Healthcare analytics, AI solutions for biological big data, providing an AI platform for the biotech, life sciences, medical and pharmaceutical industries, as well as for related technological approaches, i.e., curation and text analysis with machine learning and other activities related to AI applications to these industries.
Genetic scissors: a tool for rewriting the code of life
Emmanuelle Charpentier and Jennifer A. Doudna have discovered one of gene technology’s sharpest tools: the CRISPR/Cas9 genetic scissors. Using these, researchers can change the DNA of animals, plants and microorganisms with extremely high precision. This technology has had a revolutionary impact on the life sciences, is contributing to new cancer therapies and may make the dream of curing inherited diseases come true.
Researchers need to modify genes in cells if they are to find out about life’s inner workings. This used to be time-consuming, difficult and sometimes impossible work. Using the CRISPR/Cas9 genetic scissors, it is now possible to change the code of life over the course of a few weeks.
“There is enormous power in this genetic tool, which affects us all. It has not only revolutionised basic science, but also resulted in innovative crops and will lead to ground-breaking new medical treatments,” says Claes Gustafsson, chair of the Nobel Committee for Chemistry.
As so often in science, the discovery of these genetic scissors was unexpected. During Emmanuelle Charpentier’s studies of Streptococcus pyogenes, one of the bacteria that cause the most harm to humanity, she discovered a previously unknown molecule, tracrRNA. Her work showed that tracrRNA is part of bacteria’s ancient immune system, CRISPR/Cas, that disarms viruses by cleaving their DNA.
Charpentier published her discovery in 2011. The same year, she initiated a collaboration with Jennifer Doudna, an experienced biochemist with vast knowledge of RNA. Together, they succeeded in recreating the bacteria’s genetic scissors in a test tube and simplifying the scissors’ molecular components so they were easier to use.
In an epoch-making experiment, they then reprogrammed the genetic scissors. In their natural form, the scissors recognise DNA from viruses, but Charpentier and Doudna proved that they could be controlled so that they can cut any DNA molecule at a predetermined site. Where the DNA is cut it is then easy to rewrite the code of life.
Since Charpentier and Doudna discovered the CRISPR/Cas9 genetic scissors in 2012 their use has exploded. This tool has contributed to many important discoveries in basic research, and plant researchers have been able to develop crops that withstand mould, pests and drought. In medicine, clinical trials of new cancer therapies are underway, and the dream of being able to cure inherited diseases is about to come true. These genetic scissors have taken the life sciences into a new epoch and, in many ways, are bringing the greatest benefit to humankind.
Emmanuelle Charpentier, born 1968 in Juvisy-sur-Orge, France. Ph.D. 1995 from Institut Pasteur, Paris, France. Director of the Max Planck Unit for the Science of Pathogens, Berlin, Germany.
Jennifer A. Doudna, born 1964 in Washington, D.C, USA. Ph.D. 1989 from Harvard Medical School, Boston, USA. Professor at the University of California, Berkeley, USA and Investigator, Howard Hughes Medical Institute.
Other Articles on the Nobel Prize in this Open Access Journal Include:
Prime Editing as a New CRISPR Tool to Enhance Precision and Versatility
Curator: Stephen J. Williams, Ph.D.
The following articles is structured as follows
1: Recent update of financial statements from Editas Medicine, with a brief history of the company from formation to present
2: Main article which describes Editas Medicine’s CRISPR/Cas9 gene editing system and advantages of their system over competitors. This includes the early seminal publications showing utility of the system in humans
UPDATED 05/31/2026
The following press release announces a major public offering of shares of Editas Medicine, a biotechnology company founded in 2013 by Nobel Leaureates Dr. Jennifer Doudna, and Dr. George Church, as well as other prominent scientists to use the gene editing capabilities of the CRISPR/Cas9 system to cure genetic diseases. After initial funding of $43 million from Third Rock Ventures and Polaris Partners, Dr. Doudna left in 2014 as a result of patent disagreements. The company has had early successes with ‘in-vivo’ CRISPR in a clinical trial with EDIT-101 for Leber congenital amaurosis 10 (LCA10), however this was not able to reach market. Programs for correcting defects ex-vivo for sickle cell disease and beta-thalassemia (renicel) was terminated in 2024. Due to stiff competition from Veritas and CRSPR Therapeutics, as well as inability to secure a commercial partner, Editas Medicine pivoted their strategy in late 2024. The need for capital is reflecting in their public offering however they still maintain a strong IP portfoliol
CAMBRIDGE, Mass., May 26, 2026 (GLOBE NEWSWIRE) — Editas Medicine, Inc. (Nasdaq: EDIT), a pioneering gene editing company developing transformative medicines for serious diseases, today announced the pricing of an underwritten public offering of 55,555,556 shares of its common stock and accompanying common stock warrants to purchase an aggregate of 55,555,556 shares of common stock (or pre-funded warrants in lieu thereof). Each share of common stock and accompanying common stock warrant are being sold together at a combined public offering price of $2.25. The aggregate gross proceeds from the offering are expected to be approximately $125.0 million (assuming no exercise of the common stock warrants), before deducting underwriting discounts and commissions and offering expenses. If all of the common stock warrants are exercised at their exercise price, the Company would receive additional gross proceeds from the offering of approximately $194.4 million before deducting underwriting discounts and commissions and offering expenses.
Each common stock warrant will be exercisable for shares of common stock (or pre-funded warrants in lieu thereof), will have an exercise price of $3.50 per share (or $3.4999 per share if exercised for pre-funded warrants), will be exercisable immediately and will expire on the earlier of (i) the date that is thirty (30) days following the first public announcement by the Company of Phase 1 clinical data for the Company’s product candidate, EDIT-401, that discloses at least three patients in the trial that each demonstrated greater than 80% reduction in LDL-cholesterol as compared to baseline with at least one (1) month of follow-up and (ii) three years from the date of issuance. Any pre-funded warrants issued upon the exercise of common stock warrants will have an exercise price of $0.0001 per share of common stock, will be immediately exercisable and will expire on the date the pre-funded warrant is exercised in full.
All of the securities in the offering are being sold by Editas Medicine. The offering is expected to close on or about May 27, 2026, subject to satisfaction of customary closing conditions.Cantor and Wells Fargo Securities are acting as joint book-running managers for the offering.The securities are being offered pursuant to an effective shelf registration statement on Form S-3 (File No. 333-277471) that was filed with the Securities and Exchange Commission (SEC) on February 28, 2024, as amended by Post-Effective Amendment No. 1 to Form S-3 Registration Statement and Post-Effective Amendment No. 2 to Form S-3 Registration Statement, each filed with the SEC on March 5, 2025, and declared effective on March 21, 2025. The offering is being made only by means of a prospectus supplement and accompanying prospectus that form a part of the registration statement. A preliminary prospectus supplement and accompanying prospectus relating to and describing the terms of the offering have been filed with the SEC and are available at www.sec.gov. A final prospectus supplement relating to the offering will be filed with the SEC and will be available for free on the SEC’s website at www.sec.gov. Copies of the final prospectus supplement may be obtained, when available, by contacting Cantor Fitzgerald & Co., Attention: Capital Markets, 110 East 59th Street, 6th Floor New York, New York 10022, Email: prospectus@cantor.com; or Wells Fargo Securities, LLC, Attention: Equity Syndicate Department, 90 South 7th Street, 5th Floor, Minneapolis, Minnesota 55402, at (800) 645-3751 (option #5) or email a request to WFScustomerservice@wellsfargo.com. This press release does not constitute an offer to sell or the solicitation of an offer to buy these securities, nor shall there be any sale of these securities in any state or jurisdiction in which such offer, solicitation or sale would be unlawful prior to registration or qualification under the securities laws of any such state or jurisdiction.
About Editas Medicine As a pioneering gene editing company, Editas Medicine is focused on translating the power and potential of CRISPR genome editing systems into a robust pipeline of transformative in vivo medicines for people living with serious diseases around the world. Editas Medicine aims to discover, develop, manufacture, and commercialize durable, precision in vivo gene editing medicines for a broad class of diseases. Editas Medicine is the exclusive licensee of Broad Institute’s Cas12a patent estate and Broad Institute and Harvard University’s Cas9 patent estates for human medicines.
The following curation describes the technology developed by Editas Medicine which reduces off target effects and is the basis for the in-vivo and ex-vivo therapeutic CRSPR/Cas9 technology. Seminal publication are included,
CRISPR has become a powerful molecular for the editing of genomes tool in research, drug discovery, and the clinic
there have been many instances of off-target effects where genes, other than the selected target, are edited out. This ‘off-target’ issue has hampered much of the utility of CRISPR in gene-therapy and CART therapy
However, an article in Science by Jon Cohen explains a Nature paper’s finding of a new tool in the CRISPR arsenal called prime editing, meant to increase CRISPR specificity and precision editing capabilities.
Primeediting promises to be a cut above CRISPR Jon Cohen CRISPR, an extraordinarily powerful genome-editing tool invented in 2012, can still be clumsy. … Primeediting steers around shortcomings of both techniques by heavily modifying the Cas9 protein and the guide RNA. … ” Primeediting “well may become the way that disease-causing mutations are repaired,” he says.
The effort, led by Drs. David Liu and Andrew Anzalone at the Broad Institute (Cambridge, MA), relies on the modification of the Cas9 protein and guide RNA, so that there is only a nick in a single strand of the double helix. The canonical Cas9 cuts both strands of DNA, and so relies on an efficient gap repair activity of the cell. The second part, a new type of guide RNA called a pegRNA, contains an RNA template for a new DNA sequence to be added at the target location. This pegRNA-directed synthesis of the new template requires the attachment of a reverse transcriptase enzymes to the Cas9. So far Liu and his colleagues have tested the technology on over 175 human and rodent cell lines with great success. In addition, they had also corrected mutations which cause Tay Sachs disease, which previous CRISPR systems could not do. Liu claims that this technology could correct over 89% of pathogenic variants in human diseases.
A company Prime Medicine has been formed out of this effort.
As was announced, prime editing for human therapeutics will be jointly developed by both Prime Medicine and Beam Therapeutics, each focusing on different types of edits and distinct disease targets, which will help avoid redundancy and allow us to cover more disease territory overall. The companies will also share knowledge in prime editing as well as in accompanying technologies, such as delivery and manufacturing.
Reader of StatNews.: Can you please compare the pros and cons of prime editing versus base editing?
The first difference between base editing and prime editing is that base editing has been widely used for the past 3 1/2 years in organisms ranging from bacteria to plants to mice to primates. Addgene tells me that the DNA blueprints for base editors from our laboratory have been distributed more than 7,500 times to more than 1,000 researchers around the world, and more than 100 research papers from many different laboratories have been published using base editors to achieve desired gene edits for a wide variety of applications. While we are very excited about prime editing, it’s brand-new and there has only been one paper published thus far. So there’s much to do before we can know if prime editing will prove to be as general and robust as base editing has proven to be.
We directly compared prime editors and base editors in our study, and found that current base editors can offer higher editing efficiency and fewer indel byproducts than prime editors, while prime editors offer more targeting flexibility and greater editing precision. So when the desired edit is a transition point mutation (C to T, T to C, A to G, or G to A), and the target base is well-positioned for base editing (that is, a PAM sequence exists approximately 15 bases from the target site), then base editing can result in higher editing efficiencies and fewer byproducts. When the target base is not well-positioned for base editing, or when other “bystander” C or A bases are nearby that must not be edited, then prime editing offers major advantages since it does not require a precisely positioned PAM sequence and is a true “search-and-replace” editing capability, with no possibility of unwanted bystander editing at neighboring bases.
Of course, for classes of mutations other than the four types of point mutations that base editors can make, such as insertions, deletions, and the eight other kinds of point mutations, to our knowledge prime editing is currently the only approach that can make these mutations in human cells without requiring double-stranded DNA cuts or separate DNA templates.
Nucleases (such as the zinc-finger nucleases, TALE nucleases, and the original CRISPR-Cas9), base editors, and prime editors each have complementary strengths and weaknesses, just as scissors, pencils, and word processors each have unique and useful roles. All three classes of editing agents already have or will have roles in basic research and in applications such as human therapeutics and agriculture.
Most genetic variants that contribute to disease1 are challenging to correct efficiently and without excess byproducts2,3,4,5. Here we describe prime editing, a versatile and precise genome editing method that directly writes new genetic information into a specified DNA site using a catalytically impaired Cas9 endonuclease fused to an engineered reverse transcriptase, programmed with a prime editing guide RNA (pegRNA) that both specifies the target site and encodes the desired edit. We performed more than 175 edits in human cells, including targeted insertions, deletions, and all 12 types of point mutation, without requiring double-strand breaks or donor DNA templates. We used prime editing in human cells to correct, efficiently and with few byproducts, the primary genetic causes of sickle cell disease (requiring a transversion in HBB) and Tay–Sachs disease (requiring a deletion in HEXA); to install a protective transversion in PRNP; and to insert various tags and epitopes precisely into target loci. Four human cell lines and primary post-mitotic mouse cortical neurons support prime editing with varying efficiencies. Prime editing shows higher or similar efficiency and fewer byproducts than homology-directed repair, has complementary strengths and weaknesses compared to base editing, and induces much lower off-target editing than Cas9 nuclease at known Cas9 off-target sites. Prime editing substantially expands the scope and capabilities of genome editing, and in principle could correct up to 89% of known genetic variants associated with human diseases.
From Anzolone et al. Nature 2019 Figure 1.
Prime editing strategy
Cas9 targets DNA using a guide RNA containing a spacer sequence that hybridizes to the target DNA site. We envisioned the generation of guide RNAs that both specify the DNA target and contain new genetic information that replaces target DNA nucleotides. To transfer information from these engineered guide RNAs to target DNA, we proposed that genomic DNA, nicked at the target site to expose a 3′-hydroxyl group, could be used to prime the reverse transcription of an edit-encoding extension on the engineered guide RNA (the pegRNA) directly into the target site (Fig. 1b, c, Supplementary Discussion).. These initial steps result in a branched intermediate with two redundant single-stranded DNA flaps: a 5′ flap that contains the unedited DNA sequence and a 3′ flap that contains the edited sequence copied from the pegRNA (Fig. 1c). Although hybridization of the perfectly complementary 5′ flap to the unedited strand is likely to be thermodynamically favoured, 5′ flaps are the preferred substrate for structure-specific endonucleases such as FEN122, which excises 5′ flaps generated during lagging-strand DNA synthesis and long-patch base excision repair. The redundant unedited DNA may also be removed by 5′ exonucleases such as EXO123.
The authors reasoned that preferential 5′ flap excision and 3′ flap ligation could drive the incorporation of the edited DNA strand, creating heteroduplex DNA containing one edited strand and one unedited strand (Fig. 1c).
DNA repair to resolve the heteroduplex by copying the information in the edited strand to the complementary strand would permanently install the edit (Fig. 1c).
They had hypothesized that nicking the non-edited DNA strand might bias DNA repair to preferentially replace the non-edited strand.
Results
The authors evaluated the eukaryotic cell DNA repair outcomes of 3′ flaps produced by pegRNA-programmed reverse transcription in vitro, and performed in vitro prime editing on reporter plasmids, then transformed the reaction products into yeast cells (Extended Data Fig. 2).
Reporter plasmids encoding EGFP and mCherry separated by a linker containing an in-frame stop codon, +1 frameshift, or −1 frameshift were constructed and when plasmids were edited in vitro with Cas9 nickase, RT, and 3′-extended pegRNAs encoding a transversion that corrects the premature stop codon, 37% of yeast transformants expressed both GFP and mCherry (Fig. 1f, Extended Data Fig. 2).
They fused a variant of M—MLV-RT (reverse transcriptase) to Cas9 with an extended linker and this M-MLV RT fused to the C terminus of Cas9(H840A) nickase was designated as PE1. This strategy allowed the authors to generate a cell line containing all the required components of the primer editing system. They constructed 19 variants of PE1 containing a variety of RT mutations to evaluate their editing efficiency in human cells
Generated a pentamutant RT incorporated into PE1 (Cas9(H840A)–M-MLV RT(D200N/L603W/T330P/T306K/W313F)) is hereafter referred to as prime editor 2 (PE2). These were more thermostable versions of RT with higher efficiency.
Optimized the guide (pegRNA) using a series of permutations and recommend starting with about 10–16 nt and testing shorter and longer RT templates during pegRNA optimization.
In the previous attempts (PE1 and PE2 systems), mismatch repair resolves the heteroduplex to give either edited or non-edited products. So they next developed an optimal editing system (PE3) to produce optimal nickase activity and found nicks positioned 3′ of the edit about 40–90 bp from the pegRNA-induced nick generally increased editing efficiency (averaging 41%) without excess indel formation (6.8% average indels for the sgRNA with the highest editing efficiency) (Fig. 3b).
The cell line used to finalize and validate the system was predominantly HEK293T immortalized cell line
Together, their findings establish that PE3 systems improve editing efficiencies about threefold compared with PE2, albeit with a higher range of indels than PE2. When it is possible to nick the non-edited strand with an sgRNA that requires editing before nicking, the PE3b system offers PE3-like editing levels while greatly reducing indel formation.
Off Target Effects: Strikingly, PE3 or PE2 with the same 16 pegRNAs containing these four target spacers resulted in detectable off-target editing at only 3 out of 16 off-target sites, with only 1 of 16 showing an off-target editing efficiency of 1% or more (Extended Data Fig. 6h). Average off-target prime editing for pegRNAs targeting HEK3, HEK4, EMX1, and FANCFat the top four known Cas9 off-target sites for each protospacer was <0.1%, <2.2 ± 5.2%, <0.1%, and <0.13 ± 0.11%, respectively (Extended Data Fig. 6h).
The PE3 system was very efficient at editing the most common mutation that causes Tay-Sachs disease, a 4-bp insertion in HEXA(HEXA1278+TATC).
References
Landrum, M. J. et al. ClinVar: public archive of interpretations of clinically relevant variants. Nucleic Acids Res. 44, D862–D868 (2016).
Jinek, M. et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science337, 816–821 (2012).
Kosicki, M., Tomberg, K. & Bradley, A. Repair of double-strand breaks induced by CRISPR–Cas9 leads to large deletions and complex rearrangements. Biotechnol. 36, 765–771 (2018).
Human gene editing continues to hold a major fascination within a biomedical and biopharmaceutical industries. It’s extraordinary potential is now being realized but important questions as to who will be the beneficiaries of such breakthrough technologies remained to be answered. The session will discuss whether gene editing technologies can alleviate some of the most challenging unmet medical needs. We will discuss how research advances often never reach minority communities and how diverse patient populations will gain access to such breakthrough technologies. It is widely recognize that there are patient voids in the population and we will explore how community health centers might fill this void to ensure that state-of-the-art technologies can reach the forgotten patient groups . We also will touch ethical questions surrounding germline editing and how such research and development could impact the community at large.
Please follow LIVE on TWITTER using the following @ handles and # hashtags:
CRISPR cuts turn gels into biological watchdogs, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 2: CRISPR for Gene Editing and DNA Repair
Update 6/11/2020
CRISPR-IL used to develop next-gen genome editing products
Haifa-based Pluristem Therapeutics is a regenerative medicine company that plans to develop next-generation multi-species genome editing products for human, plant and animal DNA that could improve work done in the pharma, agriculture and aquaculture industries.
The CRISPR-IL consortium includes Sheba Medical Center and Schneider Children’s Medical Center, Bar-Ilan University, Ben-Gurion University of the Negev, Hebrew University of Jerusalem, the Weizmann Institute of Science, IDC Herzliya and Tel-Aviv University.
This consortium is also joined by Pluristem Therapeutics, which plans to bring together a team of multi-disciplinary experts to develop artificial intelligence based end-to-end genome-editing solutions.
The genome editing product designed by Pluristerm should improve existing technology.
The project also includes “the computational design of on-target DNA modification, with minimal accidental, off-target modifications, improve modification efficiency.
The product provides an accurate measuring tool to ensure the desired modification.
Genome editing if of significant interest in the prevention and treatment of human diseases including single-gene disorders such as cystic fibrosis, hemophilia and sickle cell disease. It also shows great promise for the prevention and treatment of diseases such as cancer, heart disease, mental illness and human immunodeficiency virus infection. However, ethical concerns arise when genome editing, using technologies such as CRISPR-Cas9 is used to alter human genomes.
James Collins, bioengineer at MIT and his team worked with water-filled polymers that are held together by strands of DNA, known as DNA hydrogels. To alter the properties of these materials, these scientists turned to a form of CRISPR that uses a DNA-snipping enzyme called Cas12a, which can be programed to recognize a specific DNA sequence. The enzyme then cuts its target DNA strand, then severs single strands of DNA nearby. This property lets scientists to build a series of CRISPR-controlled hydrogels encapsulating a target DNA sequence and single strands of DNA, which break up after Cas12a identifies the target sequence in a stimulus. The break-up of the single DNA strands activates the hydrogels to change shape or completely dissolve, releasing a payload.
According to Collins and his team, the programmed hydrogels will release enzymes, small molecules and human cells as part of a smart therapy in response to stimuli. However, in order to make it a smart therapeutic, the researchers in collaboration with Dan Luo, bioengineer at Cornell University placed the CRISPR- controlled hydrogels into electric circuits. The circuit is switched off in response to the detection of the genetic material of harmful pathogens such as Ebola virus and methicillin-resistant Staphylococcus aureus. The team used these hydrogels to develop a prototype diagnostic tool that sends a wireless signal to identify Ebola in lab samples.
Yet, it is evident that these CRISPR-controlled hydrogels show great potential for the prevention and treatment of diseases.
Optimization of CRISPR Gene Editing with Gold Nanoparticles
Reporter: Irina Robu, PhD
The CRISPR-Cas9 gene editing system has been welcomed as a hopeful solution to a range of genetic diseases, but the expertise has proven hard to deliver into cells. One plan is to open the cell membrane using an electric shock, but that can accidentally kill the cell. Another is to use viruses as couriers. Problem is, viruses can cause off-target side effects.
CRISPR-Cas9 is a unique technology that enables geneticists and medical researchers to edit parts of the genome by removing, adding or altering sections of DNA sequence. It is faster, cheaper and more accurate than previous techniques of editing DNA and can have a wide range of potential applications.
The CRISPR-Cas9 system consists of two key molecules that introduce a change into the DNA. One is an enzyme called Cas9 which acts as a pair of molecular scissors that can cut the two strands of DNA at a specific location in the genome where bits of DNA can be added or removed. The other one, is a piece of RNA which consists of a small piece of pre-designed RNA sequence located within a longer RNA scaffold. The scaffold part binds to the DNA and pre-designed sequence which contains Cas9. The RNA sequence is designed to find and locate specific sequence in the DNA. The Cas9 trails the guide RNA to the same location in the DNA sequence and makes a cut across both strands of DNA. At this point the cell distinguishes that the DNA is damaged and tries to repair it.
Researchers at Fred Hutchinson Cancer Research Center published new findings in Nature Materials suggested an alternative delivery method such as gold nanoparticles. The gold nanoparticles are packed with all the CRISPR components necessary to make clean gene edits. When the gold nanoparticles were tested in lab models of inherited blood disorders and HIV, between 10% and 20% of the targeted cells were effectively edited, with no toxic side effects.
The researchers use gold nanoparticles to deliver CRISPR to blood stem cells. Each gold nanoparticle contains four CRISPR components, including the enzyme needed to make the DNA cuts. But Fred Hutchinson researchers chose Cas12a, which they believed would lead to more efficient edits. Plus, Cas12a only needs one molecular guide, while Cas9 requires two.
In one experiment, they sought to disturb the gene CCR5 to make cells resistant to HIV. In the second, they created a gene mutation that can protect against blood disorders, including sickle cell disease. They observed the cells encapsulated the nanoparticles within six hours and began the gene-editing process within 48 hours. In mice, gene editing peaked eight weeks after injection, and the edited cells were still in circulation 22 weeks after the treatment.
Researchers at Fred Hutchinson are now working on improving the efficiency of the gold-based CRISPR delivery system so that 50% or more of the targeted cells are edited and are also looking for a commercial partner to bring the technology to clinical phase in the next few years.
Advances in Gene Editing and Gene Silencing | September 20-21, 2016 | Boston, MA
Reporter: Aviva Lev-Ari, PhD, RN
2.1.5.28 Advances in Gene Editing and Gene Silencing | September 20-21, 2016 | Boston, MA, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 2: CRISPR for Gene Editing and DNA Repair
KEYNOTE SESSION: GENOME EDITING FOR IN VIVO APPLICATIONS
Part 1 (of a two-part conference) will cover the use of CRISPR/Cas9 and RNAi for identifying new drug targets and therapies. It will bring together experts from all aspects of basic science and clinical research to talk about how and where gene editing and RNAi can be best applied. What are the different tools that can be used and what are their strengths and limitations? How does the CRISPR/Cas system compare to RNAi and other gene editing tools, such as Transcription Activator-like Effector Nucleases (TALENs) and zinc finger nucleases (ZFNs), and do they have any complementary uses? Scientists and clinicians from pharma/biotech as well as from academic and government labs will share their experiences leveraging the utility of gene editing for target discovery, disease modeling, and for creating cell and viral therapies. Learn more atDiscoveryOnTarget.com/RNAi-screens-functional-genomics
Keynote Session: Genome Editing for InVivo Applications
AAV for Gene Therapy and Genome Editing James Wilson, M.D., Ph.D., Professor, Department of Pathology and Laboratory Medicine, Perelman School of Medicine; Director, Orphan Disease Center and Director, Gene Therapy Program, University of Pennsylvania In vivo delivery of nucleic acid therapeutics remains the primary barrier to success. My lab has focused on the use of vectors based on adeno-associated virus (AAV) for achieving success in pre-clinical and clinical applications of gene replacement therapy. Most of the current academic and commercial applications of in vivo gene replacement therapy are based on endogenous AAVs we discovered as latent viral genomes in primates. These vectors are reasonably safe and efficient for application of gene replacement therapy. The emergence of genome editing methods has suggested more precise and effective methods to treat inherited diseases in which genes are silenced or mutations are corrected. AAV vectors have been the most efficient platform for achieving genome editing in vivo. We will review our attempts to achieve therapeutic genome editing in animal models of liver disease using AAV.
Using CRISPR/Cas to Target and Destroy Viral DNA Genomes Bryan R. Cullen, Ph.D., James B. Duke Professor of Molecular Genetics and Microbiology and Director, Center for Virology, Duke University
A number of pathogenic human DNA viruses, including HBV, HIV-1 and HSV1, cause chronic diseases in humans that remain refractory to cure, though these diseases can be controlled by antivirals. In addition the DNA virus HPV causes tumors that depend on the continued expression of viral genes. Here, I will present data demonstrating that several of these viruses can be efficiently cleaved and destroyed using viral vectors that express Cas9 and virus-specific guide RNAs, thus providing a potential novel approach to treatment.
Targeted Endonucleases as Antiviral Agents: Promises and Pitfalls Keith R. Jerome, M.D., Ph.D., Member, Vaccine and Infectious Disease Division, Fred Hutchinson Cancer Research Center; Professor and Head, Virology Division, Department of Laboratory Medicine, University of Washington
Genome editing offers the prospect of cure for infections such as HIV, hepatitis B virus, herpes simplex, and human papillomavirus, by disruption of essential viral nucleic acids or the human genes encoding receptors needed for viral entry. This talk will highlight the most recent laboratory data and the challenges still ahead in bringing this technology to the clinic.
Nucleic Acid Delivery Systems for RNA Therapy and Gene Editing Daniel Anderson, Ph.D., Professor, Department of Chemical Engineering, Institute for Medical Engineering & Science, Harvard-MIT Division of Health Sciences & Technology and David H. Koch Institute for Integrative Cancer Research, Massachusetts Institute of Technology
High throughput, combinatorial approaches have revolutionized small molecule drug discovery. Here we describe our high throughput methods for developing and characterizing RNA delivery and gene editing systems. Libraries of degradable polymers and lipid-like materials have been synthesized, formulated and screened for their ability to deliver RNA, both in vitro and in vivo. A number of delivery formulations have been developed with in vivo efficacy, and show potential applications for the treatment of genetic diseases, viral infections and cancers.
PANEL DISCUSSION: CRISPR/Cas: A Realistic and Practical Look at What the Future Could Hold
Moderator: Bryan R. Cullen, Ph.D., James B. Duke Professor of Molecular Genetics and Microbiology and Director, Center for Virology, Duke University
Participants: Session Speakers
Each speaker will spend a few minutes sharing their viewpoints and experiences on where things stand with using the CRISPR/Cas system for in vivo applications. Attendees will have an opportunity to ask questions and share their opinions.
About the Conference
Cambridge Healthtech Institute’s 13th annual two-part conference on Advances in Gene Editing and Gene Silencing will cover the latest in the use of CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas9-based gene editing and RNA interference (RNAi) for use in drug discovery and for developing novel drug therapies.
For sponsorship and exhibit sales information including podium presentation opportunities, contact:
Jon Stroup | T: +1 781-972-5483 | E: jstroup@healthtech.com
Recommended All Access Package:
Includes access to 1 Symposium and 2 Conferences
Cambridge Healthtech Institute, 250 First Avenue, Suite 300, Needham, MA, USA Tel: 781-972-5400 | Fax: 781-972-5425 | www.healthtech.com
This email is being sent to sjwilliamspa@comcast.net for marketing purposes. If it is not of interest to you, please disregard and we apologize for any inconvenience this may have caused.
WASHINGTON — At scientific meetings on genome-editing, you’d expect researchers to show pretty slides of the ribbony 3-D structure of the CRISPR-Cas9 molecules neatly snipping out disease-causing genes in order to, everyone hopes, cure illnesses from cancer to muscular dystrophy. Less expected: slides of someone kneeling on a beach with his head in the sand.
Yet that is what Dr. J. Keith Joung of Massachusetts General Hospital showed at the American Society of Hematology’s workshop on genome-editing last week in Washington. While the 150 experts from industry, academia, the National Institutes of Health, and the Food and Drug Administration were upbeat about the possibility of using genome-editing to treat and even cure sickle cell disease, leukemia, HIV/AIDS, and other blood disorders, there was a skunk at the picnic: an emerging concern that some enthusiastic CRISPR-ers are ignoring growing evidence that CRISPR might inadvertently alter regions of the genome other than the intended ones.
“In the early days of this field, algorithms were generated to predict off-target effects and [made] available on the web,” Joung said. Further research has shown, however, that such algorithms, including one from MIT and one calledE-CRISP, “miss a fair number” of off-target effects. “These tools are used in a lot of papers, but they really aren’t very good at predicting where there will be off-target effects,” he said. “We think we can get off-target effects to less than 1 percent, but we need to do better,” especially if genome-editing is to be safely used to treat patients.
Off-target effects occur because of how CRISPR works. It has two parts. RNA makes a beeline for the site in a genome specified by the RNA’s string of nucleotides, and an enzyme cuts the genome there. Trouble is, more than one site in a genome can have the same string of nucleotides. Scientists might address CRISPR to the genome version of 123 Main Street, aiming for 123 Main on chromosome 9, only to find CRISPR has instead gone to 123 Main on chromosome 14.
In one example Joung showed, CRISPR is supposed to edit a gene called VEGFA (which stimulates production of blood vessels, including those used by canceroustumors) on chromosome 6. But, studies show, this CRISPR can also hit genes on virtually every one of the other 22 human chromosomes. The same is true for CRISPRs aimed at other genes. Although each CRISPR has zero to a dozen or so “known” off-target sites (where “known” means predicted by those web-based algorithms), Joung said, there can be as many as 150 “novel” off-target sites, meaning scientists had no idea those errors were possible.
One reason for concern about off-target effects is that genome-editing might disable a tumor-suppressor gene or activate a cancer-causing one. It might also allow pieces of two different chromosomes to get together, a phenomenon called translocation, which is the cause of chronic myeloid leukemia, among other problems.
Many researchers, including those planning clinical trials, are using web-based algorithms to predict which regions of the genome might get accidentally CRISPR’d. They include the scientists whose proposal to use CRISPR in patients was the first to be approved by an NIH committee. When scientists assure regulators that they looked for off-target effects in CRISPR’d cells growing in lab dishes, what they usually mean is that they looked for CRISPR’ing of genes that the algorithms flagged.
As a result, off-target effects might be occurring but, because scientists are doing the equivalent of the drunk searching for their lost keys only under the lamppost, they’re not being found.
Other articles on CRISPR and Gene Editing on this Open Access Journal Include:
Gene Editing with CRISPR gets Crisper, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 2: CRISPR for Gene Editing and DNA Repair
Gene Editing with CRISPR gets Crisper
Curators: Larry H. Bernstein, MD, FCAP and Aviva Lev-Ari, PhD, RN
CRISPR Moves from Butchery to Surgery
More Genomes Are Going Under the CRISPR Knife, So Surgical Standards Are Rising
The Dharmacon subsidary of GE Healthcare provides the Edit-R Lentiviral Gene Engineering platform. It is based on the natural S. pyrogenes system, but unlike that system, which uses a single guide RNA (sgRNA), the platform uses two component RNAs, a gene-specific CRISPR RNA (crRNA) and a universal trans-activating crRNA (tracrRNA). Once hybridized to the universal tracrRNA (blue), the crRNA (green) directs the Cas9 nuclease to a specific genomic region to induce a double- strand break.
Scientists recently convened at the CRISPR Precision Gene Editing Congress, held in Boston, to discuss the new technology. As with any new technique, scientists have discovered that CRISPR comes with its own set of challenges, and the Congress focused its discussion around improving specificity, efficiency, and delivery.
In the naturally occurring system, CRISPR-Cas9 works like a self-vaccination in the bacterial immune system by targeting and cleaving viral DNA sequences stored from previous encounters with invading phages. The endogenous system uses two RNA elements, CRISPR RNA (crRNA) and trans-activating RNA (tracrRNA), which come together and guide the Cas9 nuclease to the target DNA.
Early publications that demonstrated CRISPR gene editing in mammalian cells combined the crRNA and tracrRNA sequences to form one long transcript called asingle-guide RNA (sgRNA). However, an alternative approach is being explored by scientists at the Dharmacon subsidiary of GE Healthcare. These scientists have a system that mimics the endogenous system through a synthetic two-component approach thatpreserves individual crRNA and tracrRNA. The tracrRNA is universal to any gene target or species; the crRNA contains the information needed to target the gene of interest.
Predesigned Guide RNAs
In contrast to sgRNAs, which are generated through either in vitro transcription of a DNA template or a plasmid-based expression system, synthetic crRNA and tracrRNA eliminate the need for additional cloning and purification steps. The efficacy of guide RNA (gRNA), whether delivered as a sgRNA or individual crRNA and tracrRNA, depends not only on DNA binding, but also on the generation of an indel that will deliver the coup de grâce to gene function.
“Almost all of the gRNAs were able to create a break in genomic DNA,” said Louise Baskin, senior product manager at Dharmacon. “But there was a very wide range in efficiency and in creating functional protein knock-outs.”
To remove the guesswork from gRNA design, Dharmacon developed an algorithm to predict gene knockout efficiency using wet-lab data. They also incorporated specificity as a component of their algorithm, using a much more comprehensive alignment tool to predict potential off-target effects caused by mismatches and bulges often missed by other alignment tools. Customers can enter their target gene to access predesigned gRNAs as either two-component RNAs or lentiviral sgRNA vectors for multiple applications.
“We put time and effort into our algorithm to ensure that our guide RNAs are not only functional but also highly specific,” asserts Baskin. “As a result, customers don’t have to do any design work.”
MilliporeSigma’s CRISPR Epigenetic Activator is based on fusion of a nuclease-deficient Cas9 (dCas9) to the catalytic histone acetyltransferase (HAT) core domain of the human E1A-associated protein p300. This technology allows researchers to target specific DNA regions or gene sequences. Researchers can localize epigenetic changes to their target of interest and see the effects of those changes in gene expression.
Knockout experiments are a powerful tool for analyzing gene function. However, for researchers who want to introduce DNA into the genome, guide design, donor DNA selection, and Cas9 activity are paramount to successful DNA integration.MilliporeSigma offers two formats for donor DNA: double-stranded DNA (dsDNA) plasmids and single-stranded DNA (ssDNA) oligonucleotides. The most appropriate format depends on cell type and length of the donor DNA. “There are some cell types that have immune responses to dsDNA,” said Gregory Davis, Ph.D., R&D manager, MilliporeSigma.
The ssDNA format can save researchers time and money, but it has a limited carrying capacity of approximately 120 base pairs.In addition to selecting an appropriate donor DNA format, controlling where, how, and when the Cas9 enzyme cuts can affect gene-editing efficiency. Scientists are playing tug-of-war, trying to pull cells toward the preferred homology-directed repair (HDR) and away from the less favored nonhomologous end joining (NHEJ) repair mechanism.One method to achieve this modifies the Cas9 enzyme to generate a nickase that cuts only one DNA strand instead of creating a double-strand break. Accordingly, MilliporeSigma has created a Cas9 paired-nickase system that promotes HDR, while also limiting off-target effects and increasing the number of sequences available for site-dependent gene modifications, such as disease-associated single nucleotide polymorphisms (SNPs).“The best thing you can do is to cut as close to the SNP as possible,” advised Dr. Davis. “As you move the double-stranded break away from the site of mutation you get an exponential drop in the frequency of recombination.”
Ribonucleo-protein Complexes
Another strategy to improve gene-editing efficiency, developed by Thermo Fisher, involves combining purified Cas9 protein with gRNA to generate a stable ribonucleoprotein (RNP) complex. In contrast to plasmid- or mRNA-based formats, which require transcription and/or translation, the Cas9 RNP complex cuts DNA immediately after entering the cell. Rapid clearance of the complex from the cell helps to minimize off-target effects, and, unlike a viral vector, the transient complex does not introduce foreign DNA sequences into the genome.
To deliver their Cas9 RNP complex to cells, Thermo Fisher has developed a lipofectamine transfection reagent called CRISPRMAX. “We went back to the drawing board with our delivery, screened a bunch of components, and got a brand-new, fully optimized lipid nanoparticle formulation,” explained Jon Chesnut, Ph.D., the company’s senior director of synthetic biology R&D. “The formulation is specifically designed for delivering the RNP to cells more efficiently.”
Besides the reagent and the formulation, Thermo Fisher has also developed a range of gene-editing tools. For example, it has introduced the Neon® transfection system for delivering DNA, RNA, or protein into cells via electroporation. Dr. Chesnut emphasized the company’s focus on simplifying complex workflows by optimizing protocols and pairing everything with the appropriate up- and downstream reagents.
From Mammalian Cells to Microbes
One of the first sources of CRISPR technology was the Feng Zhang laboratory at the Broad Institute, which counted among its first licensees a company called GenScript. This company offers a gene-editing service called GenCRISPR™ to establish mammalian cell lines with CRISPR-derived gene knockouts.
“There are a lot of challenges with mammalian cells, and each cell line has its own set of issues,” said Laura Geuss, a marketing specialist at GenScript. “We try to offer a variety of packages that can help customers who have difficult-to-work-with cells.” These packages include both viral-based and transient transfection techniques.
However, the most distinctive service offered by GenScript is its microbial genome-editing service for bacteria (Escherichia coli) and yeast (Saccharomyces cerevisiae). The company’s strategy for gene editing in bacteria can enable seamless knockins, knockouts, or gene replacements by combining CRISPR with lambda red recombineering. Traditionally one of the most effective methods for gene editing in microbes, recombineering allows editing without restriction enzymes through in vivo homologous recombination mediated by a phage-based recombination system such as lambda red.
On its own, lambda red technology cannot target multiple genes, but when paired with CRISPR, it allows the editing of multiple genes with greater efficiency than is possible with CRISPR alone, as the lambda red proteins help repair double-strand breaks in E. coli. The ability to knockout different gene combinations makes Genscript’s microbial editing service particularly well suited for the optimization of metabolic pathways.
Pooled and Arrayed Library Strategies
Scientists are using CRISPR technology for applications such as metabolic engineering and drug development. Yet another application area benefitting from CRISPR technology is cancer research. Here, the use of pooled CRISPR libraries is becoming commonplace. Pooled CRISPR libraries can help detect mutations that affect drug resistance, and they can aid in patient stratification and clinical trial design.
Pooled screening uses proliferation or viability as a phenotype to assess how genetic alterations, resulting from the application of a pooled CRISPR library, affect cell growth and death in the presence of a therapeutic compound. The enrichment or depletion of different gRNA populations is quantified using deep sequencing to identify the genomic edits that result in changes to cell viability.
MilliporeSigma provides pooled CRISPR libraries ranging from the whole human genome to smaller custom pools for these gene-function experiments. For pharmaceutical and biotech companies, Horizon Discovery offers a pooled screening service, ResponderSCREEN, which provides a whole-genome pooled screen to identify genes that confer sensitivity or resistance to a compound. This service is comprehensive, taking clients from experimental design all the way through to suggestions for follow-up studies.
Horizon Discovery maintains a Research Biotech business unit that is focused on target discovery and enabling translational medicine in oncology. “Our internal backbone gives us the ability to provide expert advice demonstrated by results,” said Jon Moore, Ph.D., the company’s CSO.
In contrast to a pooled screen, where thousands of gRNA are combined in one tube, an arrayed screen applies one gRNA per well, removing the need for deep sequencing and broadening the options for different endpoint assays. To establish and distribute a whole-genome arrayed lentiviral CRISPR library, MilliporeSigma partnered with the Wellcome Trust Sanger Institute. “This is the first and only arrayed CRISPR library in the world,” declared Shawn Shafer, Ph.D., functional genomics market segment manager, MilliporeSigma. “We were really proud to partner with Sanger on this.”
Pooled and arrayed screens are powerful tools for studying gene function. The appropriate platform for an experiment, however, will be determined by the desired endpoint assay.
The QX200 Droplet Digital PCR System from Bio-Rad Laboratories can provide researchers with an absolute measure of target DNA molecules for EvaGreen or probe-based digital PCR applications. The system, which can provide rapid, low-cost, ultra-sensitive quantification of both NHEJ- and HDR-editing events, consists of two instruments, the QX200 Droplet Generator and the QX200 Droplet Reader, and their associated consumables.
Finally, one last challenge for CRISPR lies in the detection and quantification of changes made to the genome post-editing. Conventional methods for detecting these alterations include gel methods and next-generation sequencing. While gel methods lack sensitivity and scalability, next-generation sequencing is costly and requires intensive bioinformatics.
To address this gap, Bio-Rad Laboratories developed a set of assay strategies to enable sensitive and precise edit detection with its Droplet Digital PCR (ddPCR) technology. The platform is designed to enable absolute quantification of nucleic acids with high sensitivity, high precision, and short turnaround time through massive droplet partitioning of samples.
Using a validated assay, a typical ddPCR experiment takes about five to six hours to complete. The ddPCR platform enables detection of rare mutations, and publications have reported detection of precise edits at a frequency of <0.05%, and of NHEJ-derived indels at a frequency as low as 0.1%. In addition to quantifying precise edits, indels, and computationally predicted off-target mutations, ddPCR can also be used to characterize the consequences of edits at the RNA level.
According to a recently published Science paper, the laboratory of Charles A. Gersbach, Ph.D., at Duke University used ddPCR in a study of muscle function in a mouse model of Duchenne muscular dystrophy. Specifically, ddPCR was used to assess the efficiency of CRISPR-Cas9 in removing the mutated exon 23 from the dystrophin gene. (Exon 23 deletion by CRISPR-Cas9 resulted in expression of the modified dystrophin gene and significant enhancement of muscle force.)
Quantitative ddPCR showed that exon 23 was deleted in ~2% of all alleles from the whole-muscle lysate. Further ddPCR studies found that 59% of mRNA transcripts reflected the deletion.
“There’s an overarching idea that the genome-editing field is moving extremely quickly, and for good reason,” asserted Jennifer Berman, Ph.D., staff scientist, Bio-Rad Laboratories. “There’s a lot of exciting work to be done, but detection and quantification of edits can be a bottleneck for researchers.”
The gene-editing field is moving quickly, and new innovations are finding their way into the laboratory as researchers lay the foundation for precise, well-controlled gene editing with CRISPR.
Researchers utilized a systems biology approach to develop new methods to assess drug sensitivity in cells. [The Institute for Systems Biology]
Understanding how cells respond and proliferate in the presence of anticancer compounds has been the foundation of drug discovery ideology for decades. Now, a new study from scientists at Vanderbilt University casts significant suspicion on the primary method used to test compounds for anticancer activity in cells—instilling doubt on methods employed by the entire scientific enterprise and pharmaceutical industry to discover new cancer drugs.
“More than 90% of candidate cancer drugs fail in late-stage clinical trials, costing hundreds of millions of dollars,” explained co-senior author Vito Quaranta, M.D., director of the Quantitative Systems Biology Center at Vanderbilt. “The flawed in vitro drug discovery metric may not be the only responsible factor, but it may be worth pursuing an estimate of its impact.”
The Vanderbilt investigators have developed what they believe to be a new metric for evaluating a compound’s effect on cell proliferation—called the DIP (drug-induced proliferation) rate—that overcomes the flawed bias in the traditional method.
The findings from this study were published recently in Nature Methods in an article entitled “An Unbiased Metric of Antiproliferative Drug Effect In Vitro.”
For more than three decades, researchers have evaluated the ability of a compound to kill cells by adding the compound in vitro and counting how many cells are alive after 72 hours. Yet, proliferation assays that measure cell number at a single time point don’t take into account the bias introduced by exponential cell proliferation, even in the presence of the drug.
“Cells are not uniform, they all proliferate exponentially, but at different rates,” Dr. Quaranta noted. “At 72 hours, some cells will have doubled three times and others will not have doubled at all.”
Dr. Quaranta added that drugs don’t all behave the same way on every cell line—for example, a drug might have an immediate effect on one cell line and a delayed effect on another.
The research team decided to take a systems biology approach, a mixture of experimentation and mathematical modeling, to demonstrate the time-dependent bias in static proliferation assays and to develop the time-independent DIP rate metric.
“Systems biology is what really makes the difference here,” Dr. Quaranta remarked. “It’s about understanding cells—and life—as dynamic systems.”This new study is of particular importance in light of recent international efforts to generate data sets that include the responses of thousands of cell lines to hundreds of compounds. Using the
Cancer Cell Line Encyclopedia (CCLE) and
Genomics of Drug Sensitivity in Cancer (GDSC) databases
will allow drug discovery scientists to include drug response data along with genomic and proteomic data that detail each cell line’s molecular makeup.
“The idea is to look for statistical correlations—these particular cell lines with this particular makeup are sensitive to these types of compounds—to use these large databases as discovery tools for new therapeutic targets in cancer,” Dr. Quaranta stated. “If the metric by which you’ve evaluated the drug sensitivity of the cells is wrong, your statistical correlations are basically no good.”
The Vanderbilt team evaluated the responses from four different melanoma cell lines to the drug vemurafenib, currently used to treat melanoma, with the standard metric—used for the CCLE and GDSC databases—and with the DIP rate. In one cell line, they found a glaring disagreement between the two metrics.
“The static metric says that the cell line is very sensitive to vemurafenib. However, our analysis shows this is not the case,” said co-lead study author Leonard Harris, Ph.D., a systems biology postdoctoral fellow at Vanderbilt. “A brief period of drug sensitivity, quickly followed by rebound, fools the static metric, but not the DIP rate.”
Dr. Quaranta added that the findings “suggest we should expect melanoma tumors treated with this drug to come back, and that’s what has happened, puzzling investigators. DIP rate analyses may help solve this conundrum, leading to better treatment strategies.”
The researchers noted that using the DIP rate is possible because of advances in automation, robotics, microscopy, and image processing. Moreover, the DIP rate metric offers another advantage—it can reveal which drugs are truly cytotoxic (cell killing), rather than merely cytostatic (cell growth inhibiting). Although cytostatic drugs may initially have promising therapeutic effects, they may leave tumor cells alive that then have the potential to cause the cancer to recur.
The Vanderbilt team is currently in the process of identifying commercial entities that can further refine the software and make it widely available to the research community to inform drug discovery.
An unbiased metric of antiproliferative drug effect in vitro
In vitro cell proliferation assays are widely used in pharmacology, molecular biology, and drug discovery. Using theoretical modeling and experimentation, we show that current metrics of antiproliferative small molecule effect suffer from time-dependent bias, leading to inaccurate assessments of parameters such as drug potency and efficacy. We propose the drug-induced proliferation (DIP) rate, the slope of the line on a plot of cell population doublings versus time, as an alternative, time-independent metric.
Researchers develop a technique to direct chromosome recombination with CRISPR/Cas9, allowing high-resolution genetic mapping of phenotypic traits in yeast.
Researchers used CRISPR/Cas9 to make a targeted double-strand break (DSB) in one arm of a yeast chromosome labeled with a green fluorescent protein (GFP) gene. A within-cell mechanism called homologous repair (HR) mends the broken arm using its homolog, resulting in a recombined region from the site of the break to the chromosome tip. When this cell divides by mitosis, each daughter cell will contain a homozygous section in an outcome known as “loss of heterozygosity” (LOH). One of the daughter cells is detectable because, due to complete loss of the GFP gene, it will no longer be fluorescent.REPRINTED WITH PERMISSION FROM M.J. SADHU ET AL., SCIENCE
When mapping phenotypic traits to specific loci, scientists typically rely on the natural recombination of chromosomes during meiotic cell division in order to infer the positions of responsible genes. But recombination events vary with species and chromosome region, giving researchers little control over which areas of the genome are shuffled. Now, a team at the University of California, Los Angeles (UCLA), has found a way around these problems by using CRISPR/Cas9 to direct targeted recombination events during mitotic cell division in yeast. The team described its technique today (May 5) in Science.
“Current methods rely on events that happen naturally during meiosis,” explained study coauthor Leonid Kruglyak of UCLA. “Whatever rate those events occur at, you’re kind of stuck with. Our idea was that using CRISPR, we can generate those events at will, exactly where we want them, in large numbers, and in a way that’s easy for us to pull out the cells in which they happened.”
Generally, researchers use coinheritance of a trait of interest with specific genetic markers—whose positions are known—to figure out what part of the genome is responsible for a given phenotype. But the procedure often requires impractically large numbers of progeny or generations to observe the few cases in which coinheritance happens to be disrupted informatively. What’s more, the resolution of mapping is limited by the length of the smallest sequence shuffled by recombination—and that sequence could include several genes or gene variants.
“Once you get down to that minimal region, you’re done,” said Kruglyak. “You need to switch to other methods to test every gene and every variant in that region, and that can be anywhere from challenging to impossible.”
But programmable, DNA-cutting champion CRISPR/Cas9 offered an alternative. During mitotic—rather than meiotic—cell division, rare, double-strand breaks in one arm of a chromosome preparing to split are sometimes repaired by a mechanism called homologous recombination. This mechanism uses the other chromosome in the homologous pair to replace the sequence from the break down to the end of the broken arm. Normally, such mitotic recombination happens so rarely as to be impractical for mapping purposes. With CRISPR/Cas9, however, the researchers found that they could direct double-strand breaks to any locus along a chromosome of interest (provided it was heterozygous—to ensure that only one of the chromosomes would be cut), thus controlling the sites of recombination.
Combining this technique with a signal of recombination success, such as a green fluorescent protein (GFP) gene at the tip of one chromosome in the pair, allowed the researchers to pick out cells in which recombination had occurred: if the technique failed, both daughter cells produced by mitotic division would be heterozygous, with one copy of the signal gene each. But if it succeeded, one cell would end up with two copies, and the other cell with none—an outcome called loss of heterozygosity.
“If we get loss of heterozygosity . . . half the cells derived after that loss of heterozygosity event won’t have GFP anymore,” study coauthor Meru Sadhu of UCLA explained. “We search for these cells that don’t have GFP out of the general population of cells.” If these non-fluorescent cells with loss of heterozygosity have the same phenotype as the parent for a trait of interest, then CRISPR/Cas9-targeted recombination missed the responsible gene. If the phenotype is affected, however, then the trait must be linked to a locus in the recombined, now-homozygous region, somewhere between the cut site and the GFP gene.
By systematically making cuts using CRISPR/Cas9 along chromosomes in a hybrid, diploid strain ofSaccharomyces cerevisiae yeast, picking out non-fluorescent cells, and then observing the phenotype, the UCLA team demonstrated that it could rapidly identify the phenotypic contribution of specific gene variants. “We can simply walk along the chromosome and at every [variant] position we can ask, does it matter for the trait we’re studying?” explained Kruglyak.
For example, the team showed that manganese sensitivity—a well-defined phenotypic trait in lab yeast—could be pinpointed using this method to a single nucleotide polymorphism (SNP) in a gene encoding the Pmr1 protein (a manganese transporter).
Jason Moffat, a molecular geneticist at the University of Toronto who was not involved in the work, toldThe Scientist that researchers had “dreamed about” exploiting these sorts of mechanisms for mapping purposes, but without CRISPR, such techniques were previously out of reach. Until now, “it hasn’t been so easy to actually make double-stranded breaks on one copy of a pair of chromosomes, and then follow loss of heterozygosity in mitosis,” he said, adding that he hopes to see the approach translated into human cell lines.
Applying the technique beyond yeast will be important, agreed cell and developmental biologist Ethan Bier of the University of California, San Diego, because chromosomal repair varies among organisms. “In yeast, they absolutely demonstrate the power of [this method],” he said. “We’ll just have to see how the technology develops in other systems that are going to be far less suited to the technology than yeast. . . . I would like to see it implemented in another system to show that they can get the same oomph out of it in, say, mammalian somatic cells.”
Kruglyak told The Scientist that work in higher organisms, though planned, is still in early stages; currently, his team is working to apply the technique to map loci responsible for trait differences between—rather than within—yeast species.
“We have a much poorer understanding of the differences across species,” Sadhu explained. “Except for a few specific examples, we’re pretty much in the dark there.”
Linkage and association studies have mapped thousands of genomic regions that contribute to phenotypic variation, but narrowing these regions to the underlying causal genes and variants has proven much more challenging. Resolution of genetic mapping is limited by the recombination rate. We developed a method that uses CRISPR to build mapping panels with targeted recombination events. We tested the method by generating a panel with recombination events spaced along a yeast chromosome arm, mapping trait variation, and then targeting a high density of recombination events to the region of interest. Using this approach, we fine-mapped manganese sensitivity to a single polymorphism in the transporter Pmr1. Targeting recombination events to regions of interest allows us to rapidly and systematically identify causal variants underlying trait differences.
Thank you, David, for the kind words and comments. We agree that the most immediate applications of the CRISPR-based recombination mapping will be in unicellular organisms and cell culture. We also think the method holds a lot of promise for research in multicellular organisms, although we did not mean to imply that it “will be an efficient mapping method for all multicellular organisms”. Every organism will have its own set of constraints as well as experimental tools that will be relevant when adapting a new technique. To best help experts working on these organisms, here are our thoughts on your questions.
You asked about mutagenesis during recombination. We Sanger sequenced 72 of our LOH lines at the recombination site and did not observe any mutations, as described in the supplementary materials. We expect the absence of mutagenesis is because we targeted heterozygous sites where the untargeted allele did not have a usable PAM site; thus, following LOH, the targeted site is no longer present and cutting stops. In your experiments you targeted sites that were homozygous; thus, following recombination, the CRISPR target site persisted, and continued cutting ultimately led to repair by NHEJ and mutagenesis.
As to the more general question of the optimal mapping strategies in different organisms, they will depend on the ease of generating and screening for editing events, the cost and logistics of maintaining and typing many lines, and generation time, among other factors. It sounds like in Drosophila today, your related approach of generating markers with CRISPR, and then enriching for natural recombination events that separate them, is preferable. In yeast, we’ve found the opposite to be the case. As you note, even in Drosophila, our approach may be preferable for regions with low or highly non-uniform recombination rates.
Finally, mapping in sterile interspecies hybrids should be straightforward for unicellular hybrids (of which there are many examples) and for cells cultured from hybrid animals or plants. For studies in hybrid multicellular organisms, we agree that driving mitotic recombination in the early embryo may be the most promising approach. Chimeric individuals with mitotic clones will be sufficient for many traits. Depending on the system, it may in fact be possible to generate diploid individuals with uniform LOH genotype, but this is certainly beyond the scope of our paper. The calculation of the number of lines assumes that the mapping is done in a single step; as you note in your earlier comment, mapping sequentially can reduce this number dramatically.
This is a lovely method and should find wide applicability in many settings, especially for microorganisms and cell lines. However, it is not clear that this approach will be, as implied by the discussion, an efficient mapping method for all multicellular organisms. I have performed similar experiments in Drosophila, focused on meiotic recombination, on a much smaller scale, and found that CRISPR-Cas9 can indeed generate targeted recombination at gRNA target sites. In every case I tested, I found that the recombination event was associated with a deletion at the gRNA site, which is probably unimportant for most mapping efforts, but may be a concern in some specific cases, for example for clinical applications. It would be interesting to know how often mutations occurred at the targeted gRNA site in this study.
The wider issue, however, is whether CRISPR-mediated recombination will be more efficient than other methods of mapping. After careful consideration of all the costs and the time involved in each of the steps for Drosophila, we have decided that targeted meiotic recombination using flanking visible markers will be, in most cases, considerably more efficient than CRISPR-mediated recombination. This is mainly due to the large expense of injecting embryos and the extensive effort and time required to screen injected animals for appropriate events. It is both cheaper and faster to generate markers (with CRISPR) and then perform a large meiotic recombination mapping experiment than it would be to generate the lines required for CRISPR-mediated recombination mapping. It is possible to dramatically reduce costs by, for example, mapping sequentially at finer resolution. But this approach would require much more time than marker-assisted mapping. If someone develops a rapid and cheap method of reliably introducing DNA into Drosophila embryos, then this calculus might change.
However, it is possible to imagine situations where CRISPR-mediated mapping would be preferable, even for Drosophila. For example, some genomic regions display extremely low or highly non-uniform recombination rates. It is possible that CRISPR-mediated mapping could provide a reasonable approach to fine mapping genes in these regions.
The authors also propose the exciting possibility that CRISPR-mediated loss of heterozygosity could be used to map traits in sterile species hybrids. It is not entirely obvious to me how this experiment would proceed and I hope the authors can illuminate me. If we imagine driving a recombination event in the early embryo (with maternal Cas9 from one parent and gRNA from a second parent), then at best we would end up with chimeric individuals carrying mitotic clones. I don’t think one could generate diploid animals where all cells carried the same loss of heterozygosity event. Even if we could, this experiment would require construction of a substantial number of stable transgenic lines expressing gRNAs. Mapping an ~20Mbp chromosome arm to ~10kb would require on the order of two-thousand transgenic lines. Not an undertaking to be taken lightly. It is already possible to perform similar tests (hemizygosity tests) using D. melanogaster deficiency lines in crosses with D. simulans, so perhaps CRISPR-mediated LOH could complement these deficiency screens for fine mapping efforts. But, at the moment, it is not clear to me how to do the experiment.
CRISPR-Cas9 Screening by Horizon Discovery, Cambridge, UK – HDx™ Reference Standards, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 2: CRISPR for Gene Editing and DNA Repair
CRISPR-Cas9 Screening by Horizon Discovery, Cambridge, UK – HDx™ Reference Standards
Reporters: David Orchard-Webb, PhD and Aviva Lev-Ari, PhD, RN
They have leveraged our cutting-edge technologies to build a ground-breaking platform for Immuno-Oncology drug discovery.
Their collection of high throughput assays allows you to rapidly identify single agents or combinations that affect T cell and natural killer (NK) cell function.
Gene-Editing in Immune Cells
Gene-editing in immune cells for your Immuno-Oncology project is now even faster than before. Leveraging the latest genome engineering tools, including CRISPR-Cas9, we’re able to precisely edit over 10 pre-characterized T-cell, B-cell, and monocytes lines.
Power Your Immune-Oncology Projects with Custom Engineered Immune Cells
Gene-editing in immune cell lines can be challenging due to low targeting efficiency and difficulties in single cell derivation of suspension cells. Horizon has validated 10+ immune cell lines including THP-1, Jurkat and NALM-6 cells for gene-editing projects. CRISPR, rAAV and ZFN gene-editing technologies are available depending on project requirements.
Take advantage of the largest panel of pre-characterized immune cell lines available and benefit from Horizon’s exceptional knowhow and experience in completing over 2,000 gene-editing projects.
CRISPR/Cas9, Familial Amyloid Polyneuropathy (FAP) and Neurodegenerative Disease, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 2: CRISPR for Gene Editing and DNA Repair
CRISPR/Cas9, Familial Amyloid Polyneuropathy ( FAP) and Neurodegenerative Disease
Curator: Larry H. Bernstein, MD, FCAP
CRISPR/Cas9 and Targeted Genome Editing: A New Era in Molecular Biology
The development of efficient and reliable ways to make precise, targeted changes to the genome of living cells is a long-standing goal for biomedical researchers. Recently, a new tool based on a bacterial CRISPR-associated protein-9 nuclease (Cas9) from Streptococcus pyogenes has generated considerable excitement (1). This follows several attempts over the years to manipulate gene function, including homologous recombination (2) and RNA interference (RNAi) (3). RNAi, in particular, became a laboratory staple enabling inexpensive and high-throughput interrogation of gene function (4, 5), but it is hampered by providing only temporary inhibition of gene function and unpredictable off-target effects (6). Other recent approaches to targeted genome modification – zinc-finger nucleases [ZFNs, (7)] and transcription-activator like effector nucleases [TALENs (8)]– enable researchers to generate permanent mutations by introducing doublestranded breaks to activate repair pathways. These approaches are costly and time-consuming to engineer, limiting their widespread use, particularly for large scale, high-throughput studies.
The Biology of Cas9
The functions of CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and CRISPR-associated (Cas) genes are essential in adaptive immunity in select bacteria and archaea, enabling the organisms to respond to and eliminate invading genetic material. These repeats were initially discovered in the 1980s in E. coli (9), but their function wasn’t confirmed until 2007 by Barrangou and colleagues, who demonstrated that S. thermophilus can acquire resistance against a bacteriophage by integrating a genome fragment of an infectious virus into its CRISPR locus (10).
Three types of CRISPR mechanisms have been identified, of which type II is the most studied. In this case, invading DNA from viruses or plasmids is cut into small fragments and incorporated into a CRISPR locus amidst a series of short repeats (around 20 bps). The loci are transcribed, and transcripts are then processed to generate small RNAs (crRNA – CRISPR RNA), which are used to guide effector endonucleases that target invading DNA based on sequence complementarity (Figure 1) (11).
Figure 1. Cas9 in vivo: Bacterial Adaptive Immunity
In the acquisition phase, foreign DNA is incorporated into the bacterial genome at the CRISPR loci. CRISPR loci is then transcribed and processed into crRNA during crRNA biogenesis. During interference, Cas9 endonuclease complexed with a crRNA and separate tracrRNA cleaves foreign DNA containing a 20-nucleotide crRNA complementary sequence adjacent to the PAM sequence. (Figure not drawn to scale.)
One Cas protein, Cas9 (also known as Csn1), has been shown, through knockdown and rescue experiments to be a key player in certain CRISPR mechanisms (specifically type II CRISPR systems). The type II CRISPR mechanism is unique compared to other CRISPR systems, as only one Cas protein (Cas9) is required for gene silencing (12). In type II systems, Cas9 participates in the processing of crRNAs (12), and is responsible for the destruction of the target DNA (11). Cas9’s function in both of these steps relies on the presence of two nuclease domains, a RuvC-like nuclease domain located at the amino terminus and a HNH-like nuclease domain that resides in the mid-region of the protein (13).
To achieve site-specific DNA recognition and cleavage, Cas9 must be complexed with both a crRNA and a separate trans-activating crRNA (tracrRNA or trRNA), that is partially complementary to the crRNA (11). The tracrRNA is required for crRNA maturation from a primary transcript encoding multiple pre-crRNAs. This occurs in the presence of RNase III and Cas9 (12).
During the destruction of target DNA, the HNH and RuvC-like nuclease domains cut both DNA strands, generating double-stranded breaks (DSBs) at sites defined by a 20-nucleotide target sequence within an associated crRNA transcript (11, 14). The HNH domain cleaves the complementary strand, while the RuvC domain cleaves the noncomplementary strand.
The double-stranded endonuclease activity of Cas9 also requires that a short conserved sequence, (2–5 nts) known as protospacer-associated motif (PAM), follows immediately 3´- of the crRNA complementary sequence (15). In fact, even fully complementary sequences are ignored by Cas9-RNA in the absence of a PAM sequence (16).
Cas9 and CRISPR as a New Tool in Molecular Biology
The simplicity of the type II CRISPR nuclease, with only three required components (Cas9 along with the crRNA and trRNA) makes this system amenable to adaptation for genome editing. This potential was realized in 2012 by the Doudna and Charpentier labs (11). Based on the type II CRISPR system described previously, the authors developed a simplified two-component system by combining trRNA and crRNA into a single synthetic single guide RNA (sgRNA). sgRNAprogrammed Cas9 was shown to be as effective as Cas9 programmed with separate trRNA and crRNA in guiding targeted gene alterations (Figure 2A).
To date, three different variants of the Cas9 nuclease have been adopted in genome-editing protocols. The first is wild-type Cas9, which can site-specifically cleave double-stranded DNA, resulting in the activation of the doublestrand break (DSB) repair machinery. DSBs can be repaired by the cellular Non-Homologous End Joining (NHEJ) pathway (17), resulting in insertions and/or deletions (indels) which disrupt the targeted locus. Alternatively, if a donor template with homology to the targeted locus is supplied, the DSB may be repaired by the homology-directed repair (HDR) pathway allowing for precise replacement mutations to be made (Figure 2A) (17, 18).
Cong and colleagues (1) took the Cas9 system a step further towards increased precision by developing a mutant form, known as Cas9D10A, with only nickase activity. This means it cleaves only one DNA strand, and does not activate NHEJ. Instead, when provided with a homologous repair template, DNA repairs are conducted via the high-fidelity HDR pathway only, resulting in reduced indel mutations (1, 11, 19). Cas9D10A is even more appealing in terms of target specificity when loci are targeted by paired Cas9 complexes designed to generate adjacent DNA nicks (20) (see further details about “paired nickases” in Figure 2B).
The third variant is a nuclease-deficient Cas9 (dCas9, Figure 2C) (21). Mutations H840A in the HNH domain and D10A in the RuvC domain inactivate cleavage activity, but do not prevent DNA binding (11, 22). Therefore, this variant can be used to sequence-specifically target any region of the genome without cleavage. Instead, by fusing with various effector domains, dCas9 can be used either as a gene silencing or activation tool (21, 23–26). Furthermore, it can be used as a visualization tool. For instance, Chen and colleagues used dCas9 fused to Enhanced Green Fluorescent Protein (EGFP) to visualize repetitive DNA sequences with a single sgRNA or nonrepetitive loci using multiple sgRNAs (27).
Wild-type Cas9 nuclease site specifically cleaves double-stranded DNA activating double-strand break repair machinery. In the absence of a homologous repair template non-homologous end joining can result in indels disrupting the target sequence. Alternatively, precise mutations and knock-ins can be made by providing a homologous repair template and exploiting the homology directed repair pathway.
B. Mutated Cas9 makes a site specific single-strand nick. Two sgRNA can be used to introduce a staggered double-stranded break which can then undergo homology directed repair.
C. Nuclease-deficient Cas9 can be fused with various effector domains allowing specific localization. For example, transcriptional activators, repressors, and fluorescent proteins.
Targeting Efficiency and Off-target Mutations
Targeting efficiency, or the percentage of desired mutation achieved, is one of the most important parameters by which to assess a genome-editing tool. The targeting efficiency of Cas9 compares favorably with more established methods, such as TALENs or ZFNs (8). For example, in human cells, custom-designed ZFNs and TALENs could only achieve efficiencies ranging from 1% to 50% (29–31). In contrast, the Cas9 system has been reported to have efficiencies up to >70% in zebrafish (32) and plants (33), and ranging from 2–5% in induced pluripotent stem cells (34). In addition, Zhou and colleagues were able to improve genome targeting up to 78% in one-cell mouse embryos, and achieved effective germline transmission through the use of dual sgRNAs to simultaneously target an individual gene (35).
A widely used method to identify mutations is the T7 Endonuclease I mutation detection assay (36, 37) (Figure 3). This assay detects heteroduplex DNA that results from the annealing of a DNA strand, including desired mutations, with a wildtype DNA strand (37).
Figure 3. T7 Endonuclease I Targeting Efficiency Assay
Genomic DNA is amplified with primers bracketing the modified locus. PCR products are then denatured and re-annealed yielding 3 possible structures. Duplexes containing a mismatch are digested by T7 Endonuclease I. The DNA is then electrophoretically separated and fragment analysis is used to calculate targeting efficiency.
Another important parameter is the incidence of off-target mutations. Such mutations are likely to appear in sites that have differences of only a few nucleotides compared to the original sequence, as long as they are adjacent to a PAM sequence. This occurs as Cas9 can tolerate up to 5 base mismatches within the protospacer region (36) or a single base difference in the PAM sequence (38). Off-target mutations are generally more difficult to detect, requiring whole-genome sequencing to rule them out completely.
Recent improvements to the CRISPR system for reducing off-target mutations have been made through the use of truncated gRNA (truncated within the crRNA-derived sequence) or by adding two extra guanine (G) nucleotides to the 5´ end (28, 37). Another way researchers have attempted to minimize off-target effects is with the use of “paired nickases” (20). This strategy uses D10A Cas9 and two sgRNAs complementary to the adjacent area on opposite strands of the target site (Figure 2B). While this induces DSBs in the target DNA, it is expected to create only single nicks in off-target locations and, therefore, result in minimal off-target mutations.
By leveraging computation to reduce off-target mutations, several groups have developed webbased tools to facilitate the identification of potential CRISPR target sites and assess their potential for off-target cleavage. Examples include the CRISPR Design Tool (38) and the ZiFiT Targeter, Version 4.2 (39, 40).
Applications as a Genome-editing and Genome Targeting Tool
Following its initial demonstration in 2012 (9), the CRISPR/Cas9 system has been widely adopted. This has already been successfully used to target important genes in many cell lines and organisms, including human (34), bacteria (41), zebrafish (32), C. elegans (42), plants (34), Xenopus tropicalis (43), yeast (44), Drosophila (45), monkeys (46), rabbits (47), pigs (42), rats (48) and mice (49). Several groups have now taken advantage of this method to introduce single point mutations (deletions or insertions) in a particular target gene, via a single gRNA (14, 21, 29). Using a pair of gRNA-directed Cas9 nucleases instead, it is also possible to induce large deletions or genomic rearrangements, such as inversions or translocations (50). A recent exciting development is the use of the dCas9 version of the CRISPR/Cas9 system to target protein domains for transcriptional regulation (26, 51, 52), epigenetic modification (25), and microscopic visualization of specific genome loci (27).
The CRISPR/Cas9 system requires only the redesign of the crRNA to change target specificity. This contrasts with other genome editing tools, including zinc finger and TALENs, where redesign of the protein-DNA interface is required. Furthermore, CRISPR/Cas9 enables rapid genome-wide interrogation of gene function by generating large gRNA libraries (51, 53) for genomic screening.
The Future of CRISPR/Cas9
The rapid progress in developing Cas9 into a set of tools for cell and molecular biology research has been remarkable, likely due to the simplicity, high efficiency and versatility of the system. Of the designer nuclease systems currently available for precision genome engineering, the CRISPR/Cas system is by far the most user friendly. It is now also clear that Cas9’s potential reaches beyond DNA cleavage, and its usefulness for genome locus-specific recruitment of proteins will likely only be limited by our imagination.
Scientists urge caution in using new CRISPR technology to treat human genetic disease
The bacterial enzyme Cas9 is the engine of RNA-programmed genome engineering in human cells. (Graphic by Jennifer Doudna/UC Berkeley)
A group of 18 scientists and ethicists today warned that a revolutionary new tool to cut and splice DNA should be used cautiously when attempting to fix human genetic disease, and strongly discouraged any attempts at making changes to the human genome that could be passed on to offspring.
Among the authors of this warning is Jennifer Doudna, the co-inventor of the technology, called CRISPR-Cas9, which is driving a new interest in gene therapy, or “genome engineering.” She and colleagues co-authored a perspective piece that appears in the March 20 issue of Science, based on discussions at a meeting that took place in Napa on Jan. 24. The same issue of Science features a collection of recent research papers, commentary and news articles on CRISPR and its implications. …..
A prudent path forward for genomic engineering and germline gene modification
Scientists today are changing DNA sequences to correct genetic defects in animals as well as cultured tissues generated from stem cells, strategies that could eventually be used to treat human disease. The technology can also be used to engineer animals with genetic diseases mimicking human disease, which could lead to new insights into previously enigmatic disorders.
The CRISPR-Cas9 tool is still being refined to ensure that genetic changes are precisely targeted, Doudna said. Nevertheless, the authors met “… to initiate an informed discussion of the uses of genome engineering technology, and to identify proactively those areas where current action is essential to prepare for future developments. We recommend taking immediate steps toward ensuring that the application of genome engineering technology is performed safely and ethically.”
Amyloid CRISPR Plasmids and si/shRNA Gene Silencers
Santa Cruz Biotechnology, Inc. offers a broad range of gene silencers in the form of siRNAs, shRNA Plasmids and shRNA Lentiviral Particles as well as CRISPR/Cas9 Knockout and CRISPR Double Nickase plasmids. Amyloid gene silencers are available as Amyloid siRNA, Amyloid shRNA Plasmid, Amyloid shRNA Lentiviral Particles and Amyloid CRISPR/Cas9 Knockout plasmids. Amyloid CRISPR/dCas9 Activation Plasmids and CRISPR Lenti Activation Systems for gene activation are also available. Gene silencers and activators are useful for gene studies in combination with antibodies used for protein detection. Amyloid CRISPR Knockout, HDR and Nickase Knockout Plasmids
CRISPR-Cas9-Based Knockout of the Prion Protein and Its Effect on the Proteome
The molecular function of the cellular prion protein (PrPC) and the mechanism by which it may contribute to neurotoxicity in prion diseases and Alzheimer’s disease are only partially understood. Mouse neuroblastoma Neuro2a cells and, more recently, C2C12 myocytes and myotubes have emerged as popular models for investigating the cellular biology of PrP. Mouse epithelial NMuMG cells might become attractive models for studying the possible involvement of PrP in a morphogenetic program underlying epithelial-to-mesenchymal transitions. Here we describe the generation of PrP knockout clones from these cell lines using CRISPR-Cas9 knockout technology. More specifically, knockout clones were generated with two separate guide RNAs targeting recognition sites on opposite strands within the first hundred nucleotides of the Prnp coding sequence. Several PrP knockout clones were isolated and genomic insertions and deletions near the CRISPR-target sites were characterized. Subsequently, deep quantitative global proteome analyses that recorded the relative abundance of>3000 proteins (data deposited to ProteomeXchange Consortium) were undertaken to begin to characterize the molecular consequences of PrP deficiency. The levels of ∼120 proteins were shown to reproducibly correlate with the presence or absence of PrP, with most of these proteins belonging to extracellular components, cell junctions or the cytoskeleton.
Recent advances in genome engineering technologies based on the CRISPR-associated RNA-guided endonuclease Cas9 are enabling the systematic interrogation of mammalian genome function. Analogous to the search function in modern word processors, Cas9 can be guided to specific locations within complex genomes by a short RNA search string. Using this system, DNA sequences within the endogenous genome and their functional outputs are now easily edited or modulated in virtually any organism of choice. Cas9-mediated genetic perturbation is simple and scalable, empowering researchers to elucidate the functional organization of the genome at the systems level and establish causal linkages between genetic variations and biological phenotypes. In this Review, we describe the development and applications of Cas9 for a variety of research or translational applications while highlighting challenges as well as future directions. Derived from a remarkable microbial defense system, Cas9 is driving innovative applications from basic biology to biotechnology and medicine.
The development of recombinant DNA technology in the 1970s marked the beginning of a new era for biology. For the first time, molecular biologists gained the ability to manipulate DNA molecules, making it possible to study genes and harness them to develop novel medicine and biotechnology. Recent advances in genome engineering technologies are sparking a new revolution in biological research. Rather than studying DNA taken out of the context of the genome, researchers can now directly edit or modulate the function of DNA sequences in their endogenous context in virtually any organism of choice, enabling them to elucidate the functional organization of the genome at the systems level, as well as identify causal genetic variations.
Broadly speaking, genome engineering refers to the process of making targeted modifications to the genome, its contexts (e.g., epigenetic marks), or its outputs (e.g., transcripts). The ability to do so easily and efficiently in eukaryotic and especially mammalian cells holds immense promise to transform basic science, biotechnology, and medicine (Figure 1).
For life sciences research, technologies that can delete, insert, and modify the DNA sequences of cells or organisms enable dissecting the function of specific genes and regulatory elements. Multiplexed editing could further allow the interrogation of gene or protein networks at a larger scale. Similarly, manipulating transcriptional regulation or chromatin states at particular loci can reveal how genetic material is organized and utilized within a cell, illuminating relationships between the architecture of the genome and its functions. In biotechnology, precise manipulation of genetic building blocks and regulatory machinery also facilitates the reverse engineering or reconstruction of useful biological systems, for example, by enhancing biofuel production pathways in industrially relevant organisms or by creating infection-resistant crops. Additionally, genome engineering is stimulating a new generation of drug development processes and medical therapeutics. Perturbation of multiple genes simultaneously could model the additive effects that underlie complex polygenic disorders, leading to new drug targets, while genome editing could directly correct harmful mutations in the context of human gene therapy (Tebas et al., 2014).
Eukaryotic genomes contain billions of DNA bases and are difficult to manipulate. One of the breakthroughs in genome manipulation has been the development of gene targeting by homologous recombination (HR), which integrates exogenous repair templates that contain sequence homology to the donor site (Figure 2A) (Capecchi, 1989). HR-mediated targeting has facilitated the generation of knockin and knockout animal models via manipulation of germline competent stem cells, dramatically advancing many areas of biological research. However, although HR-mediated gene targeting produces highly precise alterations, the desired recombination events occur extremely infrequently (1 in 106–109 cells) (Capecchi, 1989), presenting enormous challenges for large-scale applications of gene-targeting experiments.
Genome Editing Technologies Exploit Endogenous DNA Repair Machinery
To overcome these challenges, a series of programmable nuclease-based genome editing technologies have been developed in recent years, enabling targeted and efficient modification of a variety of eukaryotic and particularly mammalian species. Of the current generation of genome editing technologies, the most rapidly developing is the class of RNA-guided endonucleases known as Cas9 from the microbial adaptive immune system CRISPR (clustered regularly interspaced short palindromic repeats), which can be easily targeted to virtually any genomic location of choice by a short RNA guide. Here, we review the development and applications of the CRISPR-associated endonuclease Cas9 as a platform technology for achieving targeted perturbation of endogenous genomic elements and also discuss challenges and future avenues for innovation. ……
Figure 4Natural Mechanisms of Microbial CRISPR Systems in Adaptive Immunity
…… A key turning point came in 2005, when systematic analysis of the spacer sequences separating the individual direct repeats suggested their extrachromosomal and phage-associated origins (Mojica et al., 2005; Pourcel et al., 2005; Bolotin et al., 2005). This insight was tremendously exciting, especially given previous studies showing that CRISPR loci are transcribed (Tang et al., 2002) and that viruses are unable to infect archaeal cells carrying spacers corresponding to their own genomes (Mojica et al., 2005). Together, these findings led to the speculation that CRISPR arrays serve as an immune memory and defense mechanism, and individual spacers facilitate defense against bacteriophage infection by exploiting Watson-Crick base-pairing between nucleic acids (Mojica et al., 2005; Pourcel et al., 2005). Despite these compelling realizations that CRISPR loci might be involved in microbial immunity, the specific mechanism of how the spacers act to mediate viral defense remained a challenging puzzle. Several hypotheses were raised, including thoughts that CRISPR spacers act as small RNA guides to degrade viral transcripts in a RNAi-like mechanism (Makarova et al., 2006) or that CRISPR spacers direct Cas enzymes to cleave viral DNA at spacer-matching regions (Bolotin et al., 2005). …..
As the pace of CRISPR research accelerated, researchers quickly unraveled many details of each type of CRISPR system (Figure 4). Building on an earlier speculation that protospacer adjacent motifs (PAMs) may direct the type II Cas9 nuclease to cleave DNA (Bolotin et al., 2005), Moineau and colleagues highlighted the importance of PAM sequences by demonstrating that PAM mutations in phage genomes circumvented CRISPR interference (Deveau et al., 2008). Additionally, for types I and II, the lack of PAM within the direct repeat sequence within the CRISPR array prevents self-targeting by the CRISPR system. In type III systems, however, mismatches between the 5′ end of the crRNA and the DNA target are required for plasmid interference (Marraffini and Sontheimer, 2010). …..
In 2013, a pair of studies simultaneously showed how to successfully engineer type II CRISPR systems from Streptococcus thermophilus (Cong et al., 2013) andStreptococcus pyogenes (Cong et al., 2013; Mali et al., 2013a) to accomplish genome editing in mammalian cells. Heterologous expression of mature crRNA-tracrRNA hybrids (Cong et al., 2013) as well as sgRNAs (Cong et al., 2013; Mali et al., 2013a) directs Cas9 cleavage within the mammalian cellular genome to stimulate NHEJ or HDR-mediated genome editing. Multiple guide RNAs can also be used to target several genes at once. Since these initial studies, Cas9 has been used by thousands of laboratories for genome editing applications in a variety of experimental model systems (Sander and Joung, 2014). ……
The majority of CRISPR-based technology development has focused on the signature Cas9 nuclease from type II CRISPR systems. However, there remains a wide diversity of CRISPR types and functions. Cas RAMP module (Cmr) proteins identified in Pyrococcus furiosus and Sulfolobus solfataricus (Hale et al., 2012) constitute an RNA-targeting CRISPR immune system, forming a complex guided by small CRISPR RNAs that target and cleave complementary RNA instead of DNA. Cmr protein homologs can be found throughout bacteria and archaea, typically relying on a 5′ site tag sequence on the target-matching crRNA for Cmr-directed cleavage.
Unlike RNAi, which is targeted largely by a 6 nt seed region and to a lesser extent 13 other bases, Cmr crRNAs contain 30–40 nt of target complementarity. Cmr-CRISPR technologies for RNA targeting are thus a promising target for orthogonal engineering and minimal off-target modification. Although the modularity of Cmr systems for RNA-targeting in mammalian cells remains to be investigated, Cmr complexes native to P. furiosus have already been engineered to target novel RNA substrates (Hale et al., 2009, 2012). ……
Although Cas9 has already been widely used as a research tool, a particularly exciting future direction is the development of Cas9 as a therapeutic technology for treating genetic disorders. For a monogenic recessive disorder due to loss-of-function mutations (such as cystic fibrosis, sickle-cell anemia, or Duchenne muscular dystrophy), Cas9 may be used to correct the causative mutation. This has many advantages over traditional methods of gene augmentation that deliver functional genetic copies via viral vector-mediated overexpression—particularly that the newly functional gene is expressed in its natural context. For dominant-negative disorders in which the affected gene is haplosufficient (such as transthyretin-related hereditary amyloidosis or dominant forms of retinitis pigmentosum), it may also be possible to use NHEJ to inactivate the mutated allele to achieve therapeutic benefit. For allele-specific targeting, one could design guide RNAs capable of distinguishing between single-nucleotide polymorphism (SNP) variations in the target gene, such as when the SNP falls within the PAM sequence.
CRISPR/Cas9: a powerful genetic engineering tool for establishing large animal models of neurodegenerative diseases
Zhuchi Tu, Weili Yang, Sen Yan, Xiangyu Guo and Xiao-Jiang Li
Animal models are extremely valuable to help us understand the pathogenesis of neurodegenerative disorders and to find treatments for them. Since large animals are more like humans than rodents, they make good models to identify the important pathological events that may be seen in humans but not in small animals; large animals are also very important for validating effective treatments or confirming therapeutic targets. Due to the lack of embryonic stem cell lines from large animals, it has been difficult to use traditional gene targeting technology to establish large animal models of neurodegenerative diseases. Recently, CRISPR/Cas9 was used successfully to genetically modify genomes in various species. Here we discuss the use of CRISPR/Cas9 technology to establish large animal models that can more faithfully mimic human neurodegenerative diseases.
Neurodegenerative diseases — Alzheimer’s disease(AD),Parkinson’s disease(PD), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), and frontotemporal dementia (FTD) — are characterized by age-dependent and selective neurodegeneration. As the life expectancy of humans lengthens, there is a greater prevalence of these neurodegenerative diseases; however, the pathogenesis of most of these neurodegenerative diseases remain unclear, and we lack effective treatments for these important brain disorders.
CRISPR/Cas9, Non-human primates, Neurodegenerative diseases, Animal model
There are a number of excellent reviews covering different types of neurodegenerative diseases and their genetic mouse models [8–12]. Investigations of different mouse models of neurodegenerative diseases have revealed a common pathology shared by these diseases. First, the development of neuropathology and neurological symptoms in genetic mouse models of neurodegenerative diseases is age dependent and progressive. Second, all the mouse models show an accumulation of misfolded or aggregated proteins resulting from the expression of mutant genes. Third, despite the widespread expression of mutant proteins throughout the body and brain, neuronal function appears to be selectively or preferentially affected. All these facts indicate that mouse models of neurodegenerative diseases recapitulate important pathologic features also seen in patients with neurodegenerative diseases.
However, it seems that mouse models can not recapitulate the full range of neuropathology seen in patients with neurodegenerative diseases. Overt neurodegeneration, which is the most important pathological feature in patient brains, is absent in genetic rodent models of AD, PD, and HD. Many rodent models that express transgenic mutant proteins under the control of different promoters do not replicate overt neurodegeneration, which is likely due to their short life spans and the different aging processes of small animals. Also important are the remarkable differences in brain development between rodents and primates. For example, the mouse brain takes 21 days to fully develop, whereas the formation of primate brains requires more than 150 days [13]. The rapid development of the brain in rodents may render neuronal cells resistant to misfolded protein-mediated neurodegeneration. Another difficulty in using rodent models is how to analyze cognitive and emotional abnormalities, which are the early symptoms of most neurodegenerative diseases in humans. Differences in neuronal circuitry, anatomy, and physiology between rodent and primate brains may also account for the behavioral differences between rodent and primate models.
Mitochondrial dynamics–fusion, fission, movement, and mitophagy–in neurodegenerative diseases
Neurons are metabolically active cells with high energy demands at locations distant from the cell body. As a result, these cells are particularly dependent on mitochondrial function, as reflected by the observation that diseases of mitochondrial dysfunction often have a neurodegenerative component. Recent discoveries have highlighted that neurons are reliant particularly on the dynamic properties of mitochondria. Mitochondria are dynamic organelles by several criteria. They engage in repeated cycles of fusion and fission, which serve to intermix the lipids and contents of a population of mitochondria. In addition, mitochondria are actively recruited to subcellular sites, such as the axonal and dendritic processes of neurons. Finally, the quality of a mitochondrial population is maintained through mitophagy, a form of autophagy in which defective mitochondria are selectively degraded. We review the general features of mitochondrial dynamics, incorporating recent findings on mitochondrial fusion, fission, transport and mitophagy. Defects in these key features are associated with neurodegenerative disease. Charcot-Marie-Tooth type 2A, a peripheral neuropathy, and dominant optic atrophy, an inherited optic neuropathy, result from a primary deficiency of mitochondrial fusion. Moreover, several major neurodegenerative diseases—including Parkinson’s, Alzheimer’s and Huntington’s disease—involve disruption of mitochondrial dynamics. Remarkably, in several disease models, the manipulation of mitochondrial fusion or fission can partially rescue disease phenotypes. We review how mitochondrial dynamics is altered in these neurodegenerative diseases and discuss the reciprocal interactions between mitochondrial fusion, fission, transport and mitophagy.
Applications of CRISPR–Cas systems in Neuroscience
Genome-editing tools, and in particular those based on CRISPR–Cas (clustered regularly interspaced short palindromic repeat (CRISPR)–CRISPR-associated protein) systems, are accelerating the pace of biological research and enabling targeted genetic interrogation in almost any organism and cell type. These tools have opened the door to the development of new model systems for studying the complexity of the nervous system, including animal models and stem cell-derived in vitro models. Precise and efficient gene editing using CRISPR–Cas systems has the potential to advance both basic and translational neuroscience research.
Cellular neuroscience, DNA recombination, Genetic engineering, Molecular neuroscience
Figure 3: In vitro applications of Cas9 in human iPSCs.close
a | Evaluation of disease candidate genes from large-population genome-wide association studies (GWASs). Human primary cells, such as neurons, are not easily available and are difficult to expand in culture. By contrast, induced pluripo…
The development of the CRISPR/Cas9 system has made gene editing a relatively simple task. While CRISPR and other gene editing technologies stand to revolutionize biomedical research and offers many promising therapeutic avenues (such as in the treatment of HIV), a great deal of debate exists over whether CRISPR should be used to modify human embryos. As I discussed in my previous Insight article, we lack enough fundamental biological knowledge to enhance many traits like height or intelligence, so we are not near a future with genetically-enhanced super babies. However, scientists have identified a few rare genetic variants that protect against disease. One such protective variant is a mutation in the APP gene that protects against Alzheimer’s disease and cognitive decline in old age. If we can perfect gene editing technologies, is this mutation one that we should be regularly introducing into embryos? In this article, I explore the potential for using gene editing as a way to prevent Alzheimer’s disease in future generations. Alzheimer’s Disease: Medicine’s Greatest Challenge in the 21st Century Can gene editing be the missing piece in the battle against Alzheimer’s? (Source: bostonbiotech.org) I chose to assess the benefit of germline gene editing in the context of Alzheimer’s disease because this disease is one of the biggest challenges medicine faces in the 21st century. Alzheimer’s disease is a chronic neurodegenerative disease responsible for the majority of the cases of dementia in the elderly. The disease symptoms begins with short term memory loss and causes more severe symptoms – problems with language, disorientation, mood swings, behavioral issues – as it progresses, eventually leading to the loss of bodily functions and death. Because of the dementia the disease causes, Alzheimer’s patients require a great deal of care, and the world spends ~1% of its total GDP on caring for those with Alzheimer’s and related disorders. Because the prevalence of the disease increases with age, the situation will worsen as life expectancies around the globe increase: worldwide cases of Alzheimer’s are expected to grow from 35 million today to over 115 million by 2050.
Despite much research, the exact causes of Alzheimer’s disease remains poorly understood. The disease seems to be related to the accumulation of plaques made of amyloid-β peptides that form on the outside of neurons, as well as the formation of tangles of the protein tau inside of neurons. Although many efforts have been made to target amyloid-β or the enzymes involved in its formation, we have so far been unsuccessful at finding any treatment that stops the disease or reverses its progress. Some researchers believe that most attempts at treating Alzheimer’s have failed because, by the time a patient shows symptoms, the disease has already progressed past the point of no return.
While research towards a cure continues, researchers have sought effective ways to prevent Alzheimer’s disease. Although some studies show that mental and physical exercise may lower ones risk of Alzheimer’s disease, approximately 60-80% of the risk for Alzheimer’s disease appears to be genetic. Thus, if we’re serious about prevention, we may have to act at the genetic level. And because the brain is difficult to access surgically for gene therapy in adults, this means using gene editing on embryos.
With the latest CRISPR/Cas9 advance, the exhortation “turn on, tune in, drop out” comes to mind. The CRISPR/Cas9 gene-editing system was already a well-known means of “tuning in” (inserting new genes) and “dropping out” (knocking out genes). But when it came to “turning on” genes, CRISPR/Cas9 had little potency. That is, it had demonstrated only limited success as a way to activate specific genes.
A new CRISPR/Cas9 approach, however, appears capable of activating genes more effectively than older approaches. The new approach may allow scientists to more easily determine the function of individual genes, according to Feng Zhang, Ph.D., a researcher at MIT and the Broad Institute. Dr. Zhang and colleagues report that the new approach permits multiplexed gene activation and rapid, large-scale studies of gene function.
The new technique was introduced in the December 10 online edition of Nature, in an article entitled, “Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex.” The article describes how Dr. Zhang, along with the University of Tokyo’s Osamu Nureki, Ph.D., and Hiroshi Nishimasu, Ph.D., overhauled the CRISPR/Cas9 system. The research team based their work on their analysis (published earlier this year) of the structure formed when Cas9 binds to the guide RNA and its target DNA. Specifically, the team used the structure’s 3D shape to rationally improve the system.
In previous efforts to revamp CRISPR/Cas9 for gene activation purposes, scientists had tried to attach the activation domains to either end of the Cas9 protein, with limited success. From their structural studies, the MIT team realized that two small loops of the RNA guide poke out from the Cas9 complex and could be better points of attachment because they allow the activation domains to have more flexibility in recruiting transcription machinery.
Using their revamped system, the researchers activated about a dozen genes that had proven difficult or impossible to turn on using the previous generation of Cas9 activators. Each gene showed at least a twofold boost in transcription, and for many genes, the researchers found multiple orders of magnitude increase in activation.
After investigating single-guide RNA targeting rules for effective transcriptional activation, demonstrating multiplexed activation of 10 genes simultaneously, and upregulating long intergenic noncoding RNA transcripts, the research team decided to undertake a large-scale screen. This screen was designed to identify genes that confer resistance to a melanoma drug called PLX-4720.
“We … synthesized a library consisting of 70,290 guides targeting all human RefSeq coding isoforms to screen for genes that, upon activation, confer resistance to a BRAF inhibitor,” wrote the authors of the Nature paper. “The top hits included genes previously shown to be able to confer resistance, and novel candidates were validated using individual [single-guide RNA] and complementary DNA overexpression.”
A gene signature based on the top screening hits, the authors added, correlated with a gene expression signature of BRAF inhibitor resistance in cell lines and patient-derived samples. It was also suggested that large-scale screens such as the one demonstrated in the current study could help researchers discover new cancer drugs that prevent tumors from becoming resistant.
Familial amyloid polyneuropathy type I is an autosomal dominant disorder caused by mutations in the transthyretin (TTR ) gene; however, carriers of the same mutation exhibit variability in penetrance and clinical expression. We analyzed alleles of candidate genes encoding non-fibrillar components of TTR amyloid deposits and a molecule metabolically interacting with TTR [retinol-binding protein (RBP)], for possible associations with age of disease onset and/or susceptibility in a Portuguese population sample with the TTR V30M mutation and unrelated controls. We show that the V30M carriers represent a distinct subset of the Portuguese population. Estimates of genetic distance indicated that the controls and the classical onset group were furthest apart, whereas the late-onset group appeared to differ from both. Importantly, the data also indicate that genetic interactions among the multiple loci evaluated, rather than single-locus effects, are more likely to determine differences in the age of disease onset. Multifactor dimensionality reduction indicated that the best genetic model for classical onset group versus controls involved the APCS gene, whereas for late-onset cases, one APCS variant (APCSv1) and two RBP variants (RBPv1 and RBPv2) are involved. Thus, although the TTR V30M mutation is required for the disease in Portuguese patients, different genetic factors may govern the age of onset, as well as the occurrence of anticipation.
Autosomal dominant disorders may vary in expression even within a given kindred. The basis of this variability is uncertain and can be attributed to epigenetic factors, environment or epistasis. We have studied familial amyloid polyneuropathy (FAP), an autosomal dominant disorder characterized by peripheral sensorimotor and autonomic neuropathy. It exhibits variation in cardiac, renal, gastrointestinal and ocular involvement, as well as age of onset. Over 80 missense mutations in the transthyretin gene (TTR ) result in autosomal dominant disease http://www.ibmc.up.pt/~mjsaraiv/ttrmut.html). The presence of deposits consisting entirely of wild-type TTR molecules in the hearts of 10– 25% of individuals over age 80 reveals its inherent in vivo amyloidogenic potential (1).
FAP was initially described in Portuguese (2) where, until recently, the TTR V30M has been the only pathogenic mutation associated with the disease (3,4). Later reports identified the same mutation in Swedish and Japanese families (5,6). The disorder has since been recognized in other European countries and in North American kindreds in association with V30M, as well as other mutations (7).
TTR V30M produces disease in only 5–10% of Swedish carriers of the allele (8), a much lower degree of penetrance than that seen in Portuguese (80%) (9) or in Japanese with the same mutation. The actual penetrance in Japanese carriers has not been formally established, but appears to resemble that seen in Portuguese. Portuguese and Japanese carriers show considerable variation in the age of clinical onset (10,11). In both populations, the first symptoms had originally been described as typically occurring before age 40 (so-called ‘classical’ or early-onset); however, in recent years, more individuals developing symptoms late in life have been identified (11,12). Hence, present data indicate that the distribution of the age of onset in Portuguese is continuous, but asymmetric with a mean around age 35 and a long tail into the older age group (Fig. 1) (9,13). Further, DNA testing in Portugal has identified asymptomatic carriers over age 70 belonging to a subset of very late-onset kindreds in whose descendants genetic anticipation is frequent. The molecular basis of anticipation in FAP, which is not mediated by trinucleotide repeat expansions in the TTR or any other gene (14), remains elusive.
Variation in penetrance, age of onset and clinical features are hallmarks of many autosomal dominant disorders including the human TTR amyloidoses (7). Some of these clearly reflect specific biological effects of a particular mutation or a class of mutants. However, when such phenotypic variability is seen with a single mutation in the gene encoding the same protein, it suggests an effect of modifying genetic loci and/or environmental factors contributing differentially to the course of disease. We have chosen to examine age of onset as an example of a discrete phenotypic variation in the presence of the particular autosomal dominant disease-associated mutation TTR V30M. Although the role of environmental factors cannot be excluded, the existence of modifier genes involved in TTR amyloidogenesis is an attractive hypothesis to explain the phenotypic variability in FAP. ….
ATTR (TTR amyloid), like all amyloid deposits, contains several molecular components, in addition to the quantitatively dominant fibril-forming amyloid protein, including heparan sulfate proteoglycan 2 (HSPG2 or perlecan), SAP, a plasma glycoprotein of the pentraxin family (encoded by the APCS gene) that undergoes specific calcium-dependent binding to all types of amyloid fibrils, and apolipoprotein E (ApoE), also found in all amyloid deposits (15). The ApoE4 isoform is associated with an increased frequency and earlier onset of Alzheimer’s disease (Ab), the most common form of brain amyloid, whereas the ApoE2 isoform appears to be protective (16). ApoE variants could exert a similar modulatory effect in the onset of FAP, although early studies on a limited number of patients suggested this was not the case (17).
In at least one instance of senile systemic amyloidosis, small amounts of AA-related material were found in TTR deposits (18). These could reflect either a passive co-aggregation or a contributory involvement of protein AA, encoded by the serum amyloid A (SAA ) genes and the main component of secondary (reactive) amyloid fibrils, in the formation of ATTR.
Retinol-binding protein (RBP), the serum carrier of vitamin A, circulates in plasma bound to TTR. Vitamin A-loaded RBP and L-thyroxine, the two natural ligands of TTR, can act alone or synergistically to inhibit the rate and extent of TTR fibrillogenesis in vitro, suggesting that RBP may influence the course of FAP pathology in vivo (19). We have analyzed coding and non-coding sequence polymorphisms in the RBP4 (serum RBP, 10q24), HSPG2 (1p36.1), APCS (1q22), APOE (19q13.2), SAA1 and SAA2 (11p15.1) genes with the goal of identifying chromosomes carrying common and functionally significant variants. At the time these studies were performed, the full human genome sequence was not completed and systematic singlenucleotide polymorphism (SNP) analyses were not available for any of the suspected candidate genes. We identified new SNPs in APCS and RBP4 and utilized polymorphisms in SAA, HSPG2 and APOE that had already been characterized and shown to have potential pathophysiologic significance in other disorders (16,20–22). The genotyping data were analyzed for association with the presence of the V30M amyloidogenic allele (FAP patients versus controls) and with the age of onset (classical- versus late-onset patients). Multilocus analyses were also performed to examine the effects of simultaneous contributions of the six loci for determining the onset of the first symptoms. …..
The potential for different underlying models for classical and late onset is supported by the MDR analysis, which produces two distinct models when comparing each class with the controls. One could view the two onset classes as unique diseases. If this is the case, then the failure to detect a single predictive genetic model is consistent with two related, but different, diseases. This is exactly what would be expected in such a case of genetic heterogeneity (28). Using this approach, a major gene effect can be viewed as a necessary, but not sufficient, condition to explain the course of the disease. Analyzing the cases but omitting from the analysis of phenotype the necessary allele, in this case TTR V30M, can then reveal a variety of important modifiers that are distinct between the phenotypes.
The significant comparisons obtained in our study cohort indicate that the combined effects mainly result from two and three-locus interactions involving all loci except SAA1 and SAA2 for susceptibility to disease. A considerable number of four-site combinations modulate the age of onset with SAA1 appearing in a majority of significant combinations in late-onset disease, perhaps indicating a greater role of the SAA variants in the age of onset of FAP.
The correlation between genotype and phenotype in socalled simple Mendelian disorders is often incomplete, as only a subset of all mutations can reliably predict specific phenotypes (34). This is because non-allelic genetic variations and/or environmental influences underlie these disorders whose phenotypes behave as complex traits. A few examples include the identification of the role of homozygozity for the SAA1.1 allele in conferring the genetic susceptibility to renal amyloidosis in FMF (20) and the association of an insertion/deletion polymorphism in the ACE gene with disease severity in familial hypertrophic cardiomyopathy (35). In these disorders, the phenotypes arise from mutations in MEFV and b-MHC, but are modulated by independently inherited genetic variation. In this report, we show that interactions among multiple genes, whose products are confirmed or putative constituents of ATTR deposits, or metabolically interact with TTR, modulate the onset of the first symptoms and predispose individuals to disease in the presence of the V30M mutation in TTR. The exact nature of the effects identified here requires further study with potential application in the development of genetic screening with prognostic value pertaining to the onset of disease in the TTR V30M carriers.
If the effects of additional single or interacting genes dictate the heterogeneity of phenotype, as reflected in variability of onset and clinical expression (with the same TTR mutation), the products encoded by alleles at such loci could contribute to the process of wild-type TTR deposition in elderly individuals without a mutation (senile systemic amyloidosis), a phenomenon not readily recognized as having a genetic basis because of the insensitivity of family history in the elderly.
Safety and Efficacy of RNAi Therapy for Transthyretin Amyloidosis
Transthyretin amyloidosis is caused by the deposition of hepatocyte-derived transthyretin amyloid in peripheral nerves and the heart. A therapeutic approach mediated by RNA interference (RNAi) could reduce the production of transthyretin.
Methods We identified a potent antitransthyretin small interfering RNA, which was encapsulated in two distinct first- and second-generation formulations of lipid nanoparticles, generating ALN-TTR01 and ALN-TTR02, respectively. Each formulation was studied in a single-dose, placebo-controlled phase 1 trial to assess safety and effect on transthyretin levels. We first evaluated ALN-TTR01 (at doses of 0.01 to 1.0 mg per kilogram of body weight) in 32 patients with transthyretin amyloidosis and then evaluated ALN-TTR02 (at doses of 0.01 to 0.5 mg per kilogram) in 17 healthy volunteers.
Results Rapid, dose-dependent, and durable lowering of transthyretin levels was observed in the two trials. At a dose of 1.0 mg per kilogram, ALN-TTR01 suppressed transthyretin, with a mean reduction at day 7 of 38%, as compared with placebo (P=0.01); levels of mutant and nonmutant forms of transthyretin were lowered to a similar extent. For ALN-TTR02, the mean reductions in transthyretin levels at doses of 0.15 to 0.3 mg per kilogram ranged from 82.3 to 86.8%, with reductions of 56.6 to 67.1% at 28 days (P<0.001 for all comparisons). These reductions were shown to be RNAi mediated. Mild-to-moderate infusion-related reactions occurred in 20.8% and 7.7% of participants receiving ALN-TTR01 and ALN-TTR02, respectively.
ALN-TTR01 and ALN-TTR02 suppressed the production of both mutant and nonmutant forms of transthyretin, establishing proof of concept for RNAi therapy targeting messenger RNA transcribed from a disease-causing gene.
Alnylam May Seek Approval for TTR Amyloidosis Rx in 2017 as Other Programs Advance
Officials from Alnylam Pharmaceuticals last week provided updates on the two drug candidates from the company’s flagship transthyretin-mediated amyloidosis program, stating that the intravenously delivered agent patisiran is proceeding toward a possible market approval in three years, while a subcutaneously administered version called ALN-TTRsc is poised to enter Phase III testing before the end of the year.
Meanwhile, Alnylam is set to advance a handful of preclinical therapies into human studies in short order, including ones for complement-mediated diseases, hypercholesterolemia, and porphyria.
The officials made their comments during a conference call held to discuss Alnylam’s second-quarter financial results.
ATTR is caused by a mutation in the TTR gene, which normally produces a protein that acts as a carrier for retinol binding protein and is characterized by the accumulation of amyloid deposits in various tissues. Alnylam’s drugs are designed to silence both the mutant and wild-type forms of TTR.
Patisiran, which is delivered using lipid nanoparticles developed by Tekmira Pharmaceuticals, is currently in a Phase III study in patients with a form of ATTR called familial amyloid polyneuropathy (FAP) affecting the peripheral nervous system. Running at over 20 sites in nine countries, that study is set to enroll up to 200 patients and compare treatment to placebo based on improvements in neuropathy symptoms.
According to Alnylam Chief Medical Officer Akshay Vaishnaw, Alnylam expects to have final data from the study in two to three years, which would put patisiran on track for a new drug application filing in 2017.
Meanwhile, ALN-TTRsc, which is under development for a version of ATTR that affects cardiac tissue called familial amyloidotic cardiomyopathy (FAC) and uses Alnylam’s proprietary GalNAc conjugate delivery technology, is set to enter Phase III by year-end as Alnylam holds “active discussions” with US and European regulators on the design of that study, CEO John Maraganore noted during the call.
In the interim, Alnylam continues to enroll patients in a pilot Phase II study of ALN-TTRsc, which is designed to test the drug’s efficacy for FAC or senile systemic amyloidosis (SSA), a condition caused by the idiopathic accumulation of wild-type TTR protein in the heart.
Based on “encouraging” data thus far, Vaishnaw said that Alnylam has upped the expected enrollment in this study to 25 patients from 15. Available data from the trial is slated for release in November, he noted, stressing that “any clinical endpoint result needs to be considered exploratory given the small sample size and the very limited duration of treatment of only six weeks” in the trial.
Vaishnaw added that an open-label extension (OLE) study for patients in the ALN-TTRsc study will kick off in the coming weeks, allowing the company to gather long-term dosing tolerability and clinical activity data on the drug.
Enrollment in an OLE study of patisiran has been completed with 27 patients, he said, and, “as of today, with up to nine months of therapy … there have been no study drug discontinuations.” Clinical endpoint data from approximately 20 patients in this study will be presented at the American Neurological Association meeting in October.
As part of its ATTR efforts, Alnylam has also been conducting natural history of disease studies in both FAP and FAC patients. Data from the 283-patient FAP study was presented earlier this year and showed a rapid progression in neuropathy impairment scores and a high correlation of this measurement with disease severity.
During last week’s conference call, Vaishnaw said that clinical endpoint and biomarker data on about 400 patients with either FAC or SSA have already been collected in a nature history study on cardiac ATTR. Maraganore said that these findings would likely be released sometime next year.
The first medication for a rare and often fatal protein misfolding disorder has been approved in Europe. On November 16, the E gave a green light to Pfizer’s Vyndaqel (tafamidis) for treating transthyretin amyloidosis in adult patients with stage 1 polyneuropathy symptoms. [Jeffery Kelly, La Jolla]
The most clinically advanced RNA interference (RNAi) therapeutic achieved a milestone in April when Alnylam Pharmaceuticals in Cambridge, Massachusetts, reported positive results for patisiran, a small interfering RNA (siRNA) oligonucleotide targeting transthyretin for treating familial amyloidotic polyneuropathy (FAP). …
FAP is characterized by the systemic deposition of amyloidogenic variants of the transthyretin protein, especially in the peripheral nervous system, causing a progressive sensory and motor polyneuropathy.
FAP is caused by a mutation of the TTR gene, located on human chromosome 18q12.1-11.2.[5] A replacement of valine by methionine at position 30 (TTR V30M) is the mutation most commonly found in FAP.[1] The variant TTR is mostly produced by the liver.[citation needed] The transthyretin protein is a tetramer. ….

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
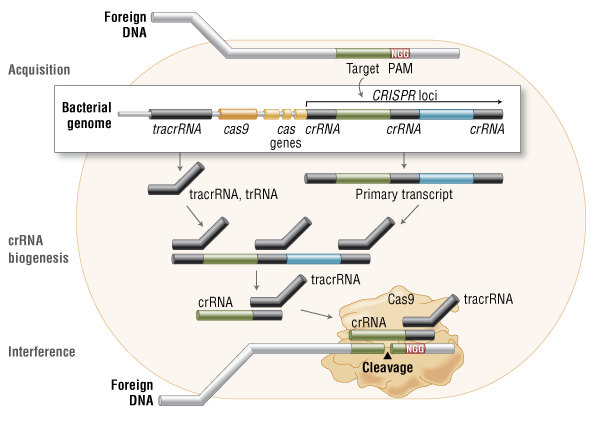{kind=link}
{kind=link}
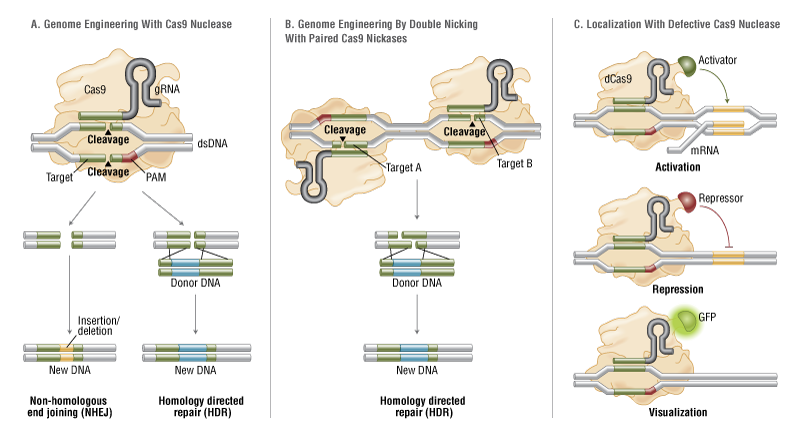{kind=link}
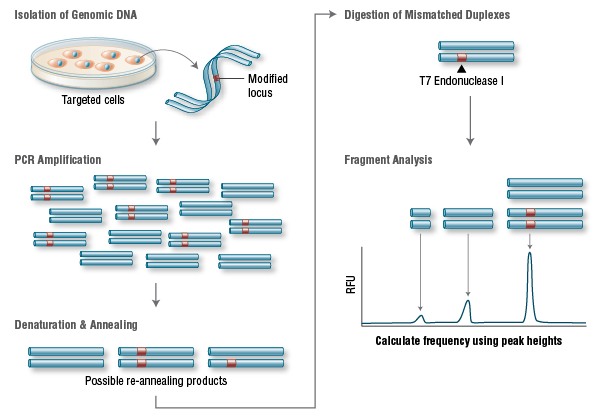{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
This is a lovely method and should find wide applicability in many settings, especially for microorganisms and cell lines. However, it is not clear that this approach will be, as implied by the discussion, an efficient mapping method for all multicellular organisms. I have performed similar experiments in Drosophila, focused on meiotic recombination, on a much smaller scale, and found that CRISPR-Cas9 can indeed generate targeted recombination at gRNA target sites. In every case I tested, I found that the recombination event was associated with a deletion at the gRNA site, which is probably unimportant for most mapping efforts, but may be a concern in some specific cases, for example for clinical applications. It would be interesting to know how often mutations occurred at the targeted gRNA site in this study.
The wider issue, however, is whether CRISPR-mediated recombination will be more efficient than other methods of mapping. After careful consideration of all the costs and the time involved in each of the steps for Drosophila, we have decided that targeted meiotic recombination using flanking visible markers will be, in most cases, considerably more efficient than CRISPR-mediated recombination. This is mainly due to the large expense of injecting embryos and the extensive effort and time required to screen injected animals for appropriate events. It is both cheaper and faster to generate markers (with CRISPR) and then perform a large meiotic recombination mapping experiment than it would be to generate the lines required for CRISPR-mediated recombination mapping. It is possible to dramatically reduce costs by, for example, mapping sequentially at finer resolution. But this approach would require much more time than marker-assisted mapping. If someone develops a rapid and cheap method of reliably introducing DNA into Drosophila embryos, then this calculus might change.
However, it is possible to imagine situations where CRISPR-mediated mapping would be preferable, even for Drosophila. For example, some genomic regions display extremely low or highly non-uniform recombination rates. It is possible that CRISPR-mediated mapping could provide a reasonable approach to fine mapping genes in these regions.
The authors also propose the exciting possibility that CRISPR-mediated loss of heterozygosity could be used to map traits in sterile species hybrids. It is not entirely obvious to me how this experiment would proceed and I hope the authors can illuminate me. If we imagine driving a recombination event in the early embryo (with maternal Cas9 from one parent and gRNA from a second parent), then at best we would end up with chimeric individuals carrying mitotic clones. I don’t think one could generate diploid animals where all cells carried the same loss of heterozygosity event. Even if we could, this experiment would require construction of a substantial number of stable transgenic lines expressing gRNAs. Mapping an ~20Mbp chromosome arm to ~10kb would require on the order of two-thousand transgenic lines. Not an undertaking to be taken lightly. It is already possible to perform similar tests (hemizygosity tests) using D. melanogaster deficiency lines in crosses with D. simulans, so perhaps CRISPR-mediated LOH could complement these deficiency screens for fine mapping efforts. But, at the moment, it is not clear to me how to do the experiment.