent mutations by introducing doublestranded breaks to activate repair pathways. These approaches are costly and time-consuming to engineer, limiting their widespread use, particularly for large scale, high-throughput studies.
The Biology of Cas9
The functions of CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and CRISPR-associated (Cas) genes are essential in adaptive immunity in select bacteria and archaea, enabling the organisms to respond to and eliminate invading genetic material. These repeats were initially discovered in the 1980s in E. coli (9), but their function wasn’t confirmed until 2007 by Barrangou and colleagues, who demonstrated that S. thermophilus can acquire resistance against a bacteriophage by integrating a genome fragment of an infectious virus into its CRISPR locus (10).
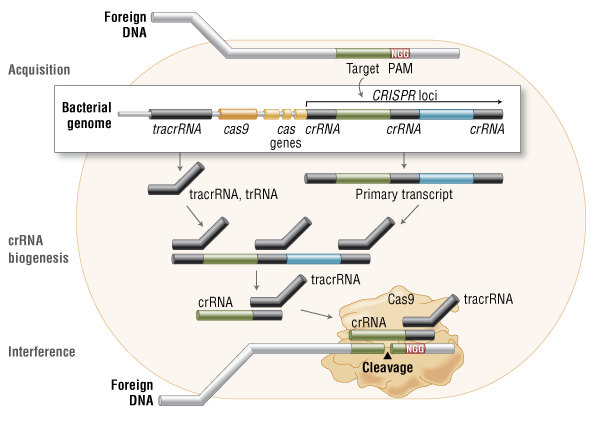
Three types of CRISPR mechanisms have been identified, of which type II is the most studied. In this case, invading DNA from viruses or plasmids is cut into small fragments and incorporated into a CRISPR locus amidst a series of short repeats (around 20 bps). The loci are transcribed, and transcripts are then processed to generate small RNAs (crRNA – CRISPR RNA), which are used to guide effector endonucleases that target invading DNA based on sequence complementarity (Figure 1) (11).
Figure 1. Cas9 in vivo: Bacterial Adaptive Immunity
https://www.neb.com/~/media/NebUs/Files/Feature%20Articles/Images/FA_Cas9_Fig1_Cas9InVivo.png
{kind=link}
In the acquisition phase, foreign DNA is incorporated into the bacterial genome at the CRISPR loci. CRISPR loci is then transcribed and processed into crRNA during crRNA biogenesis. During interference, Cas9 endonuclease complexed with a crRNA and separate tracrRNA cleaves foreign DNA containing a 20-nucleotide crRNA complementary sequence adjacent to the PAM sequence. (Figure not drawn to scale.)
https://www.neb.com/~/media/NebUs/Files/Feature%20Articles/Images/FA_Cas9_GenomeEditingGlossary.png
{kind=link}
One Cas protein, Cas9 (also known as Csn1), has been shown, through knockdown and rescue experiments to be a key player in certain CRISPR mechanisms (specifically type II CRISPR systems). The type II CRISPR mechanism is unique compared to other CRISPR systems, as only one Cas protein (Cas9) is required for gene silencing (12). In type II systems, Cas9 participates in the processing of crRNAs (12), and is responsible for the destruction of the target DNA (11). Cas9’s function in both of these steps relies on the presence of two nuclease domains, a RuvC-like nuclease domain located at the amino terminus and a HNH-like nuclease domain that resides in the mid-region of the protein (13).
To achieve site-specific DNA recognition and cleavage, Cas9 must be complexed with both a crRNA and a separate trans-activating crRNA (tracrRNA or trRNA), that is partially complementary to the crRNA (11). The tracrRNA is required for crRNA maturation from a primary transcript encoding multiple pre-crRNAs. This occurs in the presence of RNase III and Cas9 (12).
During the destruction of target DNA, the HNH and RuvC-like nuclease domains cut both DNA strands, generating double-stranded breaks (DSBs) at sites defined by a 20-nucleotide target sequence within an associated crRNA transcript (11, 14). The HNH domain cleaves the complementary strand, while the RuvC domain cleaves the noncomplementary strand.
The double-stranded endonuclease activity of Cas9 also requires that a short conserved sequence, (2–5 nts) known as protospacer-associated motif (PAM), follows immediately 3´- of the crRNA complementary sequence (15). In fact, even fully complementary sequences are ignored by Cas9-RNA in the absence of a PAM sequence (16).
Cas9 and CRISPR as a New Tool in Molecular Biology
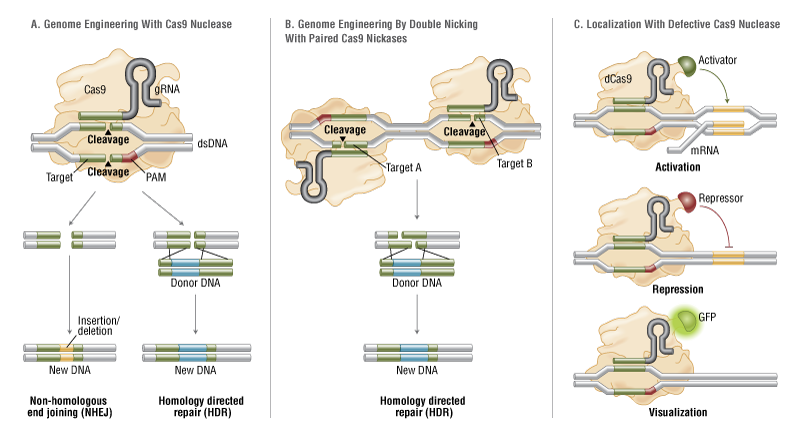
The simplicity of the type II CRISPR nuclease, with only three required components (Cas9 along with the crRNA and trRNA) makes this system amenable to adaptation for genome editing. This potential was realized in 2012 by the Doudna and Charpentier labs (11). Based on the type II CRISPR system described previously, the authors developed a simplified two-component system by combining trRNA and crRNA into a single synthetic single guide RNA (sgRNA). sgRNAprogrammed Cas9 was shown to be as effective as Cas9 programmed with separate trRNA and crRNA in guiding targeted gene alterations (Figure 2A).
To date, three different variants of the Cas9 nuclease have been adopted in genome-editing protocols. The first is wild-type Cas9, which can site-specifically cleave double-stranded DNA, resulting in the activation of the doublestrand break (DSB) repair machinery. DSBs can be repaired by the cellular Non-Homologous End Joining (NHEJ) pathway (17), resulting in insertions and/or deletions (indels) which disrupt the targeted locus. Alternatively, if a donor template with homology to the targeted locus is supplied, the DSB may be repaired by the homology-directed repair (HDR) pathway allowing for precise replacement mutations to be made (Figure 2A) (17, 18).
Cong and colleagues (1) took the Cas9 system a step further towards increased precision by developing a mutant form, known as Cas9D10A, with only nickase activity. This means it cleaves only one DNA strand, and does not activate NHEJ. Instead, when provided with a homologous repair template, DNA repairs are conducted via the high-fidelity HDR pathway only, resulting in reduced indel mutations (1, 11, 19). Cas9D10A is even more appealing in terms of target specificity when loci are targeted by paired Cas9 complexes designed to generate adjacent DNA nicks (20) (see further details about “paired nickases” in Figure 2B).
The third variant is a nuclease-deficient Cas9 (dCas9, Figure 2C) (21). Mutations H840A in the HNH domain and D10A in the RuvC domain inactivate cleavage activity, but do not prevent DNA binding (11, 22). Therefore, this variant can be used to sequence-specifically target any region of the genome without cleavage. Instead, by fusing with various effector domains, dCas9 can be used either as a gene silencing or activation tool (21, 23–26). Furthermore, it can be used as a visualization tool. For instance, Chen and colleagues used dCas9 fused to Enhanced Green Fluorescent Protein (EGFP) to visualize repetitive DNA sequences with a single sgRNA or nonrepetitive loci using multiple sgRNAs (27).
Figure 2. CRISPR/Cas9 System Applications
{kind=link}
- Wild-type Cas9 nuclease site specifically cleaves double-stranded DNA activating double-strand break repair machinery. In the absence of a homologous repair template non-homologous end joining can result in indels disrupting the target sequence. Alternatively, precise mutations and knock-ins can be made by providing a homologous repair template and exploiting the homology directed repair pathway.
B. Mutated Cas9 makes a site specific single-strand nick. Two sgRNA can be used to introduce a staggered double-stranded break which can then undergo homology directed repair.
C. Nuclease-deficient Cas9 can be fused with various effector domains allowing specific localization. For example, transcriptional activators, repressors, and fluorescent proteins.
Targeting Efficiency and Off-target Mutations
Targeting efficiency, or the percentage of desired mutation achieved, is one of the most important parameters by which to assess a genome-editing tool. The targeting efficiency of Cas9 compares favorably with more established methods, such as TALENs or ZFNs (8). For example, in human cells, custom-designed ZFNs and TALENs could only achieve efficiencies ranging from 1% to 50% (29–31). In contrast, the Cas9 system has been reported to have efficiencies up to >70% in zebrafish (32) and plants (33), and ranging from 2–5% in induced pluripotent stem cells (34). In addition, Zhou and colleagues were able to improve genome targeting up to 78% in one-cell mouse embryos, and achieved effective germline transmission through the use of dual sgRNAs to simultaneously target an individual gene (35).
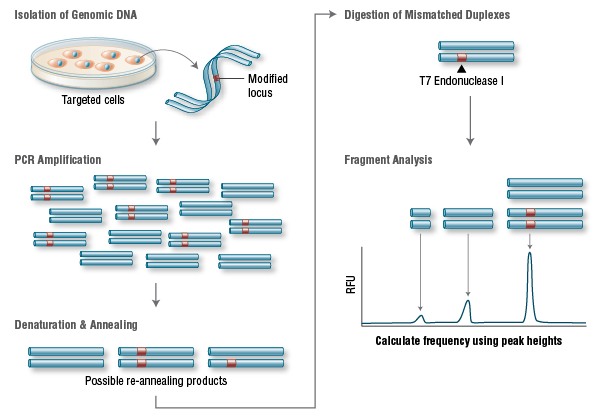
A widely used method to identify mutations is the T7 Endonuclease I mutation detection assay (36, 37) (Figure 3). This assay detects heteroduplex DNA that results from the annealing of a DNA strand, including desired mutations, with a wildtype DNA strand (37).
Figure 3. T7 Endonuclease I Targeting Efficiency Assay
{kind=link}
Genomic DNA is amplified with primers bracketing the modified locus. PCR products are then denatured and re-annealed yielding 3 possible structures. Duplexes containing a mismatch are digested by T7 Endonuclease I. The DNA is then electrophoretically separated and fragment analysis is used to calculate targeting efficiency.
Another important parameter is the incidence of off-target mutations. Such mutations are likely to appear in sites that have differences of only a few nucleotides compared to the original sequence, as long as they are adjacent to a PAM sequence. This occurs as Cas9 can tolerate up to 5 base mismatches within the protospacer region (36) or a single base difference in the PAM sequence (38). Off-target mutations are generally more difficult to detect, requiring whole-genome sequencing to rule them out completely.
Recent improvements to the CRISPR system for reducing off-target mutations have been made through the use of truncated gRNA (truncated within the crRNA-derived sequence) or by adding two extra guanine (G) nucleotides to the 5´ end (28, 37). Another way researchers have attempted to minimize off-target effects is with the use of “paired nickases” (20). This strategy uses D10A Cas9 and two sgRNAs complementary to the adjacent area on opposite strands of the target site (Figure 2B). While this induces DSBs in the target DNA, it is expected to create only single nicks in off-target locations and, therefore, result in minimal off-target mutations.
By leveraging computation to reduce off-target mutations, several groups have developed webbased tools to facilitate the identification of potential CRISPR target sites and assess their potential for off-target cleavage. Examples include the CRISPR Design Tool (38) and the ZiFiT Targeter, Version 4.2 (39, 40).
Applications as a Genome-editing and Genome Targeting Tool
Following its initial demonstration in 2012 (9), the CRISPR/Cas9 system has been widely adopted. This has already been successfully used to target important genes in many cell lines and organisms, including human (34), bacteria (41), zebrafish (32), C. elegans (42), plants (34), Xenopus tropicalis (43), yeast (44), Drosophila (45), monkeys (46), rabbits (47), pigs (42), rats (48) and mice (49). Several groups have now taken advantage of this method to introduce single point mutations (deletions or insertions) in a particular target gene, via a single gRNA (14, 21, 29). Using a pair of gRNA-directed Cas9 nucleases instead, it is also possible to induce large deletions or genomic rearrangements, such as inversions or translocations (50). A recent exciting development is the use of the dCas9 version of the CRISPR/Cas9 system to target protein domains for transcriptional regulation (26, 51, 52), epigenetic modification (25), and microscopic visualization of specific genome loci (27).
The CRISPR/Cas9 system requires only the redesign of the crRNA to change target specificity. This contrasts with other genome editing tools, including zinc finger and TALENs, where redesign of the protein-DNA interface is required. Furthermore, CRISPR/Cas9 enables rapid genome-wide interrogation of gene function by generating large gRNA libraries (51, 53) for genomic screening.
The Future of CRISPR/Cas9
The rapid progress in developing Cas9 into a set of tools for cell and molecular biology research has been remarkable, likely due to the simplicity, high efficiency and versatility of the system. Of the designer nuclease systems currently available for precision genome engineering, the CRISPR/Cas system is by far the most user friendly. It is now also clear that Cas9’s potential reaches beyond DNA cleavage, and its usefulness for genome locus-specific recruitment of proteins will likely only be limited by our imagination.
Scientists urge caution in using new CRISPR technology to treat human genetic disease
By Robert Sanders, Media relations | MARCH 19, 2015
http://news.berkeley.edu/2015/03/19/scientists-urge-caution-in-using-new-crispr-technology-to-treat-human-genetic-disease/
http://news.berkeley.edu/wp-content/uploads/2015/03/crispr350.jpg
{kind=link}
The bacterial enzyme Cas9 is the engine of RNA-programmed genome engineering in human cells. (Graphic by Jennifer Doudna/UC Berkeley)
A group of 18 scientists and ethicists today warned that a revolutionary new tool to cut and splice DNA should be used cautiously when attempting to fix human genetic disease, and strongly discouraged any attempts at making changes to the human genome that could be passed on to offspring.
Among the authors of this warning is Jennifer Doudna, the co-inventor of the technology, called CRISPR-Cas9, which is driving a new interest in gene therapy, or “genome engineering.” She and colleagues co-authored a perspective piece that appears in the March 20 issue of Science, based on discussions at a meeting that took place in Napa on Jan. 24. The same issue of Science features a collection of recent research papers, commentary and news articles on CRISPR and its implications. …..
A prudent path forward for genomic engineering and germline gene modification
David Baltimore1, Paul Berg2, …., Jennifer A. Doudna4,10,*, et al.
http://science.sciencemag.org/content/early/2015/03/18/science.aab1028.full
Science 19 Mar 2015. http://dx.doi.org:/10.1126/science.aab1028
Correcting genetic defects
Scientists today are changing DNA sequences to correct genetic defects in animals as well as cultured tissues generated from stem cells, strategies that could eventually be used to treat human disease. The technology can also be used to engineer animals with genetic diseases mimicking human disease, which could lead to new insights into previously enigmatic disorders.
The CRISPR-Cas9 tool is still being refined to ensure that genetic changes are precisely targeted, Doudna said. Nevertheless, the authors met “… to initiate an informed discussion of the uses of genome engineering technology, and to identify proactively those areas where current action is essential to prepare for future developments. We recommend taking immediate steps toward ensuring that the application of genome engineering technology is performed safely and ethically.”
Amyloid CRISPR Plasmids and si/shRNA Gene Silencers
http://www.scbt.com/crispr/table-amyloid.html
Santa Cruz Biotechnology, Inc. offers a broad range of gene silencers in the form of siRNAs, shRNA Plasmids and shRNA Lentiviral Particles as well as CRISPR/Cas9 Knockout and CRISPR Double Nickase plasmids. Amyloid gene silencers are available as Amyloid siRNA, Amyloid shRNA Plasmid, Amyloid shRNA Lentiviral Particles and Amyloid CRISPR/Cas9 Knockout plasmids. Amyloid CRISPR/dCas9 Activation Plasmids and CRISPR Lenti Activation Systems for gene activation are also available. Gene silencers and activators are useful for gene studies in combination with antibodies used for protein detection. Amyloid CRISPR Knockout, HDR and Nickase Knockout Plasmids
CRISPR-Cas9-Based Knockout of the Prion Protein and Its Effect on the Proteome
Mehrabian M, Brethour D, MacIsaac S, Kim JK, Gunawardana C.G, Wang H, et al.
PLoS ONE 2014; 9(12): e114594. http://dx.doi.org/10.1371/journal.pone.0114594
The molecular function of the cellular prion protein (PrPC) and the mechanism by which it may contribute to neurotoxicity in prion diseases and Alzheimer’s disease are only partially understood. Mouse neuroblastoma Neuro2a cells and, more recently, C2C12 myocytes and myotubes have emerged as popular models for investigating the cellular biology of PrP. Mouse epithelial NMuMG cells might become attractive models for studying the possible involvement of PrP in a morphogenetic program underlying epithelial-to-mesenchymal transitions. Here we describe the generation of PrP knockout clones from these cell lines using CRISPR-Cas9 knockout technology. More specifically, knockout clones were generated with two separate guide RNAs targeting recognition sites on opposite strands within the first hundred nucleotides of the Prnp coding sequence. Several PrP knockout clones were isolated and genomic insertions and deletions near the CRISPR-target sites were characterized. Subsequently, deep quantitative global proteome analyses that recorded the relative abundance of>3000 proteins (data deposited to ProteomeXchange Consortium) were undertaken to begin to characterize the molecular consequences of PrP deficiency. The levels of ∼120 proteins were shown to reproducibly correlate with the presence or absence of PrP, with most of these proteins belonging to extracellular components, cell junctions or the cytoskeleton.
Development and Applications of CRISPR-Cas9 for Genome Engineering
Patrick D. Hsu,1,2,3 Eric S. Lander,1 and Feng Zhang1,2,*
Cell. 2014 Jun 5; 157(6): 1262–1278. doi: 10.1016/j.cell.2014.05.010
Recent advances in genome engineering technologies based on the CRISPR-associated RNA-guided endonuclease Cas9 are enabling the systematic interrogation of mammalian genome function. Analogous to the search function in modern word processors, Cas9 can be guided to specific locations within complex genomes by a short RNA search string. Using this system, DNA sequences within the endogenous genome and their functional outputs are now easily edited or modulated in virtually any organism of choice. Cas9-mediated genetic perturbation is simple and scalable, empowering researchers to elucidate the functional organization of the genome at the systems level and establish causal linkages between genetic variations and biological phenotypes. In this Review, we describe the development and applications of Cas9 for a variety of research or translational applications while highlighting challenges as well as future directions. Derived from a remarkable microbial defense system, Cas9 is driving innovative applications from basic biology to biotechnology and medicine.
The development of recombinant DNA technology in the 1970s marked the beginning of a new era for biology. For the first time, molecular biologists gained the ability to manipulate DNA molecules, making it possible to study genes and harness them to develop novel medicine and biotechnology. Recent advances in genome engineering technologies are sparking a new revolution in biological research. Rather than studying DNA taken out of the context of the genome, researchers can now directly edit or modulate the function of DNA sequences in their endogenous context in virtually any organism of choice, enabling them to elucidate the functional organization of the genome at the systems level, as well as identify causal genetic variations.
Broadly speaking, genome engineering refers to the process of making targeted modifications to the genome, its contexts (e.g., epigenetic marks), or its outputs (e.g., transcripts). The ability to do so easily and efficiently in eukaryotic and especially mammalian cells holds immense promise to transform basic science, biotechnology, and medicine (Figure 1).
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4343198/bin/nihms659174f1.jpg
{kind=link}
For life sciences research, technologies that can delete, insert, and modify the DNA sequences of cells or organisms enable dissecting the function of specific genes and regulatory elements. Multiplexed editing could further allow the interrogation of gene or protein networks at a larger scale. Similarly, manipulating transcriptional regulation or chromatin states at particular loci can reveal how genetic material is organized and utilized within a cell, illuminating relationships between the architecture of the genome and its functions. In biotechnology, precise manipulation of genetic building blocks and regulatory machinery also facilitates the reverse engineering or reconstruction of useful biological systems, for example, by enhancing biofuel production pathways in industrially relevant organisms or by creating infection-resistant crops. Additionally, genome engineering is stimulating a new generation of drug development processes and medical therapeutics. Perturbation of multiple genes simultaneously could model the additive effects that underlie complex polygenic disorders, leading to new drug targets, while genome editing could directly correct harmful mutations in the context of human gene therapy (Tebas et al., 2014).
Eukaryotic genomes contain billions of DNA bases and are difficult to manipulate. One of the breakthroughs in genome manipulation has been the development of gene targeting by homologous recombination (HR), which integrates exogenous repair templates that contain sequence homology to the donor site (Figure 2A) (Capecchi, 1989). HR-mediated targeting has facilitated the generation of knockin and knockout animal models via manipulation of germline competent stem cells, dramatically advancing many areas of biological research. However, although HR-mediated gene targeting produces highly precise alterations, the desired recombination events occur extremely infrequently (1 in 106–109 cells) (Capecchi, 1989), presenting enormous challenges for large-scale applications of gene-targeting experiments.
Genome Editing Technologies Exploit Endogenous DNA Repair Machinery
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4343198/bin/nihms659174f2.gif
{kind=link}
To overcome these challenges, a series of programmable nuclease-based genome editing technologies have been developed in recent years, enabling targeted and efficient modification of a variety of eukaryotic and particularly mammalian species. Of the current generation of genome editing technologies, the most rapidly developing is the class of RNA-guided endonucleases known as Cas9 from the microbial adaptive immune system CRISPR (clustered regularly interspaced short palindromic repeats), which can be easily targeted to virtually any genomic location of choice by a short RNA guide. Here, we review the development and applications of the CRISPR-associated endonuclease Cas9 as a platform technology for achieving targeted perturbation of endogenous genomic elements and also discuss challenges and future avenues for innovation. ……
Figure 4 Natural Mechanisms of Microbial CRISPR Systems in Adaptive Immunity
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4343198/bin/nihms659174f4.gif
{kind=link}
…… A key turning point came in 2005, when systematic analysis of the spacer sequences separating the individual direct repeats suggested their extrachromosomal and phage-associated origins (Mojica et al., 2005; Pourcel et al., 2005; Bolotin et al., 2005). This insight was tremendously exciting, especially given previous studies showing that CRISPR loci are transcribed (Tang et al., 2002) and that viruses are unable to infect archaeal cells carrying spacers corresponding to their own genomes (Mojica et al., 2005). Together, these findings led to the speculation that CRISPR arrays serve as an immune memory and defense mechanism, and individual spacers facilitate defense against bacteriophage infection by exploiting Watson-Crick base-pairing between nucleic acids (Mojica et al., 2005; Pourcel et al., 2005). Despite these compelling realizations that CRISPR loci might be involved in microbial immunity, the specific mechanism of how the spacers act to mediate viral defense remained a challenging puzzle. Several hypotheses were raised, including thoughts that CRISPR spacers act as small RNA guides to degrade viral transcripts in a RNAi-like mechanism (Makarova et al., 2006) or that CRISPR spacers direct Cas enzymes to cleave viral DNA at spacer-matching regions (Bolotin et al., 2005). …..
As the pace of CRISPR research accelerated, researchers quickly unraveled many details of each type of CRISPR system (Figure 4). Building on an earlier speculation that protospacer adjacent motifs (PAMs) may direct the type II Cas9 nuclease to cleave DNA (Bolotin et al., 2005), Moineau and colleagues highlighted the importance of PAM sequences by demonstrating that PAM mutations in phage genomes circumvented CRISPR interference (Deveau et al., 2008). Additionally, for types I and II, the lack of PAM within the direct repeat sequence within the CRISPR array prevents self-targeting by the CRISPR system. In type III systems, however, mismatches between the 5′ end of the crRNA and the DNA target are required for plasmid interference (Marraffini and Sontheimer, 2010). …..
In 2013, a pair of studies simultaneously showed how to successfully engineer type II CRISPR systems from Streptococcus thermophilus (Cong et al., 2013) andStreptococcus pyogenes (Cong et al., 2013; Mali et al., 2013a) to accomplish genome editing in mammalian cells. Heterologous expression of mature crRNA-tracrRNA hybrids (Cong et al., 2013) as well as sgRNAs (Cong et al., 2013; Mali et al., 2013a) directs Cas9 cleavage within the mammalian cellular genome to stimulate NHEJ or HDR-mediated genome editing. Multiple guide RNAs can also be used to target several genes at once. Since these initial studies, Cas9 has been used by thousands of laboratories for genome editing applications in a variety of experimental model systems (Sander and Joung, 2014). ……
The majority of CRISPR-based technology development has focused on the signature Cas9 nuclease from type II CRISPR systems. However, there remains a wide diversity of CRISPR types and functions. Cas RAMP module (Cmr) proteins identified in Pyrococcus furiosus and Sulfolobus solfataricus (Hale et al., 2012) constitute an RNA-targeting CRISPR immune system, forming a complex guided by small CRISPR RNAs that target and cleave complementary RNA instead of DNA. Cmr protein homologs can be found throughout bacteria and archaea, typically relying on a 5′ site tag sequence on the target-matching crRNA for Cmr-directed cleavage.
Unlike RNAi, which is targeted largely by a 6 nt seed region and to a lesser extent 13 other bases, Cmr crRNAs contain 30–40 nt of target complementarity. Cmr-CRISPR technologies for RNA targeting are thus a promising target for orthogonal engineering and minimal off-target modification. Although the modularity of Cmr systems for RNA-targeting in mammalian cells remains to be investigated, Cmr complexes native to P. furiosus have already been engineered to target novel RNA substrates (Hale et al., 2009, 2012). ……
Although Cas9 has already been widely used as a research tool, a particularly exciting future direction is the development of Cas9 as a therapeutic technology for treating genetic disorders. For a monogenic recessive disorder due to loss-of-function mutations (such as cystic fibrosis, sickle-cell anemia, or Duchenne muscular dystrophy), Cas9 may be used to correct the causative mutation. This has many advantages over traditional methods of gene augmentation that deliver functional genetic copies via viral vector-mediated overexpression—particularly that the newly functional gene is expressed in its natural context. For dominant-negative disorders in which the affected gene is haplosufficient (such as transthyretin-related hereditary amyloidosis or dominant forms of retinitis pigmentosum), it may also be possible to use NHEJ to inactivate the mutated allele to achieve therapeutic benefit. For allele-specific targeting, one could design guide RNAs capable of distinguishing between single-nucleotide polymorphism (SNP) variations in the target gene, such as when the SNP falls within the PAM sequence.
CRISPR/Cas9: a powerful genetic engineering tool for establishing large animal models of neurodegenerative diseases
Zhuchi Tu, Weili Yang, Sen Yan, Xiangyu Guo and Xiao-Jiang Li
Molecular Neurodegeneration 2015; 10:35 http://dx.doi.org:/10.1186/s13024-015-0031-x
Animal models are extremely valuable to help us understand the pathogenesis of neurodegenerative disorders and to find treatments for them. Since large animals are more like humans than rodents, they make good models to identify the important pathological events that may be seen in humans but not in small animals; large animals are also very important for validating effective treatments or confirming therapeutic targets. Due to the lack of embryonic stem cell lines from large animals, it has been difficult to use traditional gene targeting technology to establish large animal models of neurodegenerative diseases. Recently, CRISPR/Cas9 was used successfully to genetically modify genomes in various species. Here we discuss the use of CRISPR/Cas9 technology to establish large animal models that can more faithfully mimic human neurodegenerative diseases.
Neurodegenerative diseases — Alzheimer’s disease(AD),Parkinson’s disease(PD), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), and frontotemporal dementia (FTD) — are characterized by age-dependent and selective neurodegeneration. As the life expectancy of humans lengthens, there is a greater prevalence of these neurodegenerative diseases; however, the pathogenesis of most of these neurodegenerative diseases remain unclear, and we lack effective treatments for these important brain disorders.
CRISPR/Cas9, Non-human primates, Neurodegenerative diseases, Animal model
There are a number of excellent reviews covering different types of neurodegenerative diseases and their genetic mouse models [8–12]. Investigations of different mouse models of neurodegenerative diseases have revealed a common pathology shared by these diseases. First, the development of neuropathology and neurological symptoms in genetic mouse models of neurodegenerative diseases is age dependent and progressive. Second, all the mouse models show an accumulation of misfolded or aggregated proteins resulting from the expression of mutant genes. Third, despite the widespread expression of mutant proteins throughout the body and brain, neuronal function appears to be selectively or preferentially affected. All these facts indicate that mouse models of neurodegenerative diseases recapitulate important pathologic features also seen in patients with neurodegenerative diseases.
However, it seems that mouse models can not recapitulate the full range of neuropathology seen in patients with neurodegenerative diseases. Overt neurodegeneration, which is the most important pathological feature in patient brains, is absent in genetic rodent models of AD, PD, and HD. Many rodent models that express transgenic mutant proteins under the control of different promoters do not replicate overt neurodegeneration, which is likely due to their short life spans and the different aging processes of small animals. Also important are the remarkable differences in brain development between rodents and primates. For example, the mouse brain takes 21 days to fully develop, whereas the formation of primate brains requires more than 150 days [13]. The rapid development of the brain in rodents may render neuronal cells resistant to misfolded protein-mediated neurodegeneration. Another difficulty in using rodent models is how to analyze cognitive and emotional abnormalities, which are the early symptoms of most neurodegenerative diseases in humans. Differences in neuronal circuitry, anatomy, and physiology between rodent and primate brains may also account for the behavioral differences between rodent and primate models.
Mitochondrial dynamics–fusion, fission, movement, and mitophagy–in neurodegenerative diseases
Hsiuchen Chen and David C. Chan
Human Molec Gen 2009; 18, Review Issue 2 R169–R176
http://dx.doi.org:/10.1093/hmg/ddp326
Neurons are metabolically active cells with high energy demands at locations distant from the cell body. As a result, these cells are particularly dependent on mitochondrial function, as reflected by the observation that diseases of mitochondrial dysfunction often have a neurodegenerative component. Recent discoveries have highlighted that neurons are reliant particularly on the dynamic properties of mitochondria. Mitochondria are dynamic organelles by several criteria. They engage in repeated cycles of fusion and fission, which serve to intermix the lipids and contents of a population of mitochondria. In addition, mitochondria are actively recruited to subcellular sites, such as the axonal and dendritic processes of neurons. Finally, the quality of a mitochondrial population is maintained through mitophagy, a form of autophagy in which defective mitochondria are selectively degraded. We review the general features of mitochondrial dynamics, incorporating recent findings on mitochondrial fusion, fission, transport and mitophagy. Defects in these key features are associated with neurodegenerative disease. Charcot-Marie-Tooth type 2A, a peripheral neuropathy, and dominant optic atrophy, an inherited optic neuropathy, result from a primary deficiency of mitochondrial fusion. Moreover, several major neurodegenerative diseases—including Parkinson’s, Alzheimer’s and Huntington’s disease—involve disruption of mitochondrial dynamics. Remarkably, in several disease models, the manipulation of mitochondrial fusion or fission can partially rescue disease phenotypes. We review how mitochondrial dynamics is altered in these neurodegenerative diseases and discuss the reciprocal interactions between mitochondrial fusion, fission, transport and mitophagy.
Applications of CRISPR–Cas systems in Neuroscience
Matthias Heidenreich & Feng Zhang
Nature Rev Neurosci 2016; 17:36–44 http://dx.doi.org:/10.1038/nrn.2015.2
Genome-editing tools, and in particular those based on CRISPR–Cas (clustered regularly interspaced short palindromic repeat (CRISPR)–CRISPR-associated protein) systems, are accelerating the pace of biological research and enabling targeted genetic interrogation in almost any organism and cell type. These tools have opened the door to the development of new model systems for studying the complexity of the nervous system, including animal models and stem cell-derived in vitro models. Precise and efficient gene editing using CRISPR–Cas systems has the potential to advance both basic and translational neuroscience research.
Cellular neuroscience, DNA recombination, Genetic engineering, Molecular neuroscience
Figure 3: In vitro applications of Cas9 in human iPSCs.close
http://www.nature.com/nrn/journal/v17/n1/carousel/nrn.2015.2-f3.jpg
{kind=link}
a | Evaluation of disease candidate genes from large-population genome-wide association studies (GWASs). Human primary cells, such as neurons, are not easily available and are difficult to expand in culture. By contrast, induced pluripo…
Molecular Therapy 12 Jan 2016
Scientific Reports 31 Mar 2016
Scientific Reports 12 Nov 2015
Alzheimer’s Disease: Medicine’s Greatest Challenge in the 21st Century
https://www.physicsforums.com/insights/can-gene-editing-eliminate-alzheimers-disease/
The development of the CRISPR/Cas9 system has made gene editing a relatively simple task. While CRISPR and other gene editing technologies stand to revolutionize biomedical research and offers many promising therapeutic avenues (such as in the treatment of HIV), a great deal of debate exists over whether CRISPR should be used to modify human embryos. As I discussed in my previous Insight article, we lack enough fundamental biological knowledge to enhance many traits like height or intelligence, so we are not near a future with genetically-enhanced super babies. However, scientists have identified a few rare genetic variants that protect against disease. One such protective variant is a mutation in the APP gene that protects against Alzheimer’s disease and cognitive decline in old age. If we can perfect gene editing technologies, is this mutation one that we should be regularly introducing into embryos? In this article, I explore the potential for using gene editing as a way to prevent Alzheimer’s disease in future generations. Alzheimer’s Disease: Medicine’s Greatest Challenge in the 21st Century Can gene editing be the missing piece in the battle against Alzheimer’s? (Source: bostonbiotech.org) I chose to assess the benefit of germline gene editing in the context of Alzheimer’s disease because this disease is one of the biggest challenges medicine faces in the 21st century. Alzheimer’s disease is a chronic neurodegenerative disease responsible for the majority of the cases of dementia in the elderly. The disease symptoms begins with short term memory loss and causes more severe symptoms – problems with language, disorientation, mood swings, behavioral issues – as it progresses, eventually leading to the loss of bodily functions and death. Because of the dementia the disease causes, Alzheimer’s patients require a great deal of care, and the world spends ~1% of its total GDP on caring for those with Alzheimer’s and related disorders. Because the prevalence of the disease increases with age, the situation will worsen as life expectancies around the globe increase: worldwide cases of Alzheimer’s are expected to grow from 35 million today to over 115 million by 2050.
Despite much research, the exact causes of Alzheimer’s disease remains poorly understood. The disease seems to be related to the accumulation of plaques made of amyloid-β peptides that form on the outside of neurons, as well as the formation of tangles of the protein tau inside of neurons. Although many efforts have been made to target amyloid-β or the enzymes involved in its formation, we have so far been unsuccessful at finding any treatment that stops the disease or reverses its progress. Some researchers believe that most attempts at treating Alzheimer’s have failed because, by the time a patient shows symptoms, the disease has already progressed past the point of no return.
While research towards a cure continues, researchers have sought effective ways to prevent Alzheimer’s disease. Although some studies show that mental and physical exercise may lower ones risk of Alzheimer’s disease, approximately 60-80% of the risk for Alzheimer’s disease appears to be genetic. Thus, if we’re serious about prevention, we may have to act at the genetic level. And because the brain is difficult to access surgically for gene therapy in adults, this means using gene editing on embryos.
Reference https://www.physicsforums.com/insights/can-gene-editing-eliminate-alzheimers-disease/
Utilising CRISPR to Generate Predictive Disease Models: a Case Study in Neurodegenerative Disorders
Dr. Bhuvaneish.T. Selvaraj – Scottish Centre for Regenerative Medicine
- Introducing the latest developments in predictive model generation
- Discover how CRISPR is being used to develop disease models to study and treat neurodegenerative disorders
- In depth Q&A session to answer your most pressing questions
Turning On Genes, Systematically, with CRISPR/Cas9
Scientists based at MIT assert that they can reliably turn on any gene of their choosing in living cells. [Feng Zhang and Steve Dixon] http://www.genengnews.com/media/images/GENHighlight/Dec12_2014_CRISPRCas9GeneActivationSystem7838101231.jpg
{kind=link}
With the latest CRISPR/Cas9 advance, the exhortation “turn on, tune in, drop out” comes to mind. The CRISPR/Cas9 gene-editing system was already a well-known means of “tuning in” (inserting new genes) and “dropping out” (knocking out genes). But when it came to “turning on” genes, CRISPR/Cas9 had little potency. That is, it had demonstrated only limited success as a way to activate specific genes.
A new CRISPR/Cas9 approach, however, appears capable of activating genes more effectively than older approaches. The new approach may allow scientists to more easily determine the function of individual genes, according to Feng Zhang, Ph.D., a researcher at MIT and the Broad Institute. Dr. Zhang and colleagues report that the new approach permits multiplexed gene activation and rapid, large-scale studies of gene function.
The new technique was introduced in the December 10 online edition of Nature, in an article entitled, “Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex.” The article describes how Dr. Zhang, along with the University of Tokyo’s Osamu Nureki, Ph.D., and Hiroshi Nishimasu, Ph.D., overhauled the CRISPR/Cas9 system. The research team based their work on their analysis (published earlier this year) of the structure formed when Cas9 binds to the guide RNA and its target DNA. Specifically, the team used the structure’s 3D shape to rationally improve the system.
In previous efforts to revamp CRISPR/Cas9 for gene activation purposes, scientists had tried to attach the activation domains to either end of the Cas9 protein, with limited success. From their structural studies, the MIT team realized that two small loops of the RNA guide poke out from the Cas9 complex and could be better points of attachment because they allow the activation domains to have more flexibility in recruiting transcription machinery.
Using their revamped system, the researchers activated about a dozen genes that had proven difficult or impossible to turn on using the previous generation of Cas9 activators. Each gene showed at least a twofold boost in transcription, and for many genes, the researchers found multiple orders of magnitude increase in activation.
After investigating single-guide RNA targeting rules for effective transcriptional activation, demonstrating multiplexed activation of 10 genes simultaneously, and upregulating long intergenic noncoding RNA transcripts, the research team decided to undertake a large-scale screen. This screen was designed to identify genes that confer resistance to a melanoma drug called PLX-4720.
“We … synthesized a library consisting of 70,290 guides targeting all human RefSeq coding isoforms to screen for genes that, upon activation, confer resistance to a BRAF inhibitor,” wrote the authors of the Nature paper. “The top hits included genes previously shown to be able to confer resistance, and novel candidates were validated using individual [single-guide RNA] and complementary DNA overexpression.”
A gene signature based on the top screening hits, the authors added, correlated with a gene expression signature of BRAF inhibitor resistance in cell lines and patient-derived samples. It was also suggested that large-scale screens such as the one demonstrated in the current study could help researchers discover new cancer drugs that prevent tumors from becoming resistant.
Susceptibility and modifier genes in Portuguese transthyretin V30M amyloid polyneuropathy: complexity in a single-gene disease
Miguel L. Soares1,2, Teresa Coelho3,6, Alda Sousa4,5, …, Maria Joa˜o Saraiva2,5 and Joel N. Buxbaum1
Human Molec Gen 2005; 14(4): 543–553 http://dx.doi.org:/10.1093/hmg/ddi051
https://www.researchgate.net/profile/Isabel_Conceicao/publication/8081351_Susceptibility_and_modifier_genes_in_Portuguese_transthyretin_V30M_amyloid_polyneuropathy_complexity_in_a_single-gene_disease/links/53e123d70cf2235f352733b3.pdf
Familial amyloid polyneuropathy type I is an autosomal dominant disorder caused by mutations in the transthyretin (TTR ) gene; however, carriers of the same mutation exhibit variability in penetrance and clinical expression. We analyzed alleles of candidate genes encoding non-fibrillar components of TTR amyloid deposits and a molecule metabolically interacting with TTR [retinol-binding protein (RBP)], for possible associations with age of disease onset and/or susceptibility in a Portuguese population sample with the TTR V30M mutation and unrelated controls. We show that the V30M carriers represent a distinct subset of the Portuguese population. Estimates of genetic distance indicated that the controls and the classical onset group were furthest apart, whereas the late-onset group appeared to differ from both. Importantly, the data also indicate that genetic interactions among the multiple loci evaluated, rather than single-locus effects, are more likely to determine differences in the age of disease onset. Multifactor dimensionality reduction indicated that the best genetic model for classical onset group versus controls involved the APCS gene, whereas for late-onset cases, one APCS variant (APCSv1) and two RBP variants (RBPv1 and RBPv2) are involved. Thus, although the TTR V30M mutation is required for the disease in Portuguese patients, different genetic factors may govern the age of onset, as well as the occurrence of anticipation.
Autosomal dominant disorders may vary in expression even within a given kindred. The basis of this variability is uncertain and can be attributed to epigenetic factors, environment or epistasis. We have studied familial amyloid polyneuropathy (FAP), an autosomal dominant disorder characterized by peripheral sensorimotor and autonomic neuropathy. It exhibits variation in cardiac, renal, gastrointestinal and ocular involvement, as well as age of onset. Over 80 missense mutations in the transthyretin gene (TTR ) result in autosomal dominant disease http://www.ibmc.up.pt/~mjsaraiv/ttrmut.html). The presence of deposits consisting entirely of wild-type TTR molecules in the hearts of 10– 25% of individuals over age 80 reveals its inherent in vivo amyloidogenic potential (1).
FAP was initially described in Portuguese (2) where, until recently, the TTR V30M has been the only pathogenic mutation associated with the disease (3,4). Later reports identified the same mutation in Swedish and Japanese families (5,6). The disorder has since been recognized in other European countries and in North American kindreds in association with V30M, as well as other mutations (7).
TTR V30M produces disease in only 5–10% of Swedish carriers of the allele (8), a much lower degree of penetrance than that seen in Portuguese (80%) (9) or in Japanese with the same mutation. The actual penetrance in Japanese carriers has not been formally established, but appears to resemble that seen in Portuguese. Portuguese and Japanese carriers show considerable variation in the age of clinical onset (10,11). In both populations, the first symptoms had originally been described as typically occurring before age 40 (so-called ‘classical’ or early-onset); however, in recent years, more individuals developing symptoms late in life have been identified (11,12). Hence, present data indicate that the distribution of the age of onset in Portuguese is continuous, but asymmetric with a mean around age 35 and a long tail into the older age group (Fig. 1) (9,13). Further, DNA testing in Portugal has identified asymptomatic carriers over age 70 belonging to a subset of very late-onset kindreds in whose descendants genetic anticipation is frequent. The molecular basis of anticipation in FAP, which is not mediated by trinucleotide repeat expansions in the TTR or any other gene (14), remains elusive.
Variation in penetrance, age of onset and clinical features are hallmarks of many autosomal dominant disorders including the human TTR amyloidoses (7). Some of these clearly reflect specific biological effects of a particular mutation or a class of mutants. However, when such phenotypic variability is seen with a single mutation in the gene encoding the same protein, it suggests an effect of modifying genetic loci and/or environmental factors contributing differentially to the course of disease. We have chosen to examine age of onset as an example of a discrete phenotypic variation in the presence of the particular autosomal dominant disease-associated mutation TTR V30M. Although the role of environmental factors cannot be excluded, the existence of modifier genes involved in TTR amyloidogenesis is an attractive hypothesis to explain the phenotypic variability in FAP. ….
ATTR (TTR amyloid), like all amyloid deposits, contains several molecular components, in addition to the quantitatively dominant fibril-forming amyloid protein, including heparan sulfate proteoglycan 2 (HSPG2 or perlecan), SAP, a plasma glycoprotein of the pentraxin family (encoded by the APCS gene) that undergoes specific calcium-dependent binding to all types of amyloid fibrils, and apolipoprotein E (ApoE), also found in all amyloid deposits (15). The ApoE4 isoform is associated with an increased frequency and earlier onset of Alzheimer’s disease (Ab), the most common form of brain amyloid, whereas the ApoE2 isoform appears to be protective (16). ApoE variants could exert a similar modulatory effect in the onset of FAP, although early studies on a limited number of patients suggested this was not the case (17).
In at least one instance of senile systemic amyloidosis, small amounts of AA-related material were found in TTR deposits (18). These could reflect either a passive co-aggregation or a contributory involvement of protein AA, encoded by the serum amyloid A (SAA ) genes and the main component of secondary (reactive) amyloid fibrils, in the formation of ATTR.
Retinol-binding protein (RBP), the serum carrier of vitamin A, circulates in plasma bound to TTR. Vitamin A-loaded RBP and L-thyroxine, the two natural ligands of TTR, can act alone or synergistically to inhibit the rate and extent of TTR fibrillogenesis in vitro, suggesting that RBP may influence the course of FAP pathology in vivo (19). We have analyzed coding and non-coding sequence polymorphisms in the RBP4 (serum RBP, 10q24), HSPG2 (1p36.1), APCS (1q22), APOE (19q13.2), SAA1 and SAA2 (11p15.1) genes with the goal of identifying chromosomes carrying common and functionally significant variants. At the time these studies were performed, the full human genome sequence was not completed and systematic singlenucleotide polymorphism (SNP) analyses were not available for any of the suspected candidate genes. We identified new SNPs in APCS and RBP4 and utilized polymorphisms in SAA, HSPG2 and APOE that had already been characterized and shown to have potential pathophysiologic significance in other disorders (16,20–22). The genotyping data were analyzed for association with the presence of the V30M amyloidogenic allele (FAP patients versus controls) and with the age of onset (classical- versus late-onset patients). Multilocus analyses were also performed to examine the effects of simultaneous contributions of the six loci for determining the onset of the first symptoms. …..
The potential for different underlying models for classical and late onset is supported by the MDR analysis, which produces two distinct models when comparing each class with the controls. One could view the two onset classes as unique diseases. If this is the case, then the failure to detect a single predictive genetic model is consistent with two related, but different, diseases. This is exactly what would be expected in such a case of genetic heterogeneity (28). Using this approach, a major gene effect can be viewed as a necessary, but not sufficient, condition to explain the course of the disease. Analyzing the cases but omitting from the analysis of phenotype the necessary allele, in this case TTR V30M, can then reveal a variety of important modifiers that are distinct between the phenotypes.
The significant comparisons obtained in our study cohort indicate that the combined effects mainly result from two and three-locus interactions involving all loci except SAA1 and SAA2 for susceptibility to disease. A considerable number of four-site combinations modulate the age of onset with SAA1 appearing in a majority of significant combinations in late-onset disease, perhaps indicating a greater role of the SAA variants in the age of onset of FAP.
The correlation between genotype and phenotype in socalled simple Mendelian disorders is often incomplete, as only a subset of all mutations can reliably predict specific phenotypes (34). This is because non-allelic genetic variations and/or environmental influences underlie these disorders whose phenotypes behave as complex traits. A few examples include the identification of the role of homozygozity for the SAA1.1 allele in conferring the genetic susceptibility to renal amyloidosis in FMF (20) and the association of an insertion/deletion polymorphism in the ACE gene with disease severity in familial hypertrophic cardiomyopathy (35). In these disorders, the phenotypes arise from mutations in MEFV and b-MHC, but are modulated by independently inherited genetic variation. In this report, we show that interactions among multiple genes, whose products are confirmed or putative constituents of ATTR deposits, or metabolically interact with TTR, modulate the onset of the first symptoms and predispose individuals to disease in the presence of the V30M mutation in TTR. The exact nature of the effects identified here requires further study with potential application in the development of genetic screening with prognostic value pertaining to the onset of disease in the TTR V30M carriers.
If the effects of additional single or interacting genes dictate the heterogeneity of phenotype, as reflected in variability of onset and clinical expression (with the same TTR mutation), the products encoded by alleles at such loci could contribute to the process of wild-type TTR deposition in elderly individuals without a mutation (senile systemic amyloidosis), a phenomenon not readily recognized as having a genetic basis because of the insensitivity of family history in the elderly.
Safety and Efficacy of RNAi Therapy for Transthyretin Amyloidosis
Coelho T, Adams D, Silva A, et al.
N Engl J Med 2013;369:819-29. http://dx.doi.org:/10.1056/NEJMoa1208760
Transthyretin amyloidosis is caused by the deposition of hepatocyte-derived transthyretin amyloid in peripheral nerves and the heart. A therapeutic approach mediated by RNA interference (RNAi) could reduce the production of transthyretin.
Methods We identified a potent antitransthyretin small interfering RNA, which was encapsulated in two distinct first- and second-generation formulations of lipid nanoparticles, generating ALN-TTR01 and ALN-TTR02, respectively. Each formulation was studied in a single-dose, placebo-controlled phase 1 trial to assess safety and effect on transthyretin levels. We first evaluated ALN-TTR01 (at doses of 0.01 to 1.0 mg per kilogram of body weight) in 32 patients with transthyretin amyloidosis and then evaluated ALN-TTR02 (at doses of 0.01 to 0.5 mg per kilogram) in 17 healthy volunteers.
Results Rapid, dose-dependent, and durable lowering of transthyretin levels was observed in the two trials. At a dose of 1.0 mg per kilogram, ALN-TTR01 suppressed transthyretin, with a mean reduction at day 7 of 38%, as compared with placebo (P=0.01); levels of mutant and nonmutant forms of transthyretin were lowered to a similar extent. For ALN-TTR02, the mean reductions in transthyretin levels at doses of 0.15 to 0.3 mg per kilogram ranged from 82.3 to 86.8%, with reductions of 56.6 to 67.1% at 28 days (P<0.001 for all comparisons). These reductions were shown to be RNAi mediated. Mild-to-moderate infusion-related reactions occurred in 20.8% and 7.7% of participants receiving ALN-TTR01 and ALN-TTR02, respectively.
ALN-TTR01 and ALN-TTR02 suppressed the production of both mutant and nonmutant forms of transthyretin, establishing proof of concept for RNAi therapy targeting messenger RNA transcribed from a disease-causing gene.
Alnylam May Seek Approval for TTR Amyloidosis Rx in 2017 as Other Programs Advance
Officials from Alnylam Pharmaceuticals last week provided updates on the two drug candidates from the company’s flagship transthyretin-mediated amyloidosis program, stating that the intravenously delivered agent patisiran is proceeding toward a possible market approval in three years, while a subcutaneously administered version called ALN-TTRsc is poised to enter Phase III testing before the end of the year.
Meanwhile, Alnylam is set to advance a handful of preclinical therapies into human studies in short order, including ones for complement-mediated diseases, hypercholesterolemia, and porphyria.
The officials made their comments during a conference call held to discuss Alnylam’s second-quarter financial results.
ATTR is caused by a mutation in the TTR gene, which normally produces a protein that acts as a carrier for retinol binding protein and is characterized by the accumulation of amyloid deposits in various tissues. Alnylam’s drugs are designed to silence both the mutant and wild-type forms of TTR.
Patisiran, which is delivered using lipid nanoparticles developed by Tekmira Pharmaceuticals, is currently in a Phase III study in patients with a form of ATTR called familial amyloid polyneuropathy (FAP) affecting the peripheral nervous system. Running at over 20 sites in nine countries, that study is set to enroll up to 200 patients and compare treatment to placebo based on improvements in neuropathy symptoms.
According to Alnylam Chief Medical Officer Akshay Vaishnaw, Alnylam expects to have final data from the study in two to three years, which would put patisiran on track for a new drug application filing in 2017.
Meanwhile, ALN-TTRsc, which is under development for a version of ATTR that affects cardiac tissue called familial amyloidotic cardiomyopathy (FAC) and uses Alnylam’s proprietary GalNAc conjugate delivery technology, is set to enter Phase III by year-end as Alnylam holds “active discussions” with US and European regulators on the design of that study, CEO John Maraganore noted during the call.
In the interim, Alnylam continues to enroll patients in a pilot Phase II study of ALN-TTRsc, which is designed to test the drug’s efficacy for FAC or senile systemic amyloidosis (SSA), a condition caused by the idiopathic accumulation of wild-type TTR protein in the heart.
Based on “encouraging” data thus far, Vaishnaw said that Alnylam has upped the expected enrollment in this study to 25 patients from 15. Available data from the trial is slated for release in November, he noted, stressing that “any clinical endpoint result needs to be considered exploratory given the small sample size and the very limited duration of treatment of only six weeks” in the trial.
Vaishnaw added that an open-label extension (OLE) study for patients in the ALN-TTRsc study will kick off in the coming weeks, allowing the company to gather long-term dosing tolerability and clinical activity data on the drug.
Enrollment in an OLE study of patisiran has been completed with 27 patients, he said, and, “as of today, with up to nine months of therapy … there have been no study drug discontinuations.” Clinical endpoint data from approximately 20 patients in this study will be presented at the American Neurological Association meeting in October.
As part of its ATTR efforts, Alnylam has also been conducting natural history of disease studies in both FAP and FAC patients. Data from the 283-patient FAP study was presented earlier this year and showed a rapid progression in neuropathy impairment scores and a high correlation of this measurement with disease severity.
During last week’s conference call, Vaishnaw said that clinical endpoint and biomarker data on about 400 patients with either FAC or SSA have already been collected in a nature history study on cardiac ATTR. Maraganore said that these findings would likely be released sometime next year.
Alnylam Presents New Phase II, Preclinical Data from TTR Amyloidosis Programs
https://www.genomeweb.com/rnai/alnylam-presents-new-phase-ii-preclinical-data-ttr-amyloidosis-programs
Amyloid disease drug approved
Nature Biotechnology 2012; (3) http://dx.doi.org:/10.1038/nbt0212-121b
The first medication for a rare and often fatal protein misfolding disorder has been approved in Europe. On November 16, the E gave a green light to Pfizer’s Vyndaqel (tafamidis) for treating transthyretin amyloidosis in adult patients with stage 1 polyneuropathy symptoms. [Jeffery Kelly, La Jolla]
Safety and Efficacy of RNAi Therapy for Transthyretin …
http://www.nejm.org/…/NEJMoa1208760?…
The New England Journal of Medicine
Aug 29, 2013 – Transthyretin amyloidosis is caused by the deposition of hepatocyte-derived transthyretin amyloid in peripheral nerves and the heart.
Alnylam’s RNAi therapy targets amyloid disease
Ken Garber
Nature Biotechnology 2015; 33(577) http://dx.doi.org:/10.1038/nbt0615-577a
RNA interference’s silencing of target genes could result in potent therapeutics.
http://www.nature.com/nbt/journal/v33/n6/images/nbt0615-577a-I1.jpg
{kind=link}
The most clinically advanced RNA interference (RNAi) therapeutic achieved a milestone in April when Alnylam Pharmaceuticals in Cambridge, Massachusetts, reported positive results for patisiran, a small interfering RNA (siRNA) oligonucleotide targeting transthyretin for treating familial amyloidotic polyneuropathy (FAP). …
Nature Biotechnology 11 April 2016
Nature Biotechnology 02 March 2014
Nature Biotechnology 04 April 2016
Translational Neuroscience: Toward New Therapies
https://books.google.com/books?isbn=0262029863
Karoly Nikolich, Steven E. Hyman – 2015 – Medical
Tafamidis for Transthyretin Familial Amyloid Polyneuropathy: A Randomized, Controlled Trial. … Multiplex Genome Engineering Using CRISPR/Cas Systems.
Is CRISPR a Solution to Familial Amyloid Polyneuropathy?
Author and Curator: Larry H. Bernstein, MD, FCAP
Originally published as
http://scholar.aci.info/view/1492518a054469f0388/15411079e5a00014c3d
FAP is characterized by the systemic deposition of amyloidogenic variants of the transthyretin protein, especially in the peripheral nervous system, causing a progressive sensory and motor polyneuropathy.
FAP is caused by a mutation of the TTR gene, located on human chromosome 18q12.1-11.2.[5] A replacement of valine by methionine at position 30 (TTR V30M) is the mutation most commonly found in FAP.[1] The variant TTR is mostly produced by the liver.[citation needed] The transthyretin protein is a tetramer. ….
Schizophrenia, broken-links
Posted in Autism Spectrum Disorders, Cell Biology, Signaling & Cell Circuits, Cerebrovascular and Neurodegenerative Diseases, Chemical Biology and its relations to Metabolic Disease, Chemical Genetics, Child and Adolescent Psychiatry, Clinical & Translational, Cognition, Genetics & Innovations in Treatment, Genome Biology, MRI, Neurodegenerative Diseases, Neurohumoral Transmission, Neurological Diseases, Neuroscience, tagged C4A1, genomics, glia and neurons, microglia, neurogenesis, neuroimmaging, Neuron, Schizophrenia on April 5, 2016| Leave a Comment »
Schizophrenia, broken-links
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Runs in the Family
New findings about schizophrenia rekindle old questions about genes and identity.
BY SIDDHARTHA MUKHERJEE
Annals of Science MARCH 28, 2016 ISSUE http://www.newyorker.com/magazine/2016/03/28/the-genetics-of-schizophrenia
http://www.newyorker.com/wp-content/uploads/2016/03/160328_r27877-690.jpg
{kind=link}
The author and his father have seen several relatives succumb to mental illness.CREDIT PHOTOGRAPH BY DAYANITA SINGH FOR THE NEW YORKER
In the winter of 2012, I travelled from New Delhi, where I grew up, to Calcutta to visit my cousin Moni. My father accompanied me as a guide and companion, but he was a sullen and brooding presence, lost in a private anguish. He is the youngest of five brothers, and Moni is his firstborn nephew—the eldest brother’s son. Since 2004, Moni, now fifty-two, has been confined to an institution for the mentally ill (a “lunatic home,” as my father calls it), with a diagnosis of schizophrenia. He is kept awash in antipsychotics and sedatives, and an attendant watches, bathes, and feeds him through the day.
My father has never accepted Moni’s diagnosis. Over the years, he has waged a lonely campaign against the psychiatrists charged with his nephew’s care, hoping to convince them that their diagnosis was a colossal error, or that Moni’s broken psyche would somehow mend itself. He has visited the institution in Calcutta twice—once without warning, hoping to see a transformed Moni, living a secretly normal life behind the barred gates. But there was more than just avuncular love at stake for him in these visits. Moni is not the only member of the family with mental illness. Two of my father’s four brothers suffered from various unravellings of the mind. Madness has been among the Mukherjees for generations, and at least part of my father’s reluctance to accept Moni’s diagnosis lies in a grim suspicion that something of the illness may be buried, like toxic waste, in himself.
Rajesh, my father’s third-born brother, had once been the most promising of the Mukherjee boys—the nimblest, the most charismatic, the most admired. But in the summer of 1946, at the age of twenty-two, he began to behave oddly, as if a wire had been tripped in his brain. The most obvious change in his personality was a volatility: good news triggered uncontained outbursts of joy; bad news plunged him into inconsolable desolation. By that winter, the sine curve of Rajesh’s psyche had tightened in its frequency and gained in its amplitude. My father recalls an altered brother: fearful at times, reckless at others, descending and ascending steep slopes of mood, irritable one morning and overjoyed the next. When Rajesh received news of a successful performance on his college exams, he vanished, elated, on a two-night excursion, supposedly “exercising” at a wrestling camp. He was feverish and hallucinating when he returned, and died of pneumonia soon afterward. Only years later, in medical school, did I realize that Rajesh was likely in the throes of an acute manic phase. His mental breakdown was the result of a near-textbook case of bipolar disorder.
Jagu, the fourth-born of my father’s siblings, came to live with us in Delhi in 1975, when I was five years old and he was forty-five. His mind, too, was failing. Tall and rail thin, with a slightly feral look in his eyes and a shock of matted, overgrown hair, he resembled a Bengali Jim Morrison. Unlike Rajesh, whose illness had surfaced in his twenties, Jagu had been troubled from his adolescence. Socially awkward, withdrawn from everyone except my grandmother, he was unable to hold a job or live by himself. By 1975, he had visions, phantasms, and voices in his head that told him what to do. He was still capable of extraordinary bursts of tenderness—when I accidentally smashed a beloved Venetian vase at home, he hid me in his bedclothes and informed my mother that he had “mounds of cash” stashed away, enough to buy “a thousand” replacement vases. But this episode was symptomatic: even his love for me extended the fabric of his psychosis and confabulation.
Unlike Rajesh, Jagu was formally diagnosed. In the late nineteen-seventies, a physician in Delhi examined him and determined that he had schizophrenia. But no medicines were prescribed. Instead, Jagu continued to live at home, half hidden away in my grandmother’s room. (As in many families in India, my grandmother lived with us.) For nearly a decade, she and my father maintained a fragile truce, with Jagu living under her care, eating meals in her room and wearing clothes that she stitched for him. At night, when Jagu was consumed by his fears and fantasies, she put him to bed like a child, with her hand on his forehead. She was his nurse, his housekeeper, his only friend, and, more important, his public defender. When my grandmother died, in 1985, Jagu joined a religious sect in Delhi and disappeared, until his death, a dozen years later.
at schizophrenia runs in families was evident even to the person who first defined the illness. In 1911, Eugen Bleuler, a Swiss-German psychiatrist, published a book describing a series of cases of men and women, typically in their teens and early twenties, whose thoughts had begun to tangle and degenerate. “In this malady, the associations lose their continuity,” Bleuler wrote. “The threads between thoughts are torn.” Psychotic visions and paranoid thoughts flashed out of nowhere. Some patients “feel themselves weak, their spirit escapes, they will never survive the day. There is a growth in their heads. Their bones have turned liquid; their hearts have turned into stone. . . . The patient’s wife must not use eggs in cooking, otherwise he will grow feathers.” His patients were often trapped between flickering emotional states, unable to choose between two radically opposed visions, Bleuler noted. “You devil, you angel, you devil, you angel,” one woman said to her lover.
Bleuler tried to find an explanation for the mysterious symptoms, but there was only one seemingly common element: schizophrenic patients tended to have first-degree relatives who were also schizophrenic. He had no tools to understand the mechanism behind the heredity. The word “gene” had been coined just two years before Bleuler published his book. The notion that a mental illness could be carried across generations by unitary, indivisible factors—corpuscles of information threading through families—would have struck most of Bleuler’s contemporaries as mad in its own right. Still, Bleuler was astonishingly prescient about the complex nature of inheritance. “If one is looking for ‘theheredity,’ one can nearly always find it,” he wrote. “We will not be able to do anything about it even later on, unless the single factor of heredity can be broken down into many hereditary factors along specific lines.”
In the nineteen-sixties, Bleuler’s hunch was confirmed by twin studies. Psychiatrists determined that if an identical twin was schizophrenic the other twin had a forty-to-fifty-per-cent chance of developing the disease—fiftyfold higher than the risk in the general population. By the early two-thousands, large population studies had revealed a strong genetic link between schizophrenia and bipolar disorder. Some of the families described in these studies had a crisscrossing history that was achingly similar to my own: one sibling affected with schizophrenia, another with bipolar disorder, and a nephew or niece also schizophrenic.
“The twin studies clarified two important features of schizophrenia and bipolar disorder,” Jeffrey Lieberman, a Columbia University psychiatrist who has studied schizophrenia for thirty years, told me. “First, it was clear that there wasn’t a single gene, but dozens of genes involved in causing schizophrenia—each perhaps exerting a small effect. And, second, even if you inherited the entire set of risk genes, as identical twins do, you still might not develop the disease. Obviously, there were other triggers or instigators involved in releasing the illness.” But while these studies established that schizophrenia had a genetic basis, they revealed nothing about the nature of the genes involved. “For doctors, patients, and families in the schizophrenia community, genetics became the ultimate mystery,” Lieberman said. “If we knew the identity of the genes, we would find the causes, and if we found the causes we could find medicines.”
In 2006, an international consortium of psychiatric geneticists launched a genomic survey of schizophrenia, hoping to advance the search for the implicated genes. With 3,322 patients and 3,587 controls, this was one of the largest and most rigorous such studies in the history of the disease. Researchers scanned through the nearly seven thousand genomes to find variations in gene segments that were correlated with schizophrenia. This strategy, termed an “association study,” does not pinpoint a gene, but it provides a general location where a disease-linked gene may be found, like a treasure map with a large “X” scratched in a corner of the genome.
The results, reported in 2009 (and updated in 2014) in the journal Nature, were a dispiriting validation of Bleuler’s hunch about multiple hereditary factors: more than a hundred independent segments of the genome were associated with schizophrenia. “There are lots of small, common genetic effects, scattered across the genome,” one researcher said. “There are many different biological processes involved.” Some of the putative culprits made biological sense—if dimly. There were genes linked to transmitters that relay messages between neurons, and genes for molecular channels that move electrical signals up and down nerve cells. But by far the most surprising association involved a gene segment on chromosome 6. This region of the genome—termed the MHC region—carries hundreds of genes typically associated with the immune system.
“The MHC-segment finding was so strange and striking that you had to sit up and take notice,” Lieberman told me. “Here was the most definitive evidence that something in the immune system might have something to do with schizophrenia. There had been hints about an immunological association before, but this was impossible to argue with. It raised an endlessly fascinating question: what was the link between immune-response genes and schizophrenia?”
Beth Stevens thinks she has solved a mystery behind brain disorders such as Alzheimer’s and schizophrenia.
by Adam Piore April 4, 2016 https://www.technologyreview.com/s/601137/the-rogue-immune-cells-that-wreck-the-brain/
In the first years of her career in brain research, Beth Stevens thought of microglia with annoyance if she thought of them at all. When she gazed into a microscope and saw these ubiquitous cells with their spidery tentacles, she did what most neuroscientists had been doing for generations: she looked right past them and focused on the rest of the brain tissue, just as you might look through specks of dirt on a windshield.
“What are they doing there?” she thought. “They’re in the way.’”
Stevens never would have guessed that just a few years later, she would be running a laboratory at Harvard and Boston’s Children’s Hospital devoted to the study of these obscure little clumps. Or that she would be arguing in the world’s top scientific journals that microglia might hold the key to understanding not just normal brain development but also what causes Alzheimer’s, Huntington’s, autism, schizophrenia, and other intractable brain disorders.
Microglia are part of a larger class of cells—known collectively as glia—that carry out an array of functions in the brain, guiding its development and serving as its immune system by gobbling up diseased or damaged cells and carting away debris. Along with her frequent collaborator and mentor, Stanford biologist Ben Barres, and a growing cadre of other scientists, Stevens, 45, is showing that these long-overlooked cells are more than mere support workers for the neurons they surround. Her work has raised a provocative suggestion: that brain disorders could somehow be triggered by our own bodily defenses gone bad.

A type of glial cell known as an oligodendrocyte
In one groundbreaking paper, in January, Stevens and researchers at the Broad Institute of MIT and Harvard showed that aberrant microglia might play a role in schizophrenia—causing or at least contributing to the massive cell loss that can leave people with devastating cognitive defects. Crucially, the researchers pointed to a chemical pathway that might be targeted to slow or stop the disease. Last week, Stevens and other researchers published a similar finding for Alzheimer’s.
This might be just the beginning. Stevens is also exploring the connection between these tiny structures and other neurological diseases—work that earned her a $625,000 MacArthur Foundation “genius” grant last September.
All of this raises intriguing questions. Is it possible that many common brain disorders, despite their wide-ranging symptoms, are caused or at least worsened by the same culprit, a component of the immune system? If so, could many of these disorders be treated in a similar way—by stopping these rogue cells?
Schizophrenia is a heritable brain illness with unknown pathogenic mechanisms. Schizophrenia’s strongest genetic association at a population level involves variation in the major histocompatibility complex (MHC) locus, but the genes and molecular mechanisms accounting for this have been challenging to identify. Here we show that this association arises in part from many structurally diverse alleles of the complement component 4 (C4) genes. We found that these alleles generated widely varying levels of C4A and C4B expression in the brain, with each common C4 allele associating with schizophrenia in proportion to its tendency to generate greater expression of C4A. Human C4 protein localized to neuronal synapses, dendrites, axons, and cell bodies. In mice, C4 mediated synapse elimination during postnatal development. These results implicate excessive complement activity in the development of schizophrenia and may help explain the reduced numbers of synapses in the brains of individuals with schizophrenia.
Complex machinery
It’s not surprising that scientists for years have ignored microglia and other glial cells in favor of neurons. Neurons that fire together allow us to think, breathe, and move. We see, hear, and feel using neurons, and we form memories and associations when the connections between different neurons strengthen at the junctions between them, known as synapses. Many neuroscientists argue that neurons create our very consciousness.
Glia, on the other hand, have always been considered less important and interesting. They have pedestrian duties such as supplying nutrients and oxygen to neurons, as well as mopping up stray chemicals and carting away the garbage.
Scientists have known about glia for some time. In the 1800s, the pathologist Rudolf Virchow noted the presence of small round cells packing the spaces between neurons and named them “nervenkitt” or “neuroglia,” which can be translated as nerve putty or glue. One variety of these cells, known as astrocytes, was defined in 1893. And then in the 1920s, the Spanish scientist Pio del Río Hortega developed novel ways of staining cells taken from the brain. This led him to identify and name two more types of glial cells, including microglia, which are far smaller than the others and are characterized by their spidery shape and multiple branches. It is only when the brain is damaged in adulthood, he suggested, that microglia spring to life—rushing to the injury, where it was thought they helped clean up the area by eating damaged and dead cells. Astrocytes often appeared on the scene as well; it was thought that they created scar tissue.
This emergency convergence of microglia and astrocytes was dubbed “gliosis,” and by the time Ben Barres entered medical school in the late 1970s, it was well established as a hallmark of neurodegenerative diseases, infection, and a wide array of other medical conditions. But no one seemed to understand why it occurred. That intrigued Barres, then a neurologist in training, who saw it every time he looked under a microscope at neural tissue in distress. “It was just really fascinating,” he says. “The great mystery was: what is the point of this gliosis? Is it good? Is it bad? Is it driving the disease process, or is it trying to repair the injured brain?”
Barres began looking for the answer. He learned how to grow glial cells in a dish and apply a new recording technique to them. He could measure their electrical qualities, which determine the biochemical signaling that all brain cells use to communicate and coördinate activity.
“From the second I started recording the glial cells, I thought ‘Oh, my God!’” Barres recalls. The electrical activity was more dynamic and complex than anyone had thought. These strange electrical properties could be explained only if the glial cells were attuned to the conditions around them, and to the signals released from nearby neurons. Barres’s glial cells, in other words, had all the machinery necessary to engage in a complex dialogue with neurons, and presumably to respond to different kinds of conditions in the brain.
Why would they need this machinery, though, if they were simply involved in cleaning up dead cells? What could they possibly be doing? It turns out that in the absence of chemicals released by glia, the neurons committed the biochemical version of suicide. Barres also showed that the astrocytes appeared to play a crucial role in forming synapses, the microscopic connections between neurons that encode memory. In isolation, neurons were capable of forming the spiny appendages necessary to reach the synapses. But without astrocytes, they were incapable of connecting to one another.
Hardly anyone believed him. When he was a young faculty member at Stanford in the 1990s, one of his grant applications to the National Institutes of Health was rejected seven times. “Reviewers kept saying, ‘Nah, there’s no way glia could be doing this,’” Barres recalls. “And even after we published two papers in Science showing that [astrocytes] had profound, almost all-or-nothing effects in controlling synapses’ formation or synapse activity, I still couldn’t get funded! I think it’s still hard to get people to think about glia as doing anything active in the nervous system.”
Marked for elimination
Beth Stevens came to study glia by accident. After graduating from Northeastern University in 1993, she followed her future husband to Washington, D.C., where he had gotten work in the U.S. Senate. Stevens had been pre-med in college and hoped to work in a lab at the National Institutes of Health. But with no previous research experience, she was soundly rebuffed. So she took a job waiting tables at a Chili’s restaurant in nearby Rockville, Maryland, and showed up at NIH with her résumé every week.
After a few months, Stevens received a call from a researcher named Doug Fields, who needed help in his lab. Fields was studying the intricacies of the process in which neurons become insulated in a coating called myelin. That insulation is essential for the transmission of electrical impulses.
As Stevens spent the following years pursuing a PhD at the University of Maryland, she was intrigued by the role that glial cells played in insulating neurons. Along the way, she became familiar with other insights into glial cells that were beginning to emerge, especially from the lab of Ben Barres. Which is why, soon after completing her PhD in 2003, Stevens found herself a postdoc in Barres’s lab at Stanford, about to make a crucial discovery.
Barres’s group had begun to identify the specific compounds astrocytes secreted that seemed to cause neurons to grow synapses. And eventually, they noticed that these compounds also stimulated production of a protein called C1q.
Conventional wisdom held that C1q was activated only in sick cells—the protein marked them to be eaten up by immune cells—and only outside the brain. But Barres had found it in the brain. And it was in healthy neurons that were arguably at their most robust stage: in early development. What was the C1q protein doing there?

https://d267cvn3rvuq91.cloudfront.net/i/images/glia33.jpg?sw=590&cx=0&cy=0&cw=2106&ch=2106
{kind=link}
A stained astrocyte.
The answer lies in the fact that marking cells for elimination is not something that happens only in diseased brains; it is also essential for development. As brains develop, their neurons form far more synaptic connections than they will eventually need. Only the ones that are used are allowed to remain. This pruning allows for the most efficient flow of neural transmissions in the brain, removing noise that might muddy the signal.
But it was unknown how exactly the process worked. Was it possible that C1q helped signal the brain to prune unused synapses? Stevens focused her postdoctoral research on finding out. “We could have been completely wrong,” she recalls. “But we went for it.”
It paid off. In a 2007 paper, Barres and Stevens showed that C1q indeed plays a role in eliminating unneeded neurons in the developing brain. And they found that the protein is virtually absent in healthy adult neurons.
Now the scientists faced a new puzzle. Does C1q show up in brain diseases because the same mechanism involved in pruning a developing brain later goes awry? Indeed, evidence was already growing that one of the earliest events in neurodegenerative diseases such as Alzheimer’s, Parkinson’s, and Huntington’s was significant loss of synapses.
When Stevens and Barres examined mice bred to develop glaucoma, a neurodegenerative disease that kills neurons in the optic system, they found that C1q appeared long before any other detectable sign that the disease was taking hold. It cropped up even before the cells started dying.
This suggested the immune cells might in fact cause the disease, or at the very least accelerate it. And that offered an intriguing possibility: that something could be made to halt the process. Barres founded a company, Annexon Biosciences, to develop drugs that could block C1q. Last week’s paper published by Barres, Stevens, and other researchers shows that a compound being tested by Annexon appears to be able to prevent the onset of Alzheimer’s in mice bred to develop the disease. Now the company hopes to test it in humans in the next two years.
Paths to treatments
To better understand the process that C1q helps trigger, Stevens and Barres wanted to figure out what actually plays the role of Pac-Man, eating up the synapses marked for death. It was well known that white blood cells known as macrophages gobbled up diseased cells and foreign invaders in the rest of the body. But macrophages are not usually present in the brain. For their theory to work, there had to be some other mechanism. And further research has shown that the cells doing the eating even in healthy brains are those mysterious clusters of material that Beth Stevens, for years, had been gazing right past in the microscope—the microglia that Río Hortega identified almost 100 years ago.
Now Stevens’s lab at Harvard, which she opened in 2008, devotes half its efforts to figuring out what microglia are doing and what causes them to do it. These cells, it turns out, appear in the mouse embryo at day eight, before any other brain cell, which suggests they might help guide the rest of brain development—and could contribute to any number of neurodevelopmental diseases when they go wrong.
Meanwhile, she is also expanding her study of the way different substances determine what happens in the brain. C1q is actually just the first in a series of proteins that accumulate on synapses marked for elimination. Stevens has begun to uncover evidence that there is a wide array of protective “don’t eat me” molecules too. It’s the balance between all these cues that regulates whether microglia are summoned to destroy synapses. Problems in any one could, conceivably, mess up the system.
Evidence is now growing that microglia are involved in several neurodevelopmental and psychiatric problems. The potential link to schizophrenia that was revealed in January emerged after researchers at the Broad Institute, led by Steven McCarroll and a graduate student named Aswin Sekar, followed a trail of genetic clues that led them directly to Stevens’s work. In 2009, three consortia from around the globe had published papers comparing DNA in people with and without schizophrenia. It was Sekar who identified a possible pattern: the more a specific type of protein was present in synapses, the higher the risk of developing the disease. The protein, C4, was closely related to C1q, the one first identified in the brain by Stevens and Barres.
McCarroll knew that schizophrenia strikes in late adolescence and early adulthood, a time when brain circuits in the prefrontal cortex undergo extensive pruning. Others had found that areas of the prefrontal cortex are among those most ravaged by the disease, which leads to massive synapse loss. Could it be that over-pruning by rogue microglia is part of what causes schizophrenia?
To find out, Sekar and McCarroll got in touch with Stevens, and the two labs began to hold joint weekly meetings. They soon demonstrated that C4 also had a role in pruning synapses in the brains of young mice, suggesting that excessive levels of the protein could indeed lead to over-pruning—and to the thinning out of brain tissue that appears to occur as symptoms such as psychotic episodes grow worse.
If the brain damage seen in Parkinson’s and Alzheimer’s stems from over-pruning that might begin early in life, why don’t symptoms of those diseases show up until later? Barres thinks he knows. He notes that the brain can normally compensate for injury by rewiring itself and generating new synapses. It also contains a lot of redundancy. That would explain why patients with Parkinson’s disease don’t show discernible symptoms until they have lost 90 percent of the neurons that produce dopamine.
It also might mean that subtle symptoms could in fact be detected much earlier. Barres points to a study of nuns published in 2000. When researchers analyzed essays the nuns had written upon entering their convents decades before, they found that women who went on to develop Alzheimer’s had shown less “idea density” even in their 20s. “I think the implication of that is they could be lifelong diseases,” Barres says. “The disease process could be going on for decades and the brain is just compensating, rewiring, making new synapses.” At some point, the microglia are triggered to remove too many cells, Barres argues, and the symptoms of the disease begin to manifest fully.
Turning this insight into a treatment is far from straightforward, because much remains unclear. Perhaps an overly aggressive response from microglia is determined by some combination of genetic variants not shared by everyone. Stevens also notes that diseases like schizophrenia are not caused by one mutation; rather, a wide array of mutations with small effects cause problems when they act in concert. The genes that control the production of C4 and other immune-system proteins may be only part of the story. That may explain why not everyone who has a C4 mutation will go on to develop schizophrenia.
Nonetheless, if Barres and Stevens are right that the immune system is a common mechanism behind devastating brain disorders, that in itself is a fundamental breakthrough. Because we have not known the mechanisms that trigger such diseases, medical researchers have been able only to alleviate the symptoms rather than attack the causes. There are no drugs available to halt or even slow neurodegeneration in diseases like Alzheimer’s. Some drugs elevate neurotransmitters in ways that briefly make it easier for individuals with dementia to form new synaptic connections, but they don’t reduce the rate at which existing synapses are destroyed. Similarly, there are no treatments that tackle the causes of autism or schizophrenia. Even slowing the progress of these disorders would be a major advance. We might finally go after diseases that have run unchecked for generations.
“We’re a ways away from a cure,” Stevens says. “But we definitely have a path forward.”
Adam Piore is a freelance writer who wrote “A Shocking Way to Fix the Brain” in November/December 2015.
Int Immunopharmacol. 2001 Mar;1(3):365-92.
Genetic, structural and functional diversities of human complement components C4A and C4B and their mouse homologues, Slp and C4.
Blanchong CA1, Chung EK, Rupert KL, Yang Y, Yang Z, Zhou B, Moulds JM, Yu CY.
Abstract
The complement protein C4 is a non-enzymatic component of the C3 and C5 convertases and thus essential for the propagation of the classical complement pathway. The covalent binding of C4 to immunoglobulins and immune complexes (IC) also enhances the solubilization of immune aggregates, and the clearance of IC through complement receptor one (CR1) on erythrocytes. Human C4 is the most polymorphic protein of the complement system. In this review, we summarize the current concepts on the 1-2-3 loci model of C4A and C4B genes in the population, factors affecting the expression levels of C4 transcripts and proteins, and the structural, functional and serological diversities of the C4A and C4B proteins. The diversities and polymorphisms of the mouse homologues Slp and C4 proteins are described and contrasted with their human homologues. The human C4 genes are located in the MHC class III region on chromosome 6. Each human C4 gene consists of 41 exons coding for a 5.4-kb transcript. The long gene is 20.6 kb and the short gene is 14.2 kb. In the Caucasian population 55% of the MHC haplotypes have the 2-locus, C4A-C4B configurations and 45% have an unequal number of C4A and C4B genes. Moreover, three-quarters of C4 genes harbor the 6.4 kb endogenous retrovirus HERV-K(C4) in the intron 9 of the long genes. Duplication of a C4 gene always concurs with its adjacent genes RP, CYP21 and TNX, which together form a genetic unit termed an RCCX module. Monomodular, bimodular and trimodular RCCX structures with 1, 2 and 3 complement C4 genes have frequencies of 17%, 69% and 14%, respectively. Partial deficiencies of C4A and C4B, primarily due to the presence of monomodular haplotypes and homo-expression of C4A proteins from bimodular structures, have a combined frequency of 31.6%. Multiple structural isoforms of each C4A and C4B allotype exist in the circulation because of the imperfect and incomplete proteolytic processing of the precursor protein to form the beta-alpha-gamma structures. Immunofixation experiments of C4A and C4B demonstrate > 41 allotypes in the two classes of proteins. A compilation of polymorphic sites from limited C4 sequences revealed the presence of 24 polymophic residues, mostly clustered C-terminal to the thioester bond within the C4d region of the alpha-chain. The covalent binding affinities of the thioester carbonyl group of C4A and C4B appear to be modulated by four isotypic residues at positions 1101, 1102, 1105 and 1106. Site directed mutagenesis experiments revealed that D1106 is responsible for the effective binding of C4A to form amide bonds with immune aggregates or protein antigens, and H1106 of C4B catalyzes the transacylation of the thioester carbonyl group to form ester bonds with carbohydrate antigens. The expression of C4 is inducible or enhanced by gamma-interferon. The liver is the main organ that synthesizes and secretes C4A and C4B to the circulation but there are many extra-hepatic sites producing moderate quantities of C4 for local defense. The plasma protein levels of C4A and C4B are mainly determined by the corresponding gene dosage. However, C4B proteins encoded by monomodular short genes may have relatively higher concentrations than those from long C4A genes. The 5′ regulatory sequence of a C4 gene contains a Spl site, three E-boxes but no TATA box. The sequences beyond–1524 nt may be completely different as the C4 genes at RCCX module I have RPI-specific sequences, while those at Modules II, III and IV have TNXA-specific sequences. The remarkable genetic diversity of human C4A and C4B probably promotes the exchange of genetic information to create and maintain the quantitative and qualitative variations of C4A and C4B proteins in the population, as driven by the selection pressure against a great variety of microbes. An undesirable accompanying byproduct of this phenomenon is the inherent deleterious recombinations among the RCCX constituents leading to autoimmune and genetic disorders.
C4A isotype is responsible for effective binding to form amide bonds with immune aggregates or protein antigens, while C4B isotype catalyzes the transacylation of the thioester carbonyl group to form ester bonds with carbohydrate antigens.
Derived from proteolytic degradation of complement C4, C4a anaphylatoxin is a mediator of local inflammatory process.
Schizophrenia and the Synapse
Genetic evidence suggests that overactive synaptic pruning drives development of schizophrenia.
By Ruth Williams | January 27, 2016
http://www.the-scientist.com/?articles.view/articleNo/45189/title/Schizophrenia-and-the-Synapse/
Compared to the brains of healthy individuals, those of people with schizophrenia have higher expression of a gene called C4, according to a paper published inNature today (January 27). The gene encodes an immune protein that moonlights in the brain as an eradicator of unwanted neural connections (synapses). The findings, which suggest increased synaptic pruning is a feature of the disease, are a direct extension of genome-wide association studies (GWASs) that pointed to the major histocompatibility (MHC) locus as a key region associated with schizophrenia risk.
“The MHC [locus] is the first and the strongest genetic association for schizophrenia, but many people have said this finding is not useful,” said psychiatric geneticist Patrick Sullivan of the University of North Carolina School of Medicine who was not involved in the study. “The value of [the present study is] to show that not only is it useful, but it opens up new and extremely interesting ideas about the biology and therapeutics of schizophrenia.”
Schizophrenia has a strong genetic component—it runs in families—yet, because of the complex nature of the condition, no specific genes or mutations have been identified. The pathological processes driving the disease remain a mystery.
Researchers have turned to GWASs in the hope of finding specific genetic variations associated with schizophrenia, but even these have not provided clear candidates.
“There are some instances where genome-wide association will literally hit one base [in the DNA],” explained Sullivan. While a 2014 schizophrenia GWAS highlighted the MHC locus on chromosome 6 as a strong risk area, the association spanned hundreds of possible genes and did not reveal specific nucleotide changes. In short, any hope of pinpointing the MHC association was going to be “really challenging,” said geneticist Steve McCarroll of Harvard who led the new study.
Nevertheless, McCarroll and colleagues zeroed in on the particular region of the MHC with the highest GWAS score—the C4 gene—and set about examining how the area’s structural architecture varied in patients and healthy people.
The C4 gene can exist in multiple copies (from one to four) on each copy of chromosome 6, and has four different forms: C4A-short, C4B-short, C4A-long, and C4B-long. The researchers first examined the “structural alleles” of the C4 locus—that is, the combinations and copy numbers of the different C4 forms—in healthy individuals. They then examined how these structural alleles related to expression of both C4Aand C4B messenger RNAs (mRNAs) in postmortem brain tissues.
…………..
Schizophrenia risk from complex variation of complement component 4
Aswin Sekar, Allison R. Bialas, Heather de Rivera, …, Schizophrenia Working Group of the Psychiatric Genomics Consortium, Mark J. Daly, Michael C. Carroll, Beth Stevens & Steven A. McCarroll
Nature (11 Feb 2016); 530: 177–183 http://dx.doi.org:/10.1038/nature16549
Schizophrenia is a heritable brain illness with unknown pathogenic mechanisms. Schizophrenia’s strongest genetic association at a population level involves variation in the major histocompatibility complex (MHC) locus, but the genes and molecular mechanisms accounting for this have been challenging to identify. Here we show that this association arises in part from many structurally diverse alleles of the complement component 4 (C4) genes. We found that these alleles generated widely varying levels of C4A and C4B expression in the brain, with each common C4 allele associating with schizophrenia in proportion to its tendency to generate greater expression of C4A. Human C4 protein localized to neuronal synapses, dendrites, axons, and cell bodies. In mice, C4 mediated synapse elimination during postnatal development. These results implicate excessive complement activity in the development of schizophrenia and may help explain the reduced numbers of synapses in the brains of individuals with schizophrenia.
- Cannon, T. D. et al. Cortex mapping reveals regionally specific patterns of genetic and disease-specific gray-matter deficits in twins discordant for schizophrenia. Proc. Natl Acad. Sci. USA 99, 3228–3233 (2002)
- Cannon, T. D. et al. Progressive reduction in cortical thickness as psychosis develops: a multisite longitudinal neuroimaging study of youth at elevated clinical risk. Biol. Psychiatry 77,147–157 (2015)
- Garey, L. J. et al. Reduced dendritic spine density on cerebral cortical pyramidal neurons in schizophrenia. J. Neurol. Neurosurg. Psychiatry 65, 446–453 (1998)
- Glantz, L. A. & Lewis, D. A. Decreased dendritic spine density on prefrontal cortical pyramidal neurons in schizophrenia. Arch. Gen. Psychiatry 57, 65–73 (2000)
- Glausier, J. R. & Lewis, D. A. Dendritic spine pathology in schizophrenia. Neuroscience 251,90–107 (2013)
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature 511, 421–427 (2014)
- Shi, J. et al. Common variants on chromosome 6p22.1 are associated with schizophrenia. Nature 460, 753–757 (2009)
- Stefansson, H. et al. Common variants conferring risk of schizophrenia. Nature 460,744–747 (2009)
- International Schizophrenia Consortium et al. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 460, 748–752 (2009)
- Schizophrenia Psychiatric Genome-Wide Association Study Consortium. Genome-wide association study identifies five new schizophrenia loci. Nature Genet . 43, 969–976 (2011)
The strongest genetic association found in schizophrenia is its association to genetic markers across the major histocompatibility complex (MHC) locus, first described in three Nature papers in 2009. …
Schizophrenia: From genetics to physiology at last
Ryan S. Dhindsa & David B. Goldstein
Nature (11 Feb 2016); 530:162–163 http://dx.doi.org:/10.1038/nature16874
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. Nature511,421–427 (2014).
- Stevens, B. et al. Cell131, 1164–1178 (2007).
- Cannon, T. D. et al. Psychiatry77, 147–157 (2015).
- Glausier, J. R. & Lewis, D. A. Neuroscience251, 90–107 (2013).
- Glantz, L. A. & Lewis, D. A. Gen. Psychiatry57, 65–73 (2000).
Jianxin Shi1, et al. Common variants on chromosome 6p22.1 are associated with schizophrenia. Nature 460, 753-757 (6 August 2009) | doi:10.1038/nature08192; Received 29 May 2009; Accepted 10 June 2009; Published online 1 July 2009; Corrected 6 August 2009
Schizophrenia, a devastating psychiatric disorder, has a prevalence of 0.5–1%, with high heritability (80–85%) and complex transmission1. Recent studies implicate rare, large, high-penetrance copy number variants in some cases2, but the genes or biological mechanisms that underlie susceptibility are not known. Here we show that schizophrenia is significantly associated with single nucleotide polymorphisms (SNPs) in the extended major histocompatibility complex region on chromosome 6. We carried out a genome-wide association study of common SNPs in the Molecular Genetics of Schizophrenia (MGS) case-control sample, and then a meta-analysis of data from the MGS, International Schizophrenia Consortium and SGENE data sets. No MGS finding achieved genome-wide statistical significance. In the meta-analysis of European-ancestry subjects (8,008 cases, 19,077 controls), significant association with schizophrenia was observed in a region of linkage disequilibrium on chromosome 6p22.1 (P = 9.54 × 10-9). This region includes a histone gene cluster and several immunity-related genes—possibly implicating aetiological mechanisms involving chromatin modification, transcriptional regulation, autoimmunity and/or infection. These results demonstrate that common schizophrenia susceptibility alleles can be detected. The characterization of these signals will suggest important directions for research on susceptibility mechanisms.
Editor’s Summary 6 August 2009
Schizophrenia risk: link to chromosome 6p22.1
A genome-wide association study using the Molecular Genetics of Schizophrenia case-control data set, followed by a meta-analysis that included over 8,000 cases and 19,000 controls, revealed that while common genetic variation that underlies risk to schizophrenia can be identified, there probably are few or no single common loci with large effects. The common variants identified here lie on chromosome 6p22.1 in a region that includes a histone gene cluster and several genes implicated in immunity.
Letter
Hreinn Stefansson1,48, et al. Common variants conferring risk of schizophrenia.
Nature 460, 744-747 (6 August 2009) | doi:10.1038/nature08186; Received 16 March 2009; Accepted 5 June 2009; Published online 1 July 2009
Schizophrenia is a complex disorder, caused by both genetic and environmental factors and their interactions. Research on pathogenesis has traditionally focused on neurotransmitter systems in the brain, particularly those involving dopamine. Schizophrenia has been considered a separate disease for over a century, but in the absence of clear biological markers, diagnosis has historically been based on signs and symptoms. A fundamental message emerging from genome-wide association studies of copy number variations (CNVs) associated with the disease is that its genetic basis does not necessarily conform to classical nosological disease boundaries. Certain CNVs confer not only high relative risk of schizophrenia but also of other psychiatric disorders1, 2, 3. The structural variations associated with schizophrenia can involve several genes and the phenotypic syndromes, or the ‘genomic disorders’, have not yet been characterized4. Single nucleotide polymorphism (SNP)-based genome-wide association studies with the potential to implicate individual genes in complex diseases may reveal underlying biological pathways. Here we combined SNP data from several large genome-wide scans and followed up the most significant association signals. We found significant association with several markers spanning the major histocompatibility complex (MHC) region on chromosome 6p21.3-22.1, a marker located upstream of the neurogranin gene (NRGN) on 11q24.2 and a marker in intron four of transcription factor 4 (TCF4) on 18q21.2. Our findings implicating the MHC region are consistent with an immune component to schizophrenia risk, whereas the association with NRGN and TCF4 points to perturbation of pathways involved in brain development, memory and cognition.
Letter
The International Schizophrenia Consortium. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 460, 748-752 (6 August 2009) | doi:10.1038/nature08185; Received 11 February 2009; Accepted 8 June 2009; Published online 1 July 2009; Corrected 6 August 2009
Schizophrenia is a severe mental disorder with a lifetime risk of about 1%, characterized by hallucinations, delusions and cognitive deficits, with heritability estimated at up to 80%1, 2. We performed a genome-wide association study of 3,322 European individuals with schizophrenia and 3,587 controls. Here we show, using two analytic approaches, the extent to which common genetic variation underlies the risk of schizophrenia. First, we implicate the major histocompatibility complex. Second, we provide molecular genetic evidence for a substantial polygenic component to the risk of schizophrenia involving thousands of common alleles of very small effect. We show that this component also contributes to the risk of bipolar disorder, but not to several non-psychiatric diseases.
The Psychiatric GWAS Consortium Steering Committee. A framework for interpreting genome-wide association studies of psychiatric disorders. Molecular Psychiatry (2009) 14, 10–17; doi:10.1038/mp.2008.126; published online 11 November 2008
Genome-wide association studies (GWAS) have yielded a plethora of new findings in the past 3 years. By early 2009, GWAS on 47 samples of subjects with attention-deficit hyperactivity disorder, autism, bipolar disorder, major depressive disorder and schizophrenia will be completed. Taken together, these GWAS constitute the largest biological experiment ever conducted in psychiatry (59 000 independent cases and controls, 7700 family trios and >40 billion genotypes). We know that GWAS can work, and the question now is whether it will work for psychiatric disorders. In this review, we describe these studies, the Psychiatric GWAS Consortium for meta-analyses of these data, and provide a logical framework for interpretation of some of the conceivable outcomes.
Keywords: genome-wide association, attention-deficit hyperactivity disorder, autism, bipolar disorder, major depressive disorder, schizophrenia
The purpose of this article is to consider the ‘big picture’ and to provide a logical framework for the possible outcomes of these studies. This is not a review of GWAS per se as many excellent reviews of this technically and statistically intricate methodological approach are available.7, 8, 9, 10, 11, 12 This is also not a review of the advantages and disadvantages of different study designs and sampling strategies for the dissection of complex psychiatric traits. We would like to consider how the dozens of GWAS papers that will soon be in the literature can be synthesized: what can integrated mega-analyses (meta-analysis is based on summary data (for example, odds ratios) from all available studies whereas ‘mega-analysis’ uses individual-level genotype and phenotype data) of all available GWAS data tell us about the etiology of these psychiatric disorders? This is an exceptional opportunity as positive or negative results will enable us to learn hard facts about these critically important psychiatric disorders. We suggest that it is not a matter of ‘success versus failure’ or ‘optimism versus pessimism’ but rather an opportunity for systematic and logical approaches to empirical data whereby both positive and appropriately qualified negative findings are informative.
The studies that comprise the Psychiatric GWAS Consortium (PGC; http://pgc.unc.edu) are shown in Table 1. GWAS data for ADHD, autism, bipolar disorder, major depressive disorder and schizophrenia from 42 samples of European subjects should be available for mega-analyses by early 2009 (>59 000 independent cases and controls and >7700 family trios). To our knowledge, the PGC will have access to the largest set of GWAS data available.
A major change in human genetics in the past 5 years has been in the growth of controlled-access data repositories, and individual phenotype and genotype data are now available for many of the studies in Table 1. When the PGC mega-analyses are completed, most data will be available to researchers via the NIMH Human Genetics Initiative (http://nimhgenetics.org). Although the ready availability of GWAS data is a benefit to the field by allowing rapid application of a wide range of analytic strategies to GWAS data, there are potential disadvantages. GWAS mega-analysis is complex and requires considerable care and expertise to be done validly. For psychiatric phenotypes, there is the additional challenge of working with disease entities based largely on clinical description, with unknown biological validity and having both substantial clinical variation within diagnostic categories as well as overlaps across categories.13 Given the urgent need to know if there are replicable genotype–phenotype associations, a new type of collaboration was required.
The purpose of the PGC is to conduct rigorous and comprehensive within- and cross-disorder GWAS mega-analyses. The PGC began in early 2007 with the principal investigators of the four GAIN GWAS,14 and within six months had grown to 110 participating scientists from 54 institutions in 11 countries. The PGC has a coordinating committee, five disease-working groups, a cross-disorder group, a statistical analysis and computational group, and a cluster computer for statistical analysis. It is remarkable that almost all investigators approached agreed to participate and that no one has left the PGC. Most effort is donated but we have obtained funding from the NIMH, the Netherlands Scientific Organization, Hersenstichting Nederland and NARSAD.
The PGC has two major specific aims. (1) Within-disorder mega-analyses: conduct separate mega-analyses of all available GWAS data for ADHD, autism, bipolar disorder, major depressive disorder, and schizophrenia to attempt to identify genetic variation convincingly associated with any one of these five disorders. (2) Cross-disorder mega-analyses: the clinically-derived DSM-IV and ICD-10 definitions may not directly reflect the fundamental genetic architecture.15 There are two subaims. (2a) Conduct mega-analysis to identify genetic variation convincingly associated with conventional definitions of two or more disorders. This nosological aim could assist in delineating the boundaries of this set of disorders. (2b) An expert working group will convert epidemiological and genetic epidemiological evidence into explicit hypotheses about overlap among these disorders, and then conduct mega-analyses based on these definitions (for example, to examine the lifetime presence of idiopathic psychotic features without regard to diagnostic context).
The goal of the PGC is to identify convincing genetic variation-disease associations. A convincing association would be extremely unlikely to result from chance, show consistent effect sizes across all or almost all samples and be impervious to vigorous attempts to disprove the finding (for example, by investigating sources of bias, confirmatory genotyping, and so on). Careful attention will be paid to the impact of potential sources of heterogeneity17 with the goal of assessing its impact without minimizing its presence.
Biological plausibility is not an initial requirement for a convincing statistical association, as there are many examples in human genetics of previously unsuspected candidate genes nonetheless showing highly compelling associations. For example, multiple SNPs in intron 1 of the FTO gene were associated with body mass index in 13 cohorts with 38 759 participants18 and yet ‘FTO’ does not appear in an exhaustive 116 page compilation of genetic studies of obesity.19 Some strong associations are in gene deserts: multiple studies have found convincing association between prostate cancer and a region on 8q24 that is ~250 kb from the nearest annotated gene.20 Both of these examples are being intensively investigated and we suspect that a compelling mechanistic ‘story’ will emerge in the near future. The presence of a compelling association without an obvious biological mechanism establishes a priority research area for molecular biology and neuroscience of a psychiatric disorder.
The PGC will use mega-analysis as the main analytic tool as individual-level data will be available from almost all samples. To wield this tool appropriately, a number of preconditions must be met. First, genotype data from different GWAS platforms must be made comparable as the direct overlap between platforms is often modest. This requires meticulous quality control for the inclusion of both SNPs and subjects and attention to the factors that can cause bias (for example, population stratification, cryptic relatedness or genotyping batch effects). Genotype harmonization can be accomplished using imputation (21, 22, for example) so that the same set of ~2 million23, 24 directly or imputed SNP genotypes are available for all subjects. Second, phenotypes need to be harmonized across studies. This is one of the most crucial components of the PGC and we are fortunate to have world experts directing the work. Third, the mega-analyses will assess potential heterogeneity of associations across samples.
A decision-tree schematic of the potential outcomes of the PGC mega-analyses is shown in Figure 1. Note that many of the possibilities in Figure 1 are not mutually exclusive and different disorders may take different paths through this framework. It is possible that there eventually will be dozens or hundreds of sequence variants strictly associated with these disorders with frequencies ranging from very rare to common.
………
GWAS has the potential to yield considerable insights but it is no panacea and may well perform differently for psychiatric disorders. Even if these psychiatric GWAS efforts are successful, the outcomes will be complex. GWAS may help us learn that clinical syndromes are actually many different things—for example, proportions of individuals with schizophrenia might evidence associations with rare CNVs of major effect,5, 6 with more common genetic variation in dozens (perhaps hundreds) of genomic regions, between genetic variation strongly modified by environmental risk factors, and some proportion may be genetically indistinguishable from the general population. Moreover, as fuel to long-standing ‘lumper versus splitter’ debates in psychiatric nosology, empirical data might show that some clinical disorders or identifiable subsets of subjects might overlap considerably.
The critical advantage of GWAS is the search of a ‘closed’ hypothesis space. If the large amount of GWAS data being generated are analyzed within a strict and coherent framework, it should be possible to establish hard facts about the fundamental genetic architecture of a set of important psychiatric disorders—which might include positive evidence of what these disorders are or exclusionary evidence of what they are not. Whatever the results, these historically large efforts should yield hard facts about ADHD, autism, bipolar disorder, major depressive disorder and schizophrenia that may help guide the next era of psychiatric research.
- Pe’er I, Yelensky R, Altshuler D, Daly MJ. Estimation of the multiple testing burden for genomewide association studies of nearly all common variants. Genet Epidemiol 2008; 32: 381–385. | Article | PubMed |
- Weiss LA, Shen Y, Korn JM, Arking DE, Miller DT, Fossdal R et al. Association between microdeletion and microduplication at 16p11.2 and autism. N Engl J Med 2008; 358: 667–675. | Article | PubMed | ChemPort |
Letter
Hreinn Stefansson1,36, et al. Large recurrent microdeletions associated with schizophrenia. Nature 455, 232-236 (11 September 2008) | doi:10.1038/nature07229; Received 17 April 2008; Accepted 8 July 2008; Corrected 11 September 2008
Reduced fecundity, associated with severe mental disorders1, places negative selection pressure on risk alleles and may explain, in part, why common variants have not been found that confer risk of disorders such as autism2, schizophrenia3 and mental retardation4. Thus, rare variants may account for a larger fraction of the overall genetic risk than previously assumed. In contrast to rare single nucleotide mutations, rare copy number variations (CNVs) can be detected using genome-wide single nucleotide polymorphism arrays. This has led to the identification of CNVs associated with mental retardation4, 5 and autism2. In a genome-wide search for CNVs associating with schizophrenia, we used a population-based sample to identify de novoCNVs by analysing 9,878 transmissions from parents to offspring. The 66 de novo CNVs identified were tested for association in a sample of 1,433 schizophrenia cases and 33,250 controls. Three deletions at 1q21.1, 15q11.2 and 15q13.3 showing nominal association with schizophrenia in the first sample (phase I) were followed up in a second sample of 3,285 cases and 7,951 controls (phase II). All three deletions significantly associate with schizophrenia and related psychoses in the combined sample. The identification of these rare, recurrent risk variants, having occurred independently in multiple founders and being subject to negative selection, is important in itself. CNV analysis may also point the way to the identification of additional and more prevalent risk variants in genes and pathways involved in schizophrenia.
The C4 gene can exist in multiple copies (from one to four) on each copy of chromosome 6, and has four different forms: C4A-short, C4B-short, C4A-long, and C4B-long. The researchers first examined the “structural alleles” of the C4 locus—that is, the combinations and copy numbers of the different C4 forms—in healthy individuals. They then examined how these structural alleles related to expression of both C4Aand C4B messenger RNAs (mRNAs) in postmortem brain tissues.
From this the researchers had a clear picture of how the architecture of the C4 locus affected expression ofC4A and C4B. Next, they compared DNA from roughly 30,000 schizophrenia patients with that from 35,000 healthy controls, and a correlation emerged: the alleles most strongly associated with schizophrenia were also those that were associated with the highest C4A expression. Measuring C4A mRNA levels in the brains of 35 schizophrenia patients and 70 controls then revealed that, on average, C4A levels in the patients’ brains were 1.4-fold higher.
C4 is an immune system “complement” factor—a small secreted protein that assists immune cells in the targeting and removal of pathogens. The discovery of C4’s association to schizophrenia, said McCarroll, “would have seemed random and puzzling if it wasn’t for work . . . showing that other complement components regulate brain wiring.” Indeed, complement protein C3 locates at synapses that are going to be eliminated in the brain, explained McCarroll, “and C4 was known to interact with C3 . . . so we thought well, actually, this might make sense.”
McCarroll’s team went on to perform studies in mice that revealed C4 is necessary for C3 to be deposited at synapses. They also showed that the more copies of the C4 gene present in a mouse, the more the animal’s neurons were pruned.
Synaptic pruning is a normal part of development and is thought to reflect the process of learning, where the brain strengthens some connections and eradicates others. Interestingly, the brains of deceased schizophrenia patients exhibit reduced neuron density. The new results, therefore, “make a lot of sense,” said Cardiff University’s Andrew Pocklington who did not participate in the work. They also make sense “in terms of the time period when synaptic pruning is occurring, which sort of overlaps with the period of onset for schizophrenia: around adolescence and early adulthood,” he added.
“[C4] has not been on anybody’s radar for having anything to do with schizophrenia, and now it is and there’s a whole bunch of really neat stuff that could happen,” said Sullivan. For one, he suggested, “this molecule could be something that is amenable to therapeutics.”
UniProtKB
Derived from proteolytic degradation of complement C4, C4a anaphylatoxin is a mediator of local inflammatory process. It induces the contraction of smooth muscle, increases vascular permeability and causes histamine release from mast cells and basophilic leukocytes.
Non-enzymatic component of C3 and C5 convertases and thus essential for the propagation of the classical complement pathway. Covalently binds to immunoglobulins and immune complexes and enhances the solubilization of immune aggregates and the clearance of IC through CR1 on erythrocytes. C4A isotype is responsible for effective binding to form amide bonds with immune aggregates or protein antigens, while C4B isotype catalyzes the transacylation of the thioester carbonyl group to form ester bonds with carbohydrate antigens.
Related Articles in Pharmaceutical Intelligence
Identical Twin Brother Develops Schizophrenia
Outstanding Achievement in Schizophrenia Research
Combining Genetic has Identified Schizophrenia Subtypes
A new relationship identified in preterm stress and development of autism or schizophrenia
Protein binding to RNAs in brain
Dopamine-β-Hydroxylase Functional Variants
An inconvenient truth about dreams
Synapse activity in neurotransmission
Role of Neurotransmitters and other such Neurosignaling Molecules
microglia and brain maintenance
To reduce symptoms of mental illness and retrain the brain
To understand what happens in the brain to cause mental illness
Social Behavior Traits Embedded in Gene Expression
News in Exploration of the Biological Causes of Mental Illness: Potential for New Treatments
Translational Research on the Mechanism of Water and Electrolyte Movements into the Cell
Signaling Pathway that Makes Young Neurons Connect was discovered @ Scripps Research Institute
Modern Society, Risk and Mentally Disordered Offender
PrecisionFDA Consistency Challenge supports projects to validate and increase reproduceability of genomic testing methods
Posted in Clinical Genomics, tagged clinical genomics, genomics, next generation sequencing on March 15, 2016| Leave a Comment »
PrecisionFDA Consistency Challenge supports projects to validate and increase reproduceability of genomic testing methods
Reporter: Stephen J. Williams, Ph.D.
PrecisionFDA
Consistency Challenge
Engage and improve DNA test results with our first community challenge

President Obama’s Precision Medicine Initiative envisions a day when an individual’s medical care will be tailored in part based on their unique characteristics and genetic make-up.
The goal of the FDA’s first precisionFDA challenge is to engage the genomics community in advancing the quality standards in order to achieve more consistent results in the context of genetic tests (related to whole human genome sequencing), advancing the goal of better personalized care.
PrecisionFDA invites all innovators to take the challenge and assess their software on the supplied reference human datasets. Participation is voluntary, but instrumental in helping the community prepare for the coming genomic data revolution.
Challenge Time Period
February 25, 2016 through April 25, 2016
AT A GLANCE
In the context of whole human genome sequencing, software pipelines typically rely on mapping sequencing reads to a reference genome and subsequently identifying variants (differences). One way of assessing the performance of such pipelines is by using well-characterized datasets such as Genome in a Bottle’s NA12878.
By supplying NA12878 whole-genome sequencing read datasets (FASTQ), and a framework for comparing variant call format (VCF) results, this challenge provides a common frame of reference for measuring some of the aspects of reproducibility and accuracy of participants’ pipelines.

The challenge begins with two precisionFDA-provided input datasets, corresponding to whole-genome sequencing of the NA12878 human sample at two different sequencing sites. Your mission is to process these FASTQ files through your mapping and variation calling pipeline and create VCF files. For one of the datasets, you are required to do a rerun of your pipeline and obtain a rerun VCF as well. You can generate those results on your own environment, and upload them to precisionFDA, or you can reconstruct your pipeline on precisionFDA and run it on the cloud.
Regardless of how you generate your VCF files, you will subsequently use the precisionFDA comparison framework to conduct several pairwise comparisons:
- By comparing the rerun VCF to the original one, you will evaluate your pipeline’s reproducibility with respect to the same exact input file.
- By comparing the VCF files of the two datasets, you will evaluate reproducibility on the same sample across different sites.
- By comparing each of your three VCF files to the NIST (Genome in a Bottle) benchmark VCF, you will get estimates for accuracy.
The complete set of these five comparisons constitutes your submission entry to the challenge. Each comparison outputs several metrics (such as precision*, recall*, f-measure, or number of non-common variants). Selected participants and winners** will be recognized on the precisionFDA website. Therefore, we hope you are willing to share your experience with others to further enhance the community’s effort to ensure consistency of tests.
The challenge runs until April 25, 2016.
CHALLENGE DETAILS
Last updated: March 2nd, 2016
If you do not yet have a contributor account on precisionFDA, file an access request with your complete information, and indicate that you are entering the challenge. The FDA acts as steward to providing the precisionFDA service to the community and ensuring proper use of the resources, so your request will be initially pending. In the meantime, you will receive an email with a link to access the precisionFDA website in browse (guest) mode. Once approved, you will receive another email with your contributor account information.
With your contributor account you can use the features required to participate in the challenge (such as transfer files or run comparisons). Everything you do on precisionFDA is initially private to you (not accessible to the FDA or the rest of the community) until you choose to publicize it. So you can immediately start working on the challenge in private, and whenever you are ready you can officially publish your results as your challenge entry.
Footnotes
* The terminology currently used in the precisionFDA comparison output (such as “precision” and “recall”) is not necessarily harmonized with definitions used by ISO, CLSI, or FDA, but are terms commonly used by NGS software developers.
** Winning a precisionFDA challenge is an acknowledgement by the precisionFDA community and does not imply FDA endorsement of any organization, tool, software, etc.
BioMEMS The Market aspects of Oligonucleotide-Chips, Products, Applications and Competition, January 21, 2016
Posted in 3D Printing for Medical Application, Big Data, Bio Instrumentation in Experimental Life Sciences Research, BioIT: BioInformatics, BioIT: BioInformatics, NGS, Clinical & Translational, Pharmaceutical R&D Informatics, Clinical Genomics, Cancer Informatics, BioTechnology - Venture Creation, BioTechnology - Venture Creation, Venture Capital, CANCER BIOLOGY & Innovations in Cancer Therapy, Cancer Informatics, Cell Biology, Cell Biology, Signaling & Cell Circuits, Chemical Genetics, Commercialization, Computational Biology/Systems and Bioinformatics, CRISPR/Cas9 & Gene Editing, Digital HealthCare – biotech & internet joint ventures, DNA repair, Exosomes, FDA Regulatory Affairs, Frontiers in Cardiology and Cardiovascular Disorders, Genetics & Pharmaceutical, Genome Biology, Genomic Expression, Genomic Testing: Methodology for Diagnosis, Intelligent Information Systems, MEMS, Mutagenesis, Proteosome, tagged DNA Sequencing, gene expression, genetics, genome, genomics, Proteomics, qPCR & SNP Genotyping Custom Oligonucleotides - highly sophisticated oligonucleotides - simple PCR primers Oligos in Plates RNA FISH, RNA, THERMO SCIENTIFIC APPLIED BIOSYSTEMS INVITROGEN FISHER SCIENTIFIC UNITY LAB SERVICES on February 2, 2016| Leave a Comment »
BioMEMS The Market aspects of Oligonucleotide-Chips, Products and Applications, Competition, January 21, 2016
Curator: Gérard LOISEAU, ESQ
BioMEMS
The Market aspects of Oligonucleotide-Chips, Products, Applications, Competition
January 21, 2016
2015-2020
The oligonucleotide synthesis market is expected to reach USD 1.918.6Billion at a CAGR of 10.1% by 2020 from USD 1.078.1Billion in 2015.
SOURCE
MARKETSANDMARKETS marketsandmarkets.com/
PLAYERS
- Agilent Technologies Inc.
- BioAutomation Corp.
- Biosearch Technologies
- Gen9 Inc.
- GenScript Inc.
- Illumina Inc.
- Integrated DNA Technologies
- New England Biolabs Inc.
- Nitto Denko Avecia Inc.
- OriGene Technologies Inc.
- Sigma-Aldrich Corporation
- Thermo Fisher Scientific Inc.
- TriLink Biotechnologies
Agilent Technologies CA NYSE :A
- Agilent was created as a spin off from Hewlett-Packard Company in 1999.
- Agilent Technologies Inc. is engaged in the life sciences, diagnostics and applied chemical markets. The Company provides application focused solutions that include instruments, software, services and consumables for the entire laboratory workflow. The Company has three business segments:
the life sciences and applied markets business,
the diagnostics and genomics business, and
the Agilent Cross Lab business
- The Company’s life sciences and applied markets business segment brings together the Company’s analytical laboratory instrumentation and informatics.
- The Company’s diagnostics and genomics business segment consists of three businesses: the Dako business, the genomics business and the nucleic acid solutions business.
- The Company’s Agilent Cross Lab business segment combines its analytical laboratory services and consumables business
SOURCE
PRODUCTS AND SERVICES
https://www.agilent.com/en-us/default#collapse-0
- October 09, 2015 03:21 PM Eastern Daylight Time
- CARPINTERIA, Calif.–(BUSINESS WIRE)–Dako, an Agilent Technologies company and a worldwide provider of cancer diagnostics, today announced the U.S. Food and Drug Administration has approved a new test that can identify PD-L1 expression levels on the surface of non-small cell lung cancer tumor cells and provide information on the survival benefit with OPDIVO® (nivolumab) for patients with non-squamous NSCLC.
SOURCE
BUSINESS WIRE busibesswire.com/
BioAutomation Corp. TX
PRODUCTS AND SERVICES
- DNA and RNA synthesis reagents for the MerMades
Note: The MerMade 192E Oligonucleotide synthesizer is designed to synthesize DNA, RNA & LNA oligonucleotides in a column format
PARTNERSHIPS
- HONGENE BIOTECH : BIOAUTOMATION is the exclusive distributor for the Americas
- EMD MILLIPORE
- BIOSEARCH TECHNOLOGIES
DISTRIBUTORS
- LINK TECHNOLOGIES : UK
- AME BIOSCIENCE : UK
- BOSUNG SCIENCE : KOREA
- DNA CHEM : CHINA
- WAKO : JAPAN
- ACE PROBE : INDIA
SOURCE
Biosearch Technologies CA
PRODUCTS
- qPCR & SNP Genotyping
- Custom Oligonucleotides
- – highly sophisticated oligonucleotides
- – simple PCR primers
- Oligos in Plates
- RNA FISH
- Synthesis Reagents
- Immunochemicals
- Primers
- Probes
- Large-Scale Synthesis Oligos
- Intermediate-Scale Synthesis Oligos
SERVICES
- GMP & Commercial Services
- OEM & Kit Manufacturing
- qPCR Design Collaborations
DISTRIBUTORS
Argentina | Australia | Austria | Brazil | Canada |Chile | China | Colombia | Czech Republic | Denmark | Ecuador | Finland | Germany |Hong Kong | Israel | Italy | Japan | Korea | Malaysia | Mexico | New Zealand | Norway | Paraguay | Peru| Philippines | Poland | Romania | Singapore | South Africa | Spain | Sweden |Switzerland | Taiwan ROC | Thailand | Turkey | United Kingdom | Uruguay | Vietnam
SOURCE
biosearchtech.com/
Gen9 Inc. MA
PRODUCTS
Gen9 is building on advances in synthetic biology to power a scalable fabrication capability that will significantly increase the world’s capacity to produce DNA content. The privately held company’s next-generation gene synthesis technology allows for the high-throughput, automated production of DNA constructs at lower cost and higher accuracy than previous methods on the market. Founded by world leaders in synthetic biology, Gen9 aims to ensure the constructive application of synthetic biology in industries ranging from enzyme and chemical production to pharmaceuticals and biofuels.
SERVICES
- Synthetic Biology
- Gene Synthesis Services
- Variant Libraries
- Gene Sequence Design Services
INVESTORS
- Agilent Technologies : Private Equity
- CAMBRIDGE, Mass. and SANTA CLARA, Calif. — April 24, 2013 —Gen9 Receives $21 Million Strategic Investment from Agilent Technologies
SOURCE
gen9bio.com/
GenScript Inc. NJ
- GenScript is the largest gene synthesis provider in the USA
- GenScript Corporation, a biology contract research organization, provides biological research and drug discovery services to pharmaceutical companies, biotech firms, and research institutions in the United States, Europe, and Japan. It offers bio-reagent, custom molecular biology, custom peptide, protein production, custom antibody production, drug candidates testing, assay development and screening, lead optimization, antibody drug development, gene synthesis, and assay-ready cell line production services.
- The company also offers molecular biology, peptide, protein, immunoassay, chemicals, and cell biology products. It offers its products through distributors in Tokyo, Japan; and Seoul, Korea. GenScript Corporation has a strategic partnership with Immunologix, Inc. The company was founded in 2002 and is based in Piscataway, New Jersey. It has subsidiaries in France, Japan, and China.
Note: As of October 24, 2011, Immunologix, Inc. was acquired by Intrexon Corporation. Immunologix, Inc. develops and produces antibody-based therapeutics for various biological targets. It produces human monoclonal antibodies against viral, bacterial, and tumor antigens, as well as human auto antigens.
Intrexon Corporation, founded in 1998, is a leader in synthetic biology focused on collaborating with companies in Health, Food, Energy, Environment and Consumer sectors to create biologically based products that improve quality of life and the health of the planet.
PRODUCTS AND SERVICES
- Gene synthesis
- Antibody services
- Protein Services
- Peptide services
INVESTORS
Note: The Balloch Group (‘TBG’) was established in 2001 by Howard Balloch (Canada‘s ambassador to China from 1996 to 2001). TBG has since grown from a market-entry consultancy working with North American clients in China to a leading advisory and merchant banking firm serving both domestic Chinese companies and multinational corporations. TBG was ranked as the number one boutique investment bank in China by ChinaVenture in 2008.
Kleiner, Perkins, Caufield and Byers
Illumina Inc. CA
Monica Heger : SAN FRANCISCO (GenomeWeb) – Illumina today announced two new next-generation sequencing platforms, a targeted sequencing system called MiniSeq and a semiconductor sequencer that is still under development.
Illumina disclosed the initiatives during a presentation at the JP Morgan Healthcare conference held here today. During the presentation, Illumina CEO Jay Flatley also announced a new genotyping array called Infinium XT; a partnership with Bio-Rad to develop a single-cell sequencing workflow; preliminary estimates of its fourth-quarter 2015 revenues; and an update on existing products. The presentation followed the company’s announcement on Sunday that it has launched a new company called Grail to develop a next-generation sequencing test for early cancer detection from patient blood samples.
The MiniSeq system, which is based on Illumina’s current sequencing technology, will begin shipping early this quarter and has a list price of $49,500. It can perform a variety of targeted DNA and RNA applications, from single-gene to pathway sequencing, and promises “all-in” prices, including library prep and sequencing, of $200 to $300 per sample, Flatley said during the JP Morgan presentation.
SOURCES
http://investor.biospace.com/biospace/quote?Symbol=ILMN
PRODUCTS AND SERVICES
- Mid to large scale manufacturing assets
- Analytical Labs
- Pre-clinical
- Clinical
- Launched products
COMPETITORS
https://finance.yahoo.com/q/co?s=ILMN+Competitors Tue, Feb 2, 2016, 2:16pm EST – US Markets
| ILMN | PVT1 | AFFX | LMNX | Industry | |
|---|---|---|---|---|---|
| Market Cap: | 22.75B | N/A | 1.13B | 835.66M | 134.14M |
| Employees: | 3,700 | 10,000 | 1,200 | 745 | 45.00 |
| Qtrly Rev Growth (yoy): | 0.14 | N/A | -0.01 | 0.07 | 0.18 |
| Revenue (ttm): | 2.14B | 3.80B1 | 357.74M | 235.37M | 8.47M |
| Gross Margin (ttm): | 0.73 | N/A | 0.63 | 0.71 | 0.58 |
| EBITDA (ttm): | 770.84M | N/A | 46.64M | 52.99M | -12.31M |
| Operating Margin (ttm): | 0.30 | N/A | 0.08 | 0.17 | -1.62 |
| Net Income (ttm): | 510.36M | 430.90M1 | 11.22M | 39.29M | N/A |
| EPS (ttm): | 3.42 | N/A | 0.13 | 0.93 | -0.34 |
| P/E (ttm): | 45.43 | N/A | 104.40 | 20.91 | 25.33 |
| PEG (5 yr expected): | 2.68 | N/A | 4.66 | 0.55 | N/A |
| P/S (ttm): | 10.87 | N/A | 3.13 | 3.45 | 13.65 |
Pvt1 = Life Technologies Corporation (privately held)
AFFX = Affymetrix Inc.
LMNX = Luminex Corporation
Integrated DNA Technologies (IDT) IOWA + CA
Integrated DNA Technologies, Inc. (IDT), the global leader in nucleic acid synthesis, serving all areas of life sciences research and development, offers products for a broad range of genomics applications. IDT’s primary business is the production of custom, synthetic nucleic acids for molecular biology applications, including qPCR, sequencing, synthetic biology, and functional genomics. The company manufactures and ships an average of 44,000 custom nucleic acids per day to more than 82,000 customers worldwide. For more information, visit idtdna.com.
PRODUCTS AND SERVICES
- DNA & RNA Synthesis
- Custom DNA Oligos 96- & 384-Well Plates Ultramer Oligos Custom RNA Oligos SameDay Oligos HotPlates ReadyMade Primers Oligo Modifications Freedom
- Dyes GMP for Molecular Diagnostics Large Scale Oligo Synthesis
Note : Skokie, IL – December 1, 2015. Integrated DNA Technologies Inc. (“IDT”), the global leader in custom nucleic acid synthesis, has entered into a definitive agreement to acquire the oligonucleotide synthesis business of AITbiotech Pte. Ltd. in Singapore (“AITbiotech”). With this acquisition, IDT expands its customer base across Southeast Asia making it possible for these additional customers to now have access to its broad range of products for genomic applications. AITbiotech will continue operations in its other core business areas.
New England Biolabs Inc.
MA
PRODUCTS AND SERVICES
- Restriction Endonucleases
- PCR, Polymerases & Amplification Technologies
- DNA Modifying Enzymes
- Library Preparation for Next Generation Sequencing
- Nucleic Acid Purification
- Markers & Ladders
- RNA Reagents
- Gene Expression
- Cellular Analysis
SOURCE
Nitto Denko Avecia Inc. MA
With over 20 years of experience in oligonucleotide development and production, and over 1000 sequences manufactured, Avecia has played an integral role in the advancing oligo therapeutic market. Our mission is to continue to build value for our customers, as they progress through drug development into commercialization. And as a member of the Nitto Denko Corporation (nitto.com), Avecia is committed to the future of the oligonucleotide market. We are driven by innovative ideas and flexible solutions, designed to provide our customers with the best in service, quality, and technology.
SOURCE
Note : 1918 Nitto Electric Industrial Co., Ltd. forms in Ohsaki, Tokyo, to produce electrical insulating materials in Japan.
2011 Acquires Avecia Biotechnology Inc. in the U.S.A.
OriGene Technologies Inc. CA
OriGene Technologies, Inc. develops, manufactures, and sells genome wide research and diagnostic products for pharmaceutical, biotechnology, and academic research applications. The company offers cDNA clones, including TrueORF cDNA, viral ORF, destination vectors, TrueClones (human), TrueClones (mouse), organelle marker plasmids, MicroRNA tools, mutant and variant clones, plasmid purification kits, transfection reagents, and gene synthesis service; and HuSH shRNA, siRNA, miRNA, qPCR reagents, plasmid purification products, transfection reagents, PolyA+ and total RNA products, first-strand cDNA synthesis, and CRISPR/Cas9 genome products. It also provides proteins and lysates, such as purified human proteins, over-expression cell lysates, mass spectrometry standard proteins, and protein purification reagents; UltraMAB IHC antibodies, TrueMAB primary antibodies, anti-tag and fluorescent proteins, ELISA antibodies, luminex antibodies, secondary antibodies, and controls and others; and anatomic pathology products, including IHC antibodies, detection systems, and IHC accessories
The company offers luminex and ELISA antibody pairs, autoantibody profiling arrays, ELISA kits, cell assay kits, assay reagents, custom development, and fluorogenic cell assays; TissueFocus search tools; tissue sections; tissue microarrays, cancer protein lysate arrays, TissueScan cDNA arrays, tissue blocks, and quality control products, as well as tissue RNA, DNA, and protein lysates; and lab essentials. Its research areas include cancer biomarker research, RNAi, pathology IHC, stem cell research, ion channels, and protein kinase products. The company provides gene synthesis and molecular biology services, genome editing, custom cloning, custom shRNA, purified protein, monoclonal antibody development, and assay development. It sells its products through distributors worldwide, as well as online. OriGene Technologies, Inc. was incorporated in 1995 and is based in Rockville, Maryland.
SOURCE
PRODUCTS AND SERVICES
- cDNA Clones Human, mouse, rat Expression validated
- RNAi shRNA, siRNA microRNA & 3’UTR clones
- Gene Synthesis Codon optimization Variant libraries
- Real-time PCR Primer pairs, panels SYBR green reagents
- Lab Essentials DNA/RNA purification kits Transfection reagents
- Anatomic Pathology UltraMAB antibodies Specificity validated
- Recombinant Proteins 10,000 human proteins from mammalian system
- Antibodies TrueMAB primary antibodies Anti-tag antibodies
- Assays and Kits ELISA & Luminex antibodies Autoantibody Profiling Array
- Cancer & Normal Tissues Pathologist verified gDNA, RNA, sections, arrays
SOURCE
origene.com/
Sigma-Aldrich Corporation MI
Louis, MO – November 18, 2015 Merck KGaA, Darmstadt, Germany, Completes Sigma-Aldrich Acquisition
Merck KGaA today announced the completion of its $17 billion acquisition of Sigma-Aldrich, creating one of the leaders in the $130 billion global industry to help solve the toughest problems in life science.
Press Release: 18-Nov-2015
Letter to our Life Science Customers from Dr. Udit Batra
The life science business of Merck KGaA, Darmstadt, Germany brings together the world-class products and services, innovative capabilities and exceptional talent of EMD Millipore and Sigma-Aldrich to create a global leader in the life science industry.
“Everything we do starts with our shared purpose – to solve the toughest problems in life science by collaborating with the global scientific community.
This combination is built on complementary strengths, which will enable us to serve you even better as one organization than either company could alone.
This means providing a broader portfolio with a catalog of more than 300,000 products, including many of the most respected brands in the industry, greater geographic reach, and an unmatched combination of industry-leading capabilities.”
PRODUCTS AND SERVICES
http://www.sigmaaldrich.com/configurator/servlet/DesignCenter?btnOpen_0.x=1
http://www.sigmaaldrich.com/content/dam/sigma-aldrich/common/quality-products.jpg
{kind=link}
Thermo Fisher Scientific Inc. MA NYSE :TMO
Thermo Fisher Scientific Inc. is a provider of analytical instruments, equipment, reagents and consumables, software and services for research, manufacturing, analysis, discovery and diagnostics. The company operates through four segments: Life Sciences Solutions, provides reagents, instruments and consumables used in biological and medical research, discovery and production of new drugs and vaccines as well as diagnosis of disease; Analytical Instruments, provides instruments, consumables, software and services that are used in the laboratory; Specialty Diagnostics, offers diagnostic test kits, reagents, culture media, instruments and associated products, and Laboratory Products and Services, offers self-manufactured and sourced products for the laboratory.
SOURCE
PRODUCTS AND SERVICES
- Oligos Value – Standard – Plate
- Primers
- Probes
- Nucleotides
BRANDS
- THERMO SCIENTIFIC
- APPLIED BIOSYSTEMS
- INVITROGEN
- FISHER SCIENTIFIC
- UNITY LAB SERVICES
PARTNERSHIPS
AFFYMETRIX : NASDAQ : AFFX : affymetrix.com/
WALTHAM, Mass. & SANTA CLARA, Calif.–(BUSINESS WIRE)–Jan. 8, 2016– Thermo Fisher Scientific Inc. (NYSE:TMO), the world leader in serving science, and Affymetrix Inc. (NASDAQ:AFFX), a leading provider of cellular and genetic analysis products, today announced that their boards of directors have unanimously approved Thermo Fisher’s acquisition of Affymetrix for $14.00 per share in cash. The transaction represents a purchase price of approximately $1.3 billion.
SOURCE
TriLink Biotechnologies CA
PRODUCTS
Oligonucleotides
- DNA Oligos
- RNA Oligos
- Modified Oligos
- Specialty Oligos
Nucleotides
- NTPs (Nucleoside Triphosphates)
- Biphosphates
- Monophosphates
SERVICES
- Custom Chemistry
- Reagents
- Aptamers
PARTNERSHIPS
- LIFE TECHNOLOGIES,
- TERMO FISHER SCIENTIFIC since July 2015 thermofisher.com/
- GENMARK genmarkdx.com/
SOURCE
Other related articles published in this Open Access Online Scientific Journal include the following:
Gene Editing: The Role of Oligonucleotide Chips
http://pharmaceuticalintelligence.com/2016/01/07/gene-editing-the-role-of-oligonucleotide-chips/
Gene Editing for Exon 51: Why CRISPR Snipping might be better than Exon Skipping for DMD
The world’s most innovative intersection
Posted in BioIT: BioInformatics, NGS, Clinical & Translational, Pharmaceutical R&D Informatics, Clinical Genomics, Cancer Informatics, tagged gene biology, gene sequencing, genetics, genomics, Harvard, MIT on January 2, 2016| Leave a Comment »
The world’s most innovative intersection
Reported by: Irina Robu
Vassar Street and Main Street, in the new world’s Cambridge, Massachusetts, would be a leading candidate.
According to the article published in Wired Magazine in November 2015 “when the Whitehead got too small for genomicist Eric Lander’s ambitions, he launched a flashier and brasher newcomer next door. The Broad Institute’s gargantuan gleaming glass lobby is filled with early gene-sequencing instruments. Its multimedia screens boast that this is one of the world’s largest gene-sequencing and research factories. The Broad’s strategy is different from that of the Whitehead; instead of concentrating a few in an ultra-exclusive bioclub, Broad bridges MIT, Harvard and most of the hospitals in Boston. Its 2,000 members extend outwards, partnering with tens of thousands of others globally. Those working at the Broad are not averse to commerce; its director alone helped to build Foundation Medicine, Verastem, Millennium, Fidelity Biosciences, Courtagen and Aclara among many other leading companies.
The sixth building on this extraordinary corner, Novartis, focuses on private research, and represents a huge migration from Basel in Switzerland towards the MIT campus, becoming Cambridge’s largest employer. Pfizer, Sanofi, Amgen, Biogen-Idec and hundreds of others cluster nearby. “
Source
Roche is developing a high-throughput low cost sequencer for NGS
Posted in Bio Instrumentation in Experimental Life Sciences Research, BioIT: BioInformatics, BioIT: BioInformatics, NGS, Clinical & Translational, Pharmaceutical R&D Informatics, Clinical Genomics, Cancer Informatics, Clinical Diagnostics, Clinical Genomics, Next Generation Sequencing (NGS), tagged clinical genomics, deep sequencing, genomics, high throughput, highthroughput, next generation sequencing, NGS, Roche, Roche Diagnostics, Roche Institute of Molecular Biology on December 16, 2015| Leave a Comment »

Roche is developing a high-throughput low cost sequencer for NGS
Reporter: Stephen J. Williams, PhD
Reported from Diagnostic World News
Long-Read Sequencing in the Age of Genomic Medicine
By Aaron Krol
December 16, 2015 | This September, Pacific Biosciences announced the creation of the Sequel, a DNA sequencer half the cost and seven times as powerful as its previous RS II instrument. PacBio, with its unique long-read sequencing technology, had already secured a place in high-end research labs, producing finished, highly accurate genomes and helping to explore the genetic “dark matter” that other next-generation sequencing (NGS) instruments miss. Now, in partnership with Roche Diagnostics, PacBio is repositioning itself as a company that can serve hospitals as well.
“Pseudogenes, large structural variants, validation, repeat disorders, polymorphic regions of the genome―all those are categories where you practically need PacBio,” says Bobby Sebra, Director of Technology Development at the Icahn School of Medicine at Mount Sinai. “Those are gaps in the system right now for short-read NGS.”
Mount Sinai’s genetic testing lab owns three RS II sequencers, running almost around the clock, and was the first lab to announce it had bought a Sequel just weeks after the new instruments were launched. (It arrived earlier this month and has been successfully tested.) Sebra’s group uses these sequencers to read parts of the genome that, thanks to their structural complexity, can only be assembled from long, continuous DNA reads.
There are a surprising number of these blind spots in the human genome. “HLA is a huge one,” Sebra says, referring to a highly variable region of the genome involved in the immune system. “It impacts everything from immune response, to pharmacogenomics, to transplant medicine. It’s a pretty important and really hard-to-genotype locus.”
Nonetheless, few clinical organizations are studying PacBio or other long-read technologies. PacBio’s instruments, even the Sequel, come with a relatively high price tag, and research on their value in treating patients is still tentative. Mount Sinai’s confidence in the technology is surely at least partly due to the influence of Sebra―an employee of PacBio for five years before coming to New York―and Genetics Department Chair Eric Schadt, at one time PacBio’s Chief Scientific Officer.
Even here, the sequencers typically can’t be used to help treat patients, as the instruments are sold for research use only. Mount Sinai is still working on a limited number of tests to submit as diagnostics to New York State regulators.
Physician Use
Roche Diagnostics, which invested $75 million in the development of the Sequel, wants to change that. The company is planning to release its own, modified version of the instrument in the second half of 2016, specifically for diagnostic use. Roche will initially promote the device for clinical studies, and eventually seek FDA clearance to sell it for routine diagnosis of patients.
In an email to Diagnostics World, Paul Schaffer, Lifecycle Leader for Roche’s sequencing platforms division, wrote that the new device will feature an integrated software pipeline to interpret test results, in support of assays that Roche will design and validate for clinical indications. The instrument will also have at least minor hardware modifications, like near field communication designed to track Roche-branded reagents used during sequencing.
This new version of the Sequel will probably not be the first instrument clinical labs turn to when they decide to start running NGS. Short-read sequencers are sure to outcompete the Roche machine on price, and can offer a pretty useful range of assays, from co-diagnostics in cancer to carrier testing for rare genetic diseases. But Roche can clear away some of the biggest barriers to entry for hospitals that want to pursue long-read sequencing.
Today, institutions like Mount Sinai that use PacBio typically have to write a lot of their own software to interpret the data that comes off the machines. Off-the-shelf analysis, with readable diagnostic reports for doctors, will make it easier for hospitals with less research focus to get on board. To this end, Roche acquired Bina, an NGS analysis company that handles structural variants and other PacBio specialties, in late 2014.
The next question will be whether Roche can design a suite of tests that clinical labs will want to run. Long-read sequencing is beloved by researchers because it can capture nearly complete genomes, finding the correct order and orientation of DNA reads. “The long-read technologies like PacBio’s are going to be, in the future, the showcase that ties it all together,” Sebra says. “You need those long reads as scaffolds to bring it together.”
But that envisions a future in which doctors will want to sequence their patients’ entire genomes. When it comes to specific medical tests, targeting just a small part of the genome connected to disease, Roche will have to content itself with some niche applications where PacBio stands out.
Early Applications
“At this time we are not releasing details regarding the specific assays under development,” Schaffer told Diagnostics World in his email. “However, virology and genetics are a key focus, as they align with other high-priority Roche Diagnostics products.”
Genetic disease is the obvious place to go with any sequencing technology. Rare hereditary disorders are much easier to understand on a genetic level than conditions like diabetes or heart disease; typically, the pathology can be traced back to a single mutation, making it easy to interpret test results.
Some of these mutations are simply intractable for short-read sequencers. A whole class of diseases, the PolyQ disorders and other repeat disorders, develop when a patient has too many copies of a single, repetitive sequence in a gene region. The gene Huntingtin, for example, contains a long stretch of the DNA code CAG; people born with 40 or more CAG repeats in a row will develop Huntington’s disease as they reach early adulthood.
These disorders would be a prime target for Roche’s sequencer. The Sequel’s long reads, spanning thousands of DNA letters at a stretch, can capture the entire repeat region of Huntingtin at a stretch, unlike short-read sequencers that would tend to produce a garbled mess of CAG reads impossible to count or put in order.
Nonetheless, the length of reads is not the only obstacle to understanding these very obstinate diseases. “The entire category of PolyQ disorders, and Fragile X and Huntington’s, is really important,” says Sebra. “But to be frank, they’re the most challenging even with PacBio.” He suggests that, even without venturing into the darkest realms of the genome, a long-read sequencer might actually be useful for diagnosing many of the same genetic diseases routinely covered by other instruments.
That’s because, even when the gene region involved in a disease is well known, there’s rarely only one way for it to go awry. “An example of that is Gaucher’s disease, in a gene called GBA,” Sebra says. “In that gene, there are hundreds of known mutations, some of which you can absolutely genotype using short reads. But others, you would need to phase the entire block to really understand.” Long-read sequencing, which is better at distinguishing maternal from paternal DNA and highlighting complex rearrangements within a gene, can offer a more thorough look at diseases with many genetic permutations, especially when tracking inheritance through a family.
“You can think of long-read sequencing as a really nice way to supplement some of the inherited panels or carrier screening panels,” Sebra says. “You can also use PacBio to verify variants that are called with short-read sequencing.”
Virology is, perhaps, a more surprising focus for Roche. Diagnosing a viral (or bacterial, or fungal) infection with NGS only requires finding a DNA read unique to a particular species or strain, something short-read sequencers are perfectly capable of.
But Mount Sinai, which has used PacBio in pathogen surveillance projects, has seen advantages to getting the full, completely assembled genomes of the organisms it’s tracking. With bacteria, for instance, key genes that confer resistance to antibiotics might be found either in the native genome, or inside plasmids, small packets of DNA that different species of bacteria freely pass between each other. If your sequencer can assemble these plasmids in one piece, it’s easier to tell when there’s a risk of antibiotic resistance spreading through the hospital, jumping from one infectious species to another.
Viruses don’t share their genetic material so freely, but a similar logic can still apply to viral infections, even in a single person. “A virus is really a mixture of different quasi-species,” says Sebra, so a patient with HIV or influenza likely has a whole constellation of subtly different viruses circulating in their body. A test that assembles whole viral genomes—which, given their tiny size, PacBio can often do in a single read—could give physicians a more comprehensive view of what they’re dealing with, and highlight any quasi-species that affect the course of treatment or how the virus is likely to spread.
The Broader View
These applications are well suited to the diagnostic instrument Roche is building. A test panel for rare genetic diseases can offer clear-cut answers, pointing physicians to any specific variants linked to a disorder, and offering follow-up information on the evidence that backs up that call.
That kind of report fits well into the workflows of smaller hospital labs, and is relatively painless to submit to the FDA for approval. It doesn’t require geneticists to puzzle over ambiguous results. As Schaffer says of his company’s overall NGS efforts, “In the past two years, Roche has been actively engaged in more than 25 partnerships, collaborations and acquisitions with the goal of enabling us to achieve our vision of sample in to results out.”
But some of the biggest ways medicine could benefit from long-read sequencing will continue to require the personal touch of labs like Mount Sinai’s.
Take cancer, for example, a field in which complex gene fusions and genetic rearrangements have been studied for decades. Tumors contain multitudes of cells with unique patchworks of mutations, and while long-read sequencing can pick up structural variants that may play a role in prognosis and treatment, many of these variants are rarely seen, little documented, and hard to boil down into a physician-friendly answer.
An ideal way to unravel a unique cancer case would be to sequence the RNA molecules produced in the tumor, creating an atlas of the “transcriptome” that shows which genes are hyperactive, which are being silenced, and which have been fused together. “When you run something like IsoSeq on PacBio and you can see truly the whole transcriptome, you’re going to figure out all possible fusions, all possible splicing events, and the true atlas of reads,” says Sebra. “Cancer is so diverse that it’s important to do that on an individual level.”
Occasionally, looking at the whole transcriptome, and seeing how a mutation in one gene affects an entire network of related genes, can reveal an unexpected treatment option―repurposing a drug usually reserved for other cancer types. But that takes a level of attention and expertise that is hard to condense into a mass-market assay.
And, Sebra suggests, there’s another reason for medical centers not to lean too heavily on off-the-shelf tests from vendors like Roche.
Devoted as he is to his onetime employer, Sebra is also a fan of other technologies now emerging to capture some of the same long-range, structural information on the genome. “You’ve now got 10X Genomics, BioNano, and Oxford Nanopore,” he says. “Often, any two or even three of those technologies, when you merge them together, can get you a much more comprehensive story, sometimes faster and sometimes cheaper.” At Mount Sinai, for example, combining BioNano and PacBio data has produced a whole human genome much more comprehensive than either platform can achieve on its own.
The same is almost certainly true of complex cases like cancer. Yet, while companies like Roche might succeed in bringing NGS diagnostics to a much larger number of patients, they have few incentives to make their assays work with competing technologies the way a research-heavy institute like Mount Sinai does.
“It actually drives the commercialization of software packages against the ability to integrate the data,” Sebra says.
Still, he’s hopeful that the Sequel can lead the industry to pay more attention to long-read sequencing in the clinic. “The RS II does a great job of long-read sequencing, but the throughput for the Sequel is so much higher that you can start to achieve large genomes faster,” he says. “It makes it more accessible for people who don’t own the RS II to get going.” And while the need for highly specialized genetics labs won’t be falling off anytime soon, most patients don’t have the luxury of being treated in a hospital with the resources of Mount Sinai. NGS companies increasingly see physicians as some of their most important customers, and as our doctors start checking into the health of our genomes, it would be a shame if ubiquitous short-read sequencing left them with blind spots.
Source: http://diagnosticsworldnews.com/2015/12/16/long-read-sequencing-age-genomic-medicine.aspx
Brain Development
Posted in Behavioral Genetics, Biological Networks, Gene Regulation and Evolution, Curation, Developmental biology, Genome Biology, Neuroscience, tagged adult neurogenesis, bHLH, brain development, genomics, interneurons, olfaction, postnatal electroporation, Transcription factor on November 19, 2015| Leave a Comment »
Brain Development
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Updated 11/22/2015
Single Gene Found to Play Huge Role in Brain Development

Figure 1: Cells in which NeuroD1 is turned on are reprogrammed to become neurons. Cell nuclei are shown in blue (Höchst stain) and neurons are shown in red (stained with neuronal marker TUJ1). [A. Pataskar,J. Jung, V. Tiwari]
Researchers at the Institute of Molecular Biology (IMB) in Mainz, Germany say they have unraveled a complex regulatory mechanism that explains how a single gene can drive the formation of brain cells. Their study (“NeuroD1 reprograms chromatin and transcription factor landscapes to induce the neuronal program”), published in The EMBO Journal, is an important step toward a better understanding of how the brain develops. It also harbors potential for regenerative medicine, according to the scientists.
Neurodegenerative disorders, such as Parkinson’s disease, are often characterized by an irreversible loss neurons. Unlike many other cell types in the body, neurons are generally not able to regenerate by themselves, so if the brain is damaged, it stays damaged. One hope of developing treatments for this kind of damage is to understand how the brain develops in the first place, and then try to imitate the process. However, the brain is also one of the most complex organs in the body, and very little is understood about the molecular pathways that guide its development.

Figure 2: Diagram showing how NeuroD1 influences the development of neurons. During brain development, expression of NeuroD1 marks the onset of neurogenesis. NeuroD1 accomplishes this via epigenetic reprogramming: neuronal genes are switched on, and the cells develop into neurons. TF: transcription factor; V: ventricle; P: pial surface. [A. Pataskar, J. Jung, V. Tiwari]
ijay Tiwari, Ph.D, and his group have been investigating a central gene in brain development, NeuroD1. This gene is expressed in the developing brain and marks the onset of neurogenesis.
In their research article, Dr. Tiwari and his colleagues have shown that during brain development NeuroD1 is not only expressed in brain stem cells but acts as a master regulator of a large number of genes that cause these cells to develop into neurons. They used a combination of neurobiology, epigenetics, and computational biology approaches to show that these genes are normally turned off in development, but NeuroD1 activity changes their epigenetic state in order to turn them on. Strikingly, the researchers show that these genes remain switched on even after NeuroD1 is later switched off. They further show that this is because NeuroD1 activity leaves permanent epigenetic marks on these genes that keep them turned on, in other words it creates an epigenetic memory of neuronal differentiation in the cell.
“Our research has shown how a single factor, NeuroD1, has the capacity to change the epigenetic landscape of the cell, resulting in a gene expression program that directs the generation of neurons,” wrote the screenplay investigators.
“This is a significant step toward understanding the relationship between DNA sequence, epigenetic changes and cell fate. It not only sheds new light on the formation of the brain during embryonic development but also opens up novel avenues for regenerative therapy,” says Dr. Tiwari.
NEUROD1 neuronal differentiation 1 [ Homo sapiens (human) ]
Official Symbol NEUROD1 provided by HGNC
Official Full Name neuronal differentiation 1 provided by HGNC
Primary source HGNC:HGNC:7762 See related Ensembl:ENSG00000162992; HPRD:03428; MIM:601724; Vega:OTTHUMG00000132583
Gene type protein coding
RefSeq status REVIEWED
OrganismHomo sapiens
Lineage Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Euarchontoglires; Primates; Haplorrhini; Catarrhini; Hominidae; Homo
Also known asBETA2; BHF-1; MODY6; NEUROD; bHLHa3
Summary This gene encodes a member of the NeuroD family of basic helix-loop-helix (bHLH) transcription factors. The protein forms heterodimers with other bHLH proteins and activates transcription of genes that contain a specific DNA sequence known as the E-box. It regulates expression of the insulin gene, and mutations in this gene result in type II diabetes mellitus. [provided by RefSeq, Jul 2008]
https://en.wikipedia.org/wiki/NEUROD1
Neurogenic differentiation 1 (NeuroD1), also called β2,[1] is a transcription factor of the NeuroD-type. It is encoded by the human gene NEUROD1.
It is a member of the NeuroD family of basic helix-loop-helix (bHLH) transcription factors. The protein forms heterodimers with other bHLH proteins and activates transcription of genes that contain a specific DNA sequence known as the E-box. It regulates expression of the insulin gene, and mutations in this gene result in type II diabetes mellitus.[2]
Contents [hide]
NeuroD1 induces terminal neuronal differentiation in olfactory neurogenesis
Camille Boutin, Olaf Hardt, Antoine de Chevigny, Nathalie Coré, Sandra Goebbels, Ralph Seidenfaden, Andreas Bosio and Harold Cremer
PNAS Jan 19, 2010; 107(3): 1201–1206. http://dx.doi.org:/10.1073/pnas.0909015107
After their generation and specification in periventricular regions, neuronal precursors maintain an immature and migratory state until their arrival in the respective target structures. Only here are terminal differentiation and synaptic integration induced. Although the molecular control of neuronal specification has started to be elucidated, little is known about the factors that control the latest maturation steps. We aimed at identifying factors that induce terminal differentiation during postnatal and adult neurogenesis, thereby focusing on the generation of periglomerular interneurons in the olfactory bulb. We isolated neuronal precursors and mature neurons from the periglomerular neuron lineage and analyzed their gene expression by microarray. We found that expression of the bHLH transcription factor NeuroD1 strikingly coincides with terminal differentiation. Using brain electroporation, we show that overexpression of NeuroD1 in the periventricular region in vivo leads to the rapid appearance of cells with morphological and molecular characteristics of mature neurons in the subventricular zone and rostral migratory stream. Conversely, shRNA-induced knockdown of NeuroD1 inhibits terminal neuronal differentiation. Thus, expression of a single transcription factor is sufficient to induce neuronal differentiation of neural progenitors in regions that normally do not show addition of new neurons. These results suggest a considerable potential of NeuroD1 for use in cell-therapeutic approaches in the nervous system.
Determination of neuronal subtypes is an early event that coincides with cell cycle exit (1, 2). However, after their generation, new neurons have to remain immature for prolonged periods, allowing their migration to final destinations where terminal differentiation occurs (3). Little is known about the factors that maintain the precursor state or induce terminal differentiation.
Olfactory neurogenesis is particularly suited to approach these late steps in neuronal differentiation. Here, stem cell populations first located in the ventricular zone and after the establishment of an ependymal layer positioned in subventricular zone (SVZ) generate migratory neuroblasts throughout life (4). These perform long-distance chain migration via the rostral migratory stream (RMS) into the olfactory bulb (OB), where they migrate into the granule cell layer (GCL) and the glomerular layer (GL) to differentiate into GABA- and dopaminergic neurons (4, 5). Thus, in this system, generation of neurons is permanent and the consecutive steps in the neurogenic sequence are spatially separated.
Determination of newly generated neurons has been studied intensively over the past years. For example, it has been demonstrated that defined areas surrounding the lateral ventricle contain predetermined stem cells that give rise to defined subsets of interneurons (6, 7). Several transcription factors have been implicated in the specification of the different neuronal populations. The zinc finger transcription factor sp8, for instance, appears to be involved in the generation of interneurons expressing calretinin (8), and analysis of Sall3 mutant mice (9) points to a role of this factor in the dopaminergic, tyrosine hydroxylase–positive lineage (10). Furthermore, it appears that interneuron diversity relies on the combinatorial expression of such transcription factors. This is exemplified by Pax6 and Dlx2, which have been shown to interact in the determination of adult generated neuronal precursors toward a dopaminergic fate (9, 11, 12). All of these transcriptional regulators are expressed early during the neurogenic process and remain present until terminal differentiation occurs.
We aimed at the identification of transcription factors that induce terminal differentiation of postnatal generated neurons in the OB. To do so we isolated neuronal precursors and differentiated interneurons from the periglomerular lineage of the OB and compared their gene expression by microarray. We established that the expression of NeuroD1, a bHLH transcription factor that has been implicated in neuronal differentiation in several experimental systems (13–17), coincides with the passage from neuronal precursor to mature interneurons. Functionally, we show that premature expression of NeuroD1 in vitro and in vivo induced highly efficiently the differentiation of forebrain progenitors. In vivo, this leads to the transitory appearance of ectopic neurons in the SVZ, RMS and striatum. Conversely, knockdown of NeuroD1 specifically inhibits terminal maturation of periglomerular neurons in the OB. Thus, NeuroD1 is both necessary and sufficient to induce key steps in terminal neuronal differentiation.
NeuroD1 Is Specifically Expressed in Mature GL Interneurons.
Subpopulations of neuronal precursors destined for the GCL and GL of the OB are generated by regionally defined stem cell populations in the periventricular region but migrate intermingled in the RMS to the OB. Once there, cells resegregate: granule cell precursors terminate their migration in the GCL, whereas the smaller population of periglomerular neuron precursors traverses this layer and the mitral cell layer (MCL) to invade the peripherally located GL (Fig. 1A). Thus, at a given time point, the GL contains both mature periglomerular neurons and their specific progenitors. Based on this spatial organization we isolated these two populations, concurrently depleting glial cells.
We devised a three-step strategy based on the following: (i) microdissection followed by enzymatic dissociation of the postnatal GL, (ii) depletion of contaminating glial cells by magnetic activated cell sorting (MACS) using an A2B5 specific antibody (18), (iii) separation of PSA-NCAM expressing cells (19) from the remaining fraction containing the mature neurons (Fig. 1B). The same purification strategy was applied to tissue microdissected from the P2 periventricular region (18). Characterization of the different cell population after sorting was performed via immunocytochemistry using the markers used for sorting (A2B5 and PSA-NCAM) as well as the differentiation marker Gad65 (18) (Fig. S1). Thus, as starting material we obtained highly enriched mature OB periglomerular interneurons (PGN), their immature progenitors (PGP), as well as a mixed population of generic progenitors (GP) from the SVZ/RMS.

Fig. 1.
Expression of NeuroD1 in the olfactory neurogenic system (A) DAPI-stained coronal section through the olfactory bulb of P5. (B) Strategy to isolate neuronal populations at different steps of their maturation. (C) Relative changes in gene expression for selected genes. Expression in GP was considered baseline, and changes are expressed as fold difference. (D–F) NeuroD1 in situ hybridization on sections from P5 mouse brain. No signal was detected along the lateral ventricle or in the RMS (D). In the olfactory bulb, individual NeuroD1+ cells were present in the GCL, whereas the MCL and the GL contained higher amounts (E, high magnification in F). A similar expression pattern was found after β-gal reaction on NeuroD1-lacZ-knockin tissue (G). (Scale bar: 200 μm in A; 100 μm inD and E; 20 μm in F and G).
Based on the purified and characterized cell populations, we performed microarray analyses to gain insight into the changes in gene expression during the neurogenic process. Investigation of expression dynamics of genes associated with either the precursor status or neuronal differentiation (Fig. S2 A and B) were used to validate the approach. Furthermore, these data were compared with those from an already available Serial Analysis of Gene Expression (SAGE) study (20).
Serial Analysis of Microarray (SAM) demonstrated the presence of groups of genes with comparable expression patterns (Fig. S2 C–E). Interestingly, only a relatively small fraction of genes were absent in the immature cell populations GP and PGP but highly represented in mature PGN (Fig. S2E). One of the genes showing such a pattern was NeuroD1, which was expressed more than 50-fold higher in PGN than in the immature populations (Fig. 1C). This was in agreement with the above-cited SAGE data, showing that NeuroD1 expression was below the detection level in neuronal precursors of the adult SVZ (20). Thus, expression of NeuroD1 was absent from precursors but coincided with terminal neuronal differentiation.
This late expression of NeuroD1 was in contrast to that of factors that have been functionally implicated in the specification of PGN, including Pax6, Sp8 and Sall3, which were expressed in both the immature populations and in the mature neurons (Fig. 1C; in situ hybridization for Pax6 in Fig. S3). Only Dlx2 showed a moderate increase in the PGN lineage outgoing, however, from an already considerable baseline level in migrating precursors (12) (Fig. 1C).
Next we analyzed the expression of NeuroD1 using in situ hybridization on P5 forebrain sections. Strong expression was found in the GL, whereas weaker expression was observed in the GCL and MCL (Fig. 1 Eand F). The transcript was undetectable in the periventricular region and the RMS (Fig. 1D). This staining was confirmed using NeuroD1-lacZ knockin mice (21) (Fig. 1G). In conclusion, these data demonstrated the absence of NeuroD1 from immature cells of the system and its strong expression in mature PGN. This pattern was coherent with a function in terminal neuronal differentiation.
NeuroD1 Induces Neuronal Differentiation in Vitro.
We studied the neurogenic potential of NeuroD1 in primary cultured neural stem cells using the neurosphere assay. In parallel to NeuroD1, we performed all experiments under the same conditions using the transcription factor Pax6, a well-described neurogenic signal in the system (9, 11, 12), to control for specificity of the observed effects. Neurosphere cells were coelectroporated with NeuroD1 or Pax6 expression vectors and GFP immediately before plating in differentiation conditions. One week after transfection, in the control condition, 14 ± 1% of the GFP-positive cells coexpressed the early neuronal marker Tuj1 (Fig. S4 A and D) whereas NeuroD1 induced Tuj1 expression in virtually all cells (98.0 ± 2%, Fig. S4 B and D). Pax6 gain-of-function led to an intermediate value (60.0 ± 3%, Fig. S4 C and D). NeuN, a later neuronal marker (22), was expressed by 21.1 ± 1% of the Tuj1-positive cells in the control situation (Fig. S4 E and H) but induced by NeuroD1 in almost all cells (93.9 ± 2%; Fig. S4 F and H). Surprisingly, Pax6 expression led to nearly complete disappearance of NeuN (1.7 ± 0.3%; Fig. S4 G and H). We investigated the induction of subtype specific markers by NeuroD1. Whereas tyrosine hydroxylase showed no augmentation, we found a 20% increase in calretinin labeling, in agreement with previous findings (23).
Next we investigated morphological parameters like process length as well as density and length of filopodia. Both NeuroD1 and Pax6 induced a significant, greater than 2-fold increase in process length (Fig. S4 I and L). We analyzed dendritic filopodia, structures that are believed to be precursors of dendritic spines (24). Expression of NeuroD1 induced a doubling in density and length of filopodia (Fig. S4 N, P, and Q). Interestingly, Pax6 reduced filopodia density to a level significantly below that of controls (Fig. S4 O and P), whereas length of the few remaining filopodia was not affected (7.0 ± 0.4 μm; Fig. S4Q).
Thus, the expression of NeuroD1 in neurosphere amplified neural stem cells induced neuronal commitment as well as morphological characteristics of mature neurons. Like NeuroD1, Pax6 favored neuronal commitment but appeared to actively suppress certain characteristics of terminal neuronal differentiation.
NeuroD1 Induces Ectopic Neurons in Vivo.
We asked whether NeuroD1 was also sufficient to induce neuronal differentiation in vivo. We used postnatal forebrain electroporation, an approach that allows efficient genetic manipulation of neural stem cells along the lateral ventricles and, consequently, of all transitory or permanent cell populations that are generated in the olfactory neurogenic process (25). The NeuroD1 expression vector or empty control plasmids were coelectroporated together with a GFP-containing vector that allowed visualization of transfected cells and their progeny at high resolution. Consequences of NeuroD1 gain-of-function were analyzed at 2, 4, 6, 8, and 15 days postelectroporation (dpe). As for the in vitro studies, results were compared with the effects of Pax6 gain-of-function.
At 2 dpe of a control vector into the lateral wall of the forebrain ventricle, 9.8 ± 1.3% (Fig. 2 A and K) of the GFP-expressing cells were localized in the VZ and had the morphology of radial glia (RG) (25). The majority of the GFP + cells, representing mainly neuronal precursors, were localized in the SVZ. Electroporation of a NeuroD1 expression vector induced a loss of GFP-positive RG cells (3.7 ± 0.5%; Fig. 2 B and K). The remaining cells in the VZ showed lower GFP levels than in controls (Fig. 2 A and B asterisks).

Fig. 2.
NeuroD1 induces neuronal morphology in vivo. Effect of NeuroD1 gain-of-function at different time points postelectroporation. (A and B) Coronal forebrain sections at the level of the lateral ventricle at 2 dpe. In the control condition, strongly GFP labeled RG are present in the VZ (A, asterisk). Expression of NeuroD1 induced a relative loss of radial glia and fainter GFP label (B, asterisk). (C and D) Coronal sections at the level of the lateral ventricle at 4 dpe. NeuroD1 expression induced an accumulation of transfected cells in the SVZ (D) and the almost total disappearance of radial glia (D). (E–F′) Sagittal sections of the RMS at 4 dpe. In the control situation, cells migrated toward the OB and presented the bipolar morphology specific of migrating precursors (E, E′, arrowheads). NeuroD1 electroporation induced loss of tangential orientation, induction of complex branching (F, F′, arrowhead), and invasion of the surrounding tissues (F, arrowheads). (G and H) Coronal section at the level of the olfactory bulb at 4 dpe. Although the majority of cells have reached the OB in the control situation (G), only a few cells were located in the center of the OB in the presence of NeuroD1 (H). (I and I′) Examples of cells presenting neuronal morphology in the SVZ at 4 dpe. (J) High magnification showing the presence of filopodia covering NeuroD1-expressing cells (arrowheads). (K) Quantification of GFP-positive cells presenting radial glia cell morphology along the lateral ventricle at 2 and 4 dpe. Control: 9.8 ± 1.3% (n= 6) at 2 dpe; 24 ± 11.8% at 4 dpe (n = 3); NeuroD1: 3.7 ± 0.5% at 2 dpe (n = 6); 1.6 ± 0.7% at 4 dpe (n = 3). (l) Distribution of the GFP-positive cells along the rostrocaudal axis. NeuroD1 expressing cells accumulated in proximal parts of the system. (M) Morphological analysis of cells in the SVZ/RMS. Three different classes were defined: (i) bipolar cells presenting tangential orientation, (ii) spherical cells, and (iii) branched cells presenting multiple processes in various directions (compare I). NeuroD1-expressing cells presented a highly branched morphology. Control: bipolar, 80.4%; spherical, 19.5%; branched, 0% (n = 133 cells). NeuroD1: bipolar, 5%; spherical, 16.8%; branched, 78% (n = 119 cells). Statistics: Mann-Whitney test. ns, not significant. **P < 0.01; ***P < 0.005. (Scale bar: 100 μm in E, F, G,and H; 25 μm in A, B, C, D,E, and F’; 10 μm in I; 5 μm in J.)
At 4 dpe, in the control situation, considerable amounts of strongly GFP+ RG cells were still present in the VZ (Fig. 2C asterisks), whereas NeuroD1 expression induced an almost complete loss of RG cells (Fig. 2 Dand K). At this time point, control cells were found along the entire SVZ and RMS. They showed generally tangential orientation and the typical morphology of migratory neuronal precursors. Large amounts of such cells were also found in the center of the OB (Fig. 2 G and L). NeuroD1 expression induced an accumulation of GFP-labeled cells in the SVZ (Fig. 2 D and L) at the expense of cells in the RMS (Fig. 2H,quantified in Fig. 2L). The accumulating cells did not have the appearance of migrating precursors but displayed complex multibranched morphologies (Fig. 2 F and F′, examples in Fig. 2 I and I′, quantified inFig. 2M). All principal processes of these cells were covered with small protrusions resembling filopodia (Fig. 2J). Such morphologically complex cells, strongly resembling neurons, were also predominant in and along proximal parts of the RMS (Fig. 2F′). Interestingly, considerable amounts of multibranched cells were found outside of the periventricular region and the RMS, invading neighboring structures such as the striatum (Fig. 2F, arrows). There was a clear correlation between the quantity of transgene expression, as visualized by GFP fluorescence, and the above parameters. Thus, NeuroD1 induced dose-dependently a neuron-like morphology in cells in the SVZ, RMS, and surrounding tissues.
We characterized the NeuroD1 induced neuron-like cell population in the periventricular region using neuronal and glial markers (Fig. 3; examples in Fig. S5). Doublecortin (DCX), a microtubule-associated protein expressed in migratory neuronal precursors (26), was seen in 75.2 ± 4.5% of the cells in the control situation but showed a significant increase after expression of NeuroD1 (91.7 ± 2.2%). NeuN, a marker for most mature neuronal cell types in the brain (22) was low in controls (5.2 ± 1.4%, n = 8) but strongly induced by NeuroD1 (65.9 ± 4.5%, n = 9). Map2, a later generic neuronal marker (27), was also rare in control cells (14.1 ± 1.4%, n = 3) but highly expressed in the NeuroD1 condition (61.9 ± 2.7%, n = 3). GFAP and Olig2 did not show significant alterations due to NeuroD1 expression. Thus, the NeuroD1-induced ectopic cells with neuronal morphology in the SVZ and RMS showed molecular characteristics of neurons.

Fig. 3.
NeuroD1 induces generic neuronal markers in vivo Molecular phenotype of the cells located in the periventricular region (level 4 in Fig. 2l). Quantification representing the percentage of GFP-positive cells expressing the respective markers. DCX: control, 75.2 ± 4.5%, n = 5; NeuroD1, 91.7 ± 2.2%, n = 5. NeuN: control, 5.2 ± 1.4%, n = 8; NeuroD1, 65.9 ± 4.5%, n = 9. Map2: control, 14.1 ± 1.4%, n = 3; NeuroD1, 61.9 ± 2.7%, n = 3. Olig2: control, 6.8 ± 5%, n = 3; NeuroD1, 2.5 ± 0.5%, n = 3. GFAP: control, 0%, n = 3; NeuroD1, 0%, n = 2. Errors bars indicate SEM. Statistics: DCX and Map2, unpaired ttest; NeuN, Mann-Whitney test. ns, not significant. *P < 0.05; **P < 0.01; ***P < 0.005.
HighWire Press-hosted articles citing this article
-
……..
NeuroD1 Is Necessary For OB Interneuron Differentiation in Vivo.
Next we asked whether NeuroD1 is essential for the generation of PGN. Given that NeuroD1 deficiency in mice is generally associated with perinatal lethality (14, 15, 21), we used a strategy based on RNAi in concert with postnatal in vivo electroporation to knock down NeuroD1 in the olfactory bulb neurogenic system. For validation, three different NeuroD1 specific shRNA vectors were cotransfected with a NeuroD1 expression construct into COS-7 cells. Western blot analysis demonstrated that two of the shRNAs, sh775 and sh776, efficiently inhibited production of the NeuroD1 protein, whereas sh777 induced a less efficient downregulation (sh775, 94.6%; sh776, 96.9%; sh777, 78.4%; corrected for loading against αtubulin; Fig. 4A). All three shRNAs were used for further in vivo studies.

Fig. 4.
In vivo terminal neuronal differentiation of PGC is impaired in absence of NeuroD1. (A) Western blot analysis of protein extracts from cos-7 cells transfected with NeuroD1 or in combination with different NeuroD1 specific shRNAs. sh775 and sh776 strongly repressed NeuroD1 protein expression (94.6% and 96.9%, respectively), whereas sh777 repressed NeuroD1 by 74.8%. (B–H′′) Consequences of loss-of-function of NeuroD1 via in vivo postnatal electroporation at 4 and 15 dpe. (B–E) No differences were observed at the level of the lateral ventricle or in the RMS at 4 dpe. (F) Cell distribution along the rostro-caudal axis was normal (definition of levels in Fig. 2l). (G and H′′) Consequences of NeuroD1 knockdown on PGN morphology at 15 dpe. (G) Whereas shRNAs showing a strong effect on NeuroD1 expression strongly inhibited morphological differentiation, the weakly active shRNA 777 had only a minor effect compared with control. (H) Examples of cells that served for classification of PGN. Class1 cells present primary and secondary branching. Dendritic spines (arrowheads) indicate their synaptic integration in OB circuitry. Class 2 cells present a single primary branch. Class 3 cells present a spherical morphology and no branching. Errors bars indicate SEM. Statistics, unpaired t test. ns, not significant. **P < 0.01; ***P < 0.005. (Scale bar: 100 μm in B–E; 20 μm in H.
………
When the two highly active NeuroD1-specific shRNAs sh775 and sh776 were electroporated, the vast majority of cells in the GL showed simple morphologies with few or no processes (classes 2 and 3), whereas cells with complex neuronal morphologies were sparse (Fig. 4G). When the less-efficient shRNA sh777 was expressed, an intermediate degree of neuronal maturation was observed (Fig. 4G), suggesting a dose-dependent action of NeuroD1 under these conditions. Comparable results were obtained for the GCL. As in the PGL, knockdown of NeuroD1 induced a dose-dependent inhibition of terminal neuronal differentiation (Fig. S9 A and B).
Thus, knockdown of NeuroD1 did not notably interfere with early steps of interneuron generation, but induced a specific defect in the acquisition of the differentiated neuronal phenotype in the OB.
Discussion
Although considerable information is available concerning the generation, specification, and migration of neurons, little is known concerning the factors and regulatory cascades that maintain the immature neuronal precursor status or induce the exit from this state and trigger terminal differentiation. Using a systematic approach, we identified NeuroD1 as a candidate for the latter function and validated this role using gain- and loss-of function approaches.
In Xenopus, a late function of NeuroD1 has been suggested based on two lines of evidence (13). First, NeuroD1 is transitorily expressed in territories where neuronal differentiation occurs. Second, misexpression of NeuroD1 causes the premature differentiation of neuronal precursors into neurons. However, the observation that NeuroD1 could also convert presumptive epidermal cells into neurons pointed toward a determination function. Therefore, a doubtless discrimination between a proneural and a terminal differentiation function was not possible.
The above-cited pioneering work in the frog has been extended through the analysis of mice with mutations in the NeuroD1 gene (14, 15, 21). In the hippocampal dentate gyrus of such animals, granule cell precursors are generated correctly in the neuroepithelium and invade the hippocampal anlage. However, in the target structure, precursors show a severe deficit in proliferation, and a defined dentate gryus is not formed (15). In the mutant cerebellum, generation and migration of early precursors appear not to be affected. Nevertheless, once these cells become postmitotic, massive cell death is observed and the cerebellum is severely affected (14). Thus, in these systems a late function of NeuroD1 is already suggested. However, because of the complexity of the models and the relatively low level of resolution, the available information is still fragmentary.
We attempted to clarify the role of NeuroD1 in neuronal differentiation by analyzing its function during olfactory neurogenesis. Using SAGE, microarray, in situ hybridization, and lacZ knockin into the NeuroD1 locus, we have demonstrated that NeuroD1 is expressed in mature neurons of the OB but is absent from immature stages. These findings are in contrast to recent expression data based on a NeuroD1 antibody, suggesting expression of the transcription factor already in the SVZ and RMS (23, 29). However, our loss-of-function approach based on RNAi shows that NeuroD1 is dispensable for generation and migration of precursors but is necessary for their transition into neurons in the target layer. These findings are in agreement with those of a recent study based on conditional NeuroD1 mutants, which showed a comparable defect in the OB (29).
……
This work demonstrates that expression of a single transcription factor can induce massive ectopic neuronal differentiation of neural stem cells in the vertebrate forebrain. The existence of postnatal and adult neurogenesis holds potential for the treatment of neurodegenerative diseases (34). However, in many experimental paradigms, transplanted or recruited cells fail to undergo differentiation into neurons and either transdifferentiate into glia or remain immature precursors (18, 35). It appears conceivable to combine such approaches with the strong neuronal differentiation inducing activity of NeuroD1.
Scientists Unveil Critical Mechanism of Memory Formation
http://www.dddmag.com/news/2015/11/scientists-unveil-critical-mechanism-memory-formation

{kind=link}
Follow Blog via Email
Join 2,041 other subscribers-
Recent Posts
- Protected: KOL on Competitive Dynamics: LPBI Group’s Founder’s Radar Screens on AI in Health, Life Sciences & BioPharma June 15, 2026
- Tumor Infiltrating Lymphocytes (TIL) as a first of kind FDA approved immunotherapy for cancer June 12, 2026
- A Systematic Analysis of Competitive Dynamics in the AI Revolution — Strategic Relevance to LPBI Group’s Mission – Alex Wissner-Gross Daily Newsletters by KOL (March 1 – June 10, 2026, to be extended to Present) June 11, 2026
- In Memoriam: Professor Yitzhak Apeloig, President and Distinguised Professor of the Technion June 4, 2026
- Top VC Investors in AI / Health AI in 2026 May 26, 2026
- Grok thinks like PhDs – Hybrid Model for Autonomous Update of Doctoral Dissertations with Original PhD Student Author editing and acceptance of Grok’s updated output. 3rd Joint article, a pilot study for Validation of an Autonomous Journal Articles Updating System (AJAUS) tested on Autonomous Update of Aviva Lev-Ari, PhD, RN Doctoral Thesis, UC, Berkeley, 1983. January 31, 2026
- 2026 World Economic Forum, 1/19 – 1/23/2026, Davos, Switzerland, LPBI Group’s KOLs interpretation of AI in Health Videos January 24, 2026
- Evolving LPBI Group’s Portfolio of Intellectual Properties (IP): From 2021 Vision to 2026 Reality January 12, 2026
- Aviva Lev-Ari, PhD, RN – Biography ->> Grokepedia Entry January 11, 2026
- List of Articles included in the Article SELECTION from Collection of Aviva Lev-Ari, PhD, RN Scientific Articles on PULSE on LinkedIn.com for Training Small Language Models (SLMs) in Domain-aware Content of Medical, Pharmaceutical, Life Sciences and Healthcare by 15 Subjects Matter January 10, 2026
Archives
Categories
Meta