Healthcare analytics, AI solutions for biological big data, providing an AI platform for the biotech, life sciences, medical and pharmaceutical industries, as well as for related technological approaches, i.e., curation and text analysis with machine learning and other activities related to AI applications to these industries.
Dr. Ahmad Masri and Dr. C Michael Gibson discuss outpatient worsening heart failure in transthyretin amyloid cardiomyopathy in the Attribute-CM trial, May 29, 2026
The following are key points to remember from this update on the treatment of cardiac transthyretin amyloidosis:
Transthyretin (TTR) is a highly conserved protein involved in transportation of thyroxine (T4) and retinol-binding protein. TTR is synthesized mostly by the liver and is rich in beta strands with an intrinsic propensity to aggregate into insoluble amyloid fibers, which deposit within tissue leading to the development of TTR-related amyloidosis (ATTR). ATTR can follow the deposition of either variant TTR (ATTRv, previously known as mutant ATTR) or wild type TTR (ATTRwt).
Cardiac ATTR has a favorable survival rate compared to light chain (AL) amyloidosis, with a median survival of 75 versus 11 months. However, ATTR cardiomyopathy is a progressive disorder but newer therapeutic options include tafamidis (positive phase 3 clinical trial), and possibly patisiran and inotersen.
Inhibition of the Synthesis of Mutated Transthyretin
Liver transplantation removes the source of mutated TTR molecules and prolongs survival, with a 20-year survival of 55.3%. However, tissue accumulation of TTR can continue after liver transplantation because TTR amyloid fibers promote subsequent deposition of ATTRwt. Combined liver–heart transplantation is feasible in younger patients with ATTRv cardiomyopathy and a small series suggests better prognosis than cardiac transplantation.
Inhibition of TTR gene expression: Patisiran is a small interfering RNA blocking the expression of both variant and wt TTR. On the basis of the APOLLO trial, it was approved for therapy of adults with ATTRv-related polyneuropathy both in the United States and European Union. In this trial, patisiran promoted favorable myocardial remodeling based on echocardiographic and N-terminal B-type natriuretic peptide (NT-BNP) changes (this effect was not demonstrated for inotersen) and is still under investigation for tafamidis.
Antisense oligonucleotides inotersen inhibits the production of both variant and wt TTR. Based on the findings of the NEURO-TTR trial, the Food and Drug Administration (FDA) approved this agent for patients with ATTRv-related polyneuropathy. In the NEURO-TTR trial, cardiomyopathy was present in 63%, but the study was not powered to measure effects of inotersen on heart disease. Inotersen can cause thrombocytopenia and must be used cautiously with bleeding risk.
Tetramer Stabilization
Selective stabilizers include tafamidis and AG10. Tafamidis is a benzoxazole and a small molecule that inhibits the dissociation of TTR tetramers by binding the T4-binding sites. The phase ATTR-ACT study showed that when comparing the pooled tafamidis arms (80 and 20 mg) with the placebo arm, tafamidis was associated with lower all-cause mortality than placebo (78 of 264 [29.5%] vs. 76 of 177 [42.9%]; hazard ratio, 0.70; 95% confidence interval, 0.51-0.96) and a lower rate of cardiovascular hospitalizations. Based on the results of the ATTR-ACT trial, it has received Breakthrough Therapy designation from the FDA for treatment of ATTR cardiomyopathy.
Nonselective agents: Diflunisal, a nonsteroidal anti-inflammatory drug, is reported to stabilize TTR tetramers. More studies are needed to confirm its clinical efficacy.
Inhibition of Oligomer Aggregation and Oligomer Disruption
Epigallocatechin gallate is the most abundant catechin in green tea. One single-center open-label 12-month study did not show survival benefits or any change in echocardiographic parameters or NT-BNP compared to baseline.
Degradation and Reabsorption of Amyloid Fibers
Doxycycline-taurosodeoxycholic acid (TUDCA) has been evaluated in two small studies and the results appear to be modest. More data are needed to confirm its efficacy.
Antibodies targeting serum amyloid P protein or amyloid fibrils: Patient enrollment for miridesap followed by anti-SAP antibodies was suspended, and this approach is not being evaluated currently. However, a monoclonal antibody designed to specifically target TTR amyloid deposits (PRX004) has entered clinical evaluation, with an ongoing phase 1 study on ATTRv.
Supportive Treatment of Cardiac Involvement
Drug therapies: Although angiotensin-converting enzyme (ACE) inhibitors/angiotensin-receptor blockers (ARBs) and beta-blockers may have been poorly tolerated in the ATTR-ACT trial, 30% of the patients were on ACE inhibitors/ARBs. There are no data with digoxin in TTR amyloid, and non-dihydropyridine calcium channel blockers are contraindicated due to negative inotropy.
Implantable cardioverter-defibrillators (ICDs): In one study, which included 53 patients with amyloid, ICD shocks occurred exclusively in the AL amyloid group and none in the TTR amyloid patients. Higher defibrillation thresholds and complication rates are of concern.
Cardiac pacing: In a large series of ATTRv-related polyneuropathy (n = 262), a pacemaker was implanted in 110 patients with His ventricular interval >700 ms. The authors recommend that any conduction disturbance on 12-lead electrocardiogram (ECG) warrants further investigation with Holter monitoring to determine candidacy for a pacemaker.
Left ventricular assist device (LVAD): Although an LVAD is technically feasible, it is associated with high short-term mortality and worse outcomes than in dilated cardiomyopathy.
Cardiac transplantation: This is a valuable option for patients with end-stage heart failure when significant extracardiac disease is excluded. In one study with 10 patients, only episodes of amyloid recurrence occurred.
This is an outstanding overview of this topic and recommended reading for anyone who cares for patients with cardiac transthyretin amyloid.
First-Ever Evidence that Patisiran Reduces Pathogenic, Misfolded TTR Monomers and Oligomers in FAP Patients
We reported data from our ongoing Phase 2 open-label extension (OLE) study of patisiran, an investigational RNAi therapeutic targeting transthyretin (TTR) for the treatment of TTR-mediated amyloidosis (ATTR amyloidosis) patients with familial amyloidotic polyneuropathy (FAP). Alnylam scientists and collaborators from The Scripps Research Institute and Misfolding Diagnostics, Inc. were able to measure the effects of patisiran on pathogenic, misfolded TTR monomers and oligomers in FAP patients. Results showed a rapid and sustained reduction in serum non-native conformations of TTR (NNTTR) of approximately 90%. Since NNTTR is pathogenic in ATTR amyloidosis and the level of NNTTR reduction correlated with total TTR knockdown, these results provide direct mechanistic evidence supporting the therapeutic hypothesis that TTR knockdown has the potential to result in clinical benefit. Furthermore, complete 12-month data from all 27 patients that enrolled in the patisiran Phase 2 OLE study showed sustained mean maximum reductions in total serum TTR of 91% for over 18 months and a mean 3.1-point decrease in mNIS+7 at 12 months, which compares favorably to an estimated increase in mNIS+7 of 13 to 18 points at 12 months based upon analysis of historical data sets in untreated FAP patients with similar baseline characteristics. Importantly, patisiran administration continues to be generally well tolerated out to 21 months of treatment.
We are encouraged by these new data that provide continued support for our hypothesis that patisiran has the potential to halt neuropathy progression in patients with FAP. If these results are replicated in a randomized, double-blind, placebo-controlled study, we believe that patisiran could emerge as an important treatment option for patients suffering from this debilitating, progressive and life-threatening disease.
Hereditary ATTR Amyloidosis with Polyneuropathy (hATTR-PN)
ATTR amyloidosis is a progressive, life-threatening disease caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in multiple organs, but primarily in the peripheral nerves and heart. ATTR amyloidosis can lead to significant morbidity, disability, and mortality. The TTR protein is produced primarily in the liver and is normally a carrier for retinol binding protein – one of the vehicles used to transport vitamin A around the body. Mutations in the TTR gene cause misfolding of the protein and the formation of amyloid fibrils that typically contain both mutant and wild-type TTR that deposit in tissues such as the peripheral nerves and heart, resulting in intractable peripheral sensory neuropathy, autonomic neuropathy, and/or cardiomyopathy.
ATTR represents a major unmet medical need with significant morbidity and mortality. There are over 100 reported TTR mutations; the particular TTR mutation and the site of amyloid deposition determine the clinical manifestations of the disease whether it is predominantly symptoms of neuropathy or cardiomyopathy.
Specifically, hereditary ATTR amyloidosis with polyneuropathy (hATTR-PN), also known as familial amyloidotic polyneuropathy (FAP), is an inherited, progressive disease leading to death within 5 to 15 years. It is due to a mutation in the transthyretin (TTR) gene, which causes misfolded TTR proteins to accumulate as amyloid fibrils predominantly in peripheral nerves and other organs. hATTR-PN can cause sensory, motor, and autonomic dysfunction, resulting in significant disability and death.
It is estimated that hATTR-PN, also known as FAP, affects approximately 10,000 people worldwide. Patients have a life expectancy of 5 to 15 years from symptom onset, and the only treatment options for early stage disease are liver transplantation and TTR stabilizers such as tafamidis (approved in Europe) and diflunisal. Unfortunately liver transplantation has limitations, including limited organ availability as well as substantial morbidity and mortality. Furthermore, transplantation eliminates the production of mutant TTR but does not affect wild-type TTR, which can further deposit after transplantation, leading to cardiomyopathy and worsening of neuropathy. There is a significant need for novel therapeutics to treat patients who have inherited mutations in the TTR gene.
Our ATTR program is the lead effort in our Genetic Medicine Strategic Therapeutic Area (STAr) product development and commercialization strategy, which is focused on advancing innovative RNAi therapeutics toward genetically defined targets for the treatment of rare diseases with high unmet medical need. We are developing patisiran (ALN-TTR02), an intravenously administered RNAi therapeutic, to treat the hATTR-PN form of the disease.
Patisiran for the Treatment hATTR-PN
APOLLO Phase 3 Trial
In 2012, Alnylam entered into an exclusive alliance with Genzyme, a Sanofi company, to develop and commercialize RNAi therapeutics, including patisiran and revusiran, for the treatment of ATTR amyloidosis in Japan and the broader Asian-Pacific region. In early 2014, this relationship was extended as a significantly broader alliance to advance RNAi therapeutics as genetic medicines. Under this new agreement, Alnylam will lead development and commercialization of patisiran in North America and Europe while Genzyme will develop and commercialize the product in the rest of world.
Hereditary ATTR Amyloidosis with Cardiomyopathy (hATTR-CM)
ATTR amyloidosis is a progressive, life-threatening disease caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in multiple organs, but primarily in the peripheral nerves and heart. ATTR amyloidosis can lead to significant morbidity, disability, and mortality. The TTR protein is produced primarily in the liver and is normally a carrier for retinol binding protein – one of the vehicles used to transport vitamin A around the body. Mutations in the TTR gene cause misfolding of the protein and the formation of amyloid fibrils that typically contain both mutant and wild-type TTR that deposit in tissues such as the peripheral nerves and heart, resulting in intractable peripheral sensory neuropathy, autonomic neuropathy, and/or cardiomyopathy.
ATTR represents a major unmet medical need with significant morbidity and mortality. There are over 100 reported TTR mutations; the particular TTR mutation and the site of amyloid deposition determine the clinical manifestations of the disease, whether it is predominantly symptoms of neuropathy or cardiomyopathy.
Specifically, hereditary ATTR amyloidosis with cardiomyopathy (hATTR-CM), also known as familial amyloidotic cardiomyopathy (FAC), is an inherited, progressive disease leading to death within 2 to 5 years. It is due to a mutation in the transthyretin (TTR) gene, which causes misfolded TTR proteins to accumulate as amyloid fibrils primarily in the heart. Hereditary ATTR amyloidosis with cardiomyopathy can result in heart failure and death.
While the exact numbers are not known, it is estimated hATTR-CM, also known as FAC affects at least 40,000 people worldwide. hATTR-CM is fatal within 2 to 5 years of diagnosis and treatment is currently limited to supportive care. Wild-type ATTR amyloidosis (wtATTR amyloidosis), also known as senile systemic amyloidosis, is a nonhereditary, progressive disease leading to death within 2 to 5 years. It is caused by misfolded transthyretin (TTR) proteins that accumulate as amyloid fibrils in the heart. Wild-type ATTR amyloidosis can cause cardiomyopathy and result in heart failure and death. There are no approved therapies for the treatment of hATTR-CM or SSA; hence there is a significant unmet need for novel therapeutics to treat these patients.
Our ATTR program is the lead effort in our Genetic Medicine Strategic Therapeutic Area (STAr) product development and commercialization strategy, which is focused on advancing innovative RNAi therapeutics toward genetically defined targets for the treatment of rare diseases with high unmet medical need. We are developing revusiran (ALN-TTRsc), a subcutaneously administered RNAi therapeutic for the treatment of hATTR-CM.
Revusiran for the Treatment of hATTR-CM
ENDEAVOUR Phase 3 Trial
In 2012, Alnylam entered into an exclusive alliance with Genzyme, a Sanofi company, to develop and commercialize RNAi therapeutics, including patisiran and revusiran, for the treatment of ATTR amyloidosis in Japan and the broader Asian-Pacific region. In early 2014, this relationship was extended as a broader alliance to advance RNAi therapeutics as genetic medicines. Under this new agreement, Alnylam and Genzyme have agreed to co-develop and co-commercialize revusiran in North America and Europe, with Genzyme developing and commercializing the product in the rest of world. This broadened relationship on revusiran is aimed at expanding and accelerating the product’s global value.
We presented pre-clinical data with ALN-TTRsc02, an investigational RNAi therapeutic targeting transthyretin (TTR) for the treatment of TTR-mediated amyloidosis (ATTR amyloidosis). In pre-clinical studies, including those in non-human primates (NHPs), ALN-TTRsc02 achieved potent and highly durable knockdown of serum TTR of up to 99% with multi-month durability achieved after just a single dose, supportive of a potentially once quarterly dose regimen. Results from studies comparing TTR knockdown activity of ALN-TTRsc02 to that of revusiran showed that ALN-TTRsc02 has a markedly superior TTR knockdown profile. Further, in initial rat toxicology studies, ALN-TTRsc02 was found to be generally well tolerated with no significant adverse events at doses as high as 100 mg/kg.
Heart failure with a preserved ejection fraction (HFPEF) is a clinical syndrome that has no pharmacologic therapies approved for this use to date. In light of failed medicines, cardiologists have refocused treatment strategies based on the theory that HFPEF is a heterogeneous clinical syndrome with different etiologies. Classification of HFPEF according to etiologic subtype may, therefore, identify cohorts with treatable pathophysiologic mechanisms and may ultimately pave the way forward for developing meaningful HFPEF therapies.1
A wealth of data now indicates that amyloid infiltration is an important mechanism underlying HFPEF. Inherited mutations in transthyretin cardiac amyloidosis (ATTRm) or the aging process in wild-type disease (ATTRwt) cause destabilization of the transthyretin (TTR) protein into monomers or oligomers, which aggregate into amyloid fibrils. These insoluble fibrils accumulate in the myocardium and result in diastolic dysfunction, restrictive cardiomyopathy, and eventual congestive heart failure (Figure 1). In an autopsy study of HFPEF patients, almost 20% without antemortem suspicion of amyloid had left ventricular (LV) TTR amyloid deposition.2 Even more resounding evidence for the contribution of TTR amyloid to HFPEF was a study in which 120 hospitalized HFPEF patients with LV wall thickness ≥12 mm underwent technetium-99m 3,3-diphosphono-1,2-propranodicarboxylic acid (99mTc-DPD) cardiac imaging,3,4 a bone isotope known to have high sensitivity and specificity for diagnosing TTR cardiac amyloidosis.5,6 Moderate-to-severe myocardial uptake indicative of TTR cardiac amyloid deposition was detected in 13.3% of HFPEF patients who did not have TTR gene mutations. Therefore, TTR cardiac amyloid deposition, especially in older adults, is not rare, can be easily identified, and may contribute to the underlying pathophysiology of HFPEF.
Figure 1
As no U.S. Food and Drug Administration-approved drugs are currently available for the treatment of HFPEF or TTR cardiac amyloidosis, the development of medications that attenuate or prevent TTR-mediated organ toxicity has emerged as an important therapeutic goal. Over the past decade, a host of therapies and therapeutic drug classes have emerged in clinical trials (Table 1), and these may herald a new direction for treating HFPEF secondary to TTR amyloid.
Table 1
TTR Silencers (siRNA and Antisense Oligonucleotides)
siRNA
Ribonucleic acid interference (RNAi) has surfaced as an endogenous cellular mechanism for controlling gene expression. Small interfering RNAs (siRNAs) delivered into cells can disrupt the production of target proteins.7,8 A formulation of lipid nanoparticle and triantennary N-acetylgalactosamine (GalNAc) conjugate that delivers siRNAs to hepatocytes is currently in clinical trials.9 Prior research demonstrated these GalNAc-siRNA conjugates result in robust and durable knockdown of a variety of hepatocyte targets across multiple species and appear to be well suited for suppression of TTR gene expression and subsequent TTR protein production.
The TTR siRNA conjugated to GalNAc, ALN-TTRSc, is now under active investigation as a subcutaneous injection in phase 3 clinical trials in patients with TTR cardiac amyloidosis.10 Prior phase 2 results demonstrated that ALN-TTRSc was generally well tolerated in patients with significant TTR disease burden and that it reduced both wild-type and mutant TTR gene expression by a mean of 87%. Harnessing RNAi technology appears to hold great promise for treating patients with TTR cardiac amyloidosis. The ability of ALN-TTRSc to lower both wild-type and mutant proteins may provide a major advantage over liver transplantation, which affects the production of only mutant protein and is further limited by donor shortage, cost, and need for immunosuppression.
Antisense Oligonucleotides
Antisense oligonucleotides (ASOs) are under clinical investigation for their ability to inhibit hepatic expression of amyloidogenic TTR protein. Currently, the ASO compound, ISIS-TTRRx, is under investigation in a phase 3 multicenter, randomized, double-blind, placebo-controlled clinical trial in patients with familial amyloid polyneuropathy (FAP).11 The primary objective is to evaluate its efficacy as measured by change in neuropathy from baseline relative to placebo. Secondary measures will evaluate quality of life (QOL), modified body mass index (mBMI) by albumin, and pharmacodynamic effects on retinol binding protein. Exploratory objectives in a subset of patients with LV wall thickness ≥13 mm without a history of persistent hypertension will examine echocardiographic parameters, N-terminal pro–B-type natriuretic peptide (NT-proBNP), and polyneuropathy disability score relative to placebo. These data will facilitate analysis of the effect of antisense oligonucleotide-mediated TTR suppression on the TTR cardiac phenotype with a phase 3 trial anticipated to begin enrollment in 2016.
TTR Stabilizers (Diflunisal, Tafamidis)
Diflunisal
Several TTR-stabilizing agents are in various stages of clinical trials. Diflunisal, a traditionally used and generically available nonsteroidal anti-inflammatory drug (NSAID), binds and stabilizes familial TTR variants against acid-mediated fibril formation in vitro and is now in human clinical trials.12,13 The use of diflunisal in patients with TTR cardiac amyloidosis is controversial given complication of chronic inhibition of cyclooxygenase (COX) enzymes, including gastrointestinal bleeding, renal dysfunction, fluid retention, and hypertension that may precipitate or exacerbate heart failure in vulnerable individuals.14-17 In TTR cardiac amyloidosis, an open-label cohort study suggested that low-dose diflunisal with careful monitoring along with a prophylactic proton pump inhibitor could be safely administered to compensated patients.18 An association was observed, however, between chronic diflunisal use and adverse changes in renal function suggesting that advanced kidney disease may be prohibitive in diflunisal therapy.In FAP patients with peripheral or autonomic neuropathy randomized to diflunisal or placebo, diflunisal slowed progression of neurologic impairment and preserved QOL over two years of follow-up.19 Echocardiography demonstrated cardiac involvement in approximately 50% of patients.20 Longer-term safety and efficacy data over an average 38 ± 31 months in 40 Japanese patients with hereditary ATTR amyloidosis who were not candidates for liver transplantation showed that diflunisal was mostly well tolerated.12 The authors cautioned the need for attentive monitoring of renal function and blood cell counts. Larger multicenter collaborations are needed to determine diflunisal’s true efficacy in HFPEF patients with TTR cardiac amyloidosis.
Tafamidis
Tafamidis is under active investigation as a novel compound that binds to the thyroxine-binding sites of the TTR tetramer, inhibiting its dissociation into monomers and blocking the rate-limiting step in the TTR amyloidogenesis cascade.21 The TTR compound was shown in an 18-month double-blind, placebo-controlled trial to slow progression of neurologic symptoms in patients with early-stage ATTRm due to the V30M mutation.22 When focusing on cardiomyopathy in a phase 2, open-label trial, tafamidis also appeared to effectively stabilize TTR tetramers in non-V30M variants, wild-type and V122I, as well as biochemical and echocardiographic parameters.23,24 Preliminary data suggests that clinically stabilized patients had shorter disease duration, lower cardiac biomarkers, less myocardial thickening, and higher EF than those who were not stabilized, suggesting early institution of therapy may be beneficial. A phase 3 trial has completed enrollment and will evaluate the efficacy, safety, and tolerability of tafamidis 20 or 80 mg orally vs. placebo.25 This will contribute to long-term safety and efficacy data needed to determine the therapeutic effects of tafamidis among ATTRm variants.
Amyloid Degraders (Doxycycline/TUDCA and Anti-SAP Antibodies)
Doxycycline/TUDCA
While silencer and stabilizer drugs are aimed at lowering amyloidogenic precursor protein production, they cannot remove already deposited fibrils in an infiltrated heart. Removal of already deposited fibrils by amyloid degraders would be an important therapeutic strategy, particularly in older adults with heavily infiltrated hearts reflected by thick walls, HFPEF, systolic heart failure, and restrictive cardiomyopathy. Combined doxycycline and tauroursodeoxycholic acid (TUDCA) disrupt TTR amyloid fibrils and appeared to have an acceptable safety profile in a small phase 2 open-label study among 20 TTR patients. No serious adverse reactions or clinical progression of cardiac or neuropathic involvement was observed over one year.26 An active phase 2, single-center, open-label, 12-month study will assess primary outcome measures including mBMI, neurologic impairment score, and NT-proBNP.27 Another phase 2 study is examining the tolerability and efficacy of doxycycline/TUDCA over an 18-month period in patients with TTR amyloid cardiomyopathy.28 Additionally, a study in patients with TTR amyloidosis is ongoing to determine the effect of doxycycline alone on neurologic function, cardiac biomarkers, echocardiographic parameters, modified body mass index, and autonomic neuropathy.29
Anti-SAP Antibodies
In order to safely clear established amyloid deposits, the role of the normal, nonfibrillar plasma glycoprotein present in all human amyloid deposits, serum amyloid P component (SAP), needs to be more clearly understood.30 In mice with amyloid AA type deposits, administration of antihuman SAP antibody triggered a potent giant cell reaction that removed massive visceral amyloid deposits without adverse effects.31 In humans with TTR cardiac amyloidosis, anti-SAP antibody treatments could be feasible because the bis-D proline compound, CPHPC, is capable of clearing circulating human SAP, which allow anti-SAP antibodies to reach residual deposited SAP. In a small, open-label, single-dose-escalation, phase 1 trial involving 15 patients with systemic amyloidosis, none of whom had clinical evidence of cardiac amyloidosis, were treated with CPHPC followed by human monoclonal IgG1 anti-SAP antibody.32 No serious adverse events were reported and amyloid deposits were cleared from the liver, kidney, and lymph node. Anti-SAP antibodies hold promise as a potential amyloid therapy because of their potential to target all forms of amyloid deposits across multiple tissue types.
Mutant or wild-type TTR cardiac amyloidoses are increasingly recognized as a cause of HFPEF. Clinicians need to be aware of this important HFPEF etiology because the diverse array of emerging disease-modifying agents for TTR cardiac amyloidosis in human clinical trials has the potential to herald a new era for the treatment of HFPEF.
References
Maurer MS, Mancini D. HFpEF: is splitting into distinct phenotypes by comorbidities the pathway forward? J Am Coll Cardiol 2014;64:550-2.
Mohammed SF, Mirzoyev SA, Edwards WD, et al. Left ventricular amyloid deposition in patients with heart failure and preserved ejection fraction. JACC Heart Fail 2014;2:113-22.
González-López E, Gallego-Delgado M, Guzzo-Merello G, et al. Wild-type transthyretin amyloidosis as a cause of heart failure with preserved ejection fraction. Eur Heart J 2015.
Castano A, Bokhari S, Maurer MS. Unveiling wild-type transthyretin cardiac amyloidosis as a significant and potentially modifiable cause of heart failure with preserved ejection fraction. Eur Heart J 2015 Jul 28. [Epub ahead of print]
Rapezzi C, Merlini G, Quarta CC, et al. Systemic cardiac amyloidoses: disease profiles and clinical courses of the 3 main types. Circulation 2009;120:1203-12.
Bokhari S, Castano A, Pozniakoff T, Deslisle S, Latif F, Maurer MS. (99m)Tc-pyrophosphate scintigraphy for differentiating light-chain cardiac amyloidosis from the transthyretin-related familial and senile cardiac amyloidoses. Circ Cardiovasc Imaging 2013;6:195-201.
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998;391:806-11.
Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001;411:494-8.
Kanasty R, Dorkin JR, Vegas A, Anderson D. Delivery materials for siRNA therapeutics. Nature Mater 2013;12:967-77.
U.S. National Institutes of Health. Phase 2 Study to Evaluate ALN-TTRSC in Patients With Transthyretin (TTR) Cardiac Amyloidosis (ClinicalTrials.gov website). 2014. Available at: https://www.clinicaltrials.gov/ct2/show/NCT01981837. Accessed 8/19/2015.
U.S. National Institutes of Health. Efficacy and Safety of ISIS-TTRRx in Familial Amyloid Polyneuropathy (Clinical Trials.gov Website. 2013. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01737398. Accessed 8/19/2015.
Sekijima Y, Dendle MA, Kelly JW. Orally administered diflunisal stabilizes transthyretin against dissociation required for amyloidogenesis. Amyloid 2006;13:236-49.
Tojo K, Sekijima Y, Kelly JW, Ikeda S. Diflunisal stabilizes familial amyloid polyneuropathy-associated transthyretin variant tetramers in serum against dissociation required for amyloidogenesis. Neurosci Res 2006;56:441-9.
Epstein M. Non-steroidal anti-inflammatory drugs and the continuum of renal dysfunction. J Hypertens Suppl 2002;20:S17-23.
Wallace JL. Pathogenesis of NSAID-induced gastroduodenal mucosal injury. Best Pract Res Clin Gastroenterol 2001;15:691-703.
Mukherjee D, Nissen SE, Topol EJ. Risk of cardiovascular events associated with selective COX-2 inhibitors. JAMA 2001;286:954-9.
Page J, Henry D. Consumption of NSAIDs and the development of congestive heart failure in elderly patients: an underrecognized public health problem. Arch Intern Med 2000;160:777-84.
Castano A, Helmke S, Alvarez J, Delisle S, Maurer MS. Diflunisal for ATTR cardiac amyloidosis. Congest Heart Fail 2012;18:315-9.
Berk JL, Suhr OB, Obici L, et al. Repurposing diflunisal for familial amyloid polyneuropathy: a randomized clinical trial. JAMA 2013;310:2658-67.
Quarta CCF, Solomon RH Suhr SD, et al. The prevalence of cardiac amyloidosis in familial amyloidotic polyneuropathy with predominant neuropathy: The Diflunisal Trial. International Symposium on Amyloidosis 2014:88-9.
Hammarstrom P, Jiang X, Hurshman AR, Powers ET, Kelly JW. Sequence-dependent denaturation energetics: A major determinant in amyloid disease diversity. Proc Natl Acad Sci U S A 2002;99 Suppl 4:16427-32.
Coelho T, Maia LF, Martins da Silva A, et al. Tafamidis for transthyretin familial amyloid polyneuropathy: a randomized, controlled trial. Neurology 2012;79:785-92.
Merlini G, Plante-Bordeneuve V, Judge DP, et al. Effects of tafamidis on transthyretin stabilization and clinical outcomes in patients with non-Val30Met transthyretin amyloidosis. J Cardiovasc Transl Res 2013;6:1011-20.
Maurer MS, Grogan DR, Judge DP, et al. Tafamidis in transthyretin amyloid cardiomyopathy: effects on transthyretin stabilization and clinical outcomes. Circ Heart Fail 2015;8:519-26.
U.S. National Institutes of Health. Safety and Efficacy of Tafamidis in Patients With Transthyretin Cardiomyopathy (ATTR-ACT) (ClinicalTrials.gov website). 2014. Available at: http://www.clinicaltrials.gov/show/NCT01994889. Accessed 8/19/2015.
Obici L, Cortese A, Lozza A, et al. Doxycycline plus tauroursodeoxycholic acid for transthyretin amyloidosis: a phase II study. Amyloid 2012;19 Suppl 1:34-6.
U.S. National Institutes of Health. Safety, Efficacy and Pharmacokinetics of Doxycycline Plus Tauroursodeoxycholic Acid in Transthyretin Amyloidosis (ClinicalTrials.gov website). 2011. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01171859. Accessed 8/19/2015.
U.S. National Institutes of Health. Tolerability and Efficacy of a Combination of Doxycycline and TUDCA in Patients With Transthyretin Amyloid Cardiomyopathy (ClinicalTrials.gov website). 2013. Available at: http://www.clinicaltrials.gov/ct2/show/NCT01855360. Accessed 8/19/2015.
U.S. National Institutes of Health. Safety and Effect of Doxycycline in Patients With Amyloidosis (ClinicalTrials.gov website).2015. Available at: https://clinicaltrials.gov/ct2/show/NCT01677286. Accessed 8/19/2015.
Pepys MB, Dash AC. Isolation of amyloid P component (protein AP) from normal serum as a calcium-dependent binding protein. Lancet 1977;1:1029-31.
Bodin K, Ellmerich S, Kahan MC, et al. Antibodies to human serum amyloid P component eliminate visceral amyloid deposits. Nature 2010;468:93-7.
Richards DB, Cookson LM, Berges AC, et al. Therapeutic Clearance of Amyloid by Antibodies to Serum Amyloid P Component. N Engl J Med 2015;373:1106-14.
…
The Acid-Mediated Denaturation Pathway of Transthyretin Yields a Conformational Intermediate That Can Self-Assemble into Amyloid†
Transthyretin (TTR) amyloid fibril formation is observed during partial acid denaturation and while refolding acid-denatured TTR, implying that amyloid fibril formation results from the self-assembly of a conformational intermediate. The acid denaturation pathway of TTR has been studied in detail herein employing a variety of biophysical methods to characterize the intermediate(s) capable of amyloid fibril formation. At physiological concentrations, tetrameric TTR remains associated from pH 7 to pH 5 and is incapable of amyloid fibril formation. Tetrameric TTR dissociates to a monomer in a process that is dependent on both pH and protein concentration below pH 5. The extent of amyloid fibril formation correlates with the concentration of the TTR monomer having an altered, but defined, tertiary structure over the pH range of 5.0−3.9. The inherent Trp fluorescence-monitored denaturation curve of TTR exhibits a plateau over the pH range where amyloid fibril formation is observed (albeit at a higher concentration), implying that a steady-state concentration of the amyloidogenic intermediate with an altered tertiary structure is being detected. Interestingly, 1-anilino-8-naphthalenesulfonate fluorescence is at a minimum at the pH associated with maximal amyloid fibril formation (pH 4.4), implying that the amyloidogenic intermediate does not have a high extent of hydrophobic surface area exposed, consistent with a defined tertiary structure. Transthyretin has two Trp residues in its primary structure, Trp-41 and Trp-79, which are conveniently located far apart in the tertiary structure of TTR. Replacement of each Trp with Phe affords two single Trp containing variants which were used to probe local pH-dependent tertiary structural changes proximal to these chromophores. The pH-dependent fluorescence behavior of the Trp-79-Phe mutant strongly suggests that Trp-41 is located near the site of the tertiary structural rearrangement that occurs in the formation of the monomeric amyloidogenic intermediate, likely involving the C-strand−loop−D-strand region. Upon further acidification of TTR (below pH 4.4), the structurally defined monomeric amyloidogenic intermediate begins to adopt alternative conformations that are not amyloidogenic, ultimately forming an A-state conformation below pH 3 which is also not amyloidogenic. In summary, analytical equilibrium ultracentrifugation, SDS−PAGE, far- and near-UV CD, fluorescence, and light scattering studies suggest that the amyloidogenic intermediate is a monomeric predominantly β-sheet structure having a well-defined tertiary structure.
Prevention of Transthyretin Amyloid Disease by Changing Protein Misfolding Energetics
Per Hammarström*, R. Luke Wiseman*, Evan T. Powers, Jeffery W. Kelly†+ Author Affiliations
Genetic evidence suggests that inhibition of amyloid fibril formation by small molecules should be effective against amyloid diseases. Known amyloid inhibitors appear to function by shifting the aggregation equilibrium away from the amyloid state. Here, we describe a series of transthyretin amyloidosis inhibitors that functioned by increasing the kinetic barrier associated with misfolding, preventing amyloidogenesis by stabilizing the native state. The trans-suppressor mutation, threonine 119 → methionine 119, which is known to ameliorate familial amyloid disease, also functioned through kinetic stabilization, implying that this small-molecule strategy should be effective in treating amyloid diseases.
Rational design of potent human transthyretin amyloid disease inhibitors
Thomas Klabunde1,2, H. Michael Petrassi3, Vibha B. Oza3, Prakash Raman3, Jeffery W. Kelly3 & James C. Sacchettini1
The human amyloid disorders, familial amyloid polyneuropathy, familial amyloid cardiomyopathy and senile systemic amyloidosis, are caused by insoluble transthyretin (TTR) fibrils, which deposit in the peripheral nerves and heart tissue. Several nonsteroidal anti-inflammatory drugs and structurally similar compounds have been found to strongly inhibit the formation of TTR amyloid fibrils in vitro. These include flufenamic acid, diclofenac, flurbiprofen, and resveratrol. Crystal structures of the protein–drug complexes have been determined to allow detailed analyses of the protein–drug interactions that stabilize the native tetrameric conformation of TTR and inhibit the formation of amyloidogenic TTR. Using a structure-based drug design approach ortho-trifluormethylphenyl anthranilic acid and N-(meta-trifluoromethylphenyl) phenoxazine 4,6-dicarboxylic acid have been discovered to be very potent and specific TTR fibril formation inhibitors. This research provides a rationale for a chemotherapeutic approach for the treatment of TTR-associated amyloid diseases.
First European consensus for diagnosis, management, and treatment of transthyretin familial amyloid polyneuropathy
Adams, Davida; Suhr, Ole B.b; Hund, Ernstc; Obici, Laurad; Tournev, Ivailoe,f; Campistol, Josep M.g; Slama, Michel S.h; Hazenberg, Bouke P.i; Coelho, Teresaj; from the European Network for TTR-FAP (ATTReuNET)
Purpose of review: Early and accurate diagnosis of transthyretin familial amyloid polyneuropathy (TTR-FAP) represents one of the major challenges faced by physicians when caring for patients with idiopathic progressive neuropathy. There is little consensus in diagnostic and management approaches across Europe.
Recent findings: The low prevalence of TTR-FAP across Europe and the high variation in both genotype and phenotypic expression of the disease means that recognizing symptoms can be difficult outside of a specialized diagnostic environment. The resulting delay in diagnosis and the possibility of misdiagnosis can misguide clinical decision-making and negatively impact subsequent treatment approaches and outcomes.
Summary: This review summarizes the findings from two meetings of the European Network for TTR-FAP (ATTReuNET). This is an emerging group comprising representatives from 10 European countries with expertise in the diagnosis and management of TTR-FAP, including nine National Reference Centres. The current review presents management strategies and a consensus on the gold standard for diagnosis of TTR-FAP as well as a structured approach to ongoing multidisciplinary care for the patient. Greater communication, not just between members of an individual patient’s treatment team, but also between regional and national centres of expertise, is the key to the effective management of TTR-FAP.
Transthyretin familial amyloid polyneuropathy (TTR-FAP) is a highly debilitating and irreversible neurological disorder presenting symptoms of progressive sensorimotor and autonomic neuropathy [1▪,2▪,3]. TTR-FAP is caused by misfolding of the transthyretin (TTR) protein leading to protein aggregation and the formation of amyloid fibrils and, ultimately, to amyloidosis (commonly in the peripheral and autonomic nervous system and the heart) [4,5]. TTR-FAP usually proves fatal within 7–12 years from the onset of symptoms, most often due to cardiac dysfunction, infection, or cachexia [6,7▪▪].
The prevalence and disease presentation of TTR-FAP vary widely within Europe. In endemic regions (northern Portugal, Sweden, Cyprus, and Majorca), patients tend to present with a distinct genotype in large concentrations, predominantly a Val30Met substitution in the TTR gene [8–10]. In other areas of Europe, the genetic footprint of TTR-FAP is more varied, with less typical phenotypic expression [6,11]. For these sporadic or scattered cases, a lack of awareness among physicians of variable clinical features and limited access to diagnostic tools (i.e., pathological studies and genetic screening) can contribute to high rates of misdiagnosis and poorer patient outcomes [1▪,11]. In general, early and late-onset variants of TTR-FAP, found within endemic and nonendemic regions, present several additional diagnostic challenges [11,12,13▪,14].
Delay in the time to diagnosis is a major obstacle to the optimal management of TTR-FAP. With the exception of those with a clearly diagnosed familial history of FAP, patients still invariably wait several years between the emergence of first clinical signs and accurate diagnosis [6,11,14]. The timely initiation of appropriate treatment is particularly pertinent, given the rapidity and irreversibility with which TTR-FAP can progress if left unchecked, as well as the limited effectiveness of available treatments during the later stages of the disease [14]. This review aims to consolidate the existing literature and present an update of the best practices in the management of TTR-FAP in Europe. A summary of the methods used to achieve a TTR-FAP diagnosis is presented, as well as a review of available treatments and recommendations for treatment according to disease status.
Patients with TTR-FAP can present with a range of symptoms [11], and care should be taken to acquire a thorough clinical history of the patient as well as a family history of genetic disease. Delay in diagnosis is most pronounced in areas where TTR-FAP is not endemic or when there is no positive family history [1▪]. TTR-FAP and TTR-familial amyloid cardiomyopathy (TTR-FAC) are the two prototypic clinical disease manifestations of a broader disease spectrum caused by an underlying hereditary ATTR amyloidosis [19]. In TTR-FAP, the disease manifestation of neuropathy is most prominent and definitive for diagnosis, whereas cardiomyopathy often suggests TTR-FAC. However, this distinction is often superficial because cardiomyopathy, autonomic neuropathy, vitreous opacities, kidney disease, and meningeal involvement all may be present with varying severity for each patient with TTR-FAP.
Among early onset TTR-FAP with usually positive family history, symptoms of polyneuropathy present early in the disease process and usually predominate throughout the progression of the disease, making neurological testing an important diagnostic aid [14]. Careful clinical examination (e.g., electromyography with nerve conduction studies and sympathetic skin response, quantitative sensation test, quantitative autonomic test) can be used to detect, characterize, and scale the severity of neuropathic abnormalities involving small and large nerve fibres [10]. Although a patient cannot be diagnosed definitively with TTR-FAP on the basis of clinical presentation alone, symptoms suggesting the early signs of peripheral neuropathy, autonomic dysfunction, and cardiac conduction disorders or infiltrative cardiomyopathy are all indicators that further TTR-FAP diagnostic investigation is warranted. Late-onset TTR-FAP often presents as sporadic cases with distinct clinical features (e.g., milder autonomic dysfunction) and can be more difficult to diagnose than early-onset TTR-FAP (Table 2) [1▪,11,12,13▪,14,20].
Genetic testing is carried out to allow detection of specific amyloidogenic TTR mutations (Table 1), using varied techniques depending on the expertise and facilities available in each country (Table S2, http://links.lww.com/CONR/A39). A targeted approach to detect a specific mutation can be used for cases belonging to families with previous diagnosis. In index cases of either endemic and nonendemic regions that do not have a family history of disease, are difficult to confirm, and have atypical symptoms, TTR gene sequencing is required for the detection of both predicted and new amyloidogenic mutations [26,27].
Following diagnosis, the neuropathy stage and systemic extension of the disease should be determined in order to guide the next course of treatment (Table 4) [3,30,31]. The three stages of TTR-FAP severity are graded according to a patient’s walking disability and degree of assistance required [30]. Systemic assessment, especially of the heart, eyes, and kidney, is also essential to ensure all aspects of potential impact of the disease can be detected [10].
The goals of cardiac investigations are to detect serious conduction disorders with the risk of sudden death and infiltrative cardiomyopathy. Electrocardiograms (ECG), Holter-ECG, and intracardiac electrophysiology study are helpful to detect conduction disorders. Echocardiograms, cardiac magnetic resonance imaging, scintigraphy with bone tracers, and biomarkers (e.g., brain natriuretic peptide, troponin) can all help to diagnose infiltrative cardiomyopathy[10]. An early detection of cardiac abnormalities has obvious benefits to the patient, given that the prophylactic implantation of pacemakers was found to prevent 25% of major cardiac events in TTR-FAP patients followed up over an average of 4 years [32▪▪]. Assessment of cardiac denervation with 123-iodine meta-iodobenzylguanidine is a powerful prognostic marker in patients diagnosed with FAP [33].
…..
Tafamidis
Tafamidis is a first-in-class therapy that slows the progression of TTR amyloidogenesis by stabilizing the mutant TTR tetramer, thereby preventing its dissociation into monomers and amyloidogenic and toxic intermediates [55,56]. Tafamidis is currently indicated in Europe for the treatment of TTR amyloidosis in adult patients with stage I symptomatic polyneuropathy to delay peripheral neurological impairment [57].
In an 18-month, double-blind, placebo-controlled study of patients with early-onset Val30Met TTR-FAP, tafamidis was associated with a 52% lower reduction in neurological deterioration (P = 0.027), a preservation of nerve function, and TTR stabilization versus placebo [58▪▪]. However, only numerical differences were found for the coprimary endpoints of neuropathy impairment [neuropathy impairment score in the lower limb (NIS-LL) responder rates of 45.3% tafamidis vs 29.5% placebo; P = 0.068] and quality of life scores [58▪▪]. A 12-month, open-label extension study showed that the reduced rates of neurological deterioration associated with tafamidis were sustained over 30 months, with earlier initiation of tafamidis linking to better patient outcomes (P = 0.0435) [59▪]. The disease-slowing effects of tafamidis may be dependent on the early initiation of treatment. In an open-label study with Val30Met TTR-FAP patients with late-onset and advanced disease (NIS-LL score >10, mean age 56.4 years), NIS-LL and disability scores showed disease progression despite 12 months of treatment with tafamidis, marked by a worsening of neuropathy stage in 20% and the onset of orthostatic hypotension in 22% of patients at follow-up [60▪].
Tafamidis is not only effective in patients exhibiting the Val30Met mutation; it also has proven efficacy, in terms of TTR stabilization, in non-Val30Met patients over 12 months [61]. Although tafamidis has demonstrated safe use in patients with TTR-FAP, care should be exercised when prescribing to those with existing digestive problems (e.g., diarrhoea, faecal incontinence) [60▪].
Diflunisal is a nonsteroidal anti-inflammatory drug (NSAID) that, similar to tafamidis, slows the rate of amyloidogenesis by preventing the dissociation, misfolding, and misassembly of the mutated TTR tetramer [62,63]. Off-label use has been reported for patients with stage I and II disease, although diflunisal is not currently licensed for the treatment of TTR-FAP.
Evidence for the clinical effectiveness of diflunisal in TTR-FAP derives from a placebo-controlled, double-blind, 24-month study in 130 patients with clinically detectable peripheral or autonomic neuropathy[64▪]. The deterioration in NIS scores was significantly more pronounced in patients receiving placebo compared with those taking diflunisal (P = 0.001), and physical quality of life measures showed significant improvement among diflunisal-treated patients (P = 0.001). Notable during this study was the high rate of attrition in the placebo group, with 50% more placebo-treated patients dropping out of this 2-year study as a result of disease progression, advanced stage of the disease, and varied mutations.
One retrospective analysis of off-label use of diflunisal in patients with TTR-FAP reported treatment discontinuation in 57% of patients because of adverse events that were largely gastrointestinal [65]. Conclusions on the safety of diflunisal in TTR-FAP will depend on further investigations on the impact of known cardiovascular and renal side-effects associated with the NSAID drug class [66,67].
When T cells receive the appropriate signals through the T cell receptor (TCR) complex and the costimulatory receptor CD28, a complex rearrangement of the cytoskeleton occurs that enables the formation of the immunological synapse, a specialized structure that forms between the antigen-presenting cell and the T cell. In this week’s issue, Roybal et al. used sophisticated imaging of live cells and computational image analysis to visualize the dynamic rearrangements of actin and various regulatory proteins in T cells activated through the TCR in the absence or presence of CD28 signaling. The regulatory proteins WAVE2 and cofilin were efficiently recruited to the immunological synapse only when both TCR and CD28 signaled. Fried et al. used a different fluorescence imaging approach, triple-color FRET (fluorescence resonance energy transfer), to visualize not the movement of proteins with the cell, but the interactions between the actin-regulatory proteins WASp and WIP in live cells. The WASp-WIP interaction is required for T cell activation. The triple-color FRET analysis revealed how changes in the interaction between WASp and WIP resulted in WASp functioning as both an on and off switch. Although most studies’ focus of T cell cytoskeletal dynamics is on the changes that occur at the immunological synapse, as González-Granado et al. showed, the nuclear cytoskeleton is also important for the immune response. In this study, live-cell imaging and immunofluorescence analysis revealed that nuclear lamin-A contributed to the polymerization of actin and, thus, immunological synapse formation. These studies highlight the insights that can be obtained about molecular dynamics from live-cell image analysis.
K. T. Roybal, T. E. Buck, X. Ruan, B. H. Cho, D. J. Clark, R. Ambler, H. M. Tunbridge, J. Zhang, P. Verkade, C. Wülfing, R. F. Murphy, Computational spatiotemporal analysis identifies WAVE2 and cofilin as joint regulators of costimulation-mediated T cell actin dynamics. Sci. Signal.9, rs3 (2016). [Abstract]
Computational spatiotemporal analysis identifies WAVE2 and cofilin as joint regulators of costimulation-mediated T cell actin dynamics
T cells must receive signals through the T cell receptor (TCR) and the costimulatory receptor CD28 to become fully activated. Critical to this process is the reorganization of plasma membrane actin at the immunological synapse, the interface between a T cell and an antigen-presenting cell. Roybal et al.imaged actin and fluorescently tagged actin regulatory proteins in T cells activated through the TCR in the absence or presence of CD28 signaling. Computational image processing to normalize differences in cell shape enabled tracking of the fluorescent proteins. The regulatory proteins WAVE2 and cofilin were efficiently recruited to the immunological synapse only when both TCR and CD28 signaled. Constitutive activation of either protein in TCR-stimulated T cells enabled normal actin reorganization even when CD28 signaling was blocked. This combination of imaging and computational analysis could be applied to other systems to determine the spatiotemporal dynamics of signaling molecules.
Fluorescence microscopy is one of the most important tools in cell biology research because it provides spatial and temporal information to investigate regulatory systems inside cells. This technique can generate data in the form of signal intensities at thousands of positions resolved inside individual live cells. However, given extensive cell-to-cell variation, these data cannot be readily assembled into three- or four-dimensional maps of protein concentration that can be compared across different cells and conditions. We have developed a method to enable comparison of imaging data from many cells and applied it to investigate actin dynamics in T cell activation. Antigen recognition in T cells by the T cell receptor (TCR) is amplified by engagement of the costimulatory receptor CD28. We imaged actin and eight core actin regulators to generate over a thousand movies of T cells under conditions in which CD28 was either engaged or blocked in the context of a strong TCR signal. Our computational analysis showed that the primary effect of costimulation blockade was to decrease recruitment of the activator of actin nucleation WAVE2 (Wiskott-Aldrich syndrome protein family verprolin-homologous protein 2) and the actin-severing protein cofilin to F-actin. Reconstitution of WAVE2 and cofilin activity restored the defect in actin signaling dynamics caused by costimulation blockade. Thus, we have developed and validated an approach to quantify protein distributions in time and space for the analysis of complex regulatory systems.
S. Fried, B. Reicher, M. H. Pauker, S. Eliyahu, O. Matalon, E. Noy, J. Chill, M. Barda-Saad, Triple-color FRET analysis reveals conformational changes in the WIP-WASp actin-regulating complex. Sci. Signal.7, ra60 (2014). [Abstract]
Triple-Color FRET Analysis Reveals Conformational Changes in the WIP-WASp Actin-Regulating Complex
Wiskott-Aldrich syndrome protein (WASp) is a key regulator of the actin cytoskeletal machinery. Binding of WASp-interacting protein (WIP) to WASp modulates WASp activity and protects it from degradation. Formation of the WIP-WASp complex is crucial for the adaptive immune response. We found that WIP and WASp interacted in cells through two distinct molecular interfaces. One interaction occurred between the WASp-homology-1 (WH1) domain of WASp and the carboxyl-terminal domain of WIP that depended on the phosphorylation status of WIP, which is phosphorylated by protein kinase C θ (PKCθ) in response to T cell receptor activation. The other interaction occurred between the verprolin homology, central hydrophobic region, and acidic region (VCA) domain of WASp and the amino-terminal domain of WIP. This latter interaction required actin, because it was inhibited by latrunculin A, which sequesters actin monomers. With triple-color fluorescence resonance energy transfer (3FRET) technology, we demonstrated that the WASp activation mechanism involved dissociation of the first interaction, while leaving the second interaction intact. This conformation exposed the ubiquitylation site on WASp, leading to degradation of WASp. Together, these data suggest that the activation and degradation of WASp are delicately balanced and depend on the phosphorylation state of WIP. Our molecular analysis of the WIP-WASp interaction provides insight into the regulation of actin-dependent processes.
J. M. González-Granado, C. Silvestre-Roig, V. Rocha-Perugini, L. Trigueros-Motos, D. Cibrián, G. Morlino, M. Blanco-Berrocal, F. G. Osorio, J. M. P. Freije, C. López-Otín, F. Sánchez-Madrid, V. Andrés, Nuclear envelope Lamin-a couples actin dynamics with immunological synapse architecture and T cell activation.Sci. Signal.7, ra37 (2014). [Abstract]
Nuclear Envelope Lamin-A Couples Actin Dynamics with Immunological Synapse Architecture and T Cell Activation
In many cell types, nuclear A-type lamins regulate multiple cellular functions, including higher-order genome organization, DNA replication and repair, gene transcription, and signal transduction; however, their role in specialized immune cells remains largely unexplored. We showed that the abundance of A-type lamins was almost negligible in resting naïve T lymphocytes, but was increased upon activation of the T cell receptor (TCR). The increase in lamin-A was an early event that accelerated formation of the immunological synapse between T cells and antigen-presenting cells. Polymerization of F-actin in T cells is a critical step for immunological synapse formation, and lamin-A interacted with the linker of nucleoskeleton and cytoskeleton (LINC) complex to promote F-actin polymerization. We also showed that lamin-A expression accelerated TCR clustering and led to enhanced downstream signaling, including extracellular signal–regulated kinase 1/2 (ERK1/2) signaling, as well as increased target gene expression. Pharmacological inhibition of the ERK pathway reduced lamin-A–dependent T cell activation. Moreover, mice lacking lamin-A in immune cells exhibited impaired T cell responses in vivo. These findings underscore the importance of A-type lamins for TCR activation and identify lamin-A as a previously unappreciated regulator of the immune response.
T cell activation by antigens involves the formation of a complex, highly dynamic, yet organized signaling complex at the site of the T cell receptors (TCRs). Srikanth et al. found that the lymphocyte-specific large guanosine triphosphatase of the Rab family CRACR2A-a associated with vesicles near the Golgi in unstimulated mouse and human CD4+ T cells. Upon TCR activation, these vesicles moved to the immunological synapse (the contact region between a T cell and an antigen-presenting cell). The guanine nucleotide exchange factor Vav1 at the TCR complex recruited CRACR2A-a to the complex. Without CRACR2A-a, T cell activation was compromised because of defective calcium and kinase signaling.
More than 60 members of the Rab family of guanosine triphosphatases (GTPases) exist in the human genome. Rab GTPases are small proteins that are primarily involved in the formation, trafficking, and fusion of vesicles. We showed that CRACR2A (Ca2+ release–activated Ca2+ channel regulator 2A) encodes a lymphocyte-specific large Rab GTPase that contains multiple functional domains, including EF-hand motifs, a proline-rich domain (PRD), and a Rab GTPase domain with an unconventional prenylation site. Through experiments involving gene silencing in cells and knockout mice, we demonstrated a role for CRACR2A in the activation of the Ca2+ and c-Jun N-terminal kinase signaling pathways in response to T cell receptor (TCR) stimulation. Vesicles containing this Rab GTPase translocated from near the Golgi to the immunological synapse formed between a T cell and a cognate antigen-presenting cell to activate these signaling pathways. The interaction between the PRD of CRACR2A and the guanidine nucleotide exchange factor Vav1 was required for the accumulation of these vesicles at the immunological synapse. Furthermore, we demonstrated that GTP binding and prenylation of CRACR2A were associated with its localization near the Golgi and its stability. Our findings reveal a previously uncharacterized function of a large Rab GTPase and vesicles near the Golgi in TCR signaling. Other GTPases with similar domain architectures may have similar functions in T cells.
Metastatic prostate cancer remains a highly lethal disease with no curative therapeutic options. A significant subset of patients with prostate cancer harbor either germline or somatic mutations in DNA repair enzyme genes such as BRCA1, BRCA2, or ATM. Emerging data suggest that drugs that target poly(adenosine diphosphate [ADP]–ribose) polymerase (PARP) enzymes may represent a novel and effective means of treating tumors with these DNA repair defects, including prostate cancers. Here we will review the molecular mechanism of action of PARP inhibitors and discuss how they target tumor cells with faulty DNA repair functions and transcriptional controls. We will review emerging data for the utility of PARP inhibition in the management of metastatic prostate cancer. Finally, we will place PARP inhibitors within the framework of precision medicine–based care of patients with prostate cancer.
In 2016, prostate cancer is expected to be diagnosed in 180,890 men, and 26,120 will die of metastatic disease.[1] While the majority of localized prostate cancers can be controlled with surgery and/or radiation, metastatic disease remains a lethal disease with no curative options. Moreover, prostate cancer is a heterogeneous disease that can be highly lethal but also slow and indolent, as reflected by a 10-year estimated survival of 17% (S9346 trial, unpublished data). The advent of affordable and efficient techniques for profiling tumors molecularly represents an unprecedented opportunity to better characterize the molecular factors that result in indolent and/or lethal disease and to tailor therapy accordingly. Many clinical trials are already underway to examine whether molecularly targeted therapies can improve outcomes.[2] In this review, we will specifically examine the molecular rationale for one of these targeted approaches, poly(adenosine diphosphate [ADP]–ribose) polymerase (PARP) inhibition, in prostate cancer. We will review how PARP inhibitors function as a class, review the molecular features that sensitize cancer cells to this therapy, and discuss the data supporting its potential for patients with prostate cancer. We will then outline a strategy for further development of PARP inhibitors in the prostate cancer field.
Metastatic prostate cancer is typically categorized as hormone-sensitive prostate cancer (HSPC), which responds to androgen ablation, or castration-resistant prostate cancer (CRPC), which develops resistance to gonadal suppression. Although bilateral orchiectomy is the historic gold-standard treatment for metastatic HSPC, gonadal suppression is currently accomplished with gonadotropin-releasing hormone agonists or antagonists with or without androgen receptor blockade. This approach remains the cornerstone of therapy for men with metastatic HSPC.[3] Emerging data from large phase III trials (CHAARTED and Systemic Therapy in Advancing or Metastatic Prostate Cancer: Evaluation of Drug Efficacy [STAMPEDE]) have also revealed a large survival benefit for the combination of docetaxel and androgen deprivation in metastatic HSPC.[4,5]
Despite these initially effective treatments, the vast majority of men with metastatic HSPC will progress to CRPC, which is the lethal stage of the disease. For these patients, several additional therapies provide benefit by further suppression of androgen signaling (enzalutamide, abiraterone), disruption of the cell cycle in replicating cells (docetaxel, cabazitaxel), targeting of bone metastases (radium-223), or activation of antitumor immunologic response (sipuleucel-T).[6] While these therapies have undoubtedly extended the median survival of patients with metastatic CRPC, their impact on survival is modest and they clearly do not work for all men. In addition, we lack validated genomic markers that would allow better selection of patients for these therapies. Therefore, a better approach that leverages the individual and unique aspects of a patient’s cancer and utilizes therapy based on these factors may allow us to improve patient outcomes.
The development of high-throughput sequencing technology has made it feasible to comprehensively analyze the genetic mutations and gene expression changes in individual prostate cancers with a high degree of resolution in real time. Many institutions now routinely perform these analyses in the hope that they might uncover molecular features that predict response to certain therapies or provide guidance for clinical trial selection.[7] This approach, colloquially termed “precision” medicine, offers the potential promise of providing the right therapy for the right patient at the right time. In the context of prostate cancer, it means molecularly characterizing a tumor and then offering patients drugs that may specifically promote tumor lethality based on these molecular features. The limitation of this approach is that it requires that the target be truly biologically relevant and that there are drugs that can effectively target these molecular changes. The discovery of both somatic and germline DNA repair deficiencies in prostate cancer, together with the development of PARP inhibitors that can kill cancer cells with these defects, is a potent example of targeting therapy to molecularly defined tumor subtypes. While much early work validating this approach has occurred in breast and ovarian cancer populations, emerging data suggest that PARP inhibition is a potentially important strategy for managing a significant subset of prostate cancer patients.
PARP Inhibition: Targeting DNA Repair Deficiency
Molecular mechanism
PARP1 catalyzes the addition of poly(ADP)-ribose (PAR) groups to target proteins in a process termed PARylation.[8] PARP1 is part of a superfamily of proteins that consists of 18 members (including the related tankyrase enzymes), which have many functions within normal and cancer cells. PARP1, the founding member of this family, is responsible for the majority of PARylation of protein targets within cells. It is primarily present in the nucleus in association with chromatin, where it participates in DNA repair and regulation of gene expression by modulating protein localization and activity.[9]
DNA damage occurs continuously in all living cells as a result of oxidative damage or DNA replicative stress.[10] When DNA damage occurs on one strand of the DNA double helix, a single-strand break (SSB) results, but if two SSBs occur in close proximity and on opposite strands, the result is a double-strand break (DSB) and discontinuity of the chromosome (Figures 1 and 2). Even a single DSB is lethal to a human cell if unrepaired because of the risk of large-scale loss of genetic information.
PARP1 plays a critical role in restoration of genomic integrity by facilitating efficient repair of DNA SSBs and DSBs. PARP1 senses DNA damage by binding to the site of SSBs and DSBs and inducing auto-PARylation, which in turn promotes recruitment of DNA repair factors (such as DNA ligase III, polymerase β, and x-ray repair cross-complementing protein 1[XRCC1]).[11] Loss of PARP1 function by means of pharmacologic or genetic mechanisms results in impaired SSB repair and, following initiation of DNA replication, creation of a DNA DSB (see Figure 1). PARP may also play an important role in DSB repair and is known to recruit the MRE11-RAD50-NBS1 complex and to promote PARylation of BRCA1, factors required for the homologous recombination (HR) pathway of DNA DSB repair. Therefore, pharmacologic inhibition of PARP1/2 in DNA repair–defective (DRD) cells that lack efficient HR repair capabilities (such as those harboring BRCA1, BRCA2, or ATM mutations) results in failure to resolve SSBs, which are then converted to DSBs that promote cellular death.
The activity of PARP1 is not limited to DNA damage response. PARP1 is also known to regulate gene expression by modulation of transcription factor activity and regulation of chromatin.[12] PARP1 binds to RNA polymerase II, regulating gene expression, and may also affect tumor suppressor and oncogenic gene expression. PARP1 can also modulate hormone-dependent gene transcription from hormone-responsive nuclear receptors, such as estrogen receptors α and β, progesterone receptor, and androgen receptor.[9]
Furthermore, PARP1 can modulate the transcriptional activity of ETS transcription factors, which suggests that pharmacologic targeting of PARP1 may be useful in TMPRSS2:ERG fusion–positive prostate cancer cells (~50% of prostate cancers).[13] PARP1 physically interacts with the TMPRSS2:ERG gene fusion and the DNA–protein kinase complex, and these interactions are required for ERG-related gene transcription. Interestingly, PARP inhibition with olaparib inhibited prostate cancer xenograft growth if tumors harbored a TMPRSS2:ERG fusion, which suggests that PARP might represent a therapeutic option for prostate cancer patients with TMPRSS2:ERG fusions.[13] This concept is being evaluated in a recently completed clinical trial (National Cancer Institute [NCI] 9012).
PARP inhibitors
Given the biologic importance of PARP1 in the context of cancer, several pharmacologic agents that target this enzyme are currently under development (Table).
Most PARP inhibitors mimic the NAD+ substrate of PARP1, competitively bind to the catalytic domain, and inhibit PAR synthesis.[14] PARP inhibitors require the expression of PARP1 and PARP2, and cells that lack expression of both genes are not sensitive to these agents. PARP inhibitors all appear to block catalytic activity and PAR synthesis in a roughly equivalent manner but may show differential ability to trap PARP1/2 at the site of DNA damage (niraparib > olaparib > veliparib), an event that blocks repair and promotes cellular lethality.[15,16] Whether these effects observed in vitro translate into clinically meaningful differences in efficacy is less clear. Furthermore, it is also now clear that the putative PARP inhibitor iniparib may not promote cytotoxicity via PARP inhibition. Several initial studies focused on iniparib, but when phase III trials failed to demonstrate the efficacy of this compound, additional mechanistic work demonstrated that iniparib may not truly be an effective PARP inhibitor.[17,18] These data illustrate the necessity of careful mechanistic characterization of any targeted agent prior to large-scale and expensive studies.
Germline DNA repair deficiency
Inherited defects in DNA repair pathways result in increased susceptibility to the development of malignancy.[19] Defects in mismatch repair proteins promote the development of tumors, including colon and uterine,[20] whereas inherited inactivating mutations in BRCA1 and BRCA2, which are required for efficient HR-based DNA DSB repair, significantly increase the risk of breast, ovarian, prostate, and other cancers.[21] Patients with these tumor types typically demonstrate homozygous inactivation of these genes, the first event occurring in the germline, with subsequent clonal somatic inactivation of the remaining allele.[21] These events presumably occur early in tumorigenesis and, by loss of robust DNA DSB repair, induce genomic instability, which causes loss of tumor suppressors, activation of oncogenes, and acceleration of tumorigenesis.
A germline mutation in BRCA1 or BRCA2 increases the risk of prostate cancer and thus may be found in 2% to 5% of prostate cancers.[22,23] The relative risk of development of prostate cancer for men ≤ age 65 with BRCA1 mutations is 1.8, but BRCA2 mutations in particular seem to increase the risk of prostate cancer formation by age 65 by about 8.6-fold. Mutations of BRCA1, BRCA2, and ATM (and perhaps other DNA repair genes) may also play a role in progression to the lethal castration-resistant state.[22,24-26] The frequency of BRCA2 germline mutations in prostate cancer alone may be as high as 2%.[22] Therefore, the development of therapies to target DNA repair is likely to benefit a relatively large and relatively young population.
Somatic DNA repair deficiency
In addition to germline defects, tumors can acquire defective DNA repair processes through somatic loss of DNA damage response genes, and these somatic mutations can also confer sensitivity to PARP inhibition.[27] This has led to the concept of “BRCAness,” which refers to somatically acquired defects in HR that, as a group, could predict tumor response to PARP inhibitors and cisplatin.[21] Somatic alterations can include either acquired mutations or epigenetic events that silence genes such as ATM; ATR; BRCA1 or -2; CHEK1 or -2; FANCA, -C, -D2, -E, -F; PALB2; MRE11 complex; or RAD51, which prevent efficient HR repair of DNA DSBs.
It is likely that a substantial proportion of men with prostate cancer may demonstrate aspects of BRCAness that could predict sensitivity to PARP inhibitors. Beltran et al performed targeted next-generation sequencing of tumors from men with advanced prostate cancer and found that 12% demonstrated BRCA2 loss and that 8% harbored ATM loss.[28] Furthermore, up to 19.3% of CRPCs demonstrate aberrations in BRCA1, BRCA2, or ATM; these events become more frequent as the disease progresses from hormone-sensitive to castration-resistant.[29] Together these data suggest that BRCAness is a reasonably frequent event in patients with advanced prostate cancer, which makes PARP inhibition an attractive target in this disease.
Synthetic lethality
The concept of promoting the killing of cancer cells by simultaneously blocking SSB repair using PARP inhibition in cells that lack efficient DSB repair is called “synthetic lethality.” In this scenario, tumor cells may harbor either germline or somatically acquired homozygous inactivation of HR. Germline defects (when present) typically affect only one allele in normal cells, and therefore normal tissues retain HR function. This difference between the DNA repair capacity of normal and cancer cells can be leveraged to produce selective cell killing of tumor cells by PARP inhibitors. Treatment of patients with PARP inhibitors will then block normal SSB repair in all cells, and these SSBs are subsequently converted to DSBs by DNA replication. In normal cells, HR restores the genome and allows survival, but in DRD cancer cells, DSBs persist, inducing cellular death selectively in the tumor cell population (see Figure 2).
PARP Inhibition in Prostate Cancer
Early-phase studies
Ample data indicate that PARP inhibitors possess antitumor activity within diverse patient populations, particularly those with BRCA1 or BRCA2 mutations.[14] One of the first studies to validate the concept of clinical benefit in patients with BRCA mutations was a phase I trial that looked at pharmacokinetic and pharmacodynamic aspects of olaparib treatment.[24] In this study, 60 patients with solid tumors were treated with various doses of olaparib (10 mg daily to 600 mg twice daily) to determine maximum tolerated dose (MTD). The study population was intentionally enriched for BRCA mutation carriers, and 22 patients of the cohort harbored BRCA1 or BRCA2 mutations. Objective tumor activity was observed in the mutation carrier population in patients with breast, ovarian, and prostate cancers. Three patients with advanced prostate cancer were included in this study cohort; the one with a BRCA2 mutation had a greater than 50% response in prostate-specific antigen (PSA) level, resolution of bone metastases, and an extended treatment course. This study suggested that there was a benefit of olaparib therapy in BRCA mutation carriers and the potential for benefit in prostate cancer patients. Further validation of olaparib efficacy in patients with BRCA mutations came from parallel proof-of-concept studies demonstrating the activity of this agent in women with breast and ovarian cancers and BRCA1 or BRCA2 mutations.[30,31] These data ultimately led to US Food and Drug Administration (FDA) approval of olaparib for women with a BRCA mutation and metastatic ovarian cancer after chemotherapy.
Additional data that demonstrate a similar spectrum of activity are available for other PARP inhibitors. Phase I data on the safety and pharmacodynamics of single-agent veliparib have been reported as an abstract,[32] and additional studies of veliparib in combination with mitomycin,[33] irinotecan,[34] and other agents have been reported.[35] VanderWeele et al published a case report of a patient with metastatic CRPC and BRCA2 mutation who had a sustained complete response to veliparib and carboplatin/gemcitabine.[36] It seems likely that many of the available PARP inhibitors may have overlapping activities, and further data will be needed to clarify which agent to use in which tumor type and the relative toxicities of each agent.
Temozolomide and veliparib in metastatic CRPC
Compelling data implicate PARP1 in the mediation of DNA repair responses to alkylating agents,[37] cellular survival in BRCA-deficient cells,[24,38] and androgen receptor–mediated prostate cancer cellular proliferation.[9,39] Furthermore, data suggest that prostate cancers that harbor the TMPRSS2:ERG fusion (present in up to 50% of prostate cancers) may be more sensitive to PARP inhibition.[13] Therefore, Hussain et al carried out a single-arm pilot study to assess the safety and efficacy of veliparib with the alkylator temozolomide (TMZ) in patients with metastatic CRPC following docetaxel therapy.[40] In this study, patients with a PSA level of ≥ 2 ng/mL were treated with veliparib, 40 mg twice daily, on days 1 to 7 and TMZ, 150 to 200 mg/m2, on days 1 to 5 on a 28-day cycle, based on tolerance data from a phase I study (ClinicalTrials.gov identifier: NCT00526617). The primary endpoint was PSA response rate (30% decline). Of the 25 patients who were evaluable for response, 2 had a confirmed response, 13 had stable PSA, and 10 had progression. The most frequent toxicities were thrombocytopenia, anemia, fatigue, neutropenia, nausea, and constipation. The investigators did assess frequency of TMPRSS2:ERG fusion but found it in only one of eight evaluable patients. Although this patient had stable disease, no conclusions could be drawn regarding the contribution of the fusion product to veliparib sensitivity. Overall, while the combination was considered tolerable, it had only modest activity. No preselection was done in the study, and because BRCAness exists in 20% of patients, it is perhaps not surprising that activity was modest. The lower dose of PARP inhibitor and the lack of established benefit for TMZ may also have contributed to less than robust clinical activity for this combination. Given the emerging molecular data, it seems that future studies will be more likely to identify activity if done in preselected patient populations.
TOPARP
The Trial of PARP Inhibition in Prostate Cancer (TOPARP-A) sought to determine whether patients with prostate cancers with molecularly identified defects in DNA repair benefited from full-dose olaparib therapy.[25] In this phase II study, 50 men with CRPC underwent biopsy of metastatic disease and targeted next-generation sequencing, exome and transcriptome analysis, and digital polymerase chain reaction. The primary endpoint was response rate (either objective response or reduction of 50% in PSA level or reduction in circulating tumor cells). All had previously received docetaxel, and most had been treated with abiraterone or enzalutamide (98%) and cabazitaxel (58%). Patients were grouped according to the presence or absence of a homozygous deletion of or deleterious mutation in DNA damage response genes, which predict sensitivity to PARP inhibition. Overall, 16 of 49 evaluable patients (33%) were biomarker positive (indicative of homozygous deleterious changes in BRCA1/2, ATM, Fanconi anemia genes, orCHEK2). Of these, five patients had germline and somatic events (three patients with germline BRCA2 and three patients with germline ATM deletions or mutations). Of the 16 patients with deleterious changes in DNA repair genes, 14 (88%) responded to olaparib. The median overall survival for patients with biomarker-positive DRD tumors who received olaparib was 13.8 months, compared with 7.5 months for those with biomarker-negative tumors (P = .05). Interestingly, two biomarker-negative patients also met criteria for response to olaparib. Although one was a longer-term responder still on therapy at the time of publication, this particular patient did harbor monoallelic deletions of bothBRCA2 and PALB2 that did not meet criteria for the prespecified biomarker-positive category but that may have contributed to tumor sensitivity. Toxicity was as expected, with patients displaying grade 3 or 4 anemia (10/50), fatigue (6/50), leukopenia (3/50), thrombocytopenia (2/50), and neutropenia (2/50). These results illustrate the feasibility of using molecular profiling to identify prostate cancers that display molecular features suggestive of sensitivity to PARP inhibition (BRCAness).
NCI 9012
ETS gene fusions—which result from gene rearrangement and juxtaposition of an androgen-responsive gene, such as TMPRSS2, to an ETS transcription factor gene, such as ERG or ETV1—occur in 50% to 60% of prostate cancers.[41,42] ETS transcription factors may also physically interact with PARP1, and PARP1 activity may be required for ETS-mediated invasion, transcription, and metastasis.[13] Androgen receptor–mediated transcription may also promote DNA DSBs and requires PARP activity for efficient repair.[43-45] Therefore, therapeutic targeting of androgen receptor signaling and PARP1 activity using abiraterone and veliparib is an attractive strategy in the management of metastatic prostate cancer.
A randomized phase II clinical trial in patients with metastatic CRPC was recently completed; it examined whether ETS fusion is a biomarker of response to abiraterone or abiraterone plus veliparib. In this study, 148 patients with metastatic CRPC underwent biopsy followed by assessment of ETS fusion status and then random assignment to either abiraterone alone or abiraterone plus veliparib. The primary endpoint was confirmed PSA response in patients receiving either abiraterone alone or combination therapy, stratified by ETS status. Secondary endpoints included safety, objective response rate, progression-free survival, and whether DNA repair gene deficiency (homozygous deletions of or deleterious mutations in: BRCA1, BRCA2, ATM, FANCA, PALB2, RAD51B, RAD51C) predicts response. This trial has now completed enrollment, and preliminary results will be presented at the American Society of Clinical Oncology 2016 Annual Meeting. Although final results are pending, the study does illustrate the feasibility of a large-scale metastatic tissue–based, biomarker-driven trial involving PARP inhibition in patients with metastatic CRPC. This study will also begin to ascertain the role of ETS fusions in determining response to PARP inhibitor therapy and will further explore the contribution of DRD to patient outcomes in those treated with standard therapy (abiraterone arm) and those treated with PARP inhibition (abiraterone plus veliparib arm).
Future studies
Given the data from the studies discussed previously and the enthusiasm for molecularly targeted trials in oncology, there is interest in further testing of PARP inhibition in prostate cancer patients. Multiple trials have recently been completed, are actively enrolling, or are nearing activation within this space (see Table, ClinicalTrials.gov).
Olaparib. Olaparib is the agent that is farthest along in clinical development and has an FDA indication in ovarian cancer. Olaparib also has the most active or pending studies in prostate cancer patients. TOPARP continues to enroll patients with metastatic CRPC, with a target accrual of 98 patients (ClinicalTrials.gov identifier: NCT01682772). There is a randomized double-blind, placebo-controlled phase II study of abiraterone plus olaparib or placebo for patients with metastatic CRPC who received prior docetaxel therapy (ClinicalTrials.gov identifier: NCT01972217). This trial, which is similar to the NCI 9012 study, has completed enrollment, but results are pending. Another trial is examining the biologic effect of olaparib on prostate cancer specimens when given alone or in combination with degarelix prior to prostatectomy (ClinicalTrials.gov identifier: NCT02324998). Furthermore, there is an open-label phase II study to assess the efficacy and safety of olaparib in patients with BRCA1 or BRCA2 mutations (regardless of tumor type), which is ongoing but no longer enrolling patients (ClinicalTrials.gov identifier: NCT01078662).
Veliparib. NCI 9012 (discussed previously) will help determine whether veliparib has potential therapeutic activity in metastatic CRPC and may identify molecularly determined subsets of disease (ie, ETS fusion–positive, DRD-positive) that might be expected to show the most benefit. The results of this study may help determine whether additional studies of this agent within the prostate cancer space are warranted.
Niraparib. The Hoosier Cancer Research Network has a planned phase I study of the combination of enzalutamide and niraparib for patients with metastatic CRPC (ClinicalTrials.gov identifier: NCT02500901), which has not yet begun enrollment. The primary endpoint of this study will be determination of MTD and dose-limiting toxicity.
Talazoparib. Although no prostate cancer–specific trials using other PARP inhibitors are currently active, several trials for molecularly targeted patient populations or phase I trials for toxicity assessment in combination with chemotherapy are ongoing; these provide some information on prostate cancer populations, depending on the types of solid tumors enrolled. There is a phase I trial of talazoparib in combination with carboplatin and paclitaxel (ClinicalTrials.gov identifier: NCT02317874) and another for patients with solid tumors and hepatic and renal dysfunction (ClinicalTrials.gov identifier: NCT02567396).
Precision Targeting of the PARP Pathway in Prostate Cancer
PARP inhibitors are a promising therapeutic option for men with prostate cancer. There is good evidence that men with either germline or somatic mutations in DNA repair pathways can derive therapeutic benefit from inhibition of PARP1/2, which blocks repair of SSB, driving persistent DSBs that lead to cancer cell lethality. Preclinical data also suggest that PARP inhibition may produce benefits by targeting chromatin and gene transcription, which implies that clinical benefits may extend beyond patients with DRD tumors.[12] To continue to develop PARP inhibitors within the prostate cancer field, we will need to develop and refine a set of biomarkers for use in selecting the right patient populations for these agents and then incorporate these biomarkers into prospective studies. As part of a precision therapy strategy, PARP inhibitors will likely play an important role in the management of prostate cancer in the near future.
It is now feasible to comprehensively profile the mutational, epigenetic, and gene expression changes in men with prostate cancer, and we are beginning to use this information to guide treatment choices.[7] Unfortunately, the functional relevance of many of the molecular features uncovered in these profiles is not completely understood. DNA repair processes are complex and require many genes for efficient repair of various types of DNA damage. Most past and ongoing studies focused on patients with specific molecular features, such as BRCA1, BRCA2, ATM, FANCA, PALB2, RAD51B, and RAD51C mutations. While mutations of these genes are likely to affect sensitivity to PARP inhibitors, mutations in other DNA repair or transcription factor genes may as well, and identification of those genes could expand the patient population that could benefit from therapy. Determination of whether other genes are susceptible to PARP inhibitor therapy will require robust preclinical models with a wide selection of genetic changes that reflect human disease; such models can be used to determine whether additional mutations and epigenetic or gene expression changes also result in PARP inhibitor sensitivity. Given the potential infrequency of many of the individual mutations that might sensitize to PARP inhibitors, large-scale registries that catalog mutations and their responsiveness to therapies may be needed.
As we define the molecular features that suggest sensitivity to PARP inhibition, the challenge will then become understanding the best strategy for incorporating these targeted agents into our standard treatment algorithms. In the context of prostate cancer, PARP inhibitors could be considered in high-risk patient populations in an adjuvant manner, before or with androgen deprivation therapy (ADT) in patients with newly metastatic disease, or in the setting of castration-resistant disease before or after the many other therapeutic options. To date, most trials in the prostate cancer space have been in the castration-resistant setting, perhaps because mutations in DNA damage genes may become more common as the disease progresses.[25] Nonetheless, there is no reason to assume that patients who harbor mutations may not benefit earlier in the disease course. Adjuvant use of PARP inhibitors in those with high-risk or micrometastatic disease could conceivably render patients disease free. Similarly, the combination of ADT and PARP inhibitors in early metastatic disease may provoke prolonged progression-free intervals similar to the situation with early docetaxel therapy but with less toxicity.[4,5] In the context of castration-resistant disease, it is reasonable to hypothesize that the combination of PARP inhibitors with hormonal agents such as abiraterone or enzalutamide or with chemotherapies might act synergistically to promote disease control.
The trials to examine these questions may be more challenging to design and execute because patients with sensitizing molecular changes represent a limited subset of total patients with prostate cancer. This means that in order to identify the subset that will benefit, many will need to be screened.[25] Because most molecular analyses are done using biopsy tissue, screening and cost may be challenging factors. In addition, the natural history of patients with DNA damage pathway mutations may also be distinct from those without such mutations. It is conceivable that mutations in DNA damage response genes may modulate patient response to standard hormonal agents, chemotherapy, or radium because all three of these therapeutic modalities have the potential to induce DNA damage in prostate cancer cells. Given these caveats, it will be essential to design an efficient precision medicine clinical trial pipeline that can rapidly molecularly profile patient tumors, assign to a therapeutic intervention, and then assess the complex resulting data and analyze results according to molecular categories.
PARP inhibitors have the potential to be a promising addition to the therapeutic arsenal used to treat prostate cancer and other solid tumors that harbor the appropriate molecular features. The transition from a standard, one-size-fits-all approach to a targeted, precision medicine strategy in which an individual prostate cancer patient’s tumor biology will guide choice of therapy will require careful planning and thought. The inclusion of PARP-targeted therapies before, after, with, or in place of standard hormonal therapies and chemotherapies will need to be defined so as to maximize antitumor effect and patient survival. Hopefully, application of these novel combinations in those most likely to benefit will ultimately lead to longer and better lives for patients with prostate cancer.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7-30.
2. McNeil C. NCI-MATCH launch highlights new trial design in precision-medicine era. J Natl Cancer Inst. 2015;107.
3. Palmbos PL, Hussain M. Non-castrate metastatic prostate cancer: Have the treatment options changed? Semin Oncol. 2013;40:337-46.
4. Sweeney CJ, Chen YH, Carducci M, et al. Chemohormonal therapy in metastatic hormone-sensitive prostate cancer. N Engl J Med. 2015;373:737-46.
5. James ND, Sydes MR, Clarke NW, et al. Addition of docetaxel, zoledronic acid, or both to first-line long-term hormone therapy in prostate cancer (STAMPEDE): survival results from an adaptive, multiarm, multistage, platform randomised controlled trial. Lancet. 2016;387:1163-77.
6. Ruch JM, Hussain MH. Evolving therapeutic paradigms for advanced prostate cancer. Oncology (Williston Park). 2011;25:496-504, 508.
7. Roychowdhury S, Iyer MK, Robinson DR, et al. Personalized oncology through integrative high-throughput sequencing: a pilot study. Sci Transl Med. 2011;3:111ra21.
8. Gibson BA, Kraus WL. New insights into the molecular and cellular functions of poly(ADP-ribose) and PARPs. Nat Rev Mol Cell Biol. 2012;13:411-24.
9. Schiewer MJ, Knudsen KE. Transcriptional roles of PARP1 in cancer. Mol Cancer Res. 2014;12:1069-80.
10. Polo SE, Jackson SP. Dynamics of DNA damage response proteins at DNA breaks: a focus on protein modifications. Genes Dev. 2011;25:409-33.
11. Plummer R. Poly(ADP-ribose)polymerase (PARP) inhibitors: from bench to bedside. Clin Oncol (R Coll Radiol). 2014;26:250-6.
12. Feng FY, de Bono JS, Rubin MA, Knudsen KE. Chromatin to clinic: the molecular rationale for PARP1 inhibitor function. Mol Cell. 2015;58:925-34.
13. Brenner JC, Ateeq B, Li Y, et al. Mechanistic rationale for inhibition of poly(ADP-ribose) polymerase in ETS gene fusion-positive prostate cancer. Cancer Cell. 2011;19:664-78.
14. Ricks TK, Chiu HJ, Ison G, et al. Successes and challenges of PARP inhibitors in cancer therapy. Front Oncol. 2015;5:222.
15. Murai J, Huang SY, Das BB, et al. Trapping of PARP1 and PARP2 by clinical PARP inhibitors. Cancer Res. 2012;72:5588-99.
16. Murai J, Huang SY, Renaud A, et al. Stereospecific PARP trapping by BMN 673 and comparison with olaparib and rucaparib. Mol Cancer Ther. 2014;13:433-43.
17. Liu X, Shi Y, Maag DX, et al. Iniparib nonselectively modifies cysteine-containing proteins in tumor cells and is not a bona fide PARP inhibitor. Clin Cancer Res. 2012;18:510-23.
18. Patel AG, De Lorenzo SB, Flatten KS, et al. Failure of iniparib to inhibit poly(ADP-ribose) polymerase in vitro. Clin Cancer Res. 2012;18:1655-62.
19. Ribezzo F, Shiloh Y, Schumacher B. Systemic DNA damage responses in aging and diseases. Semin Cancer Biol. 2016 Jan 7. [Epub ahead of print]
20. Sijmons RH, Hofstra RM. Review: clinical aspects of hereditary DNA mismatch repair gene mutations. DNA Repair (Amst). 2016;38:155-62.
21. Lord CJ, Ashworth A. BRCAness revisited. Nat Rev Cancer. 2016;16:110-20.
22. Edwards SM, Kote-Jarai Z, Meitz J, et al. Two percent of men with early-onset prostate cancer harbor germline mutations in the BRCA2 gene. Am J Hum Genet. 2003;72:1-12.
23. Castro E, Eeles R. The role of BRCA1 and BRCA2 in prostate cancer. Asian J Androl. 2012;14:409-14.
24. Fong PC, Boss DS, Yap TA, et al. Inhibition of poly(ADP-ribose) polymerase in tumors from BRCA mutation carriers. N Engl J Med. 2009;361:123-34.
25. Mateo J, Carreira S, Sandhu S, et al. DNA-repair defects and olaparib in metastatic prostate cancer. N Engl J Med. 2015;373:1697-708.
26. Grasso CS, Wu YM, Robinson DR, et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature. 2012;487:239-43.
27. McCabe N, Turner NC, Lord CJ, et al. Deficiency in the repair of DNA damage by homologous recombination and sensitivity to poly(ADP-ribose) polymerase inhibition. Cancer Res. 2006;66:8109-15.
28. Beltran H, Yelensky R, Frampton GM, et al. Targeted next-generation sequencing of advanced prostate cancer identifies potential therapeutic targets and disease heterogeneity. Eur Urol. 2013;63:920-6.
29. Robinson D, Van Allen EM, Wu YM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015;161:1215-28.
30. Audeh MW, Carmichael J, Penson RT, et al. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and recurrent ovarian cancer: a proof-of-concept trial. Lancet. 2010;376:245-51.
31. Tutt A, Robson M, Garber JE, et al. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: a proof-of-concept trial. Lancet. 2010;376:235-44.
32. Puhalla S, Beumer JH, Pahuja S, et al. Final results of a phase 1 study of single-agent veliparib in patients with either BRCA 1/2-mutated cancer, platinum-refractory ovarian, or basal-like breast cancer. J Clin Oncol. 2014;32(suppl 5):abstr 2570.
33. Villalona-Calero MA, Duan W, Zhao W, et al. Veliparib alone or in combination with mitomycin C in patients with solid tumors with functional deficiency in homologous recombination repair. J Natl Cancer Inst. 2016;108.
34. LoRusso PM, Li J, Burger A, et al. Phase I safety, pharmacokinetic and pharmacodynamic study of the poly (ADP-ribose) polymerase inhibitor veliparib with irinotecan in patients with advanced tumors. Clin Cancer Res. 2016 Feb 3. [Epub ahead of print]
35. Wagner LM. Profile of veliparib and its potential in the treatment of solid tumors. Onco Targets Ther. 2015;8:1931-9.
36. VanderWeele DJ, Paner GP, Fleming GF, Szmulewitz RZ. Sustained complete response to cytotoxic therapy and the PARP inhibitor veliparib in metastatic castration-resistant prostate cancer—a case report. Front Oncol. 2015;5:169.
37. Plummer R, Jones C, Middleton M, et al. Phase I study of the poly(ADP-ribose) polymerase inhibitor, AG014699, in combination with temozolomide in patients with advanced solid tumors. Clin Cancer Res. 2008;14:7917-23.
38. Fong PC, Yap TA, Boss DS, et al. Poly(ADP)-ribose polymerase inhibition: frequent durable responses in BRCA carrier ovarian cancer correlating with platinum-free interval. J Clin Oncol. 2010;28:2512-9.
39. Schiewer MJ, Goodwin JF, Han S, et al. Dual roles of PARP-1 promote cancer growth and progression. Cancer Discov. 2012;2:1134-49.
40. Hussain M, Carducci MA, Slovin S, et al. Targeting DNA repair with combination veliparib (ABT-888) and temozolomide in patients with metastatic castration-resistant prostate cancer. Invest New Drugs. 2014;32:904-12.
41. Kumar-Sinha C, Tomlins SA, Chinnaiyan AM. Recurrent gene fusions in prostate cancer. Nat Rev Cancer. 2008;8:497-511.
42. Tomlins SA, Rhodes DR, Perner S, et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science. 2005;310:644-8.
43. Haffner MC, De Marzo AM, Meeker AK, et al. Transcription-induced DNA double strand breaks: both oncogenic force and potential therapeutic target? Clin Cancer Res. 2011;17:3858-64.
44. Haffner MC, Aryee MJ, Toubaji A, et al. Androgen-induced TOP2B-mediated double-strand breaks and prostate cancer gene rearrangements. Nat Genet. 2010;42:668-75.
45. Lin C, Yang L, Tanasa B, et al. Nuclear receptor-induced chromosomal proximity and DNA breaks underlie specific translocations in cancer. Cell. 2009;139:1069-83.
RELATED ARTICLES
Adding ADT Fails to Improve Survival in Favorable Intermediate-Risk Prostate Cancer
Active Surveillance Follow-Up for Prostate Cancer Lacks Consistency
PARP Inhibition in Prostate Cancer: A Promising Approach
Decrease in Prostate Cancer Mortality Mirrors Declines in Cigarette Smoking
Maintaining High Physical Activity Improves Prostate Cancer Survival
Novel Discoveries in Molecular Biology and Biomedical Science, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 1: Next Generation Sequencing (NGS)
Novel Discoveries in Molecular Biology and Biomedical Science
Curator: Larry H. Bernstein, MD, FCAP
UPDATED on 6/1/2016
The following is a collection of current articles on noncoding DNA, synthetic genome engineering, protein regulation of apoptosis, drug design, and geometrics.
No longer ‘junk DNA’ — shedding light on the ‘dark matter’ of the genome
A new tool called “LIGR-Seq” enables scientists to explore in depth what non-coding RNAs actually do in human cells May 23, 2016
he LIGR-seq method for global-scale mapping of RNA-RNA interactions in vivo to reveal unexpected functions for uncharacterized RNAs that act via base-pairing interactions (credit: University of Toronto)
What used to be dismissed by many as “junk DNA” has now become vitally important, as accelerating genomic data points to the importance of non-coding RNAs (ncRNAs) — a genome’s messages that do not specifically code for proteins — in development and disease.
But our progress in understanding these molecules has been slow because of the lack of technologies that allow for systematic mapping of their functions.
Now, professor Benjamin Blencowe’s team at the University of Toronto’s Donnelly Centre has developed a method called “LIGR-seq” that enables scientists to explore in depth what ncRNAs do in human cells.
The study, described in Molecular Cell, was published on May 19, along with two other papers, in Molecular Cell and Cell, respectively, from Yue Wan’s group at the Genome Institute of Singapore and Howard Chang’s group at Stanford University in California, who developed similar methods to study RNAs in different organisms.
Of the 3 billion letters in the human genome, only two per cent make up the protein-coding genes. The genes are copied, or transcribed, into messenger RNA (mRNA) molecules, which provide templates for building proteins that do most of the work in the cell. Much of the remaining 98 per cent of the genome was initially considered by some as lacking in functional importance. However, large swaths of the non-coding genome — between half and three quarters of it — are also copied into RNA.
So then what might the resulting ncRNAs do? That depends on whom you ask. Some researchers believe that most ncRNAs have no function, that they are just a by-product of the genome’s powerful transcription machinery that makes mRNA. However, it is emerging that many ncRNAs do have important roles in gene regulation — some ncRNAs act as carriages for shuttling the mRNAs around the cell, or provide a scaffold for other proteins and RNAs to attach to and do their jobs.
But the majority of available data has trickled in piecemeal or through serendipitous discovery. And with emerging evidence that ncRNAs could drive disease progression, such as cancer metastasis, there was a great need for a technology that would allow a systematic functional analysis of ncRNAs.
“Up until now, with existing methods, you had to know what you are looking for because they all require you to have some information about the RNA of interest. The power of our method is that you don’t need to preselect your candidates; you can see what’s occurring globally in cells, and use that information to look at interesting things we have not seen before and how they are affecting biology,” says Eesha Sharma, a PhD candidate in Blencowe’s group who, along with postdoctoral fellow Tim Sterne-Weiler, co-developed the method.
The human RNA-RNA interactome, showing interactions detected by LIGR-seq (credit: University of Toronto)
The new ‘‘LIGation of interacting RNA and high-throughput sequencing’’ (LIGR-seq) tool captures interactions between different RNA molecules. When two RNA molecules have matching sequences — strings of letters copied from the DNA blueprint — they will stick together like Velcro. With LIGR-seq, the paired RNA structures are removed from cells and analyzed by state-of-the-art sequencing methods to precisely identify the RNAs that are stuck together.
“Most researchers in the life sciences agree that there’s an urgent need to understand what ncRNAs do. This technology will open the door to developing a new understanding of ncRNA function,” says Blencowe, who is also a professor in the Department of Molecular Genetics.
Not having to rely on pre-existing knowledge will boost the discovery of RNA pairs that have never been seen before. Scientists can also, for the first time, look at RNA interactions as they occur in living cells, in all their complexity, unlike in the juices of mashed up cells that they had to rely on before. This is a bit like moving on to explore marine biology from collecting shells on the beach to scuba-diving among the coral reefs, where the scope for discovery is so much bigger.
Actually, ncRNAs come in multiple flavors: there’s rRNA, tRNA, snRNA, snoRNA, piRNA, miRNA, and lncRNA, to name a few, where prefixes reflect the RNA’s place in the cell or some aspect of its function. But the truth is that no one really knows the extent to which these ncRNAs control what goes on in the cell, or how they do this.
Discoveries
Nonetheless, the new technology developed by Blencowe’s group has been able to pick up new interactions involving all classes of RNAs and has already revealed some unexpected findings.
The team discovered new roles for small nucleolar RNAs (snoRNAs), which normally guide chemical modifications of other ncRNAs. It turns out that some snoRNAs can also regulate stability of a set of protein-coding mRNAs. In this way, snoRNAs can also directly influence which proteins are made, as well as their abundance, adding a new level of control in cell biology.
And this is only the tip of the iceberg; the researchers plan to further develop and apply their technology to investigate the ncRNAs in different settings.
“We would like to understand how ncRNAs function during development. We are particularly interested in their role in the formation of neurons. But we will also use our method to discover and map changes in RNA-RNA interactions in the context of human diseases,” says Blencowe.
Abstract of Global Mapping of Human RNA-RNA Interactions
The majority of the human genome is transcribed into non-coding (nc)RNAs that lack known biological functions or else are only partially characterized. Numerous characterized ncRNAs function via base pairing with target RNA sequences to direct their biological activities, which include critical roles in RNA processing, modification, turnover, and translation. To define roles for ncRNAs, we have developed a method enabling the global-scale mapping of RNA-RNA duplexes crosslinked in vivo, “LIGation of interacting RNA followed by high-throughput sequencing” (LIGR-seq). Applying this method in human cells reveals a remarkable landscape of RNA-RNA interactions involving all major classes of ncRNA and mRNA. LIGR-seq data reveal unexpected interactions between small nucleolar (sno)RNAs and mRNAs, including those involving the orphan C/D box snoRNA, SNORD83B, that control steady-state levels of its target mRNAs. LIGR-seq thus represents a powerful approach for illuminating the functions of the myriad of uncharacterized RNAs that act via base-pairing interactions.
Understanding the unknown functions of these genes may lead to the creation of new diagnostic tests for clinical laboratories and anatomic pathology groups
Once again, J. Craig Venter, PhD, is charting new ground in gene sequencing andgenomic science. This time his research team has built upon the first synthetic cell they created in 2010 to build a more sophisticated synthetic cell. Their findings from this work may give pathologists and medical laboratory scientists new tools to diagnose disease.
Recently the research team at the J. Craig Venter Institute (JCVI) and Synthetic Genomics, Inc. (SGI) published their latest findings. Among the things they learned is that science still does not understand the functions of about a third of the genes required for their synthetic cells to function.
JCVI-syn3.0 Could Radically Alter Understanding of Human Genome
Based in La Jolla, Calif., and Rockville, Md., JCVI is a not-for-profit research institute aiming to advance genomics. Building upon its first synthetic cell—Mycoplasma mycoides (M. mycoides) JCVI-syn1.0, which JCVI constructed in 2010—the same team of scientists created the first minimal synthetic bacterial cell, which they calledJCVI-syn3.0. This new artificial cell contains 531,560 base pairs and just 473 genes, which means it is the smallest genome of any organism that can be grown in laboratory media, according to a JCVI-SGI statement.
For pathologists and medical laboratory leaders, the creation of a synthetic life form is a milestone toward better understanding genome sequencing and how this new knowledge may help advance both diagnostics and therapeutics.
“What we’ve done is important because it is a step toward completely understanding how a living cell works,” Clyde Hutchison III, PhD, told New Scientist. “If we can really understand how the cell works, then we will be able to design cells efficiently for the production of pharmaceutical and other useful products.” Hutchison is Professor Emeritus of Microbiology and Immunology at the University of North Carolina at Chapel Hill, Distinguished Professor at the J. Craig Venter Institute, a member of the National Academy of Sciences, and a fellow of the American Academy of Arts and Sciences.
Clyde Hutchison, III, PhD (above), Professor Emeritus of Microbiology and Immunology at the University of North Carolina at Chapel Hill and Distinguished Professor at the J. Craig Venter Institute, stated that his team’s “goal is to have a cell for which the precise biological function of every gene is known.” (Photo credit: JCVI.)
Understanding a Gene’s True Purpose
According to the JCVI researchers, 149 genes have no known purpose. They are, however, necessary for life and health.
“We know about two-thirds of the essential biology, and we’re missing a third,” stated J. Craig Venter, PhD, Founder and CEO of JCVI, in a story published by MedPage Today.
This knowledge is based upon decades of research. JCVI seeks to create a minimal cell operating system to understand biology, while also providing what the JCVI statement called a “chassis for use in industrial applications.”
What Do these Genes Do Anyway?
The JCVI team found that among most genes’ biological functions:
“JCVI-syn3.0 is a working approximation of a minimal cellular genome—a compromise between a small genome size and a workable growth rate for an experimental organism. It retains almost all the genes that are involved in the synthesis and processing of macromolecules. Unexpectedly, it also contains 149 genes with unknown biological functions, suggesting the presence of undiscovered functions that are essential for life,” the researchers told the journal Science.
More research is needed, the scientists say, into the 149 genes that appear to lack specific biologic functions.
Unlocking Mystery of the 149 Genes Could Lead to Advances in Genomic Science
“Finding so many genes without a known function is unsettling, but it’s exciting because it’s left us with much still to learn. It’s like the ‘dark matter’ of biology,” said Alistair Elfick, PhD, Chair of Synthetic Biological Engineering, University of Edinburgh, UK, in the New Scientist article.
Studies such as JCVI’s research is key to broadening understanding and framing appropriate questions about scientific, ethical, and economic implications of synthetic biology.
The creation of a synthetic cell will have a profound and positive impact on understanding of biology and how life works, JCVI said.
Such research may inspire new whole genome synthesis tools and semi-automated processes that could dramatically affect clinical laboratory procedures. It also could lead to new techniques and tools for advanced vaccine and pharmaceuticals, JCVI pointed out.
No single technique has set the molecular biology field ablaze with excitement and potential like the CRISPR-Cas9 genome editing system has following its introduction only a few short years ago. The following articles represent the flexibility of this technique to potentially treat a host of genetic disorders and possibly even prevent the onset of disease.
Scientists recently convened at the CRISPR Precision Gene Editing Congress, held in Boston, to discuss the new technology. As with any new technique, scientists have discovered that CRISPR comes with its own set of challenges, and the Congress focused its discussion around improving specificity, efficiency, and delivery.
With a staggering number of papers published in the past several years involving the characterization and use of the CRISPR/Cas9 gene editing system, it is surprising that researchers are still finding new features of the versatile molecular scissor enzyme.
If a Cas9 nuclease variant could be engineered that was less grabby, it might loosen its grip on DNA sequences throughout the genome—except those sequences representing on-target sites. That’s the assumption that guided a new investigation by researchers at Massachusetts General Hospital.
The gene-editing technology known as CRISPR-Cas9 is starting to raise expectations in the therapeutic realm. In fact, CRISPR-Cas9 and other CRISPR systems are moving so close to therapeutic uses that the technology’s ethical implications are starting to attract notice.
Published: Tuesday, May 24, 2016 A comparison of synthetic gene-activating Cas9 proteins can help guide research and development of therapeutic approaches.
The CRISPR-Cas9 system has come to be known as the quintessential tool that allows researchers to edit the DNA sequences of many organisms and cell types. However, scientists are also increasingly recognizing that it can be used to activate the expression of genes. To that end, they have built a number of synthetic gene activating Cas9 proteins to study gene functions or to compensate for insufficient gene expression in potential therapeutic approaches.
“The possibility to selectively activate genes using various engineered variants of the CRISPR-Cas9 system left many researchers questioning which of the available synthetic activating Cas9 proteins to use for their purposes. The main challenge was that all had been uniquely designed and tested in different settings; there was no side-by-side comparison of their relative potentials,” said George Church, Ph.D., who is Core Faculty Member at the Wyss Institute for Biologically Inspired Engineering at Harvard University, leader of its Synthetic Biology Platform, and Professor of Genetics at Harvard Medical School. “We wanted to provide that side-by-side comparison to the biomedical research community.”
In a study published on 23 May in Nature Methods, the Wyss Institute team reports how it rigorously compared and ranked the most commonly used artificial Cas9 activators in different cell types from organisms including humans, mice and flies. The findings provide a valuable guide to researchers, allowing them to streamline their endeavors.
The team also included Wyss Core Faculty Member James Collins, Ph.D., who also is the Termeer Professor of Medical Engineering & Science and Professor of Biological Engineering at the Massachusetts Institute of Technology (MIT)’s Department of Biological Engineering and Norbert Perrimon, Ph.D., a Professor of Genetics at Harvard Medical School.
Gene activating Cas9 proteins are fused to variable domains borrowed from proteins with well-known gene activation potentials and engineered so that the DNA editing ability is destroyed. In some cases, the second component of the CRISPR-Cas9 system, the guide RNA that targets the complex to specific DNA sequences, also has been engineered to bind gene-activating factors.
“We first surveyed seven advanced Cas9 activators, comparing them to each other and the original Cas9 activator that served to provide proof-of-concept for the gene activation potential of CRISPR-Cas9. Three of them, provided much higher gene activation than the other candidates while maintaining high specificities toward their target genes,” said Marcelle Tuttle, Research Fellow at the Wyss and a co-lead author of the study.
The team went on to show that the three top candidates were comparable in driving the highest level of gene expression in cells from humans, mice and fruit flies, irrespective of their tissue and developmental origins. The researchers also pinpointed ways to further maximize gene activation employing the three leading candidates.
“In some cases, maximum possible activation of a target gene is necessary to achieve a cellular or therapeutic effect. We managed to cooperatively enhance expression of specific genes when we targeted them with three copies of a top performing activator using three different guide RNAs,” said Alejandro Chavez, Ph.D., a Postdoctoral Fellow and the study’s co-first author.
“The ease of use of CRISPR-Cas9 offers enormous potential for development of genome therapeutics. This study provides valuable new design criteria that will help enable synthetic biologists and bioengineers to develop more effective targeted genome engineering technologies in the future,” said Wyss Institute Founding Director Donald Ingber, M.D., Ph.D., who is the Judah Folkman Professor of Vascular Biology at Harvard Medical School and the Vascular Biology Program at Boston Children’s Hospital, and also Professor of Bioengineering at the Harvard John A. Paulson School of Engineering and Applied Sciences.
Engineering T Cells to Functionally Cure HIV-1 Infection
Despite the ability of antiretroviral therapy to minimize human immunodeficiency virus type 1 (HIV-1) replication and increase the duration and quality of patients’ lives, the health consequences and financial burden associated with the lifelong treatment regimen render a permanent cure highly attractive. Although T cells play an important role in controlling virus replication, they are themselves targets of HIV-mediated destruction. Direct genetic manipulation of T cells for adoptive cellular therapies could facilitate a functional cure by generating HIV-1–resistant cells, redirecting HIV-1–specific immune responses, or a combination of the two strategies. In contrast to a vaccine approach, which relies on the production and priming of HIV-1–specific lymphocytes within a patient’s own body, adoptive T-cell therapy provides an opportunity to customize the therapeutic T cells prior to administration. However, at present, it is unclear how to best engineer T cells so that sustained control over HIV-1 replication can be achieved in the absence of antiretrovirals. This review focuses on T-cell gene-engineering and gene-editing strategies that have been performed in efforts to inhibit HIV-1 replication and highlights the requirements for a successful gene therapy–mediated functional cure.
Automated top-down design technique simplifies creation of DNA origami nanostructures
Nanoparticles for drug delivery and cell targeting, nanoscale robots, custom-tailored optical devices, and DNA as a storage medium are among the possible applications
May 27, 2016
The boldfaced line, known as a spanning tree, follows the desired geometric shape of the target DNA origami design method, touching each vertex just once. A spanning tree algorithm is used to map out the proper routing path for the DNA strand. (credit: Public Domain)
MIT, Baylor College of Medicine, and Arizona State University Biodesign Institute researchers have developed a radical new top-down DNA origami* design method based on a computer algorithm that allows for creating designs for DNA nanostructures by simply inputting a target shape.
DNA origami (using DNA to design and build geometric structures) has already proven wildly successful in creating myriad forms in 2- and 3- dimensions, which conveniently self-assemble when the designed DNA sequences are mixed together. The tricky part is preparing the proper DNA sequence and routing design for scaffolding and staple strands to achieve the desired target structure. Typically, this is painstaking work that must be carried out manually.
The new algorithm, which is reported together with a novel synthesis approach in the journal Science, promises to eliminate all that and expands the range of possible applications of DNA origami in biomolecular science and nanotechnology. Think nanoparticles for drug delivery and cell targeting, nanoscale robots in medicine and industry, custom-tailored optical devices, and most interesting: DNA as a storage medium, offering retention times in the millions of years.**
Shape-shifting, top-down software
Unlike traditional DNA origami, in which the structure is built up manually by hand, the team’s radical top-down autonomous design method begins with an outline of the desired form and works backward in stages to define the required DNA sequence that will properly fold to form the finished product.
“The Science paper turns the problem around from one in which an expert designs the DNA needed to synthesize the object, to one in which the object itself is the starting point, with the DNA sequences that are needed automatically defined by the algorithm,” said Mark Bathe, an associate professor of biological engineering at MIT, who led the research. “Our hope is that this automation significantly broadens participation of others in the use of this powerful molecular design paradigm.”
The algorithm, which is known as DAEDALUS (DNA Origami Sequence Design Algorithm for User-defined Structures) after the Greek craftsman and artist who designed labyrinths that resemble origami’s complex scaffold structures, can build any type of 3-D shape, provided it has a closed surface. This can include shapes with one or more holes, such as a torus.
A simplified version of the top-down procedure used to design scaffolded DNA origami nanostructures. It starts with a polygon corresponding to the target shape. Software translates a wireframe version of this structure into a plan for routing DNA scaffold and staple strands. That enables a 3D DNA-based atomic-level structural model that is then validated using 3D cryo-EM reconstruction. (credit: adapted from Biodesign Institute images)
With the new technique, the target geometric structure is first described in terms of a wire mesh made up of polyhedra, with a network of nodes and edges. A DNA scaffold using strands of custom length and sequence is generated, using a “spanning tree” algorithm — basically a map that will automatically guide the routing of the DNA scaffold strand through the entire origami structure, touching each vertex in the geometric form once. Complementary staple strands are then assigned and the final DNA structural model or nanoparticle self-assembles, and is then validated using 3D cryo-EM reconstruction.
The software allows for fabricating a variety of geometric DNA objects, including 35 polyhedral forms (Platonic, Archimedean, Johnson and Catalan solids), six asymmetric structures, and four polyhedra with nonspherical topology, using inverse design principles — no manual base-pair designs needed.
To test the method, simpler forms known as Platonic solids were first fabricated, followed by increasingly complex structures. These included objects with nonspherical topologies and unusual internal details, which had never been experimentally realized before. Further experiments confirmed that the DNA structures produced were potentially suitable for biological applications since they displayed long-term stability in serum and low-salt conditions.
Biological research uses
The research also paves the way for designing nanoscale systems mimicking the properties of viruses, photosynthetic organisms, and other sophisticated products of natural evolution. One such application is a scaffold for viral peptides and proteins for use as vaccines. The surface of the nanoparticles could be designed with any combination of peptides and proteins, located at any desired location on the structure, in order to mimic the way in which a virus appears to the body’s immune system.
The researchers demonstrated that the DNA nanoparticles are stable for more than six hours in serum, and are now attempting to increase their stability further.
The nanoparticles could also be used to encapsulate the CRISPR-Cas9 gene editing tool. The CRISPR-Cas9 tool has enormous potential in therapeutics, thanks to its ability to edit targeted genes. However, there is a significant need to develop techniques to package the tool and deliver it to specific cells within the body, Bathe says.
This is currently done using viruses, but these are limited in the size of package they can carry, restricting their use. The DNA nanoparticles, in contrast, are capable of carrying much larger gene packages and can easily be equipped with molecules that help target the right cells or tissue.
The most exciting aspect of the work, however, is that it should significantly broaden participation in the application of this technology, Bathe says, much like 3-D printing has done for complex 3-D geometric models at the macroscopic scale.
* DNA origami brings the ancient Japanese method of paper folding down to the molecular scale. The basics are simple: Take a length of single-stranded DNA and guide it into a desired shape, fastening the structure together using shorter “staple strands,” which bind in strategic places along the longer length of DNA. The method relies on the fact that DNA’s four nucleotide letters—A, T, C, & G stick together in a consistent manner — As always pairing with Ts and Cs with Gs.
The DNA molecule in its characteristic double stranded form is fairly stiff, compared with single-stranded DNA, which is flexible. For this reason, single stranded DNA makes for an ideal lace-like scaffold material. Further, its pairing properties are predictable and consistent (unlike RNA).
** A single gram of DNA can store about 700 terabytes of information — an amount equivalent to 14,000 50-gigabyte Blu-ray disks — and could potentially be operated with a fraction of the energy required for other information storage options.
Essential role of miRNAs in orchestrating the biology of the tumor microenvironment
MicroRNAs (miRNAs) are emerging as central players in shaping the biology of the Tumor Microenvironment (TME). They do so both by modulating their expression levels within the different cells of the TME and by being shuttled among different cell populations within exosomes and other extracellular vesicles. This review focuses on the state-of-the-art knowledge of the role of miRNAs in the complexity of the TME and highlights limitations and challenges in the field. A better understanding of the mechanisms of action of these fascinating micro molecules will lead to the development of new therapeutic weapons and most importantly, to an improvement in the clinical outcome of cancer patients. Keywords: Exosomes, microRNAs, Tumor microenvironment, Cancer
While cancer treatment and survival have improved worldwide, the need for further understanding of the underlying tumor biology remains. In recent years, there has been a significant shift in scientific focus towards the role of the tumor microenvironment (TME) on the development, growth, and metastatic spread of malignancies. The TME is defined as the surrounding cellular environment enmeshed around the tumor cells including endothelial cells, lymphocytes, macrophages, NK cells, other cells of the immune system, fibroblasts, mesenchymal stem cells (MSCs), and the extracellular matrix (ECM). Each of these components interacts with and influences the tumor cells, continually shifting the balance between pro- and anti-tumor phenotype. One of the predominant methods of communication between these cells is through extracellular vesicles and their microRNA (miRNA) cargo. Extracellular vesicles (EVs) are between 30 nm to a few microns in diameter, are surrounded by a phospholipid bilayer membrane, and are released from a variety of cell types into the local environment. There are three well characterized groups of EVs: 1) exosomes, typically 30–100 nm, 2) microvesicles (or ectosomes), typically 100–1000 nm, and 3) large oncosomes, typically 1–10 μm. Each of these categories has a distinctly unique biogenesis and purpose in cellcell communication despite the fact that current laboratory methods do not always allow precise differentiation. EVs are found to be enriched with membrane-bound proteins, lipid raft-associated and cytosolic proteins, lipids, DNA, mRNAs, and miRNAs, all of which can be transferred to the recipient cell upon fusion to allow cell-cell communications [1]. Of these, miRNAs have been of particular interest in cancer research, both as modifiers of transcription and translation as well as direct inhibitors or enhancers of key regulatory proteins. These miRNAs are a large family of small non-coding RNAs (19–24 nucleotides) and are known to be aberrantly expressed, both in terms of content as well as number, in both the tumor cells and the cells of the TME. Synthesis of these mature miRNA is a complex process, starting with the transcription of long, capped, and polyadenylated pri-miRNA by RNA polymerase II. These are cropped into a 60–100 nucleotide hairpinstructure pre-miRNA by the microprocessor, a heterodimer of Drosha (a ribonuclease III enzyme) and DGCR8 (DiGeorge syndrome critical region gene 8). The premiRNA is then exported to the cytoplasm by exportin 5, cleaved by Dicer, and separated into single strands by helicases. The now mature miRNA are incorporated into the RNA-induced silencing complex (RISC), a cytoplasmic effector machine of the miRNA pathway. The primary mechanism of action of the mature miRNA-RISC complex is through their binding to the 3’ untranslated region, or less commonly the 5’ untranslated region, of target mRNA, leading to protein downregulation either via translational repression or mRNA degradation. More recently, it has been shown that miRNAs can also upregulate the expression of target genes [2]. MiRNA genes are mostly intergenic and are transcribed by independent promoters [3] but can also be encoded by introns, sharing the same promoter of their host gene [4]. MiRNAs undergo the same regulatory mechanisms of any other protein coding gene (promoter methylation, histone modifications, etc.…) [5, 6]. Interestingly, each miRNA may have contradictory effects both within varying tumor cell lines and within different cells of the TME. In this review, we provide a state-of-the-art description of the key role that miRNAs have in the communication between tumor cells and the TME and their subsequent effects on the malignant phenotype. Finally, this review has made every effort to clarify, whenever possible, whether the reference is to the −3p or the -5p miRNA. Whenever such clarification has not been provided, this indicates that it was not possible to infer such information from the cited bibliography.
Angiogenesis and miRNAs Cellular plasticity, critical in the development of malignancy, includes the many diverse mechanisms elicited by cancer cells to increase their malignant potential and develop increasing treatment resistance. One such mechanism, angiogenesis, is critical to the development of metastatic disease, affecting both the growth of malignant cells locally and their survival at distant sites. In the last ten years, miRNAs, often packaged in tumor cell-derived exosomes, have emerged as important contributors to the complicated regulation and balance of pro- and anti-angiogenic factors.
Most commonly, miRNAs derived from cancer cells have oncogenic activity, promoting angiogenesis and tumor growth and survival. The most-well characterized of the pro-angiogenic miRNAs, the miR-17-92 cluster encoding six miRNAs (miR-17, −18a, −19a, −19b, −20a, and −92a), is found on chromosome 13, and is highly conserved among vertebrates [7]. The complex and multifaceted functions of the miR-17-92 cluster are summarized in Fig. 1. Amplification, both at the genetic and RNA level, of miR-17-92 was initially found in several lymphoma cell lines and has subsequently been observed in multiple mouse tumor models [7].
Central role of the miR-17-92 cluster in the biology of the TME. The miR-17-92 cluster encoding miR-17, −18a, −19b, −20a, and -92a is upregulated in multiple tumor types and interacts with various components of the TME to finely “tune” the TME through a complex combination of pro- and anti-tumoral effects
Most commonly, miRNAs derived from cancer cells have oncogenic activity, promoting angiogenesis and tumor growth and survival. The most-well characterized of the pro-angiogenic miRNAs, the miR-17-92 cluster encoding six miRNAs (miR-17, −18a, −19a, −19b, −20a, and −92a), is found on chromosome 13, and is highly conserved among vertebrates [7]. The complex and multifaceted functions of the miR-17-92 cluster are summarized in Fig. 1. Amplification, both at the genetic and RNA level, of miR-17-92 was initially found in several lymphoma cell lines and has subsequently been observed in multiple mouse tumor models [7]. Up-regulation of this particular locus has further been confirmed in miRnome analysis across multiple different tumor types, including lung, breast, stomach, prostate, colon, and pancreatic cancer [8]. The miR-17-92 cluster is directly activated by Myc and modulates a variety of downstream transcription factors important in cell cycle regulation and apoptosis including activation of E2F family and Cyclin-dependent kinase inhibitor (CDKN1A) and downregulation of BCL2L11/BIM and p21 [7]. In addition to promoting cell cycle progression and inhibiting apoptosis, the miR-17-92 cluster also downregulates thrombospondin-1 (Tsp1) and connective tissue growth factor (CTGF), important antiangiogenic proteins [7]. Similarly, microvesicles from colorectal cancer cells contain miR-1246 and TGF-β which are transferred to endothelial cells to silence promyelocytic leukemia protein (PML) and activate Smad 1/5/8 signaling promoting proliferation and migration [9]. Likewise, lung cancer cell line derived microvesicles contain miR-494, in response to hypoxia, which targets PTEN in the endothelial cells promoting angiogenesis through the Akt/eNOS pathway [10]. Lastly, exosomal miR-135b from multiple myeloma cells suppresses the HIF-1/FIH-1 pathway in endothelial cells, increasing angiogenesis [11]. A summary of the studies showing the functions of exosomal miRNAs in shaping the biology of the TME is provided in Table 1.
Table 1
Actions of exosomal miRNAs exchanged between cells of the TME
The most common target of anti-angiogenic therapy is VEGF, and not unsurprisingly, multiple miRNAs (including miR-9, miR-20b, miR-130, miR-150, and miR-497) promote angiogenesis through the induction of the VEGF pathway. The most studied of these is the up-regulation of miR-9 which has been linked to a poor prognosis in multiple tumor types, including breast cancer, non-small cell lung cancer, and melanoma [12]. The two oncogenes MYC and MYCN activate miR-9 and cause E-cadherin downregulation resulting in the upregulated transcription of VEGF [13]. In addition, miR-9 has been shown to upregulate the JAK-STAT pathway, supporting endothelial cell migration and tumor angiogenesis [13]. Both amplification of miR-20b and miR-130 as well as miR-497 suppression regulate VEGF through hypoxia inducible factor 1α (HIF-1α) supporting increased angiogenesis [14, 15, 16, 17]. …..
The pivotal discovery in 2012 by Mitra et al. laid the ground-work for our current knowledge on the interactions between tumor-derived miRNAs and fibroblasts. In combination, the down-regulation of miR-214 and miR-31 and the up-regulation of miR-155 trigger the reprogramming of quiescent fibroblasts to CAFs [32]. As expected, the reverse regulation of these miRNAs reduced the migration and invasion of co-cultured ovarian cancer cells [32]. While the pathway of miR-155’s involvement in CAF biology is still being elucidated, the pathways of miR-214 and miR-31 have been established. In endometrial cancer, miR-31 was found to target the homeobox gene SATB2, leading to enhanced tumor cell migration and invasion [33]. MiR-214 similarly has an inverse correlation with its chemokine target, C-C motif Ligand 5 (CCL5) [32]. CCL5 secretion has been associated with enhanced motility, invasion, and metastatic potential through NF-κB-mediated MMP9 activation and through generation and differentiation of myeloid-derived suppressor cells (MDSCs) [34, 35, 36]. Furthermore, miR-210 and miR-133b overexpression and miR-149 suppression have been subsequently found to independently trigger the conversion to CAFs, possibly through paracrine stimulation, and to additionally promote EMT in prostate and gastric cancer, respectively [37, 38,39]. MiR-210 additionally enlists monocytes and encourages angiogenesis [37]. …
Another function of CAFs is the destruction of the ECM and its remodeling with a tumor-supportive composition and structure which includes modulation of specific integrins and metalloproteinases as some of the most studied miRNA targets. The 23 matrix metalloproteinases (MMPs) are critical in the ECM degradation, disruption of the growth signal balance, resistance to apoptosis, establishment of a favorable metastatic niche, and promotion of angiogenesis [54]. As expected, miRNAs have been found to regulate the actions of MMPs, together working to promote cancer cell growth, invasiveness, and metastasis. In HCC, MMP2 and 9 expression is up-regulated by miR-21 via PTEN pathway downregulation. Similarly, in cholangiocarcinoma it was observed that reduced levels of miR-138 induced up-regulation of RhoC, leading to increased levels of the same two MMPs [55, 56]. ….
As has been shown throughout this review, miRNAs have an important and varied effect on human carcinogenesis by shaping the biology of the TME towards a more permissive pro-tumoral phenotype. The complex events leading to such an outcome are currently quite universally defined as the “educational” process of cancer cells on the surrounding TME. While the initial focus was on the direction from the cancer cell to the surrounding TME, increasingly interest is centered on the implications of a more dynamic bidirectional exchange of genetic information. MiRNAs represent only part of the cargo of the extracellular vesicles, but an increasing scientific literature points towards their pivotal role in creating the micro-environmental conditions for cancer cell growth and dissemination. The nearby future will have to address several questions still unanswered. First, it is absolutely necessary to clarify which miRNAs and to what extent they are involved in this process. The contradictory results of some studies can be explained by the differences in tumor-types and by different concentrations of miRNAs used for functional studies. Understanding whether different concentrations of the same miRNA elicit different target effects and therefore changes the biology of the TME, will represent a significant consideration in the development of this field. It is certainly very attractive (especially in an attempt to develop new and desperately needed better cancer biomarkers) to think that concentrations of miRNAs within the TME are reflected systemically in the circulating levels of that same miRNA, however this has not yet been irrefutably demonstrated. Moreover, the study of the paracrine interactions among different cell populations of the TME and their reciprocal effects has been limited to two, maximum three cell populations. This is still way too far from describing the complexity of the TME and only the development of new tridimensional models of the TME will be able to cast a more conclusive light on such complexity. Finally, the pharmacokinetics of miRNA-containing vesicles is in its infancy at best, and needs to be further developed if the goal is development of new therapies based on the use of exosomic miRNAs. Therefore, the future of miRNA research, particularly in its role in the TME, holds still a lot of questions that need answering. However, for these exact same reasons, this is an incredibly exciting time for research in this field. We can envision a not too far future in which these concerns will be satisfactorily addressed and our understanding of the role of miRNAs within the TME will allow us to use them as new therapeutic weapons to successfully improve the clinical outcome of cancer patients.
Researchers at the Walter and Eliza Hall Institute in Australia have discovered a new way to trigger cell death that could lead to drugs to treat cancer and autoimmune disease.
Programmed cell death (a.k.a. apoptosis) is a natural process that removes unwanted cells from the body. Failure of apoptosis can allow cancer cells to grow unchecked or immune cells to inappropriately attack the body.
The protein known as Bak is central to apoptosis. In healthy cells, Bak sits in an inert state but when a cell receives a signal to die, Bak transforms into a killer protein that destroys the cell.
Triggering the cancer-apoptosis trigger
Institute researchers Sweta Iyer, PhD, Ruth Kluck, PhD, and colleagues unexpectedly discovered that an antibody they had produced to study Bak actually bound to the Bak protein and triggered its activation. They hope to use this discovery to develop drugs that promote cell death.
The researchers used information about Bak’s three-dimensional structure to find out precisely how the antibody activated Bak. “It is well known that Bak can be activated by a class of proteins called ‘BH3-only proteins’ that bind to a groove on Bak. We were surprised to find that despite our antibody binding to a completely different site on Bak, it could still trigger activation,” Kluck said. “The advantage of our antibody is that it can’t be ‘mopped up’ and neutralized by pro-survival proteins in the cell, potentially reducing the chance of drug resistance occurring.”
Drugs that target this new activation site could be useful in combination with other therapies that promote cell death by mimicking the BH3-only proteins. The researchers are now working with collaborators to develop their antibody into a drug that can access Bak inside cells.
Their findings have just been published in the open-access journal Nature Communications. The research was supported by the National Health and Medical Research Council, the Australian Research Council, the Victorian State Government Operational Infrastructure Support Scheme, and the Victorian Life Science Computation Initiative.
Abstract of Identification of an activation site in Bak and mitochondrial Bax triggered by antibodies
During apoptosis, Bak and Bax are activated by BH3-only proteins binding to the α2–α5 hydrophobic groove; Bax is also activated via a rear pocket. Here we report that antibodies can directly activate Bak and mitochondrial Bax by binding to the α1–α2 loop. A monoclonal antibody (clone 7D10) binds close to α1 in non-activated Bak to induce conformational change, oligomerization, and cytochrome c release. Anti-FLAG antibodies also activate Bak containing a FLAG epitope close to α1. An antibody (clone 3C10) to the Bax α1–α2 loop activates mitochondrial Bax, but blocks translocation of cytosolic Bax. Tethers within Bak show that 7D10 binding directly extricates α1; a structural model of the 7D10 Fab bound to Bak reveals the formation of a cavity under α1. Our identification of the α1–α2 loop as an activation site in Bak paves the way to develop intrabodies or small molecules that directly and selectively regulate these proteins.
“Cure” is a word that’s dominated the rhetoric in the war on cancer for decades. But it’s a word that medical professionals tend to avoid. While the American Cancer Society reports that cancer treatment has improved markedly over the decades and the five-year survival rate is impressively high for many cancers, oncologists still refrain from declaring their cancer-free patients cured. Why?
Patients are declared cancer-free (also called complete remission) when there are no more signs of detectable disease.
However, minuscule clusters of cancer cells below the detection level can remain in a patient’s body after treatment. Moreover, such small clusters of straggler cells may undergo metastasis, where they escape from the initial tumor into the bloodstream and ultimately settle in a distant site, often a vital organ such as the lungs, liver or brain.
When a colony of these metastatic cells reaches a detectable size, the patient is diagnosed with recurrent metastatic cancer. About one in three breast cancer patients diagnosed with early-stage cancer later develop metastatic disease, usually within five years of initial remission.
By the time metastatic cancer becomes evident, it is much more difficult to treat than when it was originally diagnosed.
What if these metastatic cells could be detected earlier, before they established a “foothold” in a vital organ? Better yet, could these metastatic cancer cells be intercepted, preventing them them from lodging in a vital organ in the first place?
The implant is a tiny porous polymer disc (basically a miniature sponge, no larger than a pencil eraser) that can be inserted just under a patient’s skin. Implantation triggers the immune system’s “foreign body response,” and the implant starts to soak up immune cells that travel to it. If the implant can catch mobile immune cells, then why not mobile metastatic cancer cells?
We gave implants to mice specially bred to model metastatic breast cancer. When the mice had palpable tumors but no evidence of metastatic disease, the implant was removed and analyzed.
Cancer cells were indeed present in the implant, while the other organs (potential destinations for metastatic cells) still appeared clean. This means that the implant can be used to spot previously undetectable metastatic cancer before it takes hold in an organ.
For patients with cancer in remission, an implant that can detect tumor cells as they move through the body would be a diagnostic breakthrough. But having to remove it to see if it has captured any cancer cells is not the most convenient or pleasant detection method for human patients.
Detecting cancer cells with noninvasive imaging
There could be a way around this, though: a special imaging method under development at Northwestern University called Inverse Spectroscopic Optical Coherence Tomography (ISOCT). ISOCT detects molecular-level differences in the way cells in the body scatter light. And when we scan our implant with ISOCT, the light scatter pattern looks different when it’s full of normal cells than when cancer cells are present. In fact, the difference is apparent when even as few as 15 out of the hundreds of thousands of cells in the implant are cancer cells.
There’s a catch – ISOCT cannot penetrate deep into tissue. That means it is not a suitable imaging technology for finding metastatic cells buried deep in internal organs. However, when the cancer cell detection implant is located just under the skin, it may be possible to detect cancer cells trapped in it using ISOCT. This could offer an early warning sign that metastatic cells are on the move.
This early warning could prompt doctors to monitor their patients more closely or perform additional tests. Conversely, if no cells are detected in the implant, a patient still in remission could be spared from unneeded tests.
The ISOCT results show that noninvasive imaging of the implant is feasible. But it’s a method still under development, and thus it’s not widely available. To make scanning easier and more accessible, we’re working to adapt more ubiquitous imaging technologies like ultrasound to detect tiny quantities of tumor cells in the implant.
Besides providing a way to detect tiny numbers of cancer cells before they can form new tumors in other parts of the body, our implant offers an even more intriguing possibility: diverting metastatic cells away from vital organs, and sequestering them where they cannot cause any damage.
In our mouse studies, we found that metastatic cells got caught in the implant before they were apparent in vital organs. When metastatic cells eventually made their way into the organs, the mice with implants still had significantly fewer tumor cells in their organs than implant-free controls. Thus, the implant appears to provide a therapeutic benefit, most likely by taking the metastatic cells it catches out of the circulation, preventing them from lodging anywhere vital.
Interestingly, we have not seen cancer cells leave the implant once trapped, or form a secondary tumor in the implant. Ongoing work aims to learn why this is. Whether the cells can stay safely immobilized in the implant or if it would need to be removed periodically will be important questions to answer before the implant could be used in human patients.
What the future may hold
For now, our work aims to make the implant more effective at drawing and detecting cancer cells. Since we tested the implant with metastatic breast cancer cells, we also want to see if it will work on other types of cancer. Additionally, we’re studying the cells the implant traps, and learning how the implant interacts with the body as a whole. This basic research should give us insight into the process of metastasis and how to treat it.
In the future (and it might still be far off), we envision a world where recovering cancer patients can receive a detector implant to stand guard for disease recurrence and prevent it from happening. Perhaps the patient could even scan their implant at home with a smartphone and get treatment early, when the disease burden is low and the available therapies may be more effective. Better yet, perhaps the implant could continually divert all the cancer cells away from vital organs on its own, like Iron Man’s electromagnet that deflects shrapnel from his heart.
This solution is still not a “cure.” But it would transform a formidable disease that one out of three cancer survivors would otherwise ultimately die from into a condition with which they could easily live.
New PSA Test Examines Protein Structures to Detect Prostate Cancers
5/16/2016 by Cleveland Clinic
A promising new test is detecting prostate cancer more precisely than current tests, by identifying molecular changes in the prostate specific antigen (PSA) protein, according to Cleveland Clinic research presented today at the American Urological Association annual meeting.
The study – part of an ongoing multicenter prospective clinical trial – found that the IsoPSATM test can also differentiate between high-risk and low-risk disease, as well as benign conditions.
Although widely used, the current PSA test relies on detection strategies that have poor specificity for cancer – just 25 percent of men who have a prostate biopsy due to an elevated PSA level actually have prostate cancer, according to the National Cancer Institute – and an inability to determine the aggressiveness of the disease.
The IsoPSA test, however, identifies prostate cancer in a new way. Developed by Cleveland Clinic, in collaboration with Cleveland Diagnostics, Inc., IsoPSA identifies the molecular structural changes in protein biomarkers. It is able to detect cancer by identifying these structural changes, as opposed to current tests that simply measure the protein’s concentration in a patient’s blood.
“While the PSA test has undoubtedly been one of the most successful biomarkers in history, its limitations are well known. Even currently available prostate cancer diagnostic tests rely on biomarkers that can be affected by physiological factors unrelated to cancer,” said Eric Klein, M.D., chair of Cleveland Clinic’s Glickman Urological & Kidney Institute. “These study results show that using structural changes in PSA protein to detect cancer is more effective and can help prevent unneeded biopsies in low-risk patients.”
The clinical trial involves six healthcare institutions and 132 patients, to date. It examined the ability of IsoPSA to distinguish patients with and without biopsy-confirmed evidence of cancer. It also evaluated the test’s precision in differentiating patients with high-grade (Gleason = 7) cancer from those with low-grade (Gleason = 6) disease and benign findings after standard ultrasound-guided biopsy of the prostate.
Substituting the IsoPSA structure-based composite index for the standard PSA resulted in improvement in diagnostic accuracy. Compared with serum PSA testing, IsoPSA performed better in both sensitivity and specificity.
“We took an ‘out of the box’ approach that has shown success in detecting prostate cancer but also has the potential to address other clinically important questions such as clinical surveillance of patients after treatment,” said Mark Stovsky, M.D., staff member, Cleveland Clinic Glickman Urological & Kidney Institute’s Department of Urology. Stovsky has a leadership position (Chief Medical Officer) and investment interest in Cleveland Diagnostics, Inc. “In general, the clinical utility of prostate cancer early detection and screening tests is often limited by the fact that biomarker concentrations may be affected by physiological processes unrelated to cancer, such as inflammation, as well as the relative lack of specificity of these biomarkers to the cancer phenotype. In contrast, clinical research data suggests that the IsoPSA assay can interrogate the entire PSA isoform distribution as a single stand-alone diagnostic tool which can reliably identify structural changes in the PSA protein that correlate with the presence or absence and aggressiveness of prostate cancer.”
Point of Care, Highly Accurate Cervical Cancer Screening
Fifty-five million times a year, American women go to their gynecologist for a Pap Smear. After waiting a few weeks for the results, more than 3.5 million of them are called back to the physician for a follow up visualization of the cervix. Beyond the stress related to possibly having cancer, the women are then subjected to a colposcopic exam, and all too often, a painful biopsy. Then more stressful waiting for a final diagnosis from the pathologist.
Cervical cancer develops slowly, allowing for successful treatment, when identified on time. Regions with high screening compliancy have low mortality rates from this cancer. In the US, for instance, where screening rates are close to 90%, only 4,200 women die from cervical cancer, annually, or 2.6 women per 100,000. However, the screening process in the developed world is long, complicated and not optimized.
In developing regions however, cervical cancer is a leading cause of women death. Over 85% of the total deaths from this cancer are in developing countries. Regions suffering from low screening rates include not only Africa, India and China, but many Eastern European countries as well. According to an OECD report from 2014, the cervical cancer screening rates in Romania and Hungary are as low as 14.6% and 36.7% respectively. The mortality rates in these countries are high, 16 in 100,000 women in Romania and 7.7 in 100,000 in Hungary.
The current screening process for cervical cancer detection is long, beginning with a Pap or HPV test. Cytology results take weeks to receive. A positive result requires follow-up testing by colposcopy and often biopsy. In countries where there is little access to medical care, or where screening compliancy is low, the chances of successful detection via this multi-step process are small. Developing regions and non-compliant countries require a point of care diagnostic method, which eliminates the need for return visits.
Additional limitations to cervical cancer screening are the low sensitivity and specificity rates of Pap tests and the high false positive rates of HPV test, leading to unnecessary colposcopies. Both cytology and colposcopy testing are highly dependent on operator proficiency for accurate diagnosis.
Biop has developed a new technology for the optimization of this process, into one, three minute, painless optical scan. The vaginal probe uses advanced optical, imaging and non-imaging technologies to identify and classify epithelium based cancers and pre-cancerous lesions. The probe is inserted into the vaginal canal, and scans the entire cervix. The resulting images and optical signatures created from the light, and captured by the sensors, are analyzed by the proprietary algorithm. The result is two pictures, on the physician’s screen; a high resolution photograph of the patient’s cervix, immediately next to a hot/cold map indicating a precise classification and location of any diseased lesions.
Deep learning applied to drug discovery and repurposing
Deep neural networks for drug discovery (credit: Insilico Medicine, Inc.)
Scientists from Insilico Medicine, Inc. have trained deep neural networks (DNNs) to predict the potential therapeutic uses of 678 drugs, using gene-expression data obtained from high-throughput experiments on human cell lines from Broad Institute’s LINCS databases and NIH MeSH databases.
The supervised deep-learning drug-discovery engine used the properties of small molecules, transcriptional data, and literature to predict efficacy, toxicity, tissue-specificity, and heterogeneity of response.
“We used LINCS data from Broad Institute to determine the effects on cell lines before and after incubation with compounds, co-author and research scientist Polina Mamoshina explained to KurzweilIAI.
“We used gene expression data of total mRNA from cell lines extracted and measured before incubation with compound X and after incubation with compound X to identify the response on a molecular level. The goal is to understand how gene expression (the transcriptome) will change after drug uptake. It is a differential value, so we need a reference (molecular state before incubation) to compare.”
The research is described in a paper in the upcoming issue of the journal Molecular Pharmaceutics.
Helping pharmas accelerate R&D
Alex Zhavoronkov, PhD, Insilico Medicine CEO, who coordinated the study, said the initial goal of their research was to help pharmaceutical companies significantly accelerate their R&D and increase the number of approved drugs. “In the process we came up with more than 800 strong hypotheses in oncology, cardiovascular, metabolic, and CNS spaces and started basic validation,” he said.
The team measured the “differential signaling pathway activation score for a large number of pathways to reduce the dimensionality of the data while retaining biological relevance.” They then used those scores to train the deep neural networks.*
“This study is a proof of concept that DNNs can be used to annotate drugs using transcriptional response signatures, but we took this concept to the next level,” said Alex Aliper, president of research, Insilico Medicine, Inc., lead author of the study.
Via Pharma.AI, a newly formed subsidiary of Insilico Medicine, “we developed a pipeline for in silico drug discovery — which has the potential to substantially accelerate the preclinical stage for almost any therapeutic — and came up with a broad list of predictions, with multiple in silico validation steps that, if validated in vitro and in vivo, can almost double the number of drugs in clinical practice.”
Despite the commercial orientation of the companies, the authors agreed not to file for intellectual property on these methods and to publish the proof of concept.
Deep-learning age biomarkers
According to Mamoshina, earlier this month, Insilico Medicine scientists published the first deep-learned biomarker of human age — aiming to predict the health status of the patient — in a paper titled “Deep biomarkers of human aging: Application of deep neural networks to biomarker development” by Putin et al, in Aging; and an overview of recent advances in deep learning in a paper titled “Applications of Deep Learning in Biomedicine” by Mamoshina et al., also in Molecular Pharmaceutics.
Insilico Medicine is located in the Emerging Technology Centers at Johns Hopkins University in Baltimore, Maryland, in collaboration with Datalytic Solutions and Mind Research Network.
* In this study, scientists used the perturbation samples of 678 drugs across A549, MCF-7 and PC-3 cell lines from the Library of Integrated Network-Based Cellular Signatures (LINCS) project developed by the National Institutes of Health (NIH) and linked those to 12 therapeutic use categories derived from MeSH (Medical Subject Headings) developed and maintained by the National Library of Medicine (NLM) of the NIH.
To train the DNN, scientists utilized both gene level transcriptomic data and transcriptomic data processed using a pathway activation scoring algorithm, for a pooled dataset of samples perturbed with different concentrations of the drug for 6 and 24 hours. Cross-validation experiments showed that DNNs achieve 54.6% accuracy in correctly predicting one out of 12 therapeutic classes for each drug.
One peculiar finding of this experiment was that a large number of drugs misclassified by the DNNs had dual use, suggesting possible application of DNN confusion matrices in drug repurposing. FutureTechnologies Media Group | Video presentation Insilico medicine
Abstract of Deep learning applications for predicting pharmacological properties of drugs and drug repurposing using transcriptomic data
Deep learning is rapidly advancing many areas of science and technology with multiple success stories in image, text, voice and video recognition, robotics and autonomous driving. In this paper we demonstrate how deep neural networks (DNN) trained on large transcriptional response data sets can classify various drugs to therapeutic categories solely based on their transcriptional profiles. We used the perturbation samples of 678 drugs across A549, MCF-7 and PC-3 cell lines from the LINCS project and linked those to 12 therapeutic use categories derived from MeSH. To train the DNN, we utilized both gene level transcriptomic data and transcriptomic data processed using a pathway activation scoring algorithm, for a pooled dataset of samples perturbed with different concentrations of the drug for 6 and 24 hours. When applied to normalized gene expression data for “landmark genes,” DNN showed cross-validation mean F1 scores of 0.397, 0.285 and 0.234 on 3-, 5- and 12-category classification problems, respectively. At the pathway level DNN performed best with cross-validation mean F1 scores of 0.701, 0.596 and 0.546 on the same tasks. In both gene and pathway level classification, DNN convincingly outperformed support vector machine (SVM) model on every multiclass classification problem. For the first time we demonstrate a deep learning neural net trained on transcriptomic data to recognize pharmacological properties of multiple drugs across different biological systems and conditions. We also propose using deep neural net confusion matrices for drug repositioning. This work is a proof of principle for applying deep learning to drug discovery and development.
A novel nanoscale organic transistor-based biosensor that can detect molecules associated with neurodegenerative diseases and some types of cancer has been developed by researchers at the National Nanotechnology Laboratory (LNNano) in Brazil.
The transistor, mounted on a glass slide, contains the reduced form of the peptide glutathione (GSH), which reacts in a specific way when it comes into contact with the enzyme glutathione S-transferase (GST), linked to Parkinson’s, Alzheimer’s and breast cancer, among other diseases.
Sensitive water-gated copper phthalocyanine (CuPc) thin-film transistor (credit: Rafael Furlan de Oliveira et al./Organic Electronics)
“The device can detect such molecules even when they’re present at very low levels in the examined material, thanks to its nanometric sensitivity,” explained Carlos Cesar Bof Bufon, Head of LNNano’s Functional Devices & Systems Lab (DSF).
Bufon said the system can be adapted to detect other substances by replacing the analytes (detection compounds). The team is working on paper-based biosensors to further lower the cost, improve portability, and facilitate fabrication and disposal.
The research is published in the journal Organic Electronics.
Abstract of Water-gated phthalocyanine transistors: Operation and transduction of the peptide–enzyme interaction
The use of aqueous solutions as the gate medium is an attractive strategy to obtain high charge carrier density (1012 cm−2) and low operational voltages (<1 V) in organic transistors. Additionally, it provides a simple and favorable architecture to couple both ionic and electronic domains in a single device, which is crucial for the development of novel technologies in bioelectronics. Here, we demonstrate the operation of transistors containing copper phthalocyanine (CuPc) thin-films gated with water and discuss the charge dynamics at the CuPc/water interface. Without the need for complex multilayer patterning, or the use of surface treatments, water-gated CuPc transistors exhibited low threshold (100 ± 20 mV) and working voltages (<1 V) compared to conventional CuPc transistors, along with similar charge carrier mobilities (1.2 ± 0.2) x 10−3 cm2 V−1 s−1. Several device characteristics such as moderate switching speeds and hysteresis, associated with high capacitances at low frequencies upon bias application (3.4–12 μF cm−2), indicate the occurrence of interfacial ion doping. Finally, water-gated CuPc OTFTs were employed in the transduction of the biospecific interaction between tripeptide reduced glutathione (GSH) and glutathione S-transferase (GST) enzyme, taking advantage of the device sensitivity and multiparametricity.
The study offers understanding of potential therapeutic targets.
Building on data from The Cancer Genome Atlas (TCGA) project, a multi-institutional team of scientists have completed the first large-scale “proteogenomic” study of breast cancer, linking DNA mutations to protein signaling and helping pinpoint the genes that drive cancer. Conducted by members of the National Cancer Institute’s Clinical Proteomic Tumor Analysis Consortium (CPTAC), including Baylor College of Medicine, Broad Institute of MIT and Harvard, Fred Hutchinson Cancer Research Center, New York University Langone Medical Center, and Washington University School of Medicine, the study takes aim at proteins, the workhorses of the cell, and their modifications to better understand cancer.
Appearing in the Advance Online Publication of Nature, the study illustrates the power of integrating genomic and proteomic data to yield a more complete picture of cancer biology than either analysis could do alone. The effort produced a broad overview of the landscape of the proteome (all the proteins found in a cell) and the phosphoproteome (the sites at which proteins are tagged by phosphorylation, a chemical modification that drives communication in the cell) across a set of 77 breast cancer tumors that had been genomically characterized in the TCGA project. Although the TCGA produced an extensive catalog of somatic mutations found in cancer, the effects of many of those mutations on cellular functions or patients’ outcomes are unknown.
In addition, not all mutated genes are true “drivers” of cancer — some are merely “passenger” mutations that have little functional consequence. And some mutations are found within very large DNA regions that are deleted or present in extra copies, so winnowing the list of candidate genes by studying the activity of their protein products can help identify therapeutic targets. “We don’t fully understand how complex cancer genomes translate into the driving biology that causes relapse and mortality,” said Matthew Ellis, director of the Lester and Sue Smith Breast Center at Baylor College of Medicine and a senior author of the paper.
“These findings show that proteogenomic integration could one day prove to be a powerful clinical tool, allowing us to traverse the large knowledge gap between cancer genomics and clinical action.” In this study, the researchers at the Broad Institute analyzed breast tumors using accurate mass, high-resolution mass spectrometry, a technology that extends the coverage of the proteome far beyond the coverage that can be achieved by traditional antibody-based methods. This allowed them to scale their efforts and quantify more than 12,000 proteins and 33,000 phosphosites, an extremely deep level of coverage.
Scripps scientists have designed a drug candidate that decreases growth of breast cancer cells.
In a development that could lead to a new generation of drugs to precisely treat a range of diseases, scientists from the Florida campus of The Scripps Research Institute (TSRI) have for the first time designed a drug candidate that decreases the growth of tumor cells in animal models in one of the hardest to treat cancers—triple negative breast cancer.
“This is the first example of taking a genetic sequence and designing a drug candidate that works effectively in an animal model against triple negative breast cancer,” said TSRI Professor Matthew Disney. “The study represents a clear breakthrough in precision medicine, as this molecule only kills the cancer cells that express the cancer-causing gene—not healthy cells. These studies may transform the way the lead drugs are identified—by using the genetic makeup of a disease.”
The study, published by the journal Proceedings of the National Academy of Sciences, demonstrates that the Disney lab’s compound, known as Targaprimir-96, triggers breast cancer cells to kill themselves via programmed cell death by precisely targeting a specific RNA that ignites the cancer.
Short-Cut to Drug Candidates
While the goal of precision medicine is to identify drugs that selectively affect disease-causing biomolecules, the process has typically involved time-consuming and expensive high-throughput screens to test millions of potential drug candidates to identify those few that affect the target of interest. Disney’s approach eliminates these screens.
The new study uses the lab’s computational approach called Inforna, which focuses on developing designer compounds that bind to RNA folds, particularly microRNAs.
MicroRNAs are short molecules that work within all animal and plant cells, typically functioning as a “dimmer switch” for one or more genes, binding to the transcripts of those genes and preventing protein production. Some microRNAs have been associated with diseases. For example, microRNA-96, which was the target of the new study, promotes cancer by discouraging programmed cell death, which can rid the body of cells that grow out of control.
In the new study, the drug candidate was tested in animal models over a 21-day course of treatment. Results showed decreased production of microRNA-96 and increased programmed cell death, significantly reducing tumor growth. Since targaprimir-96 was highly selective in its targeting, healthy cells were unaffected.
In contrast, Disney noted, a typical cancer therapeutic targets and kills cells indiscriminately, often leading to side effects that can make these drugs difficult for patients to tolerate.
Benjamin Zealley and Aubrey D.N.J. de Grey Commentary on Some Recent Theses Relevant to Combating Aging: June 2015
Cancer Autoantibody Biomarker Discovery and Validation Using Nucleic Acid Programmable Protein Array Jie Wang, PhD, Arizona State University
Currently in the United States, many patients with cancer do not benefit from population-based screening due to challenges associated with the existing cancer screening scheme. Blood-based diagnostic assays have the potential to detect diseases in a non-invasive way. Proteins released from small early tumors may only be present intermittently and are diluted to tiny concentrations in the blood, making them difficult to use as biomarkers. However, they can induce autoantibody (AAb) responses, which can amplify the signal and persist in the blood even if the antigen is gone. Circulating autoantibodies are a promising class of molecules that have the potential to serve as early detection biomarkers for cancers. This PhD thesis aims to screen for autoantibody biomarkers for the early detection of two deadly cancers, basal-like breast cancer and lung adenocarcinoma. First, a method was developed to display proteins in both native and denatured conformations on a protein array. This method adopted a novel protein tag technology, called a HaloTag, to immobilize proteins covalently on the surface of a glass slide. The covalent attachment allowed these proteins to endure harsh treatment without becoming dissociated from the slide surface, which enabled the profiling of antibody responses against both conformational and linear epitopes. Next, a plasma screening protocol was optimized to increase significantly the signal-to-noise ratio of protein array–based AAb detection. Following this, the AAb responses in basal-like breast cancer were explored using nucleic acid programmable protein arrays (NAPPA) containing 10,000 full-length human proteins in 45 cases and 45 controls. After verification in a large sample set (145 basal-like breast cancer cases, 145 controls, 70 non-basal breast cancer) by enzyme-linked immunosorbent assay (ELISA), a 13-AAb classifier was developed to differentiate patients from controls with a sensitivity of 33% at 98% specificity. A similar approach was also applied to the lung cancer study to identify AAbs that distinguished lung cancer patients from computed tomography–positive benign pulmonary nodules (137 lung cancer cases, 127 smoker controls, 170 benign controls). In this study, two panels of AAbs were discovered that showed promising sensitivity and specificity. Six out of eight AAb targets were also found to have elevated mRNA levels in lung adenocarcinoma patients using TCGA data. These projects as a whole provide novel insights into the association between AAbs and cancer, as well as general B cell antigenicity against self-proteins.
Comment: There are two widely supported models for cancer development and progression—the clonal evolution (CE) model and the cancer stem cell (CSC) model. Briefly, the former claims that most or all cells in a tumor contribute to its maintenance; as newer and more aggressive clones develop by random mutation, they become responsible for driving growth. The range of different mutational profiles generated is assumed to be large enough to account for disease recurrence after therapy (due to rare resistant clones) and metastasis (clones arising with the ability to travel to distant sites). The CSC model instead asserts that a small number of mutated stem cells are the origin of the primary cell mass, drive metastasis through the intermittent release of undifferentiated, highly mobile progeny, and account for recurrence due to a generally quiescent metabolic profile conferring potent resistance to chemotherapy. In either case, the immunological visibility of an early tumor may be highly sporadic. Clones arising early in CE differ little in proteomic terms from healthy host cells; those that do trigger a response are unlikely to have acquired robust resistance to immune attack, so are destroyed quickly in favor of their stealthier brethren. Likewise, CSCs share some of the immune privilege of normal stem cells and, due to their inherent ability to produce differentiated progeny with distinct proteomic signatures, are partially protected from attacks on their descendants. Consequently, such well-hidden cells may remain in the body for years to decades. The autoantibody panel developed in this study for basal-like breast cancer exhibits exceptional specificity despite a comparatively small training set. Given its ease of application, this suggests great promise for a more exhaustively trained classifier as a populationlevel screening tool.
Condition-Specific Differential Sub-Network Analysis for Biological Systems Deepali Jhamb, PhD, Indiana University
Biological systems behave differently under different conditions. Advances in sequencing technology over the last decade have led to the generation of enormous amounts of condition-specific data. However, these measurements often fail to identify low-abundance genes and proteins that can be biologically crucial. In this work, a novel textmining system was first developed to extract condition-specific proteins from the biomedical literature. The literaturederived data was then combined with proteomics data to construct condition-specific protein interaction networks. Furthermore, an innovative condition-specific differential analysis approach was designed to identify key differences, in the form of sub-networks, between any two given biological systems. The framework developed here was implemented to understand the differences between limb regenerationcompetent Ambystoma mexicanum and regeneration-deficient Xenopus laevis. This study provides an exhaustive systems-level analysis to compare regeneration competent and deficient sub-networks to show how different molecular entities inter-connect with each other and are rewired during the formation of an accumulation blastema in regenerating axolotl limbs. This study also demonstrates the importance of literature-derived knowledge, specific to limb regeneration, to augment the systems biology analysis. Our findings show that although the proteins might be common between the two given biological conditions, they can have a high dissimilarity based on their biological and topological properties in the sub-network. The knowledge gained from the distinguishing features of limb regeneration in amphibians can be used in future to induce regeneration chemically in mammalian systems. The approach developed in this dissertation is scalable and adaptable to understanding differential sub-networks between any two biological systems. This methodology will not only facilitate the understanding of biological processes and molecular functions that govern a given system, but will also provide novel intuitions about the pathophysiology of diseases/conditions.
Comment: We have long advocated a principle of directly comparing young and old bodies as a means to identify the classes of physical damage that accumulate in the body during aging. This approach circumvents our ignorance of the full etiology of each particular disease manifestation, a phenomenally difficult question given the ethical issues of experimenting on human subjects, the lengthy ‘‘incubation time’’ of aging-related diseases, and the complex interconnections between their risk factors—innate and environmental. Repairing such damage has the potential to prevent pathology before symptoms appear, an approach now becoming increasingly mainstream.11 However, a naı¨ve comparison faces a number of difficulties, even given a sufficiently large sample set to compensate for inter-individual variation. Most importantly, the causal significance of a given species cannot be reliably determined from its simple prevalence.12 The catalytic nature of cell biology means that those entities whose abundance changes the most profoundly in absolute terms are quite unlikely to be the drivers of that change and may even spontaneously revert to baseline levels in the absence of on-going stimulation. Meanwhile, functionality is often heavily influenced independently of abundance by post-translational modifications that may escape direct detection. Sub-network analysis uses computational means to identify groups of genes and/or proteins that vary in a synchronized way with some parameter, indicating functional connectivity. The application of methods such as those developed here to the comparison of a wide range of younger and older conditions will facilitate the identification of processes—not merely individual factors—that are impaired with age, and thus will help greatly in identifying the optimal points for intervention.
Development of a Light Actuated Drug Delivery-on-Demand System Chase Linsley, PhD, University of California, Los Angeles
The need for temporal–spatial control over the release of biologically active molecules has motivated efforts to engineer novel drug delivery-on-demand strategies actuated via light irradiation. Many systems, however, have been limited to in vitro proof-of-concept due to biocompatibility issues with the photo-responsive moieties or the light wavelength, intensity, and duration. To overcome these limitations, the objective of this dissertation was to design a light-actuated drug delivery-on-demand strategy that uses biocompatible chromophores and safe wavelengths of light, thereby advancing the clinical prospects of light-actuated drug delivery-on-demand systems. This was achieved by: (1) Characterizing the photothermal response of biocompatible visible light and near-infrared-responsive chromophores and demonstrating the feasibility and functionality of the light actuated on-demand drug delivery system in vitro; and (2) designing a modular drug delivery-on-demand system that could control the release of biologically active molecules over an extended period of time. Three biocompatible chromophores—Cardiogreen, Methylene Blue, and riboflavin—were identified and demonstrated significant photothermal response upon exposure to near-infrared and visible light, and the amount of temperature change was dependent upon light intensity, wavelength, as well as chromophore concentration. As a proof-of-concept, pulsatile release of a model protein from a thermally responsive delivery vehicle fabricated from poly(N-isopropylacrylamide) was achieved over 4 days by loading the delivery vehicle with Cardiogreen and irradiating with near-infrared light. To extend the useful lifetime of the light-actuated drug delivery-on-demand system, a modular, reservoir-valve system was designed. Using poly(ethylene glycol) as a reservoir for model small molecule drugs combined with a poly(N-isopropylacrylamide) valve spiked with chromophore-loaded liposomes, pulsatile release was achieved over 7 days upon light irradiation. Ultimately, this drug delivery strategy has potential for clinical applications that require explicit control over the presentation of biologically active molecules. Further research into the design and fabrication of novel biocompatible thermally responsive delivery vehicles will aid in the advancement of the light-actuated drug delivery-on-demand strategy described here. Comment: Our combined comments on this thesis and the next one appear after the next abstract.
Light-Inducible Gene Regulation in Mammalian Cells Lauren Toth, PhD, Duke University
The growing complexity of scientific research demands further development of advanced gene regulation systems. For instance, the ultimate goal of tissue engineering is to develop constructs that functionally and morphologically resemble the native tissue they are expected to replace. This requires patterning of gene expression and control of cellular phenotype within the tissue-engineered construct. In the field of synthetic biology, gene circuits are engineered to elucidate mechanisms of gene regulation and predict the behavior of more complex systems. Such systems require robust gene switches that can quickly turn gene expression on or off. Similarly, basic science requires precise genetic control to perturb genetic pathways or understand gene function. Additionally, gene therapy strives to replace or repair genes that are responsible for disease. The safety and efficacy of such therapies require control of when and where the delivered gene is expressed in vivo.
Unfortunately, these fields are limited by the lack of gene regulation systems that enable both robust and flexible cellular control. Most current gene regulation systems do not allow for the manipulation of gene expression that is spatially defined, temporally controlled, reversible, and repeatable. Rather, they provide incomplete control that forces the user to choose to control gene expression in either space or time, and whether the system will be reversible or irreversible. The recent emergence of the field of optogenetics—the ability to control gene expression using light—has made it possible to regulate gene expression with spatial, temporal, and dynamic control. Light-inducible systems provide the tools necessary to overcome the limitations of other gene regulation systems, which can be slow, imprecise, or cumbersome to work with. However, emerging light-inducible systems require further optimization to increase their efficiency, reliability, and ease of use.
Initially, we engineered a light-inducible gene regulation system that combines zinc finger protein technology and the light-inducible interaction between Arabidopsis thaliana plant proteins GIGANTEA (GI) and the light oxygen voltage (LOV) domain of FKF1. Zinc finger proteins (ZFPs) can be engineered to target almost any DNA sequence through tandem assembly of individual zinc finger domains that recognize a specific 3-bp DNA sequence. Fusion of three different ZFPs to GI (GI-ZFP) successfully targeted the fusion protein to the specific DNA target sequence of the ZFP. Due to the interaction between GI and LOV, co-expression of GI-ZFP with a fusion protein consisting of LOV fused to three copies of the VP16 transactivation domain (LOV-VP16) enabled blue-light dependent recruitment of LOV-VP16 to the ZFP target sequence. We showed that placement of three to nine copies of a ZFP target sequence upstream of a luciferase or enhanced green fluorescent protein (eGFP) transgene enabled expression of the transgene in response to blue light. Gene activation was both reversible and tunable on the basis of duration of light exposure, illumination intensity, and the number of ZFP binding sites upstream of the transgene. Gene expression could also be patterned spatially by illuminating the cell culture through photomasks containing various patterns.
Although this system was useful for controlling the expression of a transgene, for many applications it is useful to control the expression of a gene in its natural chromosomal position. Therefore, we capitalized on recent advances in programmed gene activation to engineer an optogenetic tool that could easily be targeted to new, endogenous DNA sequences without re-engineering the light inducible proteins. This approach took advantage of CRISPR/Cas9 technology, which uses a gene-specific guide RNA (gRNA) to facilitate Cas9 targeting and binding to a desired sequence, and the light-inducible heterodimerizers CRY2 and CIB1 from Arabidopsis thaliana to engineer a lightactivated CRISPR/Cas9 effector (LACE) system. We fused the full-length (FL) CRY2 to the transcriptional activator VP64 (CRY2FL-VP64) and the amino-terminal fragment of CIB1 to the amino, carboxyl, or amino and carboxyl terminus of a catalytically inactive Cas9. When CRY2-VP64 and one of the CIBN/dCas9 fusion proteins are expressed with a gRNA, the CIBN/dCas9 fusion protein localizes to the gRNA target. In the presence of blue light, CRY2FL binds to CIBN, which translocates CRY2FL-VP64 to the gene target and activates transcription. Unlike other optogenetic systems, the LACE system can be targeted to new endogenous loci by solely manipulating the specificity of the gRNA without having to re-engineer the light-inducible proteins. We achieved light-dependent activation of the IL1RN, HBG1/2, or ASCL1 genes by delivery of the LACE system and four gene-specific gRNAs per promoter region. For some gene targets, we achieved equivalent activation levels to cells that were transfected with the same gRNAs and the synthetic transcription factor dCas9-VP64. Gene activation was also shown to be reversible and repeatable through modulation of the duration of blue light exposure, and spatial patterning of gene expression was achieved using an eGFP reporter and a photomask.
Finally, we engineered a light-activated genetic ‘‘on’’ switch (LAGOS) that provides permanent gene expression in response to an initial dose of blue light illumination. LAGOS is a lentiviral vector that expresses a transgene only upon Cre recombinase–mediated DNA recombination. We showed that this vector, when used in conjunction with a light-inducible Cre recombinase system, could be used to express MyoD or the synthetic transcription factor VP64- MyoD in response to light in multiple mammalian cell lines, including primary mouse embryonic fibroblasts. We achieved light-mediated up-regulation of downstream myogenic markers myogenin, desmin, troponin T, and myosin heavy chains I and II as well as fusion of C3H10T1/2 cells into myotubes that resembled a skeletal muscle cell phenotype. We also demonstrated LAGOS functionality in vivo by engineering the vector to express human VEGF165 and human ANG1 in response to light. HEK 293T cells stably expressing the LAGOS vector and transiently expressing the light-inducible Cre recombinase proteins were implanted into mouse dorsal window chambers. Mice that were illuminated with blue light had increased micro-vessel density compared to mice that were not illuminated. Analysis of human vascular endothelial growth factor (VEGF) and human ANG1 levels by enzyme-linked immunosorbent assay (ELISA) revealed statistically higher levels of VEGF and ANG1 in illuminated mice compared to non-illuminated mice.
In summary, the objective of this work was to engineer robust light-inducible gene regulation systems that can control genes and cellular fate in a spatial and temporal manner. These studies combine the rapid advances in gene targeting and activation technology with natural light-inducible plant protein interactions. Collectively, this thesis presents several optogenetic systems that are expected to facilitate the development of multicellular cell and tissue constructs for use in tissue engineering, synthetic biology, gene therapy, and basic science both in vitro and in vivo.
Comment: Although it is easy to characterize technological progress as following in the wake of scientific discoveries, the reverse is almost equally true; advances in technique open the door to types of experiment previously intractable or impossible. Such is currently the case for the field of optically controlled biotechnology, which has exploded into prominence, particularly over the last half-decade. Light of an appropriate wavelength can penetrate mammalian tissues to a depth of up to a couple of centimeters, rendering much of the living body accessible to optical study and control—still more if the detector/source is integrated into an endoscopic or fiber optic probe. Techniques borrowed from the semiconductor industry allow patterns of illumination to be controlled down to the nanometer scale, ideal for addressing individual cells. The highly controlled time course of such experiments, as compared to traditional means of gene activation, such as the addition of a chemical agent to the medium, eliminates confounding variables, and simplifies data analysis. Furthermore, this level of immediate control opens the door to closed-loop systems where the activity of entities under optical control can be continuously tuned in relation to some parameter(s). In the first of these two illuminating theses, a vehicle is developed that permits light-driven release of a small molecule. Such a system could be employed to target a systemically administered antibiotic or anti-neoplastic agent to a site of infection or cancer while sparing other bodily tissues from toxicity. Because most modern drugs cannot be produced in the body, even given arbitrarily good control of cellular biochemistry, this technique will have lasting value in numerous clinical contexts. In the second thesis, the level of precision achieved is even more profound; the CRISPR/Cas9 system has received much recent attention13 in its own right for its capacity to target arbitrary genetic sequences without an arduous protein-engineering step. The LACE system described stands to permit genetic manipulation with almost arbitrarily good spatial, temporal, and genomic site-specific control, using only means available to a typical university laboratory.
Targeting T Cells for the Immune-Modulation of Human Diseases Regina Lin, PhD, Duke University
Dysregulated inflammation underlies the pathogenesis of a myriad of human diseases ranging from cancer to autoimmunity. As coordinators, executers, and sentinels of host immunity, T cells represent a compelling target population for immune-modulation. In fact, the antigen-specificity, cytotoxicity, and promise of long-lived of immune-protection make T cells ideal vehicles for cancer immunotherapy. Interventions for autoimmune disorders, on the other hand, aim to dampen T cell–mediated inflammation and promote their regulatory functions. Although significant strides have been made in targeting T cells for immune modulation, current approaches remain less than ideal and leave room for improvement. In this dissertation, I seek to improve on current T cell-targeted immunotherapies, by identifying and pre-clinically characterizing their mechanisms of action and in vivo therapeutic efficacy.
CD8+ cytotoxic T cells have potent anti-tumor activity and therefore are leading candidates for use in cancer immunotherapy. The application of CD8+ T cells for clinical use has been limited by the susceptibility of ex vivo– expanded CD8+ T cells to become dysfunctional in response to immunosuppressive microenvironments. To enhance the efficacy of adoptive cell transfer therapy (ACT), we established a novel microRNA (miRNA)-targeting approach that augments CTL cytotoxicity and preserves immunocompetence. Specifically, we screened for miRNAs that modulate cytotoxicity and identified miR-23a as a strong functional repressor of the transcription factor Blimp-1, which promotes CTL cytotoxicity and effector cell differentiation. In a cohort of advanced lung cancer patients, miR- 23a was up-regulated in tumor-infiltrating CD8+ T cells, and its expression correlated with impaired anti-tumor potential of patient CD8+ T cells. We determined that tumor-derived transforming growth factor-b (TGF-b) directly suppresses CD8+ T cell immune function by elevating miR-23a and down-regulating Blimp-1. Functional blockade of miR-23a in human CD8+ T cells enhanced granzyme B expression; and in mice with established tumors, immunotherapy with just a small number of tumor-specific CD8+ T cells in which miR-23a was inhibited robustly hindered tumor progression. Together, our findings provide a miRNA-based strategy that subverts the immunosuppression of CD8+ T cells that is often observed during adoptive cell transfer tumor immunotherapy and identify a TGF-bmediated tumor immune-evasion pathway
Having established that miR-23a-inhibition can enhance the quality and functional resilience of anti-tumor CD8+ T cells, especially within the immune-suppressive tumor microenvironment, we went on to interrogate the translational applicability of this strategy in the context of chimeric antigen receptor (CAR)-modified CD8+ T cells. Although CAR T cells hold immense promise for ACT, CAR T cells are not completely curative due to their in vivo functional suppression by immune barriers—such as TGF-b—within the tumor microenvironment. Because TGF-b poses a substantial immune barrier in the tumor microenvironment, we sought to investigate whether inhibiting miR-23a in CAR T cells can confer immune competence to afford enhanced tumor clearance. To this end, we retrovirally transduced wild-type and miR-23a–deficient CD8+ T cells with the EGFRvIII-CAR, which targets the PepvIII tumorspecific epitope expressed by glioblastomas (GBM). Our in vitro studies demonstrated that while wild-type EGFRvIIICAR T cells were vulnerable to functional suppression by TGF-b, miR-23a abrogation rendered EGFRvIII-CAR T cells immune-resistant to TGF-b. Rigorous preclinical studies are currently underway to evaluate the efficacy of miR-23adeficient EGFRvIII-CAR T cells for GBM immunotherapy.
Last, we explored novel immune-suppressive therapies by the biological characterization of pharmacological agents that could target T cells. Although immune-suppressive drugs are classical therapies for a wide range of autoimmune diseases, they are accompanied by severe adverse effects. This motivated our search for novel immunesuppressive agents that are efficacious and lack undesirable side effects. To this end, we explored the potential utility of subglutinol A, a natural product isolated from the endophytic fungus Fusarium subglutinans. We showed that subglutinol A exerts multimodal immune-suppressive effects on activated T cells in vitro. Subglutinol A effectively blocked T cell proliferation and survival, while profoundly inhibiting pro-inflammatory interferon-c (IFN-c) and interleukin-17 (IL-17) production by fully differentiated effector Th1 and Th17 cells. Our data further revealed that subglutinol A might exert its anti-inflammatory effects by exacerbating mitochondrial damage in T cells, but not in innate immune cells or fibroblasts. Additionally, we demonstrated that subglutinol A significantly reduced lymphocytic infiltration into the footpad and ameliorated footpad swelling in the mouse model of Th1-driven delayed-type hypersensitivity. These results suggest the potential of subglutinol A as a novel therapeutic for inflammatory diseases.
Comment: Immunotherapy is among the most promising approaches to cancer treatment, having the specificity and scope to selectively target transformed cells wherever they may reside within the body and the potential to install a permanent defense against disease recurrence. By the time a typical cancer is clinically diagnosed, however, it has already found means to survive a prolonged period of potential immune attack. The mechanisms by which tumors evade immune surveillance are beginning to be elucidated,15,16 and include both direct suppression of effector cells and progressive editing of the host’s immune repertoire to disfavor future attack. It is inherently difficult to interfere with these defenses directly, due to the selection pressures in genetically heterogeneous neoplastic tissue. Much effort is thus being focused on methods for rendering therapeutically delivered immune cells resistant to their effects. The cytokine TGF-b is paradoxically known to function as both a tumor suppressor in healthy tissue and as a tumorderived species associated with multiple cancer-promoting activities, including enhanced immune evasion. This work identifies the pathway by which TGF-b compromises cytotoxic T cell function in the tumor microenvironment, and demonstrates an effective method for blocking this signal. In many clinical cases, however, editing of the patient’s immune repertoire has already removed or rendered anergic those immune cells able to recognize their cancer. Thus, the finding that blocking TGF-b signaling also appears to enhance the effectiveness of CAR-modified T cells— engineered with an antibody fragment targeting them with high affinity to a particular tumor-associated epitope—is a welcome addition to these already promising results.
Novel Fibonacci and non-Fibonacci structure in the sunflower: results of a citizen science experiment
Jonathan Swinton, Erinma Ochu, The MSI Turing’s Sunflower Consortium
This citizen science study evaluates the occurrence of Fibonacci structure in the spirals of sunflower (Helianthus annuus) seedheads. This phenomenon has competing biomathematical explanations, and our core premise is that observation of both Fibonacci and non-Fibonacci structure is informative for challenging such models. We collected data on 657 sunflowers. In our most reliable data subset, we evaluated 768 clockwise or anticlockwise parastichy numbers of which 565 were Fibonacci numbers, and a further 67 had Fibonacci structure of a predefined type. We also found more complex Fibonacci structures not previously reported in sunflowers. This is the third, and largest, study in the literature, although the first with explicit and independently checkable inclusion and analysis criteria and fully accessible data. This study systematically reports for the first time, to the best of our knowledge, seedheads without Fibonacci structure. Some of these are approximately Fibonacci, and we found in particular that parastichy numbers equal to one less than a Fibonacci number were present significantly more often than those one more than a Fibonacci number. An unexpected further result of this study was the existence of quasi-regular heads, in which no parastichy number could be definitively assigned.
Introduction
Fibonacci structure can be found in hundreds of different species of plants [1]. This has led to a variety of competing conceptual and mathematical models that have been developed to explain this phenomenon. It is not the purpose of this paper to survey these: reviews can be found in [1–4], with more recent work including [5–10]. Instead, we focus on providing empirical data useful for differentiating them.
These models are in some ways now very mathematically satisfying in that they can explain high Fibonacci numbers based on a small number of plausible assumptions, though they are not so satisfying to experimental scientists [11]. Despite an increasingly detailed molecular and biophysical understanding of plant organ positioning [12–14], the very parsimony and generality of the mathematical explanations make the generation and testing of experimental hypotheses difficult. There remains debate about the appropriate choice of mathematical models, and whether they need to be central to our understanding of the molecular developmental biology of the plant. While sunflowers provide easily the largest Fibonacci numbers in phyllotaxis, and thus, one might expect, some of the stronger constraints on any theory, there is a surprising lack of systematic data to support the debate. There have been only two large empirical studies of spirals in the capitulum, or head, of the sunflower: Weisse [15] and Schoute [16], which together counted 459 heads; Schoute found numbers from the main Fibonacci sequence 82% of the time and Weise 95%. The original motivation of this study was to add a third replication to these two historical studies of a widely discussed phenomenon. Much more recently, a study of a smaller sample of 21 seedheads was carried out by Couder [17], who specifically searched for non-Fibonacci examples, whereas Ryan et al. [18] studied the arrangement of seeds more closely in a small sample of Helianthus annuus and a sample of 33 of the related perennial H. tuberosus.
Neither the occurrence of Fibonacci structure nor the developmental biology leading to it are at all unique to sunflowers. As common in other species, the previous sunflower studies found not only Fibonacci numbers, but also the occasional occurrence of the double Fibonacci numbers, Lucas numbers and F4 numbers defined below [1]. It is worth pointing out the warning of Cooke [19] that numbers from these sequences make up all but three of the first 17 integers. This means that it is particularly valuable to look at specimens with large parastichy numbers, such as the sunflowers, where the prevalence of Fibonacci structure is at its most striking.
Neither Schoute nor Weisse reported their precise technique for assigning parastichy numbers to their samples, and it is noteworthy that neither author reported any observation of non-Fibonacci structure. One of the objectives of this study was to rigorously define Fibonacci structure in advance and to ensure that the assignment method, though inevitably subjective, was carefully documented.
This paper concentrates on the patterning of seeds towards the outer rim of sunflower seedheads. The number of ray florets (the ‘petals’, typically bright yellow) or the green bracts behind them tends to have a looser distribution around a Fibonacci number. In the only mass survey of these, Majumder & Chakravarti [20] counted ray florets on 1002 sunflower heads and found a distribution centred on 21.
This citizen science study evaluates the occurrence of Fibonacci structure in the spirals of sunflower (Helianthus annuus) seedheads. This phenomenon has competing biomathematical explanations, and our core premise is that observation of both Fibonacci and non-Fibonacci structure is informative for challenging such models. We collected data on 657 sunflowers. In our most reliable data subset, we evaluated 768 clockwise or anticlockwise parastichy numbers of which 565 were Fibonacci numbers, and a further 67 had Fibonacci structure of a predefined type. We also found more complex Fibonacci structures not previously reported in sunflowers. This is the third, and largest, study in the literature, although the first with explicit and independently checkable inclusion and analysis criteria and fully accessible data. This study systematically reports for the first time, to the best of our knowledge, seedheads without Fibonacci structure. Some of these are approximately Fibonacci, and we found in particular that parastichy numbers equal to one less than a Fibonacci number were present significantly more often than those one more than a Fibonacci number. An unexpected further result of this study was the existence of quasi-regular heads, in which no parastichy number could be definitively assigned.
Incorporation of irregularity into the mathematical models of phyllotaxis is relatively recent: [17] gave an example of a disordered pattern arising directly from the deterministic model while more recently the authors have begun to consider the effects of stochasticity [10,21]. Differentiating between these models will require data that go beyond capturing the relative prevalence of different types of Fibonacci structure, so this study was also designed to yield the first large-scale sample of disorder in the head of the sunflower.
The Fibonacci sequence is the sequence of integers 1,2,3,5,8,13,21,34,55,89,144… in which each member after the second is the sum of the two preceding. The Lucas sequence is the sequence of integers 1,3,4,7,11,18,29,47,76,123… obeying the same rule but with a different starting condition; the F4 sequence is similarly 1,4,5,9,14,23,37,60,97,…. The double Fibonacci sequence 2,4,6,10,16,26,42,68,110,… is double the Fibonacci sequence. We say that a parastichy number which is any of these numbers has Fibonacci structure. The sequencesF5=1,5,6,11,17,28,45,73,… and F8=1,8,9,17,26,43,69,112… also arise from the same rule, but as they had not been previously observed in sunflowers we did not include these in the pre-planned definition of Fibonacci structure for parsimony. One example of adjacent pairs from each of these sequences was, in fact, observed but both examples are classified as non-Fibonacci below. A parastichy number which is any of 12,20,33,54,88,143 is also not classed as having Fibonacci structure but is distinguished as a Fibonacci number minus one in some of the analyses, and similarly 14,22,35,56,90,145 as Fibonacci plus one.
When looking at a seedhead such as in figure 1 the eye naturally picks out at least one family of parastichies or spirals: in this case, there is a clockwise family highlighted in blue in the image on the right-hand side.
Figure 5 plots the individual pairs observed. On the reference line, the ratio of the numbers is equal to the golden ratio so departures from the line mark departures from Fibonacci structure, which are less evident in the more reliable photoreviewed dataset. It can be seen from table 3 that Fibonacci pairings dominate the dataset.
Observed pairings of Fibonacci types of clockwise and anticlockwise parastichy numbers. Other means any parastichy number which neither has Fibonacci structure nor is Fibonacci ±1. Of all the Fibonacci ±1/Fibonacci pairs, only sample 191, a (21,20) pair, was not close to an adjacent Fibonacci pair.
One typical example of a Fibonacci pair is shown in figure 6, with a double Fibonacci case infigure 1 and a Lucas one in figure 7. There was no photoreviewed example of an F4 pairing. The sole photoreviewed assignment of a parastichy number to the F4 sequence was the anticlockwise parastichy number 37 in sample 570, which was relatively disordered. The clockwise parastichy number was 55, lending support to the idea this may have been a perturbation of a (34,55) pattern. We also found adjacent members of higher-order Fibonacci series. Figures 8 and 9 each show well-ordered examples with parastichy counts found adjacent in the F5 and F8 series, respectively: neither of these have been previously reported in the sunflower.
Sunflower 095. An (89,55) example with 89 clockwise parastichies and 55 anticlockwise ones, extending right to the rim of the head. Because these are clear and unambiguous, the other parastichy families which are visible towards the centre are not counted here.
Sunflower 667. Anticlockwise parastichies only, showing competing parastichy families which are distinct but in some places overlapping.
Our core results are twofold. First, and unsurprisingly, Fibonacci numbers, and Fibonacci structure more generally, are commonly found in the patterns in the seedheads of sunflowers. Given the extent to which Fibonacci patterns have attracted pseudo-scientific attention [33], this substantial replication of limited previous studies needs no apology. We have also published, for the first time, examples of seedheads related to the F5 and F8 sequences but by themselves they do not add much to the evidence base. Our second core result, though, is a systematic survey of cases where Fibonacci structure, defined strictly or loosely, did not appear. Although not common, such cases do exist and should shed light on the underlying developmental mechanisms. This paper does not attempt to shed that light, but we highlight the observations that any convincing model should explain. First, the prevalence of Lucas numbers is higher than those of double Fibonacci numbers in all three large datasets in the literature, including ours, and there are sporadic appearances of F4, F5 and F8 sequences. Second, counts near to but not exactly equal to Fibonacci structure are also observable: we saw a parastichy count of 54 more often than the most common Lucas count of 47. Sometimes, ambiguity arises in the counting process as to whether an exact Fibonacci-structured number might be obtained instead, but there are sufficiently many unambiguous cases to be confident this is a genuine phenomenon. Third, among these approximately Fibonacci counts, those which are a Fibonacci number minus one are significantly more likely to be seen than a Fibonacci number plus one. Fourth, it is not uncommon for the parastichy families in a seedhead to have strong departures from rotational symmetry: this can have the effect of yielding parastichy numbers which have large departures from Fibonacci structure or which are completely uncountable. This is related to the appearance of competing parastichy families. Fifth, it is common for the parastichy count in one direction to be more orderly and less ambiguous than that in the other. Sixth, seedheads sometimes possess completely disordered regions which make the assignment of parastichy numbers impossible. Some of these observations are unsurprising, some can be challenged by different counting protocols, and some are likely to be easily explained by the mathematical properties of deformed lattices, but taken together they pose a challenge for further research.
It is in the nature of this crowd-sourced experiment with multiple data sources that it is much easier to show variability than it is to find correlates of that variability. We tried a number of cofactor analyses that found no significant effect of geography, growing conditions or seed type but if they do influence Fibonacci structure, they are likely to be much easier to detect in a single-experimenter setting.
We have been forced by our results to extend classifications of seedhead patterns beyond structured Fibonacci to approximate Fibonacci ones. Clearly, the more loose the definition of approximate Fibonacci, the easier it is to explain away departures from model predictions. Couder [17] found one case of a (54,87) pair that he interpreted as a triple Lucas pair 3×(18,29). While mathematically true, in the light of our data, it might be more compellingly be thought of as close to a (55,89) ideal than an exact triple Lucas one. Taken together, this need to accommodate non-exact patterns, the dominance of one less over one more than Fibonacci numbers, and the observation of overlapping parastichy families suggest that models that explicitly represent noisy developmental processes may be both necessary and testable for a full understanding of this fascinating phenomenon. In conclusion, this paper provides a testbed against which a new generation of mathematical models can and should be built.
UC Berkeley researchers have found a long-elusive Achilles’ heel within “triple-negative” breast tumors, a common type of breast cancer that is difficult to treat. The scientists then used a drug-like molecule to successfully target this vulnerability, killing cancer cells in the lab and shrinking tumors in mice.
“We were looking for targets that drive cancer metabolism in triple-negative breast cancer, and we found one that was very specific to this type of cancer,” said Daniel K. Nomura, an associate professor of chemistry and of nutritional sciences and toxicology at UC Berkeley and senior author for the study, which is published online ahead of print in Cell Chemical Biology.
Triple-negative breast cancers account for about one in five breast cancers, and they are deadlier than other forms of breast cancer, in part because no drugs have been developed to specifically target these tumors.
Triple-negative breast cancers do not rely on the hormones estrogen and progesterone for growth, nor on human epidermal growth factor receptor 2 (HER2). Because they do not depend on these three targets, they are not vulnerable to modern hormonal therapies or to the HER2-targeted drug Herceptin (trastuzumab).
Instead, oncologists treat triple-negative breast cancer with older chemotherapies that target all dividing cells. If triple-negative breast cancer spreads beyond the breast to distant sites within the body, an event called metastasis, there are few treatment options.
Tumor cells develop abnormal metabolism, which they rely on to get the energy boost they need to fuel their rapid growth. In their new study, the research team used an innovative approach to search for active enzymes that triple-negative breast cancers use differently for metabolism in comparison to other cells and even other tumors.
Inhibiting cancer metabolism
They discovered that cells from triple-negative breast cancer cells rely on vigorous activity by an enzyme called glutathione-S-transferase Pi1 (GSTP1). They showed that in cancer cells, GSTP1 regulates a type of metabolism called glycolysis, and that inhibition of GSTP1 impairs glycolytic metabolism in triple-negative cancer cells, starving them of energy, nutrients and signaling capability. Normal cells do not rely as much on this particular metabolic pathway to obtain usable chemical energy, but cells within many tumors heavily favor glycolysis.
Co-author Eranthie Weerapana, an associate professor of chemistry at Boston College, developed a molecule named LAS17 that tightly and irreversibly attaches to the target site on the GSTP1 molecule. By binding tightly to GSTP1, LAS17 inhibits activity of the enzyme. The researchers found that LAS17 was highly specific for GSTP1, and did not attach to other proteins in cells.
According to Nomura, LAS17 did not appear to have toxic side effects in mice, where it shrank tumors grown to an invasive stage from surgically transplanted, human, triple-negative breast cancer cells that had long been maintained in lab cultures.
The research team intends to continue studying LAS17, Nomura said, with the next step being to study tumor tissue resected from human triple-negative breast cancers and transplanted directly into mice.
“Inhibiting GSTP1 impairs glycolytic metabolism,” Nomura said. “More broadly, this inhibition starves triple-negative breast cancer cells, preventing them from making the macromolecules they need, including the lipids they need to make membranes and the nucleic acids they need to make DNA. It also prevents these cells from making enough ATP, the molecule that is the basic energy fuel for cells.”
Beyond the metabolic role they first sought to track down, GSTP1 also appears to aid signaling within triple-negative breast cancer cells, helping to spur tumor growth, the researchers found.
Technique identifies Achilles’ heels
Nomura said it was surprising that a single, unique target emerged from the research team’s search.
The method used by the researchers, called “reactivity-based chemoproteomics,” can quickly lead to specific targetable sites — the Achilles’ heels — on proteins of interest, and eventually to drug development strategies, Nomura said.
The approach is to search for protein targets that are actively functioning within cells, instead of first using the well-trod path of surveying all genes to identify the specific genes that have taken the first step toward protein production. With that more conventional strategy, the switching on, or “expression,” of genes is evidenced by the easily quantified molecule called messenger RNA, made by the cell from a gene’s DNA template.
Nomura’s team instead first used chemical probes that can react with certain configurations of two of the amino acid building blocks of protein — cysteine and lysine — known to be involved in several kinds of important structural and functional transitions that active proteins can undergo.
“A lot can happen after the first step in protein production, and we believe our method for identifying fully formed, active proteins is more useful for tracking down relevant differences in cellular physiology,” Nomura said.
The researchers analyzed and compared cells from five distinct triple-negative breast cancers that had been grown in cell cultures for generations, along with cells from four distinct breast cancers that were not triple negative.
The scientists used a chemical identification technique known as mass spectrometry to narrow down the set of proteins that had active lysines and cysteines to just those that were metabolic enzymes. Only then did they use the more conventional approach of measuring gene expression in the different cancer cell types.
GSTP1 was the only metabolically active enzyme that was specifically expressed only in triple-negative breast cancer cells compared to other breast cancer cell types, the researchers found. Separate analysis of databases of human breast cancer by UC San Francisco co-authors confirmed that GSTP1 is overexpressed in patients with triple-negative breast cancers in comparison to patients with other breast cancers.
In addition to Nomura and Weerapana, study authors included Sharon Louie, Elizabeth Grossman, Lucky Ding, Tucker Huffman and David Miyamoto, from UC Berkeley; Roman Camarda and Andrei Goga, from UC San Francisco, and Lisa Crawford, from Boston College. Study funders included the National Institutes of Health, the American Cancer Society, the U.S. Department of Defense, and the Searle Scholar Foundation.
Breast cancer metastasized to the liver (Courtesy of National Cancer Institute)
UC Berkeley researchers have found a long-elusive Achilles’ heel within “triple-negative” breast tumors, a common type of breast cancer that is difficult to treat. The scientists then used a drug-like molecule to successfully target this vulnerability, killing cancer cells in the lab and shrinking tumors in mice.
“We were looking for targets that drive cancer metabolism in triple-negative breast cancer, and we found one that was very specific to this type of cancer,” said Daniel K. Nomura, an associate professor of chemistry and of nutritional sciences and toxicology at UC Berkeley and senior author for the study, which is published online ahead of print on May 12 in Cell Chemical Biology.
Triple-negative breast cancers account for about one in five breast cancers, and they are deadlier than other forms of breast cancer, in part because no drugs have been developed to specifically target these tumors.
Triple-negative breast cancers do not rely on the hormones estrogen and progesterone for growth, nor on human epidermal growth factor receptor 2 (HER2). Because they do not depend on these three targets, they are not vulnerable to modern hormonal therapies or to the HER2-targeted drug Herceptin (trastuzumab).
Instead, oncologists treat triple-negative breast cancer with older chemotherapies that target all dividing cells. If triple-negative breast cancer spreads beyond the breast to distant sites within the body, an event called metastasis, there are few treatment options.
Tumor cells develop abnormal metabolism, which they rely on to get the energy boost they need to fuel their rapid growth. In their new study, the research team used an innovative approach to search for active enzymes that triple-negative breast cancers use differently for metabolism in comparison to other cells and even other tumors.
Inhibiting cancer metabolism
They discovered that cells from triple-negative breast cancer cells rely on vigorous activity by an enzyme called glutathione-S-transferase Pi1 (GSTP1). They showed that in cancer cells, GSTP1 regulates a type of metabolism called glycolysis, and that inhibition of GSTP1 impairs glycolytic metabolism in triple-negative cancer cells, starving them of energy, nutrients and signaling capability. Normal cells do not rely as much on this particular metabolic pathway to obtain usable chemical energy, but cells within many tumors heavily favor glycolysis.
•We used chemoproteomics to profile metabolic drivers of breast cancer
•GSTP1 is a novel triple-negative breast cancer-specific target
•GSTP1 inhibition impairs triple-negative breast cancer pathogenicity
•GSTP1 inhibition impairs GAPDH activity to affect metabolism and signaling
Breast cancers possess fundamentally altered metabolism that fuels their pathogenicity. While many metabolic drivers of breast cancers have been identified, the metabolic pathways that mediate breast cancer malignancy and poor prognosis are less well understood. Here, we used a reactivity-based chemoproteomic platform to profile metabolic enzymes that are enriched in breast cancer cell types linked to poor prognosis, including triple-negative breast cancer (TNBC) cells and breast cancer cells that have undergone an epithelial-mesenchymal transition-like state of heightened malignancy. We identified glutathione S-transferase Pi 1 (GSTP1) as a novel TNBC target that controls cancer pathogenicity by regulating glycolytic and lipid metabolism, energetics, and oncogenic signaling pathways through a protein interaction that activates glyceraldehyde-3-phosphate dehydrogenase activity. We show that genetic or pharmacological inactivation of GSTP1 impairs cell survival and tumorigenesis in TNBC cells. We put forth GSTP1 inhibitors as a novel therapeutic strategy for combatting TNBCs through impairing key cancer metabolism and signaling pathways.
Switching on genes, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 1: Next Generation Sequencing (NGS)
Switching on genes
Curator: Larry H. Bernstein, MD, FCAP
LPBI
UPDATED 3/17/2020
Gene Expression Controls Revealed
Researchers have modelled every atom in a key part of the process for switching on genes, revealing a whole new area for potential drug targets.
Proteins are essential for processes that sustain life. They are created in cells through a process called gene expression, which uses instructions from stretches of DNA called genes to build proteins. Sometimes genes are faulty and create proteins that contain errors, preventing the cell from functioning properly. These lead to genetic diseases like cystic fibrosis and haemophilia.
Gene expression is controlled by molecules called transcription factors, which bind to the start of a gene sequence at its ‘basal machinery’ and tell it to switch on and start creating certain proteins.
The way transcription factors bind to the basal machinery is a ‘fuzzy’ process, meaning the exact sequence of events is unknown because the steps do not exist for long enough to be captured by traditional imaging techniques.
But now, by creating a computer simulation of all of the tens of thousands of atoms making up the process and modelling their movements in 50 million separate steps, researchers at Imperial College London have been able to determine the sequence of events that lead to genes being switched on.
DISRUPTING DETRIMENTAL GENES
The simulated process revealed ‘pockets’ in the gene basal machinery, which the transcription factors move in and out of during binding. Knowing how these structures fit together could lead to the design of molecules that interfere with or disrupt the process, potentially tackling diseases.
Lead researcher Dr Robert Weinzierl from Imperial’s Department of Life Sciences said: “For the first time, we can fill in the dynamic landscape of interaction between transcription factors and basal machinery. This is a central mechanism for gene expression – the interactions here determine whether a gene gets switched on and creates proteins.”
“Gene regulation is a completely new drug target that has previously been too challenging to explore,” added Dr Weinzierl. “This process influences biology on a really fundamental level, and could allow us to prevent the expression of detrimental genes.”
FASTER DRUG SCREENING
The researchers’ new technique predicts the movements of all the atoms in order to build up a picture of the structures involved changing every couple of femtoseconds – quadrillionths of a second. The results of the first trial of the technique are reported today in PLOS Computational Biology.
Dr Weinzierl has submitted a patent application for his computer-based approach to studying gene expression interactions. Using this, compounds could be screened for possible fit into the basal machinery pockets.
“With computer simulation, it becomes easy to identify candidate compounds that could target these interactions without the need to test them first in real life, cutting down the time required to sift for new drugs,” said Dr Weinzierl.
Steps that lead to genes being switched on revealed in atomic simulation
Researchers have modelled every atom in a key part of the process for switching on genes, revealing a whole new area for potential drug targets.
Proteins are essential for processes that sustain life. They are created in cells through a process called gene expression, which uses instructions from stretches of DNA called genes to build proteins. Sometimes genes are faulty and create proteins that contain errors, preventing the cell from functioning properly. These lead to genetic diseases like cystic fibrosis and haemophilia.
Gene regulation is a completely new drug target that has previously been too challenging to explore.
Gene expression is controlled by molecules called transcription factors, which bind to the start of a gene sequence at its ‘basal machinery’ and tell it to switch on and start creating certain proteins.
The way transcription factors bind to the basal machinery is a ‘fuzzy’ process, meaning the exact sequence of events is unknown because the steps do not exist for long enough to be captured by traditional imaging techniques.
But now, by creating a computer simulation of all of the tens of thousands of atoms making up the process and modelling their movements in 50 million separate steps, researchers at Imperial College London have been able to determine the sequence of events that lead to genes being switched on.
Transcriptional activation domains (ADs) are generally thought to be intrinsically unstructured, but capable of adopting limited secondary structure upon interaction with a coactivator surface. The indeterminate nature of this interface made it hitherto difficult to study structure/function relationships of such contacts. Here we used atomistic accelerated molecular dynamics (aMD) simulations to study the conformational changes of the GCN4 AD and variants thereof, either free in solution, or bound to the GAL11 coactivator surface. We show that the AD-coactivator interactions are highly dynamic while obeying distinct rules. The data provide insights into the constant and variable aspects of orientation of ADs relative to the coactivator, changes in secondary structure and energetic contributions stabilizing the various conformers at different time points. We also demonstrate that a prediction of α-helical propensity correlates directly with the experimentally measured transactivation potential of a large set of mutagenized ADs. The link between α-helical propensity and the stimulatory activity of ADs has fundamental practical and theoretical implications concerning the recruitment of ADs to coactivators.Author Summary
The regulated transcription of eukaryotic genes is governed by gene-specific transcription factors that contain activation domains to stimulate the expression of nearby genes. Activation domains are unable to take up a defined three-dimensional conformation. Nevertheless, as we demonstrate in our study, molecular dynamics simulations reveal that the key docking point of such domains (centered around several large hydrophobic amino acid sidechains) folds into fluctuating α-helical conformations. Analysis of published data shows that this tendency of adopting such local structures correlates directly with stimulation activity. We also investigate the interaction of these structurally unstable domains with a coactivator interaction partner. Computational simulations are ideally suited for analysing the rapidly changing, “fuzzy” interactions occurring between these protein partners. We gained new insights into the competitive nature of the key hydrophobic sidechains in binding to a pocket on the coactivator surface and documented for the first time the rapidly changing movements of an activation domain during these interactions.
The last decade has seen an incredible breakthrough in technologies that allow histones, transcription factors (TFs), and RNA polymerases to be precisely mapped throughout the genome. From this research, it is clear that there is a complex interaction between the chromatin landscape and the general transcriptional machinery and that the dynamic control of this interface is central to gene regulation. However, the chromatin remodeling enzymes and general TFs cannot, on their own, recognize and stably bind to promoter or enhancer regions. Rather, they are recruited to cis regulatory regions through interaction with site-specific DNA binding TFs and/or proteins that recognize epigenetic marks such as methylated cytosines or specifically modified amino acids in histones. These “recruitment” factors are modular in structure, reflecting their ability to interact with the genome via one region of the protein and to simultaneously bind to other regulatory proteins via “effector” domains. In this chapter, we provide examples of common effector domains that can function in transcriptional regulation via their ability to (a) interact with the basal transcriptional machinery and general co-activators, (b) interact with other TFs to allow cooperative binding, and (c) directly or indirectly recruit histone and chromatin modifying enzymes.
Transcriptional activation is a stepwise process that requires (a) creating and maintaining an open chromatin structure, (b) assembly of the preinitiation complex, and (c) transition to productive elongation (Fig. 12.1). Successful completion of each of these steps involves a diverse group of proteins, some of which function in a relatively promoter-specific manner whereas others regulate large sets of genes. Recent advances in molecular and computational biology allow histone and DNA modifications, TFs, and RNA polymerases to be precisely mapped throughout the genome, relative to active or silent promoters (see [1–3] for reviews). From this research, it is becoming clear that there is a complex interaction between the chromatin landscape and the transcriptional machinery and that the dynamic relationship of this interface is central to biological control over gene expression [4]. It is now recognized that regulatory factors can exert their influence on transcriptional activation either via co-localization with other proteins that are bound at or near core promoter regions or they can be recruited to distal enhancer regions and interact with promoter-bound proteins via looping mechanisms. However, generally speaking, the chromatin remodeling enzymes and the general transcription factors involved in initiation and elongation cannot, on their own, recognize and stably bind to the promoter or enhancer regions.
Regulation of transcription. Shown is a schematic representing the three steps needed for productive transcription, including Step 1: the creation of open chromatin, which involves interactions between DNA-bound proteins and histone modifying enzymes …
One way in which chromatin remodeling enzymes and general transcription factors are recruited to cis-regulatory regions is through interaction with site-specific DNA binding TFs (Fig. 12.2a). The three largest classes of site-specific DNA binding proteins in mammals contact the genome via conserved DNA binding domains called zinc fingers, homeodomains, and helix–loop–helix domains [5] (Chapter 3 of this volume provides a catalog of eukaryotic DNA binding domains, and Chapters 4 and 5 specifically review C2H2 zinc fingers and homeodomains). Each of these classes of site-specific DNA binding factors contains many different proteins; for example, in humans there are over 650 zinc finger proteins, ~ 250 homeodomain proteins, and ~80 helix-loop-helix proteins [5]. Within each class, individual TFs can bind to and regulate hundreds to thousands of different genes. Site-specific TFs are modular in their structure reflecting their ability to bind to DNA via their DNA binding domains and simultaneously bind to other transcriptional regulatory proteins via so-called effector domains. The modular nature of site-specific TFs has been repeatedly demonstrated using in vitro and in vivo reporter assays. In these experiments, effector domains are separated from their natural DNA binding domains and then engineered to be part of a fusion protein having a heterologous DNA binding domain. Numerous studies have shown that simply bringing such effector domains to promoter regions can modulate transcription [6–8].
Modular structure of effector domain-containing proteins. Effector domains can be recruited to specific genomic regions via a DNA binding domains that recognize short DNA sequence motifs (TFBS), b recognition of a methylated cytosine (shown as a black…
Another way in which chromatin remodeling enzymes and general transcription factors can be brought to the genome is via effector domains that reside in proteins that can recognize epigenomic marks. Similar to recognition of a short nucleotide motif by a DNA binding protein, other proteins can distinguish distinctively modified DNA and histone protein “motifs”. For example, methylated cytosine in the 5′-CpG-3′ dinucleotide sequence is specifically recognized by members of a family of proteins containing a conserved methyl-CpG binding domain (MBD). MBD-containing proteins, which include MeCP2, MBD1, MBD2 and MBD4, bind specifically to methyl-CpG motifs located throughout the genome [9]; see Fig. 12.2b. MBD-containing proteins function by recruiting various co-regulators to methyl-CpG sites. For example, MeCP2 simultaneously binds promoter regions containing methyl-CpG motifs and the Sin3-containing histone deacetylase complex via a transcriptional repression domain (TRD), resulting in histone deacetylation and transcriptional silencing [10, 11]. Likewise, MBD1 and MBD2 copurify with distinct cellular complexes which link DNA methylation with chromatin modification and transcriptional repression. Similarly, posttranslational modifications of the amino termini of core histones are correlated to transcriptional states and are recognized by relevant chromatin-associated proteins (Fig. 12.2c). Several different histone modifications have been identified, including acetylation, phosphorylation, and methylation, and specific protein domains have evolved to recognize several of these different modifications. For example, different methylation states of histone H3 at lysine 4 can be recognized by tudor, chromo, and plant homeodomains (PHD), by malignant brain tumor (MBT) domains, and by WD40 repeat domains (many of these domains are structurally related and are collectively referred to as the “royal family” [12], reviewed [13, 14]). Other examples of this family include the chromodomain of HP1, which interacts with lower (mono- and di-) methylation states of lysine 9 of histone H3 but preferentially binds to the trimethylated state [15, 16] and the tudor domain of 53BP1, which can discriminate between the diand tri-methyl state of H4K20, preferring the dimethyl form [17, 18]. Acetylated lysine is also recognized by specific protein modules called the bromodomain [19], which is found in many chromatin-associated proteins and in nearly all known nuclear histone acetyltransferases (HATs). Of course, epigenetic marks such as DNA methylation and histone modifications are located at specific genomic regions (which can vary in different cell types), indicating that DNA methylases and histone modifying enzymes must be recruited to the genome by sequence-specific mechanisms such as site-specific TFs or RNAs. For example, KRAB-ZNFs can recruit the KAP1/SETDB1 histone methylating complex and long non-coding RNAs can recruit the PRC2 histone methylation complex [20–23].
The focus of this chapter is on the effector domains that are brought to specific sites of the genome by DNA binding proteins, methyl-CpG binding proteins, or histone binding proteins. (The interaction of TFs with chromatin more generally is discussed in Chapter 11). We provide examples of common effector domains that can function in transcriptional regulation via their ability to influence each of the steps outlined in Fig. 12.1. Specifically, we discuss effector domains that can: (a) interact with the basal transcriptional machinery and general co-activators, (b) interact with other TFs to allow cooperative binding, and (c) directly or indirectly recruit histone and chromatin modifying enzymes.
Eukaryotic transcriptional dynamics: from single molecules to cell populations
Transcriptional regulation is achieved through combinatorial interactions between regulatory elements in the human genome and a vast range of factors that modulate the recruitment and activity of RNA polymerase. Experimental approaches for studying transcription in vivo now extend from single-molecule techniques to genome-wide measurements. Parallel to these developments is the need for testable quantitative and predictive models for understanding gene regulation. These conceptual models must also provide insight into the dynamics of transcription and the variability that is observed at the single-cell level. In this Review, we discuss recent results on transcriptional regulation and also the models those results engender. We show how a non-equilibrium description informs our view of transcription by explicitly considering time-and energy-dependence at the molecular level.
Transcriptional regulation in the nucleus is the culmination of the actions of a diverse range of factors, such as transcription factors, chromatin remodellers, polymerases, helicases, topoisomerases, kinases, chaperones, proteasomes, acetyltransferases, deacetylases and methyltransferases. Determining how these molecules work in concert in the eukaryotic nucleus to regulate genes remains a central challenge in molecular biology. Dynamics lie at the heart of this mystery. Megadalton complexes assemble and disassemble on genes within seconds1,2; nucleosome turnover ranges from minutes to hours3; and gene activity demonstrates complex temporal patterns such as oscillation and transcriptional bursting4,5. Exciting new experimental advances have enabled the study of dynamic transcriptional regulation at the single-molecule6 and genome-wide7levels, thus enhancing our understanding of transcriptional regulation in vivo. These approaches also necessitate new models for describing gene expression. In this Review, we discuss recent in vivo results and the quantitative models that are motivated by those results.
Chromatin immunoprecipitation (ChIP) provides genome-wide occupancy profiles for chromatin-interacting factors at near base-pair resolution in populations of cells8,9. Using this approach on a genome-wide level has generated comprehensive maps of regulation on a gene-by-gene basis7,8,10. This population approach has been complemented by single-cell imaging techniques. Almost all factors that have been studied by live-cell microscopy exhibit dwell times on chromatin on the order of seconds11, and single-cell studies demonstrate a great variability in gene expression among cells in a population, owing in part to the stochastic nature of transcription12. Despite these tremendous advances in understanding the behaviour of individual factors, both methods fall short of capturing the sequence of events that is required to activate or repress a gene in vivo. Ideally, the occupancy of many factors that are coincident on a single stretch of DNA would be measured to obtain a sense of the complexes and intermediates that assemble in vivo. However, this experimental challenge is a daunting one. Current re-ChIP (also known as sequential ChIP) experiments usually look at two factors4,13 but it would be necessary to look at an order of magnitude more factors to begin to capture the combinatorial complexity of transcriptional regulation in metazoans4,14–16.
The gulf between actual mechanisms of transcriptional regulation and experimental capabilities could be bridged by using quantitative models of transcription. Decades of biochemical, structural and genetic data have spawned multiple models of transcriptional regulation, several of which we discuss below (FIG. 1). Even though these views are not mutually exclusive and boundaries between them are not clear, they reflect fundamental differences regarding the mechanisms of the underlying molecular processes. Currently, most quantitative theoretical models describe transcriptional regulation as an equilibrium thermodynamic phenomenon — an assumption that allows model building without explicitly considering the dynamics. Here we explain how this description is fundamentally inconsistent with the canonical view of gene regulation based on a sequential, ordered recruitment of factors, which is an example of a non-equilibrium model. In the context of a non-equilibrium model, the transcriptional dynamics can exhibit a form of molecular memory so that the future behaviour of the system depends on its history. We will outline this gap between the molecular biologist’s canonical view of transcription and the quantitative approaches that are often used to describe it. We argue for a non-equilibrium view of transcriptional regulation that is informed and constrained by single-cell observations. With the ability to observe single transcription factors17 and single transcribing genes18 in living cells, new experimental and modelling possibilities are emerging for understanding transcription dynamics in vivo.
A model of how twist defects, stimulated by the remodeler ATPase, shift DNA around the nucleosome . From the Science article G.D. Bowman and S. Deindl. Remodeling the Genome. Science (2020); 166(6461:p35-36.
In complex organisms such as humans, a single genetic blueprint can give rise to a multitude of different cell types, from nerve to liver to muscle. Such cellular diversity relies on restricting which portions of genomic DNA are accessible and therefore can be read by cellular machinery. Ultimately, access to DNA depends on placement of a repetitive, spool-like structure called the nucleosome, the basic packaging unit of chromosomes. The nucleosome occludes two tight loops of DNA and thus represents a fundamentally repressive element. When and where nucleosomes are positioned can affect complex transcriptional programs, and therefore disruptions in the factors responsible for nucleosome positioning often result in cancers and multisystem developmental diseases. Although the mechanism of shifting nucleosomes along DNA has long proved elusive, a recent flurry of structural, biophysical, and biochemical work has revealed a core mechanistic framework explaining how nucleosomes are actively repositioned throughout the genome.
Nucleosomes are the most ubiquitous protein-DNA complexes in all eukaryotic cells. The core of each nucleosome is a symmetric, disk-like structure made of histone proteins that provides a scaffold around which two loops of the DNA helix are snugly wrapped (1). Histones are often modified through, for example, acetylation, methylation, and phosphorylation, which add an additional layer of information on top of the genetic code. This epigenetic information demarcates functionally distinct regions of the genome—for instance, whether a gene is active or designated to remain silent—for each cell type.
Owing to their extensive protein-DNA interface, nucleosomes are relatively stable structures. Active placement and reorganization of nucleosomes depend on chromatin remodelers. As the gatekeepers of nucleosome packaging, these enzymes participate both in activating and repressing gene expression. Remodelers can assemble, disassemble, and exchange histones within the nucleosome, as well as shift the position of the histone core along DNA. Acting on either face of the nucleosome disk, remodelers can move the histone core back and forth on DNA, changing which parts of DNA are exposed and which are wrapped up in the nucleosome. Increased exposure of DNA occurs when remodelers shift adjacent nucleosomes into each other, resulting in histone ejection (2).
The ability of remodelers to manipulate nucleosome structure stems from a highly conserved adenosine triphosphatase (ATPase) motor that belongs to a larger superfamily of helicase-like ATPases called superfamily 2 (SF2).
Consistent with an inchworm-type translocation mechanism, nucleosomal DNA is shifted by remodelers with an elementary step of a single base pair (4). Given the spiral structure of the duplex, nucleosomal DNA must shift around the histone core in a corkscrew fashion. Remodeler ATPases remain in a fixed location on the histone core during DNA translocation, which means that DNA all around the nucleosome must shift in response to localized action at the ATPase binding site. When poised for hydrolysis, the ATPase in its closed state eliminates this bulge, corkscrewing DNA toward the nucleosome midpoint (dyad) on the other side. This creation and elimination of a DNA bulge is equivalent to altering DNA twist.
These snapshots of remodelers altering DNA twist are complemented by the observation of active movement of nucleosomal DNA by remodelers during three-color single-molecule FRET (Förster resonance energy transfer) experiments, where DNA movements were coupled to ATP hydrolysis (13).
The authors suggest some questions that might direct future research. For example, in addition to DNA geometry and energetics, to what extent is twist diffusion dependent on other characteristics of the nucleosome?
Histone proteins come in a variety of flavors, and an important goal is identifying whether distinct biophysical properties of histone variants and other epigenetic signatures can determine or bias the outcomes of remodeling reactions.
Massachusetts General Hospital (MGH) investigators have identified the first potential molecular treatment target for the most common form of pancreatic cancer, which kills more than 90 percent of patients. Along with finding that the tumor suppressor protein SIRT6 is inactive in around 30 percent of cases of pancreatic ductal adenocarcinoma (PDAC), the team identified the precise pathway by which SIRT6 suppresses PDAC development, a mechanism different from the way it suppresses colorectal cancer. The paper will appear in the June 2 issue of Cell and have been published online.
“With the advance of cancer genomics, it has become evident that alterations in epigenetic factors – those that control whether and when other genes are expressed – represent some of the most frequent alterations in cancer,” says Raul Mostoslavsky, MD, PhD, of the MGH Cancer Center, senior author of the report. “Yet, not many of those factors have been described before, and those that have been identified have not been linked to specific downstream targets. Not only did more than a third of analyzed PDAC patient samples exhibit the molecular signature we identified, those patients also turned out to have very poor prognoses.”
Among its other functions, SIRT6 is known to control how cells process glucose, and a 2012 study by Mostoslavsky’s team found that its ability to suppress colorectal cancer involves control of a process called glycolysis. But while that study also found reduced SIRT6 expression in PDAC tumor cells, the current investigation indicated that SIRT6 deficiency promotes PDAC through a different mechanism. Experiments in cell lines and animal models revealed that low SIRT6 levels in PDAC were correlated with increased expression of Lin28b, an oncoprotein normally expressed during fetal development.
Lin28b expression proved to be essential to the growth and survival of SIRT6-deficient PDAC cells and acted by preventing a family of tumor-suppressing mRNAs called let-7 from blocking expression of three genes previously associated with increased aggressiveness and metastasis in pancreatic cancers. All of these hallmarks – reduced SIRT6, increased Lin28b and reduced let-7 expression – were found in tumor samples from patients who died more quickly.
“A general message from these studies is that cancer cells benefit from modulating epigenetic factors like SIRT6 by acquiring the ability to override normal cellular growth control patterns,” says Mostoslavsky, an associate professor of Medicine at Harvard Medical School and an associate member at the Broad Institute. “Each tumor type may acquire a unique set of capabilities that may provide tumor-specific growth and survival advantages, which may need to be determined for each kind of cancer. In terms of our findings regarding PDAC, we are intrigued by the downstream pathways controlled by Lin28b and how they increase aggressiveness and metastasis, and we are hopeful that developing in the future Lin28b inhibitors could benefit this subset of PDAC patients, who currently have very few treatment options.”
SIRT6 Suppresses Pancreatic Cancer through Control of Lin28b
Sita Kugel, Carlos Sebastián, Julien Fitamant,…., Alon Goren, Vikram Deshpande, Nabeel Bardeesy, Raul Mostoslavsky
•Loss of SIRT6 cooperates with oncogenic Kras to drive pancreatic cancer
•SIRT6 regulates the oncofetal protein Lin28b through promoter histone deacetylation
•Lin28b drives the growth and survival of SIRT6-deficient pancreatic cancer
•SIRT6 and Lin28b expression define prognosis in specific pancreatic cancer subsets
Chromatin remodeling proteins are frequently dysregulated in human cancer, yet little is known about how they control tumorigenesis. Here, we uncover an epigenetic program mediated by the NAD+-dependent histone deacetylase Sirtuin 6 (SIRT6) that is critical for suppression of pancreatic ductal adenocarcinoma (PDAC), one of the most lethal malignancies. SIRT6 inactivation accelerates PDAC progression and metastasis via upregulation of Lin28b, a negative regulator of the let-7 microRNA. SIRT6 loss results in histone hyperacetylation at theLin28b promoter, Myc recruitment, and pronounced induction of Lin28b and downstream let-7 target genes, HMGA2, IGF2BP1, and IGF2BP3. This epigenetic program defines a distinct subset with a poor prognosis, representing 30%–40% of human PDAC, characterized by reduced SIRT6 expression and an exquisite dependence on Lin28b for tumor growth. Thus, we identify SIRT6 as an important PDAC tumor suppressor and uncover the Lin28b pathway as a potential therapeutic target in a molecularly defined PDAC subset.
The sirtuins (SIRTs; of which there are seven in mammals) are NAD+-dependent enzymes that regulate a large number of cellular pathways and forestall the progression of ageing and age-associated diseases. In recent years, the role of sirtuins in cancer biology has become increasingly apparent, and growing evidence demonstrates that sirtuins regulate many processes that go awry in cancer cells, such as cellular metabolism, the regulation of chromatin structure and the maintenance of genomic stability. In this article, we review recent advances in our understanding of how sirtuins affect cancer metabolism, DNA repair and the tumour microenvironment and how activating or inhibiting sirtuins may be important in preventing or treating cancer.
Figure 1: Overview of the role of sirtuins in the regulation of cancer metabolism
The inhibitory effects of sirtuin 3 (SIRT3), SIRT4 and SIRT6 on metabolic pathways that drive cancer cells are depicted. In normal cells, SIRT6 functions as a co-repressor for the transcription factors hypoxia-inducible factor 1α (HIF1α…
Tumor suppressor p53 cooperates with SIRT6 to regulate gluconeogenesis by promoting FoxO1 nuclear exclusion
In mammalian cells, tumor suppressor p53 plays critical roles in the regulation of glucose metabolism, including glycolysis and oxidative phosphorylation, but whether and how p53 also regulates gluconeogenesis is less clear. Here, we report that p53 efficiently down-regulates the expression of phosphoenolpyruvate carboxykinase (PCK1) and glucose-6-phosphatase (G6PC), which encode rate-limiting enzymes in gluconeogenesis. Cell-based assays demonstrate the p53-dependent nuclear exclusion of forkhead box protein O1 (FoxO1), a key transcription factor that mediates activation of PCK1 and G6PC, with consequent alleviation of FoxO1- dependent gluconeogenesis. Further mechanistic studies show that p53 directly activates expression of the NAD+-dependent histone deacetylase sirtuin 6 (SIRT6), whose interaction with FoxO1 leads to FoxO1 deacetylation and export to the cytoplasm. In support of these observations, p53-mediated FoxO1 nuclear exclusion, down-regulation of PCK1 and G6PC expression, and regulation of glucose levels were confirmed in C57BL/J6 mice and in liver-specific Sirt6 conditional knockout mice. Our results provide insights into mechanisms of metabolism-related p53 functions that may be relevant to tumor suppression.
As the “guardian of the genome,” tumor suppressor p53 has been reported to coordinate diverse cellular responses to a broad range of environment stresses (1) and to play antineoplastic roles by activating downstream target genes involved in DNA damage repair, apoptosis, and cell-cycle arrest (2). Recent studies have indicated broader roles for p53 in mediating metabolic changes in cells under various physiological and pathological conditions (3–7). For example, p53 was reported to influence the balance between glycolysis and oxidative phosphorylation by inducing the p53-induced glycolysis and apoptosis regulator (TIGAR) and by regulating the synthesis of cytochrome c oxidase 2 (SCO2) (3), respectively, thus promoting the switch from glycolysis to oxidative phosphorylation. p53 also may impede metabolism by reducing glucose import (4) or by inhibiting the pentose phosphate pathway (PPP) (5). More recently, context-dependent inhibitory (6) or stimulatory (7, 8) effects of p53 on gluconeogenesis have been reported. It thus is clear that p53 plays important roles in glucose regulation in mammalian cells. Glucose homeostasis is maintained by a delicate balance between intestinal absorption of sugar, gluconeogenesis, and the utilization of glucose by the peripheral tissues, irrespective of feeding or fasting (9). The gluconeogenesis pathway is catalyzed by several key enzymes that include the first and last rate-limiting enzymes of the process, phosphoenolpyruvate carboxykinase (PCK1) and glucose-6-phosphatase (G6PC), respectively. The expression of both PCK1 and G6PC is controlled mainly at the transcription level. For example, the transcription factor forkhead box protein O1 (FoxO1) activates gluconeogenesis through direct binding to the promoters of G6PC and PCK1 (10). A dominant negative FoxO1 lacking its transactivation domain significantly decreases gluconeogenesis (11) whereas FoxO1 ablation impairs fasting- and cAMP-induced PCK1 and G6PC expression (12). Therefore, factors influencing expression of FoxO1 or its binding activity to the PCK1 and G6PC promoters are potential targets for gluconeogenesis regulation. The transcription activity of FoxO family members is regulated by a sophisticated signaling network. Various environmental stimuli cause different posttranslational modifications of FoxO proteins, including phosphorylation, acetylation, ubiquitination, and methylation (13–15). The phosphorylation of FoxO proteins is known to be essential for their shuttling between the nucleus and cytoplasm. For example, kinase Akt/PKB phosphorylates FoxO1 at threonine 24 and at serines 256 and 319, which in turn leads to 14-3-3 binding and subsequent cytoplasmic sequestration. The acetylation of FoxO proteins also affects their trafficking and DNA-binding activities (15–17). Sirtuin (SIRT)1, a homolog of the yeast silent information regulator-2 (Sir2), has been identified as a deacetylase for FoxO proteins (15, 17, 18). Of the seven mammalian sirtuins, SIRT1, SIRT6, and SIRT7 are localized to the nucleus (19), and SIRT6 was recently reported to act as a central player in regulating the DNA damage response, glucose metabolism, and aging (20–26). Using a knockout mouse model, it was found that SIRT6 functions as a corepressor of the transcription factor Hif1α to suppress glucose uptake and glycolysis.
Significance: Beyond its canonical functions in processes such as cell-cycle arrest, apoptosis, and senescence, the tumor suppressor p53 has been increasingly implicated in metabolism. Here, in vitro and in vivo studies establish a role for p53 in gluconeogenesis through a previously unidentified mechanism involving (i) direct activation of the gene encoding the NAD-dependent deacetylase sirtuin 6 (SIRT6), (ii) SIRT6-dependent deacetylation and nuclear exclusion of forkhead box protein O1 (FoxO1), and (iii) downregulation of FoxO1-activated genes (G6PC and PCK1) that are rate-limiting for gluconeogenesis. These results have implications for proposed tumor-suppressor functions of p53 through regulation of metabolic pathways.
Among a variety of other functions, SIRT6 was previously connected to glucose metabolism. For example, SIRT6 acts as a corepressor of the transcription factor Hif1-α to suppress glycolysis (23). Conversely, the deletion of Sirt6 in mice results in severe hypoglycemia (33) whereas the liver-specific deletion of Sirt6 leads to increased glycolysis and triglyceride synthesis (23, 34). Our study adds further evidence that SIRT6 plays an important role in glucose metabolism by connecting p53 transcription activity and gluconeogenesis. Our data also reemphasize a previously established role for SIRT6 in regulating the acetylation state and nuclear localization of FoxO proteins, albeit in a divergent manner. Thus, the Caenorhabditis elegans SIRT6/7 homolog SIR-2.4 was implicated in DAF-16 deacetylation and consequent nuclear localization and function in stress responses (35); and the effect was reported to be indirect and to involve a stress-induced inhibition by SIR-2.4 of CBP-mediated acetylation of DAF-16 that is independent of its deacetylase activity. These results, emphasizing context-dependent SIRT6 mechanisms, contrast with the SIRT6 deacetylase activity requirement for FoxO1 nuclear exclusion in the present study and a likely direct effect of SIRT6 on FoxO1 deacetylation based on their direct interaction, the SIRT6 deacetylase activity requirement, and precedent (15, 17, 18) from direct SIRT1-mediated deacetylation of FoxO proteins.
Despite a high genetic diversity, cancer cells exhibit a common set of functional characteristics, one being the “Warburg effect”: i.e., continuous high glucose uptake and a higher rate of glycolysis than that in normal cells (36). To favor the rapid proliferation requirement for high ATP/ADP and ATP/AMP ratios, cancer cells use large amounts of glucose. p53, as one of the most important tumor suppressors, exerts its antineoplastic function through diverse pathways that include the regulation of glucose metabolism. Thus, p53 regulates glucose metabolism by activation of TIGAR (3), which lowers the intracellular concentrations of fructose-2,6-bisphosphate and decreases glycolysis. On the other hand, p53 activation causes down-regulation of several glycolysisrelated factors such as phosphoglycerate mutase (PGM) (37) and the glucose transporters (4). Expression of p53 also can limit the activity of IκBα and IκBβ, thereby restricting the activation of NFκB and dampening the expression of glycolysis-promoting genes such as GLUT3 (38). As a reverse glycolysis pathway, gluconeogenesis generates glucose from noncarbohydrate precursors and is conceivably essential for tumor cell growth. However, the current study further supports the notion (6) that p53 is also involved in a gluconeogenesis inhibition pathway, which in this case is executed by enhanced SIRT6 expression and subsequent FoxO1 nuclear exclusion. These results raise the interesting possibility that an inhibition of gluconeogenesis may contribute to the tumorsuppressive function of p53.
http://www.jcpjournal.org/journal/DOIx.php?id=10.15430/JCP.2013.18.3.221 Keywords : Oncogenes, Tumor suppressors, Glutamine metabolism, Cancer cells … p53 is a well-known protein which is involved in many cellular functionsincluding cell … deprivation activates p53 by regulating protein phosphatase 2A ( PP2A). …. and tumor suppressors may affect glutamine metabolism in cancercells
Protein controlling glucose metabolism also a tumor suppressor
Finding supports metabolic strategies to control tumor growth
A protein known to regulate how cells process glucose also appears to be a tumor suppressor, adding to the potential that therapies directed at cellular metabolism may help suppress tumor growth. In their report in the Dec. 7 issue of Cell, a multi-institutional research team describes finding that cells lacking the enzyme SIRT6, which controls how cells process glucose, quickly become cancerous. They also found evidence that uncontrolled glycolysis, a stage in normal glucose metabolism, may drive tumor formation in the absence of SIRT6 and that suppressing glycolysis can halt tumor formation.
“Our study provides solid evidence that SIRT6 may function as a tumor suppressor by regulating glycolytic metabolism in cancer cells,” says Raul Mostoslavsky, MD, PhD, of the Massachusetts General Hospital (MGH) Cancer Center, senior author of the report. “Critically, our findings indicate that, in tumors driven by low SIRT6 levels, drugs that may inhibit glycolysis – currently a hot research topic among biotechnology companies – could have therapeutic benefits.”
The hypothesis that a switch in the way cells process glucose could set off tumor formation was first proposed in the 1920s by German researcher Otto Warburg, who later received the Nobel Prize for discoveries in cellular respiration. He observed that, while glucose metabolism is normally a two-step process involving glycolysis in the cellular cytoplasm followed by cellular respiration in the mitochondria, in cancer cells rates of glycolysis are up to 200 times higher. Warburg’s proposition that this switch in glucose processing was a primary cause of cancer did not hold up, as subsequent research supported the role of mutations in oncogenes, which can spur tumor growth if overexpressed, and tumor suppressors, which keep cell proliferation under control. But recent studies have suggested that alterations in cellular metabolism may be part of the process through which activated oncogenes or inactivated tumor suppressors stimulate cancer formation.
A 2010 study led by Mostoslavsky found that the absence of SIRT6 – one of a family of proteins called sirtuins that regulate many important biological pathways – appears to “flip the switch” from normal glucose processing to the excess rates of glycolysis seen in cancer cells. The current study was specifically designed to investigate whether SIRT6’s control of glucose metabolism also suppresses tumor formation. The research team first showed that cultured skin cells from embryonic mice lacking SIRT6 proliferated rapidly and quickly formed tumors when injected into adult mice. They also confirmed elevated glycolysis levels in both cells lacking SIRT6 and tumor cells and found that formation of tumors through SIRT6 deficiency did not appear to involve oncogene activation.
Analysis of tumor samples from patients found reduced SIRT6 expression in many – particularly in colorectal and pancreatic tumors. Even among patients whose tumors appeared to be more aggressive, higher levels of SIRT6 expression may have delayed or, for some, prevented relapse. In a mouse model programmed to develop numerous colon polyps, the researchers showed that lack of intestinal SIRT6 expression tripled the formation of polyps, many of which became invasive tumors. Treating the animals with a glycolytic inhibitor significantly reduced tumor formation, even in the absence of SIRT6.
“Our results indicate that, at least in certain cancers, inhibiting glycolytic metabolism could provide a strong alternative way to halt cancer growth, possibly acting synergistically with current anti-tumor therapies,” says Mostoslavsky, an assistant professor of Medicine at Harvard Medical School. “Cancer metabolism has only recently emerged as a hallmark of tumorigenesis, and the field is rapidly expanding. With the current pace of research and the speed at which some basic discoveries are moving into translational studies, it is likely that drugs targeting cancer metabolism may be available to patients in the near future.”
THE HISTONE DEACETYLASE SIRT6 IS A NOVEL TUMOR SUPPRESSOR THAT CONTROLS CANCER METABOLISM
Reprogramming of cellular metabolism is a key event during tumorigenesis. Despite being known for decades (Warburg effect), the molecular mechanisms regulating this switch remained unexplored. Here, we identify SIRT6 as a novel tumor suppressor that regulates aerobic glycolysis in cancer cells. Importantly, loss of SIRT6 leads to tumor formation without activation of known oncogenes, while transformed SIRT6-deficient cells display increased glycolysis and tumor growth, suggesting that SIRT6 plays a role in both establishment and maintenance of cancer. Using a conditional SIRT6 allele, we show that SIRT6 deletion in vivoincreases the number, size and aggressiveness of tumors. SIRT6 also functions as a novel regulator of ribosome metabolism by co-repressing MYC transcriptional activity. Lastly, SIRT6 is selectively downregulated in several human cancers, and expression levels of SIRT6 predict prognosis and tumor-free survival rates, highlighting SIRT6 as a critical modulator of cancer metabolism. Our studies reveal SIRT6 to be a potent tumor suppressor acting to suppress cancer metabolism.
Cancer cells are characterized by the acquisition of several characteristics that enable them to become tumorigenic (Hanahan and Weinberg, 2000). Among them, the ability to sustain uncontrolled proliferation represents the most fundamental trait of cancer cells. This hyperproliferative state involves the deregulation of proliferative signaling pathways as well as loss of cell cycle regulation. In addition, tumor cells need to readjust their energy metabolism to fuel cell growth and division. This metabolic adaptation is directly regulated by many oncogenes and tumor suppressors, and is required to support the energetic and anabolic demands associated with cell growth and proliferation (Lunt and Vander Heiden, 2011).
Alteration in glucose metabolism is the best-known example of metabolic reprogramming in cancer cells. Under aerobic conditions, normal cells convert glucose to pyruvate through glycolysis, which enters the mitochondria to be further catabolized in the tricarboxylic acid cycle (TCA) to generate adenosine-5’-triphosphate (ATP). Under anaerobic conditions, mitochondrial respiration is abated; glucose metabolism is shifted towards glycolytic conversion of pyruvate into lactate. This metabolic reprogramming is also observed in cancer cells even in the presence of oxygen and was first described by Otto Warburg several decades ago (Warburg, 1956; Warburg et al., 1927). By switching their glucose metabolism towards “aerobic glycolysis”, cancer cells accumulate glycolytic intermediates that will be used as building blocks for macromolecular synthesis (Vander Heiden et al., 2009). Most cancer cells exhibit increased glucose uptake, which is due, in part, to the upregulation of glucose transporters, mainly GLUT1 (Yamamoto et al., 1990; Younes et al., 1996). Moreover, cancer cells display a high expression and activity of several glycolytic enzymes, including phospho-fructose kinase (PFK)-1, pyruvate kinase M2, lactate dehydrogenase (LDH)-A and pyruvate dehydrogenase kinase (PDK)-1 (Lunt and Vander Heiden, 2011), leading to the high rate of glucose catabolism and lactate production characteristic of these cells. Importantly, downregulation of either LDH-A or PDK1 decreases tumor growth (Bonnet et al., 2007; Fantin et al., 2006; Le et al., 2010) suggesting an important role for these proteins in the metabolic reprogramming of cancer cells.
Traditionally, cancer-associated alterations in metabolism have been considered a secondary response to cell proliferation signals. However, growing evidence has demonstrated that metabolic reprogramming of cancer cells is a primary function of activated oncogenes and inactivated tumor suppressors (Dang et al., 2012;DeBerardinis et al., 2008; Ward and Thompson, 2012). Despite this evidence, whether the metabolic reprogramming observed in cancer cells is a driving force for tumorigenesis remains as yet poorly understood.
Sirtuins are a family of NAD+-dependent protein deacetylases involved in stress resistance and metabolic homeostasis (Finkel et al., 2009). In mammals, there are seven members of this family (SIRT1-7). SIRT6 is a chromatin-bound factor that was first described as a suppressor of genomic instability (Mostoslavsky et al., 2006). SIRT6 also localizes to telomeres in human cells and controls cellular senescence and telomere structure by deacetylating histone H3 lysine 9 (H3K9) (Michishita et al., 2008). However, the main phenotype SIRT6 deficient mice display is an acute and severe metabolic abnormality. At 20 days of age, they develop a degenerative phenotype that includes complete loss of subcutaneous fat, lymphopenia, osteopenia, and acute onset of hypoglycemia, leading to death in less than ten days (Mostoslavsky et al., 2006). Recently, we have demonstrated that the lethal hypoglycemia exhibited by SIRT6 deficient mice is caused by an increased glucose uptake in muscle and brown adipose tissue (Zhong et al., 2010). Specifically, SIRT6 co-represses HIF-1α by deacetylating H3K9 at the promoters of several glycolytic genes and, consequently, SIRT6 deficient cells exhibit increased glucose uptake and upregulated glycolysis even under normoxic conditions (Zhong et al., 2010). Such a phenotype, reminiscent of the “Warburg Effect” in tumor cells, prompted us to investigate whether SIRT6 may protect against tumorigenesis by inhibiting glycolytic metabolism.
Here, we demonstrate that SIRT6 is a novel tumor suppressor that regulates aerobic glycolysis in cancer cells. Strikingly, SIRT6 acts as a first hit tumor suppressor and lack of this chromatin factor leads to tumor formation even in non-transformed cells. Notably, inhibition of glycolysis in SIRT6 deficient cells completely rescues their tumorigenic potential, suggesting that enhanced glycolysis is the driving force for tumorigenesis in these cells. Furthermore, we provide new data demonstrating that SIRT6 regulates cell proliferation by acting as a co-repressor of c-Myc, inhibiting the expression of ribosomal genes. Finally, SIRT6 expression is downregulated in human cancers, strongly reinforcing the idea that SIRT6 is a novel tumor suppressor.
…..
In addition to controlling glucose metabolism in cancer cells, our current work unravels a novel function of SIRT6 as a regulator of ribosomal gene expression. One of the main features of cancer cells is their high proliferative potential. In order to proliferate, cancer cells readjust their metabolism to generate biosynthetic precursors for macromolecular synthesis (Deberardinis et al., 2008). However, protein synthesis also requires the activation of a transcriptional program leading to ribosome biogenesis and mRNA translation (van Riggelen et al., 2010). As a master regulator of cell proliferation, MYC regulates ribosome biogenesis and protein synthesis by controlling the transcription and assembly of ribosome components as well as translation initiation (Dang et al., 2012; van Riggelen et al., 2010). Our results show that SIRT6 specifically regulates the expression of ribosomal genes. In keeping with this, SIRT6-deficient tumor cells exhibit high levels of ribosomal protein gene expression. Beyond ribosome biosynthesis, MYC regulates glucose and glutamine metabolism (Dang et al., 2012). Our results show that glutamine – but not glucose – metabolism is rescued in SIRT6-deficient/MYC knockdown cells, suggesting that SIRT6 and MYC might have redundant roles in regulating glucose metabolism.
Overall, our results indicate that SIRT6 represses tumorigenesis by inhibiting a glycolytic switch required for cancer cell proliferation. Inhibition of glycolysis in SIRT6-deficient cells abrogates tumor formation, providing proof of concept that inhibition of glycolytic metabolism in tumors with low SIRT6 levels could provide putative alternative approaches to modulate cancer growth. Furthermore, we uncover a new role for SIRT6 as a regulator of ribosome biosynthesis by co-repressing MYC transcriptional activity. Our results indicate that SIRT6 sits at a critical metabolic node, modulating both glycolytic metabolism and ribosome biosynthesis (Figure 7L). SIRT6 deficiency deregulates both pathways, leading to robust metabolic reprogramming that is sufficient to promote tumorigenesis bypassing major oncogenic signaling pathway activation.
Lack of cellular enzyme triggers switch in glucose processing
Understanding mechanism underlying SIRT6 activity may help treat diabetes, cancer
A study investigating how a cellular enzyme affects blood glucose levels in mice provides clues to pathways that may be involved in processes including the regulation of longevity and the proliferation of tumor cells. In their report in the January 22 issue of Cell, a Massachusetts General Hospital (MGH)-based team of researchers describes the mechanism by which absence of the enzyme SIRT6 induces a fatal drop in blood sugar in mice by triggering a switch between two critical cellular processes.
“We found that SIRT6 functions as a master regulator of glucose levels by maintaining the normal processes by which cells convert glucose into energy,” says Raul Mostoslavsky, MD, PhD, of the MGH Cancer Center, who led the study. “Learning more about how this protein controls the way cells handle glucose could lead to new approaches to treating type 2 diabetes and even cancer.”
SIRT6 belongs to a family of proteins called sirtuins, which regulate important biological pathways in organisms from bacteria to humans. Originally discovered in yeast, sirtuins in mammals have been shown to have important roles in metabolic regulation, programmed cell death and adaptation to stress. SIRT6 is one of seven mammalian sirtuins, and Mostoslavsky’s team previously showed that mice lacking the protein die in the first month of life from acute hypoglycemia. The current study was designed to investigate exactly how lack of SIRT6 causes this radical drop in blood sugar.
Normally cells convert glucose into energy through a two-step process. The first stage called glycolysis takes place in the cytoplasm, where glucose is broken down into an acid called pyruvate and a few molecules of ATP, the enzyme that provides the energy to power most biological processes. Pyruvate is taken into cellular structures called mitochondria, where it is further processed to release much greater amounts of ATP through a process called cellular respiration.
In a series of experiments in mouse cells, the researchers showed that SIRT6-deficiency hypoglycemia is caused by increased cellular uptake of glucose and not by elevated insulin levels or defects in the absorption of glucose from food. They then found increased levels of glycolysis and reduced mitochondrial respiration in SIRT6-knockout cells, something usually seen when cells are starved for oxygen or glucose, and showed that activation of the switch from cellular respiration to glycolysis is controlled through SIRT6’s regulation of a protein called HIF1alpha. Normally, SIRT6 represses glycolytic genes through its role as a compactor of chromatin – the tightly wound combination of DNA and a protein backbone that makes up chromosomes. In the absence of SIRT6, this structure is opened, causing activation of these glycolytic genes. The investigators’ finding increased expression of glycolytic genes in living SIRT6-knockout mice – which also had elevated levels of lactic acid, characteristic of a switch to glycolytic glucose processing – supported their cellular findings.
Studies in yeast, worms and flies have suggested a role for sirtuins in aging and longevity, and while much of the enzymes’ activity in mammals is unclear, SIRT6’s control of critical glucose-metabolic pathways could signify a contribution to lifespan regulation. Elevated glycolysis also is commonly found in tumor cells, suggesting that a lack of SIRT6 could contribute to tumor growth. Conversely, since knocking out SIRT6 causes blood sugar to drop, limited SIRT6 inhibition could be a novel strategy for treating type 2 diabetes.
“There’s a lot we still don’t know about SIRT6,” adds Mostoslavsky, who is an assistant professor of Medicine at Harvard Medical School. “We need to identify the factors that interact with SIRT6 and determine how it is regulated; investigate whether it acts as a tumor suppressor and how it might help lower glucose levels in diabetes; and determine its target organs in living animals, all of which we are investigating.”
A tale of metabolites: the crosstalk between chromatin and energy metabolism
Mitochondrial metabolism influences histone and DNA modifications by retrograde signaling and activation of transcriptional programs. Considering the high number of putative sites for acetylation and methylation in chromatin, we propose in this Perspective that epigenetic modifications might impinge on cellular metabolism by affecting the pool of acetyl-CoA and SAM.
Metabolism can be defined as the sum of chemical reactions that occur within a cell to sustain life. It is also the way that a cell interacts with energy sources: in other words, it is the coordination of energy intake, its utilization and storage that ultimately allows growth and cell division. In animal cells, mitochondria have evolved to become the most efficient system to generate energy. This organelle consumes carbon sources via oxidative phosphorylation to produce ATP, the energy currency of the cell. Additionally, the mitochondria produces intermediate metabolites for the biosynthesis of DNA, proteins and lipids.
Under basic dividing conditions, uptake of nutrients is tightly regulated through growth signaling pathways, thus differentiated cells engage in oxidative metabolism, the most efficient mechanism to produce energy from nutrients. Cells metabolize glucose to pyruvate through glycolysis in the cytoplasm, and this pyruvate is then oxidized into CO2 through the mitochondrial TCA cycle. The electrochemical gradient generated across the inner mitochondrial membrane facilitates ATP production in a highly efficient manner. Studies in recent years indicate that under conditions of nutrient excess, cells increase their nutrient uptake, adopting instead what is known as aerobic glycolysis, an adaptation that convert pyruvate into lactate, enabling cells to produce intermediate metabolites to sustain growth (anabolic metabolism) (1). Interestingly, most cancer cells undergo the same metabolic switch (Warburg Effect), a unique evolutionary trait that allows them to grow unabated. Although aerobic glycolysis generates much less ATP from glucose compared to oxidative phosphorylation, it provides critical intermediate metabolites that are used for anaplerotic reactions, and therefore is an obligatory adaptation among highly proliferative cells. In response to variations in nutrient availability, cells regulate their metabolic output, coordinating biochemical reactions and mitochondrial activity by altering transcription of mitochondrial genes through both activation of transcription factors, such as PGC1α, and chromatin modulators that exert epigenetic changes on metabolic genes.
Mitochondrial dysfunction has been implicated in aging, degenerative diseases and cancer. Proper mitochondrial function can be compromised by the accumulation of mutations in mitochondrial DNA that occur during aging. In addition, reactive oxygen species (ROS) produced during oxidative phosphorylation can promote oxidative damage to DNA, protein and lipids, in turn adversely affecting global cellular functions. In recent years, several studies have illustrated a novel unexpected link between metabolism and gene activity: fluctuations in mitochondrial and cytoplasmic metabolic reactions can reprogram global metabolism by means of impacting epigenetic dynamics. These studies will be briefly summarized in the first part of this article. In the second part, we will propose a provocative novel hypothesis: the crosstalk between metabolism and epigenetics is a two-way street, and defects in chromatin modulators may affect availability of intermediate metabolites, in turn influencing energy metabolism.
Metabolism impacts epigenetics
A regulated crosstalk between metabolic pathways in the mitochondria and epigenetic mechanisms in the nucleus allows cellular adaptations to new environmental conditions. Fine-tuning of gene expression is achieved by changes in chromatin dynamics, including methylation of DNA and posttranslational modifications of histones: acetyl, methyl and phosphate groups can be added by acetyltransferases, methyltransferases and kinases, respectively, to different residues on histones. Given the number of residues that can potentially undergo modifications in histone tails and in the DNA, it is reasonable to consider that metabolic changes affecting the availability of these metabolites will impact epigenetics (as discussed below).
Recently, acetylation of proteins was revealed to be as abundant as phosphorylation (2). This posttranslational modification involves the covalent binding of an acetyl group obtained from acetyl-CoA to a lysine. In histones, acetylation can modify higher order chromatin structure and serve as a docking site for histone code readers. Recent mass spectrometry studies have uncovered the complete acetylome in human cells and revealed that protein acetylation occurs broadly in the nucleus, cytoplasm and mitochondria, affecting more than 1700 proteins (2). Acetylation of proteins depends on the availability of acetyl-CoA in each cellular compartment, but this metabolite is produced in the mitochondria and cannot cross the mitochondrial membrane. In single cell eukaryotes, the pool of acetyl groups required for histone acetylation comes from the production of acetyl-CoA by the enzyme acetyl-CoA synthetase (Acs2p), which is responsible of converting acetate into acetyl-CoA. In mammalian cells, although they have a homolog enzyme to Acs2p, AceCS1, the majority of acetyl-CoA is produced from mitochondrion-derived citrate by the enzyme adenosine triphosphate (ATP)-citrate lyase (ACL) (3). ACL is present in the cytoplasm and in the nucleus, and is responsible for the production of acetyl-CoA from citrate in both compartments. Citrate is generated in the metabolism of glucose and glutamine in the TCA cycle. In contrast to acetyl-CoA, citrate can cross the mitochondrial membrane and diffuse through the nuclear pores into the nucleus, where it can be converted into acetyl-CoA by ACL. Wellen and colleagues found that ACL is required for acetylation of histones under normal growth conditions; knockdown of ACL decreases the pool of acetyl-CoA in the nucleus and reduces the level of histone acetylation (3). Strikingly, reduction in histone acetylation occurs preferentially around glycolytic genes, leading to downregulation of their transcription and therefore inhibition of glycolysis. These observations reveal a process where glucose metabolism dictates histone acetylation that in a feedback mechanism controls the rate of glycolysis.
Notably, deacetylation of histones also exhibits a metabolic influence. Deacetylation of histones is achieved by class I and class II histone deacetylases (HDACs) and by a separate class (class III), also known as sirtuins. Sirtuins use NAD+ as a cofactor for deacetylation, and the ratio of NAD+/NADH regulates their activity. In diets rich in carbohydrates, growth factors stimulate cellular glucose uptake and the production of energy is carried out through glycolysis. In this context, the NAD+/NADH ratio decreases, in turn inhibiting, in theory, sirtuins in the cytoplasm (Sirt2) and nucleus (Sirt1, Sirt6 and Sirt7). In fact, low Sirt1 and Sirt6 activity generates a global increase in protein acetylation. Interestingly, Sirt6, which is exclusively nuclear, deacetylates H3K9 Hif1α target genes, repressing their transcription. Since most of these genes are glycolytic, deacetylation of histones by Sirt6 modulates glycolysis. Indeed, SIRT6-deficient mice experience a dramatic increase in glucose uptake for glycolysis, triggering a fatal hypoglycemia in few weeks (4).
In animal cells, both histone acetylation and deacetylation are under the control of glucose metabolism through the availability of acetyl-CoA and NAD+, respectively. However, is this metabolic control restricted to acetylation, or can other reactions in the nucleus be influenced by the energy status of the cell?
Histone methyltransferases (HMTs) use S-adenosylmethionine (SAM) to transfer a methyl group onto lysine and arginine residues on histone tails. SAM is produced from methionine by the enzyme S-adenosyl methionine transferase (MAT) in a reaction that uses ATP. The recent finding of MAT in the nucleus suggests that the SAM pool could also be controlled locally in this compartment (5). The reverse reaction catalyzed by histone demethylases (HDMs) uses flavin adenine dinucleotide (FAD+) and α-ketoglutarate as coenzymes. FAD is a common redox coenzyme that exists in two different redox states. In its reduced state, FADH2 is a carrier of energy and when oxidized, FAD+ is consumed in the oxidation of succinate to fumarate by the enzyme succinate dehydrogenase (complex II) in one of the last steps of the TCA cycle. On the other hand, α-ketoglutarate is an intermediate in the TCA cycle. It is generated from isocitrate by the enzymes isocitrate dehydrogenase 1 and 2 (IDH1-cytosolic and IDH2-mitochondrial) (Figure 1A–B). Based on these findings, it is easy to infer that the amount of coenzymes used for histone methylation and demethylation could also be controlled by metabolic reactions. Moreover, the different cellular compartments compete for the same metabolites. Indeed, changes in diet that affect the biosynthesis of SAM, FAD and α-ketoglutarate in the mitochondria and cytoplasm have been shown to impact histone methylation (6).
Figure 1 A) Diagram depicting two-way crosstalk between metabolites in cytoplasm/mitochondria and chromatin.
More recently, some of the metabolic enzymes responsible for producing cofactors for nuclear biochemical reactions have been found mutated in cancer. For instance, IDH1 and IDH2 somatic mutations are recurrent in gliomas and acute myeloid leukemias (AML). These mutations lead not only to a decreased production of α-ketoglutarate but also to a new activity: α-ketoglutarate is in fact converted into 2-hydroxyglutarate (2-HG), a metabolite rarely found in normal cells. The new metabolite is a competitive inhibitor of α-ketoglutarate-dependent dioxygenase enzymes, including the Jumonji C (JmjC) domain containing histone demethylases and the recently discovered TET family of 5-methylcytosine (5mC) hydroxylases involved in DNA demethylation (7). By inhibiting JmjC and TET enzymes, the aberrant production of 2-HG generates a genome-wide histone and DNA hypermethylation phenotype. This is considered to be, at least in part, at the origin of tumorigenesis in IDH1 and IDH2 mutated cells and for this reason, 2-HG may earn its place as an oncometabolite. The discovery that mutations in metabolic enzymes may influence tumorigenesis by means of controlling genome-wide epigenetic changes caused a paradigm shift, indicating that such metabolic abnormalities may affect cancer beyond the Warburg Effect. ….
Chromatin modifications and cellular metabolism are tightly connected. Thus far the only aspects that have been considered are the retrograde signaling, with mitochondrial metabolites affecting histone modifications, and the anterograde transcriptional regulation of metabolism. A third aspect of the link between nucleus and metabolism has been, in our opinion, omitted so far: a direct influence of chromatin on acetyl-CoA and SAM availability, which may have an essential role also in cancer establishment and development (Figure 1A–B). Notably, a shift towards glycolytic metabolism is now considered a hallmark of cancer cells. It is also true that multiple tumors carry mutations in chromatin modifiers. However, new studies suggest that those two processes may be much more intertwined that previously appreciated, further blurring the limits on their respective roles in tumorigenesis. There is no doubt that changes in metabolite availability can drastically impact chromatin modifications. We believe that the opposite may be true as well. At least in mouse models, deficiency in two chromatin modifiers, SIRT6 and Jhdm2, causes drastic metabolic abnormalities. Even though some of those phenotypes depend on changes in gene-expression, we would like to propose that severe attrition of metabolite pools might as well play a role, a possibility that awaits experimental proof.
……
Investigators at UC San Diego say that when they blocked a well known signaling molecule that plays a major role in driving colorectal cancer, an escape pathway emerged that allowed tumors to continue to grow.
The pathway they explored, ERK1/2, is a problem for about a third of all colorectal cancer patients, says Petrus R. de Jong, MD, PhD, a co-first author on the paper.
“Since we were genetically deleting the ERK1/2 pathway, we expected to see less cell proliferation,” said de Jong. “Instead, the opposite occurred. There was more cell growth and loss of organization within the cells.”
The problem was ERK5, the investigators add. And when that was blocked as well in animal models and cell lines for the disease, the combination approach proved more effective in halting cancer growth.
“If you block one pathway, cancer cells usually mutate and find another pathway that ultimately allows for a recurrence of cancer growth,” said co-first author Koji Taniguchi. “Usually, mutations occur over weeks or months. But other times, as in this case, the tumor does not need to develop mutations to find an escape route from targeted therapy. When you find the compensatory pathway and block both, there is no more escape.”
Immunofluorescent staining of intestinal epithelium tissue shows cell growth (green). In a normal mouse model (left), cell growth is controlled, but in a mouse model with the ERK1/2 pathway blocked (right) increased cell proliferation and loss of organization occurred. [UC San Diego Health]
An international research team lead by scientists at the University of California San Diego School of Medicine uncovered some surprising results while investigating a potential therapeutic target for the ERK1 and two pathways. These signaling pathways are widely expressed and known to drive cancer growth in one-third of patients with colorectal cancer (CRC). The UCSD team found that an alternative pathway immediately emerges when ERK1/2 is halted, thus allowing tumor cell proliferation to continue.
“Since we were genetically deleting the ERK1/2 pathway, we expected to see less cell proliferation,” explained co-lead study author Petrus R. de Jong, M.D., Ph.D., translational scientist at Sanford Burnham Prebys Medical Discovery Institute. “Instead, the opposite occurred. There was more cell growth and loss of organization within the cells.”
The exciting part of this new study is investigators found that treating both ERK1/2 and the compensatory pathway ERK5 concomitantly with a combination of drug inhibitors halted CRC growth more effectively in both mouse models and human CRC cell lines.
“We show that loss of Erk1/2 in intestinal epithelial cells results in defects in nutrient absorption, epithelial cell migration, and secretory cell differentiation,” the authors wrote. “However, intestinal epithelial cell proliferation is not impeded, implying compensatory mechanisms. Genetic deletion ofErk1/2 or pharmacological targeting of MEK1/2 results in supraphysiological activity of the ERK5 pathway. Furthermore, targeting both pathways causes a more effective suppression of cell proliferation in murine intestinal organoids and human CRC lines.”
The findings from this study were published recently in Nature Communications in an article entitled “ERK5 Signalling Rescues Intestinal Epithelial Turnover and Tumour Cell Proliferation upon ERK1/2 Abrogation.”
The ERK pathway plays a critical role in embryonic development and tissue repair because it instructs cells to multiply and start dividing, but when overactivated cancer growth often occurs.
“Therapies aimed at targeting ERK1/2 likely fail because this mechanism is allowing proliferation through a different pathway,” noted senior study author Eyal Raz, M.D., professor of medicine at UC San Diego School of Medicine. “Previously, ERK5 didn’t seem important in colorectal cancer. This is an underappreciated escape pathway for tumor cells. Hence, the combination of ERK1/2 and ERK5 inhibitors may lead to more effective treatments for colorectal cancer patients.”
Currently, there are 1.2 million people living with CRC in the United States, making it the third most common cancer among men and women. In 2016 alone, an estimated 134,490 new cases are expected to be diagnosed, so understanding the molecular mechanisms that drive tumor promotion are paramount to treating this disease effectively.
“If you block one pathway, cancer cells usually mutate and find another pathway that ultimately allows for a recurrence of cancer growth,” remarked co-lead study author Koji Taniguchi, M.D., Ph.D., senior researcher at the Keio University School of Medicine in Tokyo. “Usually, mutations occur over weeks or months. But other times, as in this case, the tumor does not need to develop mutations to find an escape route from targeted therapy. When you find the compensatory pathway and block both, there is no more escape.”
The researchers were excited by their findings but urged caution at over interpretation of their initial findings and suggested that other classes of inhibitors be tested in combination with ERK5 inhibitors in human CRC cells in preclinical mouse models before any patient trial can begin.
ERK5 signalling rescues intestinal epithelial turnover and tumour cell proliferation upon ERK1/2 abrogation
The ERK1/2 MAPK signalling module integrates extracellular cues that induce proliferation and differentiation of epithelial lineages, and is an established oncogenic driver, particularly in the intestine. However, the interrelation of the ERK1/2 module relative to other signalling pathways in intestinal epithelial cells and colorectal cancer (CRC) is unclear. Here we show that loss of Erk1/2in intestinal epithelial cells results in defects in nutrient absorption, epithelial cell migration and secretory cell differentiation. However, intestinal epithelial cell proliferation is not impeded, implying compensatory mechanisms. Genetic deletion of Erk1/2 or pharmacological targeting of MEK1/2 results in supraphysiological activity of the ERK5 pathway. Furthermore, targeting both pathways causes a more effective suppression of cell proliferation in murine intestinal organoids and human CRC lines. These results suggest that ERK5 provides a common bypass route in intestinal epithelial cells, which rescues cell proliferation upon abrogation of ERK1/2 signalling, with therapeutic implications in CRC.
The extracellular signal-regulated kinases 1 and 2 (ERK1/2) are part of the classical family of mammalian mitogen-activated protein kinases (MAPKs), which also include three c-Jun amino-terminal kinases (JNK1/2/3), four p38 isoforms and its lesser-known counterpart, ERK5. The serine/threonine kinases ERK1 (MAPK3, also known as p44 MAPK) and ERK2 (MAPK1, also known as p42 MAPK) show 83% amino acid identity, are ubiquitously expressed and typically activated by growth factors and phorbol esters, whereas the p38 and JNK pathways are mainly activated by inflammatory cytokines and stress1. MAPKs are involved in regulation of mitosis, gene expression, cell metabolism, cell motility and apoptosis. ERK1/2 are activated by MEK1 and MEK2, which themselves are activated by Raf-1, A-Raf or B-Raf1, 2. Ras proteins (K-Ras, H-Ras or N-Ras) are small GTPases that can be activated by receptor tyrosine kinases (RTKs) or G-protein coupled receptors (GPCRs), which recruit Raf proteins to the plasma membrane where they are activated. Together, these modules constitute the Ras–Raf–MEK–ERK pathway3.
The activation of ERK1/2 results in their nuclear translocation where they can phosphorylate a variety of nuclear targets such as Elk-1, c-Fos and c-Myc1, in addition to p90 ribosomal S6 kinases (p90RSKs) and mitogen- and stress-activated protein kinases, MSK1/2. The full repertoire of substrates for ERK1/2 consists of at least 160 cellular proteins4. These proteins are typically involved in the regulation of cell proliferation—more specifically, G1/S-phase cell cycle progression—and differentiation. However, their cellular effects are context-dependent and determined by the spatial and temporal dynamics of ERK1/2 activity5, which are highly regulated by scaffolding proteins and phosphatases3, 6, 7.
Despite vast literature on the role of ERK1/2 in cell proliferation, the absolute requirement of this signalling module in rapidly dividing tissues relative to other signalling pathways is unknown. The small intestinal epithelium is particularly suitable to address this question given the short (4–8 days) and dynamic life cycle of intestinal epithelial cells (IECs). Lgr5+ intestinal stem cells at the intestinal crypt base produce transit-amplifying cells, which then undergo a number of proliferative cycles before terminal differentiation into absorptive enterocytes at the crypt–villus border. Enterocytes then migrate to the villus tip where they undergo anoikis and are shed into the gut lumen8. All of these cellular events are tightly coordinated by the Wnt, Notch, bone morphogenetic protein (BMP) and Hedgehog pathways9, whereas the roles of ERK1/2 remain to be charted. In the intestines, the ERK1/2 pathway is likely activated by autocrine and paracrine factors downstream of RTKs, such as epidermal growth factor receptor (EGFR)10, and by exogenous microbial-derived substrates that signal through the Toll-like receptor (TLR)/MyD88 pathway11.
To study the effects of ERK1/2 in the adult intestinal epithelium, we generated mice with a conditional (IEC-specific) and tamoxifen-inducible deletion of Erk2 on the Erk1−/− background, which completely abrogates this pathway. We show that the ERK1/2 signalling module, surprisingly, is dispensable for IEC proliferation. Genetic deletion of Erk1/2 in primary IEC or treatment of colorectal cancer (CRC) cell lines with MEK1/2 inhibitors results in compensatory activation of the ERK5 pathway. Moreover, the treatment of human CRC lines with a combination of MEK1/2 and ERK5 inhibitors is more efficacious in the inhibition of cancer cell growth. Thus, compensatory signalling by ERK5 suggests a potential rescue pathway that has clinical implications for targeted therapy in colorectal cancer.
….
Figure 1: Wasting disease associated with malabsorption in Erk1/2ΔIEC mice.
ERK1/2ΔIEC causes wasting and enterocyte dysfunction
Here we show that ERK1/2 signalling in mouse intestinal epithelium is dispensable for cell proliferation, while it resulted in abnormal differentiation of enterocytes, wasting disease and ultimately lethality (Fig. 1). Consistent with these findings, ERK1/2 MAPKs were shown to be associated with the enterocyte brush border and activated upon RTK stimulation or feeding27 or electrical field stimulation in polarized epithelium28. This seems at odds with literature that suggest that maintained ERK1/2 signalling precludes enterocyte differentiation29, 30. A possible explanation for this discrepancy could be that cycling IEC in the transit amplifying zone of the crypt require relatively high levels of active ERK1/2, which is readily blocked by pharmacological intervention, whereas a transition to low level ERK1/2 activity in IEC migrating into the villus compartment promotes the absorptive enterocyte differentiation program that is only perturbed upon complete genetic deletion of Erk1/2. Little is known about the role of ERK1/2 signalling in the life cycle of secretory cells in the gut. A recent report by Heuberger et al.15 described that IEC-specific deletion of non-receptor tyrosine phosphatase, Shp2, resulted in the loss of p-ERK1/2 levels in the small intestine. This coincided with an increased number of Paneth cells at the expense of goblet cells in the small intestine, as well as shortening of villi. They also observed the strongest staining for epithelial p-ERK1/2 in the TA zone. This p-ERK1/2+ staining pattern and the architectural organization of the TA zone were lost in Shp2 knockout mice. Interestingly, the deleterious effects of Shp2 deficiency were rescued by expression of constitutively active MEK1. A model was proposed in which the balance between Wnt/β-catenin and MAPK signalling determines Paneth cell versus goblet cell differentiation, respectively15. This proposed crucial role for ERK1/2 MAPK signalling in intestinal secretory cell differentiation is consistent with our observations inERK1/2ΔIECmice.
Migration and differentiation are functionally intertwined in the intestines, as demonstrated by the immature phenotype of mislocalized Paneth cells observed in ΔIEC mice (Fig. 2). Critical to epithelial cell migration is proper cytoskeleton reorganization mediated by the small GTPases of the Rho family, cell polarization regulated by Cdc42 and dynamic adhesion through cell–matrix and cell–cell interaction via integrin/FAK/Src signalling31. The ERK1/2 module is used as a downstream effector of many of these pathways in the intestine, including Rho GTPases32, FAK33and Src34, and has been suggested to promote cell motility33, 35. RTK signalling also contributes to cell migration, for example, Eph–Ephrin receptor interactions are crucial for correct positioning of Paneth cells36. Ephrin receptor-induced epithelial cell migration has been shown to be mediated by Src and ERK1/2 activation37, 38, which may explain the Paneth cell mislocalization observed in ΔIEC mice. In summary, the ERK1/2 module is indispensable for full maturation of both absorptive enterocytes and the secretory lineage (Fig. 7a), confirming its crucial role in the integration of cellular cues required for determination of epithelial cell fate.
Figure 7: Roles of ERK1/2 and ERK5 in intestinal homeostasis and tumorigenesis.
(a) When the ERK1/2 pathway is intact, extracellular cues that are transduced via RTKs or GPCRs activate Ras under physiological conditions, or alternatively, Ras is constitutively active in colorectal cancer (RasΔ*), which preferentially activates the Raf–MEK1/2–ERK1/2 module. The nuclear and transcriptional targets of ERK1/2 are crucial for enterocyte and secretory cell differentiation, IEC migration, as well as cell proliferation under homeostatic and oncogenic conditions. Importantly, ERK1/2 activation also results in the activation of negative feedback mechanisms that suppress its upstream kinases (for example, RTKs, son of sevenless, Raf) and activate dual specificity phosphatases (DUSPs), resulting in the silencing of the ERK5 module. (b) Upon abrogation of MEK1/2 or genetic knockout ofErk1/2, the lack of negative feedback mechanisms (that is, feedback activation) results in upregulation of the Ras–Raf–MEK5–ERK5 module, which maintains cell proliferation under physiological conditions, or results in continued tumour cell proliferation in colorectal cancer, respectively. However, the lack of activation of ERK1/2-specific targets results in differentiation and migration defects of intestinal epithelial cells culminating in malabsorption, wasting disease and mortality. Compensatory upregulation of the ERK5 pathway in CRC can be reversed by targeted treatment with its specific inhibitor, XMD8-92.
An unexpected finding was the redundancy of ERK1/2 in the gut with regard to cell proliferation.Erk1/2 deletion was compensated by upregulated ERK5 signalling. Genetic targeting of ERK1/2 in vitro previously showed that Erk2 knockdown is more effective than Erk1 knockdown in suppressing cell proliferation, although this may be related to higher expression levels of the former39. The effect of gene dosage was demonstrated in vivo by the observations that whileErk1−/− mice are viable12 and Erk2−/− mice die in utero13, Erk2+/− mice are only viable when at least one copy of Erk1 is present. However, mice heterozygous (+/−) for both Erk1 and Erk2 alleles were born at lower than Mendelian ratio39. More recently, it was reported that transgenic expression of ERK1 can compensate for Erk2 deletion40, demonstrating functional redundancy between both family members. Deletion of Erk1/2 in adult skin tissue resulted in hypoplasia, which was associated with G2/M cell cycle arrest, without notable differentiation defects of keratinocytes41. These data differ from our observations in the intestines, which might be explained by incomplete and transient siRNA-mediated knockdown of ERK1/2 in primary keratinocyte cultures41, compared with more efficient genomic deletion of Erk1 and Erk2 that is typically achieved by the Villin-Cre-ERT2 system14, possibly resulting in different outcomes.
Both ERK1/2 and ERK5 have been described to promote cell cycle progression, although they have different upstream signalling partners, MEK1/2 and MEK5, respectively1. Furthermore, ERK2 and ERK5 proteins share only about 66% sequence identity, and MEK5 is phosphorylated by MEKK2/3, which can also activate the p38 and JNK pathways42. The ERK5 pathway is classically activated by stress stimuli, in addition to mitogens; thus, it shares features of both the ERK1/2, and p38 and JNK pathways, respectively43. ERK5 induces expression of cyclin D1 (refs 44, 45), and suppresses expression of cyclin dependent kinase inhibitors46, thereby promoting G1/S-phase cell cycle progression. Importantly, the role of ERK5 in IEC differentiation and intestinal homeostasis is currently unknown. Knockout of Erk1/2 in IEC induced activity of ERK5, which was not detectable in naive mice (Fig. 3). These data suggest that the ERK1/2 and ERK5 modules may share proximal signalling components. Although EGFR is a likely candidate in this context19, 20, we found that abrogation of EGFR signalling did not prevent enhanced ERK5 activity upon MEK1/2 inhibition. Although it was originally suggested that ERK5 signalling is independent of Ras20, other groups established that Ras, either through physiological signalling47, or by its oncogenic activity48,49, activates the MEK5–ERK5 signalling axis. Thus, rewiring of signalling networks downstream of Ras could explain the supraphysiological activity of ERK5 upon conditional deletion of Erk1/2 in the intestines. In fact, it has been shown that ERK1/2 signalling mediates negative feedback on ERK5 activity50, possibly through transcriptional activation of dual specificity phosphatases (DUSPs)51. Alternatively, ERK1/2-induced FOS-like antigen 1 (Fra-1) may negatively regulate MEK5 (ref. 52). These data suggest that ERK5 is a default bypass route downstream of RTK-Ras and activated upon loss of ERK1/2-mediated repression, thereby ensuring the transduction of mitogenic signals to the nucleus (Fig. 7b). Consistent with this concept, we found that ERK5 inhibition induces atrophy of ΔIEC intestinal organoids (Fig. 4). In addition, important downstream transcriptional targets of ERK5 and ERK1/2 overlap, such as immediate-early gene Fra1 and oncogene c-Myc, whereas c-Fos and Egr1 were specifically induced by ERK1/2 (Fig. 6 and Supplementary Fig. 7). Specificity of ERK1/2 over ERK5 and other MAPK family members for the activation of c-Fos has been previously described53, demonstrating their differential biological output despite the shared ability to transduce potent mitogenic signals.
Our findings may be relevant for the use of MAPK inhibitors in the treatment of colorectal cancer. Although there was only a mild phenotype in the colons of ΔIEC mice under homeostatic conditions, the Ras–RAF–MEK–ERK pathway is generally upregulated in malignant cells including CRC54. Targeted therapy typically results in feedback activation of upstream players of the targeted kinase, which are then able to reactivate the same pathway or utilize bypass signalling routes55. For example, on activation, ERK1/2 phosphorylates EGFR, son of sevenless56, and Raf57, thereby terminating upstream signalling activity. Knockout of Erk1/2 eliminates this negative feedback. Our data suggest that ERK5 is a putative resistance pathway in the context of targeted treatment with MEK1/2 or ERK1/2 inhibitors (Fig. 7b). Different classes of MEK1/2 inhibitors display various modes of resistance to therapy (innate, adaptive and acquired)58. Since we have only used one MEK1/2 inhibitor (PD0325901) in our studies, it will be necessary to evaluate other classes of inhibitors in combination with ERK5 inhibitors. Importantly, while treatment with either the MEK1/2 or ERK5 inhibitor suppressed tumour growth in murine Apc−/− organoids, only the latter was able to inhibit the proliferation of Apc−/−;KRASG12V organoids (Fig. 6), which are more representative of human CRC. In line with this, suppression of ERK5 expression by forced expression of miR-143/145 inhibited intestinal adenoma formation in the ApcMin/+ model59, and activated MEK5 correlated with more invasive CRC in human60. ERK5 has been previously reported to mediate resistance to cytotoxic chemotherapy-induced apoptosis61. The highly specific and bioavailable ERK5 inhibitor, XMD8-92, has shown antitumour effects in multiple preclinical cancer models by inhibiting tumour angiogenesis, metastasis and chemo-resistance62. Furthermore, ERK5 inhibition does not induce feedback activation of upstream or parallel signalling pathways62. In conclusion, the ERK1/2 and ERK5 MAPK modules display a high degree of signalling plasticity in the intestinal epithelium, which has implications for targeted treatment of colorectal cancer.
Researchers Reveal Role of Transcription Factor Isoforms in Colon Diseases
Balance between the two isoforms, P1 and P2, of nuclear receptor HNF4a in the colonic crypt influences susceptibility to colitis and colon cancer. P1 is seen here in green. P2 is seen in red. [Poonamjot Deol, Sladek lab, UC Riverside]
Scientists at the University of California, Riverside have determined the distribution of the P1 and P2 isoforms of hepatocyte nuclear factor 4α (HNF4α) in the colons of mice. They report (“Opposing Roles of Nuclear Receptor HNF4α Isoforms in Colitis and Colitis-Associated Colon Cancer”) in eLife that maintaining a balance of P1 and P2 is crucial for reducing risk of contracting colon cancer and colitis.
What is already known in the field of cell biology is that the HNF4α transcription factor plays a key role in both diseases. HNF4α comes in two major isoforms, P1-HNF4α and P2-HNF4α (P1 and P2), but just how each isoform is involved in colitis and colon cancer is not understood.
“P1 and P2 have been conserved between mice and humans for 70 million years,” said Frances M. Sladek, Ph.D., professor of cell biology, who led the research project. “Both isoforms are important and we want to keep an appropriate balance between them in our gut by avoiding foods that would disrupt this balance and consuming foods that help preserve it. What these foods are is our next focus in the lab.”
The intestine is the only adult tissue in the body that expresses both P1 and P2. Dr. Sladek and her team have shown for the first time that these isoforms perform nonredundant functions in the intestine and are relevant to colitis and colitis-associated colon cancer.
“Our study also suggests that finding a drug to stabilize one isoform should be more effective than targeting both isoforms for treating colitis and colon cancer,” said Karthikeyani Chellappa, Ph.D., the first author of the research paper and a former postdoctoral researcher in Sladek’s lab.
Dr. Sladek explained that the colonic epithelial surface has finger-like invaginations (into the colonic wall) called colonic crypts that house stem cells at their base. These stem cells help regenerate new epithelial cells that continuously migrate up toward the surface, thus ensuring complete renewal of the intestinal lining every 3–5 days.
The researchers observed that the P1-positive cells were found in the surface lining and the top portion of the crypt (green in the accompanying image) whereas P2-positive cells were mostly in the proliferative compartment in the lower half of the crypt (the proliferation marker is red in the image.) Furthermore, when transgenic mice genetically engineered to have only either P1 or P2 were subjected to a carcinogen and, subsequently, to an irritant to stress the epithelial lining of the colon, the researchers found that the P1 mice showed fewer tumors than wild-type control mice. When treated with irritant alone, these mice were resistant to colitis. In sharp contrast, mice with only P2 showed more tumors and were much more susceptible to colitis.
The researchers explain these findings by invoking the “barrier function,” a mucosal barrier generated by the colon’s epithelial cells that prevents bacteria in the gut from entering the body. In the case of P1 mice, this barrier function was enhanced. The P2 mice, on the other hand, showed a compromised barrier function, presumably allowing bacteria to pass through.
Next, the researchers examined genes expressed in the P1 and P2 mice. They found that resistin-like molecule (RELM)-beta, a cytokine (a signaling molecule of the immune system) expressed in the gastrointestinal tract and implicated in colitis, was expressed far more in the P2 mice than the P1 mice.
“This makes sense since a reduced barrier function means bacteria can go across the barrier, which activates RELM-beta,” Dr. Sladek said. “We also found that the P2 protein transcribes RELM-beta more effectively than the P1 protein.”
Next, Poonamjot Deol, Ph.D., an assistant project scientist in Dr. Slaked’s lab and the second author of the eLife study, will lead a project aimed at understanding how diet affects the distribution of P1 and P2 in the gut. She and others in the lab also plan to investigate how obesity and colitis may be linked. (Diet studies performed in Dr. Sladek’s lab in the past illustrated soybean oil’s adverse effect on obesity.)
“In the case of colitis, could soybean oil be playing a part in allowing bacteria to get across the barrier function?” Dr. Deol said. “We do not know. We know its detrimental effect on obesity. But more research needs to be done where colitis is concerned.”
Opposing roles of nuclear receptor HNF4α isoforms in colitis and colitis-associated colon cancer
KarthikeyaniChellappa,
PoonamjotDeol
Department of Cell Biology and Neuroscience, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>PoonamjotDeol,
Jane REvans
Department of Cell Biology and Neuroscience, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>Jane REvans,
Linh MVuong
Department of Cell Biology and Neuroscience, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>Linh M, Vuong
GangChen
Division of Biomedical Sciences, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>GangChen,
NadegeBriançon
Department of Cell Biology, Harvard Medical School, Boston, United States
No competing interests declared
</div>”>NadegeBriançon,
EugeneBolotin
Department of Cell Biology and Neuroscience, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>EugeneBolotin,
ChristianLytle
Division of Biomedical Sciences, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>ChristianLytle,
Meera GNair
Division of Biomedical Sciences, University of California, Riverside, Riverside, United States
No competing interests declared
</div>”>Meera GNair,
Frances MSladek
Department of Cell Biology and Neuroscience, University of California Riverside, Riverside, United Statesfrances.sladek@ucr.edu
HNF4α has been implicated in colitis and colon cancer in humans but the role of the different HNF4α isoforms expressed from the two different promoters (P1 and P2) active in the colon is not clear. Here, we show that P1-HNF4α is expressed primarily in the differentiated compartment of the mouse colonic crypt and P2-HNF4α in the proliferative compartment. Exon swap mice that express only P1- or only P2-HNF4α have different colonic gene expression profiles, interacting proteins, cellular migration, ion transport and epithelial barrier function. The mice also exhibit altered susceptibilities to experimental colitis (DSS) and colitis-associated colon cancer (AOM+DSS). When P2-HNF4α-only mice (which have elevated levels of the cytokine resistin-like β, RELMβ, and are extremely sensitive to DSS) are crossed with Retnlb-/- mice, they are rescued from mortality. Furthermore, P2-HNF4α binds and preferentially activates the RELMβ promoter. In summary, HNF4α isoforms perform non-redundant functions in the colon under conditions of stress, underscoring the importance of tracking them both in colitis and colon cancer.
Disease related changes in proteomics, protein folding, protein-protein interaction, Volume 2 (Volume Two: Latest in Genomics Methodologies for Therapeutics: Gene Editing, NGS and BioInformatics, Simulations and the Genome Ontology), Part 1: Next Generation Sequencing (NGS)
Disease related changes in proteomics, protein folding, protein-protein interaction
Curator: Larry H. Bernstein, MD, FCAP
LPBI
Frankenstein Proteins Stitched Together by Scientists
The Frankenstein monster, stitched together from disparate body parts, proved to be an abomination, but stitched together proteins may fare better. They may, for example, serve specific purposes in medicine, research, and industry. At least, that’s the ambition of scientists based at the University of North Carolina. They have developed a computational protocol called SEWING that builds new proteins from connected or disconnected pieces of existing structures. [Wikipedia]
Unlike Victor Frankenstein, who betrayed Promethean ambition when he sewed together his infamous creature, today’s biochemists are relatively modest. Rather than defy nature, they emulate it. For example, at the University of North Carolina (UNC), researchers have taken inspiration from natural evolutionary mechanisms to develop a technique called SEWING—Structure Extension With Native-substructure Graphs. SEWING is a computational protocol that describes how to stitch together new proteins from connected or disconnected pieces of existing structures.
“We can now begin to think about engineering proteins to do things that nothing else is capable of doing,” said UNC’s Brian Kuhlman, Ph.D. “The structure of a protein determines its function, so if we are going to learn how to design new functions, we have to learn how to design new structures. Our study is a critical step in that direction and provides tools for creating proteins that haven’t been seen before in nature.”
Traditionally, researchers have used computational protein design to recreate in the laboratory what already exists in the natural world. In recent years, their focus has shifted toward inventing novel proteins with new functionality. These design projects all start with a specific structural “blueprint” in mind, and as a result are limited. Dr. Kuhlman and his colleagues, however, believe that by removing the limitations of a predetermined blueprint and taking cues from evolution they can more easily create functional proteins.
Dr. Kuhlman’s UNC team developed a protein design approach that emulates natural mechanisms for shuffling tertiary structures such as pleats, coils, and furrows. Putting the approach into action, the UNC team mapped 50,000 stitched together proteins on the computer, and then it produced 21 promising structures in the laboratory. Details of this work appeared May 6 in the journal Science, in an article entitled, “Design of Structurally Distinct Proteins Using Strategies Inspired by Evolution.”
“Helical proteins designed with SEWING contain structural features absent from other de novo designed proteins and, in some cases, remain folded at more than 100°C,” wrote the authors. “High-resolution structures of the designed proteins CA01 and DA05R1 were solved by x-ray crystallography (2.2 angstrom resolution) and nuclear magnetic resonance, respectively, and there was excellent agreement with the design models.”
Essentially, the UNC scientists confirmed that the proteins they had synthesized contained the unique structural varieties that had been designed on the computer. The UNC scientists also determined that the structures they had created had new surface and pocket features. Such features, they noted, provide potential binding sites for ligands or macromolecules.
“We were excited that some had clefts or grooves on the surface, regions that naturally occurring proteins use for binding other proteins,” said the Science article’s first author, Tim M. Jacobs, Ph.D., a former graduate student in Dr. Kuhlman’s laboratory. “That’s important because if we wanted to create a protein that can act as a biosensor to detect a certain metabolite in the body, either for diagnostic or research purposes, it would need to have these grooves. Likewise, if we wanted to develop novel therapeutics, they would also need to attach to specific proteins.”
Currently, the UNC researchers are using SEWING to create proteins that can bind to several other proteins at a time. Many of the most important proteins are such multitaskers, including the blood protein hemoglobin.
Histone Mutation Deranges DNA Methylation to Cause Cancer
In some cancers, including chondroblastoma and a rare form of childhood sarcoma, a mutation in histone H3 reduces global levels of methylation (dark areas) in tumor cells but not in normal cells (arrowhead). The mutation locks the cells in a proliferative state to promote tumor development. [Laboratory of Chromatin Biology and Epigenetics at The Rockefeller University]
They have been called oncohistones, the mutated histones that are known to accompany certain pediatric cancers. Despite their suggestive moniker, oncohistones have kept their oncogenic secrets. For example, it has been unclear whether oncohistones are able to cause cancer on their own, or whether they need to act in concert with additional DNA mutations, that is, mutations other than those affecting histone structures.
While oncohistone mechanisms remain poorly understood, this particular question—the oncogenicity of lone oncohistones—has been resolved, at least in part. According to researchers based at The Rockefeller University, a change to the structure of a histone can trigger a tumor on its own.
This finding appeared May 13 in the journal Science, in an article entitled, “Histone H3K36 Mutations Promote Sarcomagenesis Through Altered Histone Methylation Landscape.” The article describes the Rockefeller team’s study of a histone protein called H3, which has been found in about 95% of samples of chondoblastoma, a benign tumor that arises in cartilage, typically during adolescence.
The Rockefeller scientists found that the H3 lysine 36–to–methionine (H3K36M) mutation impairs the differentiation of mesenchymal progenitor cells and generates undifferentiated sarcoma in vivo.
After the scientists inserted the H3 histone mutation into mouse mesenchymal progenitor cells (MPCs)—which generate cartilage, bone, and fat—they watched these cells lose the ability to differentiate in the lab. Next, the scientists injected the mutant cells into living mice, and the animals developed the tumors rich in MPCs, known as an undifferentiated sarcoma. Finally, the researchers tried to understand how the mutation causes the tumors to develop.
The scientists determined that H3K36M mutant nucleosomes inhibit the enzymatic activities of several H3K36 methyltransferases.
“Depleting H3K36 methyltransferases, or expressing an H3K36I mutant that similarly inhibits H3K36 methylation, is sufficient to phenocopy the H3K36M mutation,” the authors of the Science study wrote. “After the loss of H3K36 methylation, a genome-wide gain in H3K27 methylation leads to a redistribution of polycomb repressive complex 1 and de-repression of its target genes known to block mesenchymal differentiation.”
Essentially, when the H3K36M mutation occurs, the cell becomes locked in a proliferative state—meaning it divides constantly, leading to tumors. Specifically, the mutation inhibits enzymes that normally tag the histone with chemical groups known as methyls, allowing genes to be expressed normally.
In response to this lack of modification, another part of the histone becomes overmodified, or tagged with too many methyl groups. “This leads to an overall resetting of the landscape of chromatin, the complex of DNA and its associated factors, including histones,” explained co-author Peter Lewis, Ph.D., a professor at the University of Wisconsin-Madison and a former postdoctoral fellow in laboratory of C. David Allis, Ph.D., a professor at Rockefeller.
The finding—that a “resetting” of the chromatin landscape can lock the cell into a proliferative state—suggests that researchers should be on the hunt for more mutations in histones that might be driving tumors. For their part, the Rockefeller researchers are trying to learn more about how this specific mutation in histone H3 causes tumors to develop.
“We want to know which pathways cause the mesenchymal progenitor cells that carry the mutation to continue to divide, and not differentiate into the bone, fat, and cartilage cells they are destined to become,” said co-author Chao Lu, Ph.D., a postdoctoral fellow in the Allis lab.
Once researchers understand more about these pathways, added Dr. Lewis, they can consider ways of blocking them with drugs, particularly in tumors such as MPC-rich sarcomas—which, unlike chondroblastoma, can be deadly. In fact, drugs that block these pathways may already exist and may even be in use for other types of cancers.
“One long-term goal of our collaborative team is to better understand fundamental mechanisms that drive these processes, with the hope of providing new therapeutic approaches,” concluded Dr. Allis.
Histone H3K36 mutations promote sarcomagenesis through altered histone methylation landscape
Missense mutations (that change one amino acid for another) in histone H3 can produce a so-called oncohistone and are found in a number of pediatric cancers. For example, the lysine-36–to-methionine (K36M) mutation is seen in almost all chondroblastomas. Lu et al. show that K36M mutant histones are oncogenic, and they inhibit the normal methylation of this same residue in wild-type H3 histones. The mutant histones also interfere with the normal development of bone-related cells and the deposition of inhibitory chromatin marks.
Several types of pediatric cancers reportedly contain high-frequency missense mutations in histone H3, yet the underlying oncogenic mechanism remains poorly characterized. Here we report that the H3 lysine 36–to–methionine (H3K36M) mutation impairs the differentiation of mesenchymal progenitor cells and generates undifferentiated sarcoma in vivo. H3K36M mutant nucleosomes inhibit the enzymatic activities of several H3K36 methyltransferases. Depleting H3K36 methyltransferases, or expressing an H3K36I mutant that similarly inhibits H3K36 methylation, is sufficient to phenocopy the H3K36M mutation. After the loss of H3K36 methylation, a genome-wide gain in H3K27 methylation leads to a redistribution of polycomb repressive complex 1 and de-repression of its target genes known to block mesenchymal differentiation. Our findings are mirrored in human undifferentiated sarcomas in which novel K36M/I mutations in H3.1 are identified.
Mitochondria? We Don’t Need No Stinking Mitochondria!
Diagram comparing typical eukaryotic cell to the newly discovered mitochondria-free organism. [Karnkowska et al., 2016, Current Biology 26, 1–11]
The organelle that produces a significant portion of energy for eukaryotic cells would seemingly be indispensable, yet over the years, a number of organisms have been discovered that challenge that biological pretense. However, these so-called amitochondrial species may lack a defined organelle, but they still retain some residual functions of their mitochondria-containing brethren. Even the intestinal eukaryotic parasite Giardia intestinalis, which was for many years considered to be mitochondria-free, was proven recently to contain a considerably shriveled version of the organelle.
Now, an international group of scientists has released results from a new study that challenges the notion that mitochondria are essential for eukaryotes—discovering an organism that resides in the gut of chinchillas that contains absolutely no trace of mitochondria at all.
“In low-oxygen environments, eukaryotes often possess a reduced form of the mitochondrion, but it was believed that some of the mitochondrial functions are so essential that these organelles are indispensable for their life,” explained lead study author Anna Karnkowska, Ph.D., visiting scientist at the University of British Columbia in Vancouver. “We have characterized a eukaryotic microbe which indeed possesses no mitochondrion at all.”
Mysterious Eukaryote Missing Mitochondria
Researchers uncover the first example of a eukaryotic organism that lacks the organelles.
Monocercomonoides sp. PA203VLADIMIR HAMPL, CHARLES UNIVERSITY, PRAGUE, CZECH REPUBLIC
Scientists have long thought that mitochondria—organelles responsible for energy generation—are an essential and defining feature of a eukaryotic cell. Now, researchers from Charles University in Prague and their colleagues are challenging this notion with their discovery of a eukaryotic organism,Monocercomonoides species PA203, which lacks mitochondria. The team’s phylogenetic analysis, published today (May 12) in Current Biology,suggests that Monocercomonoides—which belong to the Oxymonadida group of protozoa and live in low-oxygen environments—did have mitochondria at one point, but eventually lost the organelles.
“This is quite a groundbreaking discovery,” said Thijs Ettema, who studies microbial genome evolution at Uppsala University in Sweden and was not involved in the work.
“This study shows that mitochondria are not so central for all lineages of living eukaryotes,” Toni Gabaldonof the Center for Genomic Regulation in Barcelona, Spain, who also was not involved in the work, wrote in an email to The Scientist. “Yet, this mitochondrial-devoid, single-cell eukaryote is as complex as other eukaryotic cells in almost any other aspect of cellular complexity.”
Charles University’s Vladimir Hampl studies the evolution of protists. Along with Anna Karnkowska and colleagues, Hampl decided to sequence the genome of Monocercomonoides, a little-studied protist that lives in the digestive tracts of vertebrates. The 75-megabase genome—the first of an oxymonad—did not contain any conserved genes found on mitochondrial genomes of other eukaryotes, the researchers found. It also did not contain any nuclear genes associated with mitochondrial functions.
“It was surprising and for a long time, we didn’t believe that the [mitochondria-associated genes were really not there]. We thought we were missing something,” Hampl told The Scientist. “But when the data kept accumulating, we switched to the hypothesis that this organism really didn’t have mitochondria.”
Because researchers have previously not found examples of eukaryotes without some form of mitochondria, the current theory of the origin of eukaryotes poses that the appearance of mitochondria was crucial to the identity of these organisms.
“We now view these mitochondria-like organelles as a continuum from full mitochondria to very small . Some anaerobic protists, for example, have only pared down versions of mitochondria, such as hydrogenosomes and mitosomes, which lack a mitochondrial genome. But these mitochondrion-like organelles perform essential functions of the iron-sulfur cluster assembly pathway, which is known to be conserved in virtually all eukaryotic organisms studied to date.
Yet, in their analysis, the researchers found no evidence of the presence of any components of this mitochondrial pathway.
Like the scaling down of mitochondria into mitosomes in some organisms, the ancestors of modernMonocercomonoides once had mitochondria. “Because this organism is phylogenetically nested among relatives that had conventional mitochondria, this is most likely a secondary adaptation,” said Michael Gray, a biochemist who studies mitochondria at Dalhousie University in Nova Scotia and was not involved in the study. According to Gray, the finding of a mitochondria-deficient eukaryote does not mean that the organelles did not play a major role in the evolution of eukaryotic cells.
To be sure they were not missing mitochondrial proteins, Hampl’s team also searched for potential mitochondrial protein homologs of other anaerobic species, and for signature sequences of a range of known mitochondrial proteins. While similar searches with other species uncovered a few mitochondrial proteins, the team’s analysis of Monocercomonoides came up empty.
“The data is very complete,” said Ettema. “It is difficult to prove the absence of something but [these authors] do a convincing job.”
To form the essential iron-sulfur clusters, the team discovered that Monocercomonoides use a sulfur mobilization system found in the cytosol, and that an ancestor of the organism acquired this system by lateral gene transfer from bacteria. This cytosolic, compensating system allowed Monocercomonoides to lose the otherwise essential iron-sulfur cluster-forming pathway in the mitochondrion, the team proposed.
“This work shows the great evolutionary plasticity of the eukaryotic cell,” said Karnkowska, who participated in the study while she was a postdoc at Charles University. Karnkowska, who is now a visiting researcher at the University of British Columbia in Canada, added: “This is a striking example of how far the evolution of a eukaryotic cell can go that was beyond our expectations.”
“The results highlight how many surprises may await us in the poorly studied eukaryotic phyla that live in under-explored environments,” Gabaldon said.
Ettema agreed. “Now that we’ve found one, we need to look at the bigger picture and see if there are other examples of eukaryotes that have lost their mitochondria, to understand how adaptable eukaryotes are.”
Karnkowska et al., “A eukaryote without a mitochondrial organelle,” Current Biology,doi:10.1016/j.cub.2016.03.053, 2016.
•Monocercomonoides sp. is a eukaryotic microorganism with no mitochondria
•The complete absence of mitochondria is a secondary loss, not an ancestral feature
•The essential mitochondrial ISC pathway was replaced by a bacterial SUF system
The presence of mitochondria and related organelles in every studied eukaryote supports the view that mitochondria are essential cellular components. Here, we report the genome sequence of a microbial eukaryote, the oxymonad Monocercomonoides sp., which revealed that this organism lacks all hallmark mitochondrial proteins. Crucially, the mitochondrial iron-sulfur cluster assembly pathway, thought to be conserved in virtually all eukaryotic cells, has been replaced by a cytosolic sulfur mobilization system (SUF) acquired by lateral gene transfer from bacteria. In the context of eukaryotic phylogeny, our data suggest that Monocercomonoides is not primitively amitochondrial but has lost the mitochondrion secondarily. This is the first example of a eukaryote lacking any form of a mitochondrion, demonstrating that this organelle is not absolutely essential for the viability of a eukaryotic cell.
This method catches a bait protein together with its associated protein partners in virus-like particles that are budded from human cells. Like this, cell lysis is not needed and protein complexes are preserved during purification.
With his feet in both a proteomics lab and an interactomics lab, VIB/UGent professor Sven Eyckerman is well aware of the shortcomings of conventional approaches to analyze protein complexes. The lysis conditions required in mass spectrometry–based strategies to break open cell membranes often affect protein-protein interactions. “The first step in a classical study on protein complexes essentially turns the highly organized cellular structure into a big messy soup”, Eyckerman explains.
Inspired by virus biology, Eyckerman came up with a creative solution. “We used the natural process of HIV particle formation to our benefit by hacking a completely safe form of the virus to abduct intact protein machines from the cell.” It is well known that the HIV virus captures a number of host proteins during its particle formation. By fusing a bait protein to the HIV-1 GAG protein, interaction partners become trapped within virus-like particles that bud from mammalian cells. Standard proteomic approaches are used next to reveal the content of these particles. Fittingly, the team named the method ‘Virotrap’.
The Virotrap approach is exceptional as protein networks can be characterized under natural conditions. By trapping protein complexes in the protective environment of a virus-like shell, the intact complexes are preserved during the purification process. The researchers showed the method was suitable for detection of known binary interactions as well as mass spectrometry-based identification of novel protein partners.
Virotrap is a textbook example of bringing research teams with complementary expertise together. Cross-pollination with the labs of Jan Tavernier (VIB/UGent) and Kris Gevaert (VIB/UGent) enabled the development of this platform.
Jan Tavernier: “Virotrap represents a new concept in co-complex analysis wherein complex stability is physically guaranteed by a protective, physical structure. It is complementary to the arsenal of existing interactomics methods, but also holds potential for other fields, like drug target characterization. We also developed a small molecule-variant of Virotrap that could successfully trap protein partners for small molecule baits.”
Kris Gevaert: “Virotrap can also impact our understanding of disease pathways. We were actually surprised to see that this virus-based system could be used to study antiviral pathways, like Toll-like receptor signaling. Understanding these protein machines in their natural environment is essential if we want to modulate their activity in pathology.“
Trapping mammalian protein complexes in viral particles
Cell lysis is an inevitable step in classical mass spectrometry–based strategies to analyse protein complexes. Complementary lysis conditions, in situ cross-linking strategies and proximal labelling techniques are currently used to reduce lysis effects on the protein complex. We have developed Virotrap, a viral particle sorting approach that obviates the need for cell homogenization and preserves the protein complexes during purification. By fusing a bait protein to the HIV-1 GAG protein, we show that interaction partners become trapped within virus-like particles (VLPs) that bud from mammalian cells. Using an efficient VLP enrichment protocol, Virotrap allows the detection of known binary interactions and MS-based identification of novel protein partners as well. In addition, we show the identification of stimulus-dependent interactions and demonstrate trapping of protein partners for small molecules. Virotrap constitutes an elegant complementary approach to the arsenal of methods to study protein complexes.
Proteins mostly exert their function within supramolecular complexes. Strategies for detecting protein–protein interactions (PPIs) can be roughly divided into genetic systems1 and co-purification strategies combined with mass spectrometry (MS) analysis (for example, AP–MS)2. The latter approaches typically require cell or tissue homogenization using detergents, followed by capture of the protein complex using affinity tags3 or specific antibodies4. The protein complexes extracted from this ‘soup’ of constituents are then subjected to several washing steps before actual analysis by trypsin digestion and liquid chromatography–MS/MS analysis. Such lysis and purification protocols are typically empirical and have mostly been optimized using model interactions in single labs. In fact, lysis conditions can profoundly affect the number of both specific and nonspecific proteins that are identified in a typical AP–MS set-up. Indeed, recent studies using the nuclear pore complex as a model protein complex describe optimization of purifications for the different proteins in the complex by examining 96 different conditions5. Nevertheless, for new purifications, it remains hard to correctly estimate the loss of factors in a standard AP–MS experiment due to washing and dilution effects during treatments (that is, false negatives). These considerations have pushed the concept of stabilizing PPIs before the actual homogenization step. A classical approach involves cross-linking with simple reagents (for example, formaldehyde) or with more advanced isotope-labelled cross-linkers (reviewed in ref. 2). However, experimental challenges such as cell permeability and reactivity still preclude the widespread use of cross-linking agents. Moreover, MS-generated spectra of cross-linked peptides are notoriously difficult to identify correctly. A recent lysis-independent solution involves the expression of a bait protein fused to a promiscuous biotin ligase, which results in labelling of proteins proximal to the activity of the enzyme-tagged bait protein6. When compared with AP–MS, this BioID approach delivers a complementary set of candidate proteins, including novel interaction partners7, 8. Such particular studies clearly underscore the need for complementary approaches in the co-complex strategies.
The evolutionary stress on viruses promoted highly condensed coding of information and maximal functionality for small genomes. Accordingly, for HIV-1 it is sufficient to express a single protein, the p55 GAG protein, for efficient production of virus-like particles (VLPs) from cells9, 10. This protein is highly mobile before its accumulation in cholesterol-rich regions of the membrane, where multimerization initiates the budding process11. A total of 4,000–5,000 GAG molecules is required to form a single particle of about 145 nm (ref. 12). Both VLPs and mature viruses contain a number of host proteins that are recruited by binding to viral proteins. These proteins can either contribute to the infectivity (for example, Cyclophilin/FKBPA13) or act as antiviral proteins preventing the spreading of the virus (for example, APOBEC proteins14).
We here describe the development and application of Virotrap, an elegant co-purification strategy based on the trapping of a bait protein together with its associated protein partners in VLPs that are budded from the cell. After enrichment, these particles can be analysed by targeted (for example, western blotting) or unbiased approaches (MS-based proteomics). Virotrap allows detection of known binary PPIs, analysis of protein complexes and their dynamics, and readily detects protein binders for small molecules.
Concept of the Virotrap system
Classical AP–MS approaches rely on cell homogenization to access protein complexes, a step that can vary significantly with the lysis conditions (detergents, salt concentrations, pH conditions and so on)5. To eliminate the homogenization step in AP–MS, we reasoned that incorporation of a protein complex inside a secreted VLP traps the interaction partners under native conditions and protects them during further purification. We thus explored the possibility of protein complex packaging by the expression of GAG-bait protein chimeras (Fig. 1) as expression of GAG results in the release of VLPs from the cells9, 10. As a first PPI pair to evaluate this concept, we selected the HRAS protein as a bait combined with the RAF1 prey protein. We were able to specifically detect the HRAS–RAF1 interaction following enrichment of VLPs via ultracentrifugation (Supplementary Fig. 1a). To prevent tedious ultracentrifugation steps, we designed a novel single-step protocol wherein we co-express the vesicular stomatitis virus glycoprotein (VSV-G) together with a tagged version of this glycoprotein in addition to the GAG bait and prey. Both tagged and untagged VSV-G proteins are probably presented as trimers on the surface of the VLPs, allowing efficient antibody-based recovery from large volumes. The HRAS–RAF1 interaction was confirmed using this single-step protocol (Supplementary Fig. 1b). No associations with unrelated bait or prey proteins were observed for both protocols.
Figure 1: Schematic representation of the Virotrap strategy.
Expression of a GAG-bait fusion protein (1) results in submembrane multimerization (2) and subsequent budding of VLPs from cells (3). Interaction partners of the bait protein are also trapped within these VLPs and can be identified after purification by western blotting or MS analysis (4).
Virotrap for the detection of binary interactions
We next explored the reciprocal detection of a set of PPI pairs, which were selected based on published evidence and cytosolic localization15. After single-step purification and western blot analysis, we could readily detect reciprocal interactions between CDK2 and CKS1B, LCP2 and GRAP2, and S100A1 and S100B (Fig. 2a). Only for the LCP2 prey we observed nonspecific association with an irrelevant bait construct. However, the particle levels of the GRAP2 bait were substantially lower as compared with those of the GAG control construct (GAG protein levels in VLPs; Fig. 2a, second panel of the LCP2 prey). After quantification of the intensities of bait and prey proteins and normalization of prey levels using bait levels, we observed a strong enrichment for the GAG-GRAP2 bait (Supplementary Fig. 2).
…..
Virotrap for unbiased discovery of novel interactions
For the detection of novel interaction partners, we scaled up VLP production and purification protocols (Supplementary Fig. 5 and Supplementary Note 1 for an overview of the protocol) and investigated protein partners trapped using the following bait proteins: Fas-associated via death domain (FADD), A20 (TNFAIP3), nuclear factor-κB (NF-κB) essential modifier (IKBKG), TRAF family member-associated NF-κB activator (TANK), MYD88 and ring finger protein 41 (RNF41). To obtain specific interactors from the lists of identified proteins, we challenged the data with a combined protein list of 19 unrelated Virotrap experiments (Supplementary Table 1 for an overview). Figure 3 shows the design and the list of candidate interactors obtained after removal of all proteins that were found in the 19 control samples (including removal of proteins from the control list identified with a single peptide). The remaining list of confident protein identifications (identified with at least two peptides in at least two biological repeats) reveals both known and novel candidate interaction partners. All candidate interactors including single peptide protein identifications are given in Supplementary Data 2 and also include recurrent protein identifications of known interactors based on a single peptide; for example, CASP8 for FADD and TANK for NEMO. Using alternative methods, we confirmed the interaction between A20 and FADD, and the associations with transmembrane proteins (insulin receptor and insulin-like growth factor receptor 1) that were captured using RNF41 as a bait (Supplementary Fig. 6). To address the use of Virotrap for the detection of dynamic interactions, we activated the NF-κB pathway via the tumour necrosis factor (TNF) receptor (TNFRSF1A) using TNFα (TNF) and performed Virotrap analysis using A20 as bait (Fig. 3). This resulted in the additional enrichment of receptor-interacting kinase (RIPK1), TNFR1-associated via death domain (TRADD), TNFRSF1A and TNF itself, confirming the expected activated complex20.
Figure 3: Use of Virotrap for unbiased interactome analysis
Lysis conditions used in AP–MS strategies are critical for the preservation of protein complexes. A multitude of lysis conditions have been described, culminating in a recent report where protein complex stability was assessed under 96 lysis/purification protocols5. Moreover, the authors suggest to optimize the conditions for every complex, implying an important workload for researchers embarking on protein complex analysis using classical AP–MS. As lysis results in a profound change of the subcellular context and significantly alters the concentration of proteins, loss of complex integrity during a classical AP–MS protocol can be expected. A clear evolution towards ‘lysis-independent’ approaches in the co-complex analysis field is evident with the introduction of BioID6 and APEX25 where proximal proteins, including proteins residing in the complex, are labelled with biotin by an enzymatic activity fused to a bait protein. A side-by-side comparison between classical AP–MS and BioID showed overlapping and unique candidate binding proteins for both approaches7, 8, supporting the notion that complementary methods are needed to provide a comprehensive view on protein complexes. This has also been clearly demonstrated for binary approaches15 and is a logical consequence of the heterogenic nature underlying PPIs (binding mechanism, requirement for posttranslational modifications, location, affinity and so on).
In this report, we explore an alternative, yet complementary method to isolate protein complexes without interfering with cellular integrity. By trapping protein complexes in the protective environment of a virus-like shell, the intact complexes are preserved during the purification process. This constitutes a new concept in co-complex analysis wherein complex stability is physically guaranteed by a protective, physical structure. A comparison of our Virotrap approach with AP–MS shows complementary data, with specific false positives and false negatives for both methods (Supplementary Fig. 7).
The current implementation of the Virotrap platform implies the use of a GAG-bait construct resulting in considerable expression of the bait protein. Different strategies are currently pursued to reduce bait expression including co-expression of a native GAG protein together with the GAG-bait protein, not only reducing bait expression but also creating more ‘space’ in the particles potentially accommodating larger bait protein complexes. Nevertheless, the presence of the bait on the forming GAG scaffold creates an intracellular affinity matrix (comparable to the early in vitro affinity columns for purification of interaction partners from lysates26) that has the potential to compete with endogenous complexes by avidity effects. This avidity effect is a powerful mechanism that aids in the recruitment of cyclophilin to GAG27, a well-known weak interaction (Kd=16 μM (ref. 28)) detectable as a background association in the Virotrap system. Although background binding may be increased by elevated bait expression, weaker associations are readily detectable (for example, MAL—MYD88-binding study; Fig. 2c).
The size of Virotrap particles (around 145 nm) suggests limitations in the size of the protein complex that can be accommodated in the particles. Further experimentation is required to define the maximum size of proteins or the number of protein complexes that can be trapped inside the particles.
….
In conclusion, Virotrap captures significant parts of known interactomes and reveals new interactions. This cell lysis-free approach purifies protein complexes under native conditions and thus provides a powerful method to complement AP–MS or other PPI data. Future improvements of the system include strategies to reduce bait expression to more physiological levels and application of advanced data analysis options to filter out background. These developments can further aid in the deployment of Virotrap as a powerful extension of the current co-complex technology arsenal.
New Autism Blood Biomarker Identified
Researchers at UT Southwestern Medical Center have identified a blood biomarker that may aid in earlier diagnosis of children with autism spectrum disorder, or ASD
In a recent edition of Scientific Reports, UT Southwestern researchers reported on the identification of a blood biomarker that could distinguish the majority of ASD study participants versus a control group of similar age range. In addition, the biomarker was significantly correlated with the level of communication impairment, suggesting that the blood test may give insight into ASD severity.
“Numerous investigators have long sought a biomarker for ASD,” said Dr. Dwight German, study senior author and Professor of Psychiatry at UT Southwestern. “The blood biomarker reported here along with others we are testing can represent a useful test with over 80 percent accuracy in identifying ASD.”
ASD1 – was 66 percent accurate in diagnosing ASD. When combined with thyroid stimulating hormone level measurements, the ASD1-binding biomarker was 73 percent accurate at diagnosis
A Search for Blood Biomarkers for Autism: Peptoids
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by impairments in social interaction and communication, and restricted, repetitive patterns of behavior. In order to identify individuals with ASD and initiate interventions at the earliest possible age, biomarkers for the disorder are desirable. Research findings have identified widespread changes in the immune system in children with autism, at both systemic and cellular levels. In an attempt to find candidate antibody biomarkers for ASD, highly complex libraries of peptoids (oligo-N-substituted glycines) were screened for compounds that preferentially bind IgG from boys with ASD over typically developing (TD) boys. Unexpectedly, many peptoids were identified that preferentially bound IgG from TD boys. One of these peptoids was studied further and found to bind significantly higher levels (>2-fold) of the IgG1 subtype in serum from TD boys (n = 60) compared to ASD boys (n = 74), as well as compared to older adult males (n = 53). Together these data suggest that ASD boys have reduced levels (>50%) of an IgG1 antibody, which resembles the level found normally with advanced age. In this discovery study, the ASD1 peptoid was 66% accurate in predicting ASD.
….
Peptoid libraries have been used previously to search for autoantibodies for neurodegenerative diseases19 and for systemic lupus erythematosus (SLE)21. In the case of SLE, peptoids were identified that could identify subjects with the disease and related syndromes with moderate sensitivity (70%) and excellent specificity (97.5%). Peptoids were used to measure IgG levels from both healthy subjects and SLE patients. Binding to the SLE-peptoid was significantly higher in SLE patients vs. healthy controls. The IgG bound to the SLE-peptoid was found to react with several autoantigens, suggesting that the peptoids are capable of interacting with multiple, structurally similar molecules. These data indicate that IgG binding to peptoids can identify subjects with high levels of pathogenic autoantibodies vs. a single antibody.
In the present study, the ASD1 peptoid binds significantly lower levels of IgG1 in ASD males vs. TD males. This finding suggests that the ASD1 peptoid recognizes antibody(-ies) of an IgG1 subtype that is (are) significantly lower in abundance in the ASD males vs. TD males. Although a previous study14 has demonstrated lower levels of plasma IgG in ASD vs. TD children, here, we additionally quantified serum IgG levels in our individuals and found no difference in IgG between the two groups (data not shown). Furthermore, our IgG levels did not correlate with ASD1 binding levels, indicating that ASD1 does not bind IgG generically, and that the peptoid’s ability to differentiate between ASD and TD males is related to a specific antibody(-ies).
ASD subjects underwent a diagnostic evaluation using the ADOS and ADI-R, and application of the DSM-IV criteria prior to study inclusion. Only those subjects with a diagnosis of Autistic Disorder were included in the study. The ADOS is a semi-structured observation of a child’s behavior that allows examiners to observe the three core domains of ASD symptoms: reciprocal social interaction, communication, and restricted and repetitive behaviors1. When ADOS subdomain scores were compared with peptoid binding, the only significant relationship was with Social Interaction. However, the positive correlation would suggest that lower peptoid binding is associated with better social interaction, not poorer social interaction as anticipated.
The ADI-R is a structured parental interview that measures the core features of ASD symptoms in the areas of reciprocal social interaction, communication and language, and patterns of behavior. Of the three ADI-R subdomains, only the Communication domain was related to ASD1 peptoid binding, and this correlation was negative suggesting that low peptoid binding is associated with greater communication problems. These latter data are similar to the findings of Heuer et al.14 who found that children with autism with low levels of plasma IgG have high scores on the Aberrant Behavior Checklist (p < 0.0001). Thus, peptoid binding to IgG1 may be useful as a severity marker for ASD allowing for further characterization of individuals, but further research is needed.
It is interesting that in serum samples from older men, the ASD1 binding is similar to that in the ASD boys. This is consistent with the observation that with aging there is a reduction in the strength of the immune system, and the changes are gender-specific25. Recent studies using parabiosis26, in which blood from young mice reverse age-related impairments in cognitive function and synaptic plasticity in old mice, reveal that blood constituents from young subjects may contain important substances for maintaining neuronal functions. Work is in progress to identify the antibody/antibodies that are differentially binding to the ASD1 peptoid, which appear as a single band on the electrophoresis gel (Fig. 4).
……..
The ADI-R is a structured parental interview that measures the core features of ASD symptoms in the areas of reciprocal social interaction, communication and language, and patterns of behavior. Of the three ADI-R subdomains, only the Communication domain was related to ASD1 peptoid binding, and this correlation was negative suggesting that low peptoid binding is associated with greater communication problems. These latter data are similar to the findings of Heuer et al.14 who found that children with autism with low levels of plasma IgG have high scores on the Aberrant Behavior Checklist (p < 0.0001). Thus, peptoid binding to IgG1 may be useful as a severity marker for ASD allowing for further characterization of individuals, but further research is needed.
Titration of IgG binding to ASD1 using serum pooled from 10 TD males and 10 ASD males demonstrates ASD1’s ability to differentiate between the two groups. (B)Detecting IgG1 subclass instead of total IgG amplifies this differentiation. (C) IgG1 binding of individual ASD (n=74) and TD (n=60) male serum samples (1:100 dilution) to ASD1 significantly differs with TD>ASD. In addition, IgG1 binding of older adult male (AM) serum samples (n=53) to ASD1 is significantly lower than TD males, and not different from ASD males. The three groups were compared with a Kruskal-Wallis ANOVA, H = 10.1781, p<0.006. **p<0.005. Error bars show SEM. (D) Receiver-operating characteristic curve for ASD1’s ability to discriminate between ASD and TD males.
Association between peptoid binding and ADOS and ADI-R subdomains
Higher scores in any domain on the ADOS and ADI-R are indicative of more abnormal behaviors and/or symptoms. Among ADOS subdomains, there was no significant relationship between Communication and peptoid binding (z = 0.04, p = 0.966), Communication + Social interaction (z = 1.53, p = 0.127), or Stereotyped Behaviors and Restrictive Interests (SBRI) (z = 0.46, p = 0.647). Higher scores on the Social Interaction domain were significantly associated with higher peptoid binding (z = 2.04, p = 0.041).
Among ADI-R subdomains, higher scores on the Communication domain were associated with lower levels of peptoid binding (z = −2.28, p = 0.023). There was not a significant relationship between Social Interaction (z = 0.07, p = 0.941) or Restrictive/Repetitive Stereotyped Behaviors (z = −1.40, p = 0.162) and peptoid binding.
Computational Model Finds New Protein-Protein Interactions
Researchers at University of Pittsburgh have discovered 500 new protein-protein interactions (PPIs) associated with genes linked to schizophrenia.
Using a computational model they developed, researchers at the University of Pittsburgh School of Medicine have discovered more than 500 new protein-protein interactions (PPIs) associated with genes linked to schizophrenia. The findings, published online in npj Schizophrenia, a Nature Publishing Group journal, could lead to greater understanding of the biological underpinnings of this mental illness, as well as point the way to treatments.
There have been many genome-wide association studies (GWAS) that have identified gene variants associated with an increased risk for schizophrenia, but in most cases there is little known about the proteins that these genes make, what they do and how they interact, said senior investigator Madhavi Ganapathiraju, Ph.D., assistant professor of biomedical informatics, Pitt School of Medicine.
“GWAS studies and other research efforts have shown us what genes might be relevant in schizophrenia,” she said. “What we have done is the next step. We are trying to understand how these genes relate to each other, which could show us the biological pathways that are important in the disease.”
Each gene makes proteins and proteins typically interact with each other in a biological process. Information about interacting partners can shed light on the role of a gene that has not been studied, revealing pathways and biological processes associated with the disease and also its relation to other complex diseases.
Dr. Ganapathiraju’s team developed a computational model called High-Precision Protein Interaction Prediction (HiPPIP) and applied it to discover PPIs of schizophrenia-linked genes identified through GWAS, as well as historically known risk genes. They found 504 never-before known PPIs, and noted also that while schizophrenia-linked genes identified historically and through GWAS had little overlap, the model showed they shared more than 100 common interactors.
“We can infer what the protein might do by checking out the company it keeps,” Dr. Ganapathiraju explained. “For example, if I know you have many friends who play hockey, it could mean that you are involved in hockey, too. Similarly, if we see that an unknown protein interacts with multiple proteins involved in neural signaling, for example, there is a high likelihood that the unknown entity also is involved in the same.”
Dr. Ganapathiraju and colleagues have drawn such inferences on protein function based on the PPIs of proteins, and made their findings available on a website Schizo-Pi. This information can be used by biologists to explore the schizophrenia interactome with the aim of understanding more about the disease or developing new treatment drugs.
Schizophrenia interactome with 504 novel protein–protein interactions
(GWAS) have revealed the role of rare and common genetic variants, but the functional effects of the risk variants remain to be understood. Protein interactome-based studies can facilitate the study of molecular mechanisms by which the risk genes relate to schizophrenia (SZ) genesis, but protein–protein interactions (PPIs) are unknown for many of the liability genes. We developed a computational model to discover PPIs, which is found to be highly accurate according to computational evaluations and experimental validations of selected PPIs. We present here, 365 novel PPIs of liability genes identified by the SZ Working Group of the Psychiatric Genomics Consortium (PGC). Seventeen genes that had no previously known interactions have 57 novel interactions by our method. Among the new interactors are 19 drug targets that are targeted by 130 drugs. In addition, we computed 147 novel PPIs of 25 candidate genes investigated in the pre-GWAS era. While there is little overlap between the GWAS genes and the pre-GWAS genes, the interactomes reveal that they largely belong to the same pathways, thus reconciling the apparent disparities between the GWAS and prior gene association studies. The interactome including 504 novel PPIs overall, could motivate other systems biology studies and trials with repurposed drugs. The PPIs are made available on a webserver, called Schizo-Pi at http://severus.dbmi.pitt.edu/schizo-pi with advanced search capabilities.
Schizophrenia (SZ) is a common, potentially severe psychiatric disorder that afflicts all populations.1 Gene mapping studies suggest that SZ is a complex disorder, with a cumulative impact of variable genetic effects coupled with environmental factors.2 As many as 38 genome-wide association studies (GWAS) have been reported on SZ out of a total of 1,750 GWAS publications on 1,087 traits or diseases reported in the GWAS catalog maintained by the National Human Genome Research Institute of USA3 (as of April 2015), revealing the common variants associated with SZ.4 The SZ Working Group of the Psychiatric Genomics Consortium (PGC) identified 108 genetic loci that likely confer risk for SZ.5 While the role of genetics has been clearly validated by this study, the functional impact of the risk variants is not well-understood.6,7 Several of the genes implicated by the GWAS have unknown functions and could participate in possibly hitherto unknown pathways.8 Further, there is little or no overlap between the genes identified through GWAS and ‘candidate genes’ proposed in the pre-GWAS era.9
Interactome-based studies can be useful in discovering the functional associations of genes. For example,disrupted in schizophrenia 1 (DISC1), an SZ related candidate gene originally had no known homolog in humans. Although it had well-characterized protein domains such as coiled-coil domains and leucine-zipper domains, its function was unknown.10,11 Once its protein–protein interactions (PPIs) were determined using yeast 2-hybrid technology,12 investigators successfully linked DISC1 to cAMP signaling, axon elongation, and neuronal migration, and accelerated the research pertaining to SZ in general, and DISC1 in particular.13 Typically such studies are carried out on known protein–protein interaction (PPI) networks, or as in the case of DISC1, when there is a specific gene of interest, its PPIs are determined by methods such as yeast 2-hybrid technology.
Knowledge of human PPI networks is thus valuable for accelerating discovery of protein function, and indeed, biomedical research in general. However, of the hundreds of thousands of biophysical PPIs thought to exist in the human interactome,14,15 <100,000 are known today (Human Protein Reference Database, HPRD16 and BioGRID17 databases). Gold standard experimental methods for the determination of all the PPIs in human interactome are time-consuming, expensive and may not even be feasible, as about 250 million pairs of proteins would need to be tested overall; high-throughput methods such as yeast 2-hybrid have important limitations for whole interactome determination as they have a low recall of 23% (i.e., remaining 77% of true interactions need to be determined by other means), and a low precision (i.e., the screens have to be repeated multiple times to achieve high selectivity).18,19Computational methods are therefore necessary to complete the interactome expeditiously. Algorithms have begun emerging to predict PPIs using statistical machine learning on the characteristics of the proteins, but these algorithms are employed predominantly to study yeast. Two significant computational predictions have been reported for human interactome; although they have had high false positive rates, these methods have laid the foundation for computational prediction of human PPIs.20,21
We have created a new PPI prediction model called High-Confidence Protein–Protein Interaction Prediction (HiPPIP) model. Novel interactions predicted with this model are making translational impact. For example, we discovered a PPI between OASL and DDX58, which on validation showed that an increased expression of OASL could boost innate immunity to combat influenza by activating the RIG-I pathway.22 Also, the interactome of the genes associated with congenital heart disease showed that the disease morphogenesis has a close connection with the structure and function of cilia.23Here, we describe the HiPPIP model and its application to SZ genes to construct the SZ interactome. After computational evaluations and experimental validations of selected novel PPIs, we present here 504 highly confident novel PPIs in the SZ interactome, shedding new light onto several uncharacterized genes that are associated with SZ.
We developed a computational model called HiPPIP to predict PPIs (see Methods and Supplementary File 1). The model has been evaluated by computational methods and experimental validations and is found to be highly accurate. Evaluations on a held-out test data showed a precision of 97.5% and a recall of 5%. 5% recall out of 150,000 to 600,000 estimated number of interactions in the human interactome corresponds to 7,500–30,000 novel PPIs in the whole interactome. Note that, it is likely that the real precision would be higher than 97.5% because in this test data, randomly paired proteins are treated as non-interacting protein pairs, whereas some of them may actually be interacting pairs with a small probability; thus, some of the pairs that are treated as false positives in test set are likely to be true but hitherto unknown interactions. In Figure 1a, we show the precision versus recall of our method on ‘hub proteins’ where we considered all pairs that received a score >0.5 by HiPPIP to be novel interactions. In Figure 1b, we show the number of true positives versus false positives observed in hub proteins. Both these figures also show our method to be superior in comparison to the prediction of membrane-receptor interactome by Qi et al’s.24 True positives versus false positives are also shown for individual hub proteins by our method in Figure 1cand by Qi et al’s.23 in Figure 1d. These evaluations showed that our predictions contain mostly true positives. Unlike in other domains where ranked lists are commonly used such as information retrieval, in PPI prediction the ‘false positives’ may actually be unlabeled instances that are indeed true interactions that are not yet discovered. In fact, such unlabeled pairs predicted as interactors of the hub gene HMGB1 (namely, the pairs HMGB1-KL and HMGB1-FLT1) were validated by experimental methods and found to be true PPIs (See the Figures e–g inSupplementary File 3). Thus, we concluded that the protein pairs that received a score of ⩾0.5 are highly confident to be true interactions. The pairs that receive a score less than but close to 0.5 (i.e., in the range of 0.4–0.5) may also contain several true PPIs; however, we cannot confidently say that all in this range are true PPIs. Only the PPIs predicted with a score >0.5 are included in the interactome.
Computational evaluation of predicted protein–protein interactions on hub proteins: (a) precision recall curve. (b) True positive versus false positives in ranked lists of hub type membrane receptors for our method and that by Qi et al. True positives versus false positives are shown for individual membrane receptors by our method in (c) and by Qi et al. in (d). Thick line is the average, which is also the same as shown in (b). Note:x-axis is recall in (a), whereas it is number of false positives in (b–d). The range of y-axis is observed by varying the threshold from 1.0–0 in (a), and to 0.5 in (b–d).
SZ interactome
By applying HiPPIP to the GWAS genes and Historic (pre-GWAS) genes, we predicted over 500 high confidence new PPIs adding to about 1400 previously known PPIs.
Schizophrenia interactome: network view of the schizophrenia interactome is shown as a graph, where genes are shown as nodes and PPIs as edges connecting the nodes. Schizophrenia-associated genes are shown as dark blue nodes, novel interactors as red color nodes and known interactors as blue color nodes. The source of the schizophrenia genes is indicated by its label font, where Historic genes are shown italicized, GWAS genes are shown in bold, and the one gene that is common to both is shown in italicized and bold. For clarity, the source is also indicated by the shape of the node (triangular for GWAS and square for Historic and hexagonal for both). Symbols are shown only for the schizophrenia-associated genes; actual interactions may be accessed on the web. Red edges are the novel interactions, whereas blue edges are known interactions. GWAS, genome-wide association studies of schizophrenia; PPI, protein–protein interaction.
We have made the known and novel interactions of all SZ-associated genes available on a webserver called Schizo-Pi, at the addresshttp://severus.dbmi.pitt.edu/schizo-pi. This webserver is similar to Wiki-Pi33 which presents comprehensive annotations of both participating proteins of a PPI side-by-side. The difference between Wiki-Pi which we developed earlier, and Schizo-Pi, is the inclusion of novel predicted interactions of the SZ genes into the latter.
Despite the many advances in biomedical research, identifying the molecular mechanisms underlying the disease is still challenging. Studies based on protein interactions were proven to be valuable in identifying novel gene associations that could shed new light on disease pathology.35 The interactome including more than 500 novel PPIs will help to identify pathways and biological processes associated with the disease and also its relation to other complex diseases. It also helps identify potential drugs that could be repurposed to use for SZ treatment.
Functional and pathway enrichment in SZ interactome
When a gene of interest has little known information, functions of its interacting partners serve as a starting point to hypothesize its own function. We computed statistically significant enrichment of GO biological process terms among the interacting partners of each of the genes using BinGO36 (see online at http://severus.dbmi.pitt.edu/schizo-pi).
Protein aggregation and aggregate toxicity: new insights into protein folding, misfolding diseases and biological evolution
Massimo Stefani · Christopher M. Dobson
Abstract The deposition of proteins in the form of amyloid fibrils and plaques is the characteristic feature of more than 20 degenerative conditions affecting either the central nervous system or a variety of peripheral tissues. As these conditions include Alzheimer’s, Parkinson’s and the prion diseases, several forms of fatal systemic amyloidosis, and at least one condition associated with medical intervention (haemodialysis), they are of enormous importance in the context of present-day human health and welfare. Much remains to be learned about the mechanism by which the proteins associated with these diseases aggregate and form amyloid structures, and how the latter affect the functions of the organs with which they are associated. A great deal of information concerning these diseases has emerged, however, during the past 5 years, much of it causing a number of fundamental assumptions about the amyloid diseases to be reexamined. For example, it is now apparent that the ability to form amyloid structures is not an unusual feature of the small number of proteins associated with these diseases but is instead a general property of polypeptide chains. It has also been found recently that aggregates of proteins not associated with amyloid diseases can impair the ability of cells to function to a similar extent as aggregates of proteins linked with specific neurodegenerative conditions. Moreover, the mature amyloid fibrils or plaques appear to be substantially less toxic than the prefibrillar aggregates that are their precursors. The toxicity of these early aggregates appears to result from an intrinsic ability to impair fundamental cellular processes by interacting with cellular membranes, causing oxidative stress and increases in free Ca2+ that eventually lead to apoptotic or necrotic cell death. The ‘new view’ of these diseases also suggests that other degenerative conditions could have similar underlying origins to those of the amyloidoses. In addition, cellular protection mechanisms, such as molecular chaperones and the protein degradation machinery, appear to be crucial in the prevention of disease in normally functioning living organisms. It also suggests some intriguing new factors that could be of great significance in the evolution of biological molecules and the mechanisms that regulate their behaviour.
The genetic information within a cell encodes not only the specific structures and functions of proteins but also the way these structures are attained through the process known as protein folding. In recent years many of the underlying features of the fundamental mechanism of this complex process and the manner in which it is regulated in living systems have emerged from a combination of experimental and theoretical studies [1]. The knowledge gained from these studies has also raised a host of interesting issues. It has become apparent, for example, that the folding and unfolding of proteins is associated with a whole range of cellular processes from the trafficking of molecules to specific organelles to the regulation of the cell cycle and the immune response. Such observations led to the inevitable conclusion that the failure to fold correctly, or to remain correctly folded, gives rise to many different types of biological malfunctions and hence to many different forms of disease [2]. In addition, it has been recognised recently that a large number of eukaryotic genes code for proteins that appear to be ‘natively unfolded’, and that proteins can adopt, under certain circumstances, highly organised multi-molecular assemblies whose structures are not specifically encoded in the amino acid sequence. Both these observations have raised challenging questions about one of the most fundamental principles of biology: the close relationship between the sequence, structure and function of proteins, as we discuss below [3].
It is well established that proteins that are ‘misfolded’, i.e. that are not in their functionally relevant conformation, are devoid of normal biological activity. In addition, they often aggregate and/or interact inappropriately with other cellular components leading to impairment of cell viability and eventually to cell death. Many diseases, often known as misfolding or conformational diseases, ultimately result from the presence in a living system of protein molecules with structures that are ‘incorrect’, i.e. that differ from those in normally functioning organisms [4]. Such diseases include conditions in which a specific protein, or protein complex, fails to fold correctly (e.g. cystic fibrosis, Marfan syndrome, amyotonic lateral sclerosis) or is not sufficiently stable to perform its normal function (e.g. many forms of cancer). They also include conditions in which aberrant folding behaviour results in the failure of a protein to be correctly trafficked (e.g. familial hypercholesterolaemia, α1-antitrypsin deficiency, and some forms of retinitis pigmentosa) [4]. The tendency of proteins to aggregate, often to give species extremely intractable to dissolution and refolding, is of course also well known in other circumstances. Examples include the formation of inclusion bodies during overexpression of heterologous proteins in bacteria and the precipitation of proteins during laboratory purification procedures. Indeed, protein aggregation is well established as one of the major difficulties associated with the production and handling of proteins in the biotechnology and pharmaceutical industries [5].
Considerable attention is presently focused on a group of protein folding diseases known as amyloidoses. In these diseases specific peptides or proteins fail to fold or to remain correctly folded and then aggregate (often with other components) so as to give rise to ‘amyloid’ deposits in tissue. Amyloid structures can be recognised because they possess a series of specific tinctorial and biophysical characteristics that reflect a common core structure based on the presence of highly organised βsheets [6]. The deposits in strictly defined amyloidoses are extracellular and can often be observed as thread-like fibrillar structures, sometimes assembled further into larger aggregates or plaques. These diseases include a range of sporadic, familial or transmissible degenerative diseases, some of which affect the brain and the central nervous system (e.g. Alzheimer’s and Creutzfeldt-Jakob diseases), while others involve peripheral tissues and organs such as the liver, heart and spleen (e.g. systemic amyloidoses and type II diabetes) [7, 8]. In other forms of amyloidosis, such as primary or secondary systemic amyloidoses, proteinaceous deposits are found in skeletal tissue and joints (e.g. haemodialysis-related amyloidosis) as well as in several organs (e.g. heart and kidney). Yet other components such as collagen, glycosaminoglycans and proteins (e.g. serum amyloid protein) are often present in the deposits protecting them against degradation [9, 10, 11]. Similar deposits to those in the amyloidoses are, however, found intracellularly in other diseases; these can be localised either in the cytoplasm, in the form of specialised aggregates known as aggresomes or as Lewy or Russell bodies or in the nucleus (see below).
The presence in tissue of proteinaceous deposits is a hallmark of all these diseases, suggesting a causative link between aggregate formation and pathological symptoms (often known as the amyloid hypothesis) [7, 8, 12]. At the present time the link between amyloid formation and disease is widely accepted on the basis of a large number of biochemical and genetic studies. The specific nature of the pathogenic species, and the molecular basis of their ability to damage cells, are however, the subject of intense debate [13, 14, 15, 16, 17, 18, 19, 20]. In neurodegenerative disorders it is very likely that the impairment of cellular function follows directly from the interactions of the aggregated proteins with cellular components [21, 22]. In the systemic non-neurological diseases, however, it is widely believed that the accumulation in vital organs of large amounts of amyloid deposits can by itself cause at least some of the clinical symptoms [23]. It is quite possible, however, that there are other more specific effects of aggregates on biochemical processes even in these diseases. The presence of extracellular or intracellular aggregates of a specific polypeptide molecule is a characteristic of all the 20 or so recognised amyloid diseases. The polypeptides involved include full length proteins (e.g. lysozyme or immunoglobulin light chains), biological peptides (amylin, atrial natriuretic factor) and fragments of larger proteins produced as a result of specific processing (e.g. the Alzheimer βpeptide) or of more general degradation [e.g. poly(Q) stretches cleaved from proteins with poly(Q) extensions such as huntingtin, ataxins and the androgen receptor]. The peptides and proteins associated with known amyloid diseases are listed in Table 1. In some cases the proteins involved have wild type sequences, as in sporadic forms of the diseases, but in other cases these are variants resulting from genetic mutations associated with familial forms of the diseases. In some cases both sporadic and familial diseases are associated with a given protein; in this case the mutational variants are usually associated with early-onset forms of the disease. In the case of the neurodegenerative diseases associated with the prion protein some forms of the diseases are transmissible. The existence of familial forms of a number of amyloid diseases has provided significant clues to the origins of the pathologies. For example, there are increasingly strong links between the age at onset of familial forms of disease and the effects of the mutations involved on the propensity of the affected proteins to aggregate in vitro. Such findings also support the link between the process of aggregation and the clinical manifestations of disease [24, 25].
The presence in cells of misfolded or aggregated proteins triggers a complex biological response. In the cytosol, this is referred to as the ‘heat shock response’ and in the endoplasmic reticulum (ER) it is known as the ‘unfolded protein response’. These responses lead to the expression, among others, of the genes for heat shock proteins (Hsp, or molecular chaperone proteins) and proteins involved in the ubiquitin-proteasome pathway [26]. The evolution of such complex biochemical machinery testifies to the fact that it is necessary for cells to isolate and clear rapidly and efficiently any unfolded or incorrectly folded protein as soon as it appears. In itself this fact suggests that these species could have a generally adverse effect on cellular components and cell viability. Indeed, it was a major step forward in understanding many aspects of cell biology when it was recognised that proteins previously associated only with stress, such as heat shock, are in fact crucial in the normal functioning of living systems. This advance, for example, led to the discovery of the role of molecular chaperones in protein folding and in the normal ‘housekeeping’ processes that are inherent in healthy cells [27, 28]. More recently a number of degenerative diseases, both neurological and systemic, have been linked to, or shown to be affected by, impairment of the ubiquitin-proteasome pathway (Table 2). The diseases are primarily associated with a reduction in either the expression or the biological activity of Hsps, ubiquitin, ubiquitinating or deubiquitinating enzymes and the proteasome itself, as we show below [29, 30, 31, 32], or even to the failure of the quality control mechanisms that ensure proper maturation of proteins in the ER. The latter normally leads to degradation of a significant proportion of polypeptide chains before they have attained their native conformations through retrograde translocation to the cytosol [33, 34].
….
It is now well established that the molecular basis of protein aggregation into amyloid structures involves the existence of ‘misfolded’ forms of proteins, i.e. proteins that are not in the structures in which they normally function in vivo or of fragments of proteins resulting from degradation processes that are inherently unable to fold [4, 7, 8, 36]. Aggregation is one of the common consequences of a polypeptide chain failing to reach or maintain its functional three-dimensional structure. Such events can be associated with specific mutations, misprocessing phenomena, aberrant interactions with metal ions, changes in environmental conditions, such as pH or temperature, or chemical modification (oxidation, proteolysis). Perturbations in the conformational properties of the polypeptide chain resulting from such phenomena may affect equilibrium 1 in Fig. 1 increasing the population of partially unfolded, or misfolded, species that are much more aggregation-prone than the native state.
Fig. 1 Overview of the possible fates of a newly synthesised polypeptide chain. The equilibrium ① between the partially folded molecules and the natively folded ones is usually strongly in favour of the latter except as a result of specific mutations, chemical modifications or partially destabilising solution conditions. The increased equilibrium populations of molecules in the partially or completely unfolded ensemble of structures are usually degraded by the proteasome; when this clearance mechanism is impaired, such species often form disordered aggregates or shift equilibrium ② towards the nucleation of pre-fibrillar assemblies that eventually grow into mature fibrils (equilibrium ③). DANGER! indicates that pre-fibrillar aggregates in most cases display much higher toxicity than mature fibrils. Heat shock proteins (Hsp) can suppress the appearance of pre-fibrillar assemblies by minimising the population of the partially folded molecules by assisting in the correct folding of the nascent chain and the unfolded protein response target incorrectly folded proteins for degradation.
……
Little is known at present about the detailed arrangement of the polypeptide chains themselves within amyloid fibrils, either those parts involved in the core βstrands or in regions that connect the various β-strands. Recent data suggest that the sheets are relatively untwisted and may in some cases at least exist in quite specific supersecondary structure motifs such as β-helices [6, 40] or the recently proposed µ-helix [41]. It seems possible that there may be significant differences in the way the strands are assembled depending on characteristics of the polypeptide chain involved [6, 42]. Factors including length, sequence (and in some cases the presence of disulphide bonds or post-translational modifications such as glycosylation) may be important in determining details of the structures. Several recent papers report structural models for amyloid fibrils containing different polypeptide chains, including the Aβ40 peptide, insulin and fragments of the prion protein, based on data from such techniques as cryo-electron microscopy and solid-state magnetic resonance spectroscopy [43, 44]. These models have much in common and do indeed appear to reflect the fact that the structures of different fibrils are likely to be variations on a common theme [40]. It is also emerging that there may be some common and highly organised assemblies of amyloid protofilaments that are not simply extended threads or ribbons. It is clear, for example, that in some cases large closed loops can be formed [45, 46, 47], and there may be specific types of relatively small spherical or ‘doughnut’ shaped structures that can result in at least some circumstances (see below).
…..
The similarity of some early amyloid aggregates with the pores resulting from oligomerisation of bacterial toxins and pore-forming eukaryotic proteins (see below) also suggest that the basic mechanism of protein aggregation into amyloid structures may not only be associated with diseases but in some cases could result in species with functional significance. Recent evidence indicates that a variety of micro-organisms may exploit the controlled aggregation of specific proteins (or their precursors) to generate functional structures. Examples include bacterial curli [52] and proteins of the interior fibre cells of mammalian ocular lenses, whose β-sheet arrays seem to be organised in an amyloid-like supramolecular order [53]. In this case the inherent stability of amyloid-like protein structure may contribute to the long-term structural integrity and transparency of the lens. Recently it has been hypothesised that amyloid-like aggregates of serum amyloid A found in secondary amyloidoses following chronic inflammatory diseases protect the host against bacterial infections by inducing lysis of bacterial cells [54]. One particularly interesting example is a ‘misfolded’ form of the milk protein α-lactalbumin that is formed at low pH and trapped by the presence of specific lipid molecules [55]. This form of the protein has been reported to trigger apoptosis selectively in tumour cells providing evidence for its importance in protecting infants from certain types of cancer [55]. ….
Amyloid formation is a generic property of polypeptide chains ….
It is clear that the presence of different side chains can influence the details of amyloid structures, particularly the assembly of protofibrils, and that they give rise to the variations on the common structural theme discussed above. More fundamentally, the composition and sequence of a peptide or protein affects profoundly its propensity to form amyloid structures under given conditions (see below).
Because the formation of stable protein aggregates of amyloid type does not normally occur in vivo under physiological conditions, it is likely that the proteins encoded in the genomes of living organisms are endowed with structural adaptations that mitigate against aggregation under these conditions. A recent survey involving a large number of structures of β-proteins highlights several strategies through which natural proteins avoid intermolecular association of β-strands in their native states [65]. Other surveys of protein databases indicate that nature disfavours sequences of alternating polar and nonpolar residues, as well as clusters of several consecutive hydrophobic residues, both of which enhance the tendency of a protein to aggregate prior to becoming completely folded [66, 67].
……
Precursors of amyloid fibrils can be toxic to cells
It was generally assumed until recently that the proteinaceous aggregates most toxic to cells are likely to be mature amyloid fibrils, the form of aggregates that have been commonly detected in pathological deposits. It therefore appeared probable that the pathogenic features underlying amyloid diseases are a consequence of the interaction with cells of extracellular deposits of aggregated material. As well as forming the basis for understanding the fundamental causes of these diseases, this scenario stimulated the exploration of therapeutic approaches to amyloidoses that focused mainly on the search for molecules able to impair the growth and deposition of fibrillar forms of aggregated proteins. ….
Structural basis and molecular features of amyloid toxicity
The presence of toxic aggregates inside or outside cells can impair a number of cell functions that ultimately lead to cell death by an apoptotic mechanism [95, 96]. Recent research suggests, however, that in most cases initial perturbations to fundamental cellular processes underlie the impairment of cell function induced by aggregates of disease-associated polypeptides. Many pieces of data point to a central role of modifications to the intracellular redox status and free Ca2+ levels in cells exposed to toxic aggregates [45, 89, 97, 98, 99, 100, 101]. A modification of the intracellular redox status in such cells is associated with a sharp increase in the quantity of reactive oxygen species (ROS) that is reminiscent of the oxidative burst by which leukocytes destroy invading foreign cells after phagocytosis. In addition, changes have been observed in reactive nitrogen species, lipid peroxidation, deregulation of NO metabolism [97], protein nitrosylation [102] and upregulation of heme oxygenase-1, a specific marker of oxidative stress [103]. ….
Results have recently been reported concerning the toxicity towards cultured cells of aggregates of poly(Q) peptides which argues against a disease mechanism based on specific toxic features of the aggregates. These results indicate that there is a close relationship between the toxicity of proteins with poly(Q) extensions and their nuclear localisation. In addition they support the hypotheses that the toxicity of poly(Q) aggregates can be a consequence of altered interactions with nuclear coactivator or corepressor molecules including p53, CBP, Sp1 and TAF130 or of the interaction with transcription factors and nuclear coactivators, such as CBP, endowed with short poly(Q) stretches ([95] and references therein)…..
Concluding remarks
The data reported in the past few years strongly suggest that the conversion of normally soluble proteins into amyloid fibrils and the toxicity of small aggregates appearing during the early stages of the formation of the latter are common or generic features of polypeptide chains. Moreover, the molecular basis of this toxicity also appears to display common features between the different systems that have so far been studied. The ability of many, perhaps all, natural polypeptides to ‘misfold’ and convert into toxic aggregates under suitable conditions suggests that one of the most important driving forces in the evolution of proteins must have been the negative selection against sequence changes that increase the tendency of a polypeptide chain to aggregate. Nevertheless, as protein folding is a stochastic process, and no such process can be completely infallible, misfolded proteins or protein folding intermediates in equilibrium with the natively folded molecules must continuously form within cells. Thus mechanisms to deal with such species must have co-evolved with proteins. Indeed, it is clear that misfolding, and the associated tendency to aggregate, is kept under control by molecular chaperones, which render the resulting species harmless assisting in their refolding, or triggering their degradation by the cellular clearance machinery [166, 167, 168, 169, 170, 171, 172, 173, 175, 177, 178].
Misfolded and aggregated species are likely to owe their toxicity to the exposure on their surfaces of regions of proteins that are buried in the interior of the structures of the correctly folded native states. The exposure of large patches of hydrophobic groups is likely to be particularly significant as such patches favour the interaction of the misfolded species with cell membranes [44, 83, 89, 90, 91, 93]. Interactions of this type are likely to lead to the impairment of the function and integrity of the membranes involved, giving rise to a loss of regulation of the intracellular ion balance and redox status and eventually to cell death. In addition, misfolded proteins undoubtedly interact inappropriately with other cellular components, potentially giving rise to the impairment of a range of other biological processes. Under some conditions the intracellular content of aggregated species may increase directly, due to an enhanced propensity of incompletely folded or misfolded species to aggregate within the cell itself. This could occur as the result of the expression of mutational variants of proteins with decreased stability or cooperativity or with an intrinsically higher propensity to aggregate. It could also occur as a result of the overproduction of some types of protein, for example, because of other genetic factors or other disease conditions, or because of perturbations to the cellular environment that generate conditions favouring aggregation, such as heat shock or oxidative stress. Finally, the accumulation of misfolded or aggregated proteins could arise from the chaperone and clearance mechanisms becoming overwhelmed as a result of specific mutant phenotypes or of the general effects of ageing [173, 174].
The topics discussed in this review not only provide a great deal of evidence for the ‘new view’ that proteins have an intrinsic capability of misfolding and forming structures such as amyloid fibrils but also suggest that the role of molecular chaperones is even more important than was thought in the past. The role of these ubiquitous proteins in enhancing the efficiency of protein folding is well established [185]. It could well be that they are at least as important in controlling the harmful effects of misfolded or aggregated proteins as in enhancing the yield of functional molecules.
Nutritional Status is Associated with Faster Cognitive Decline and Worse Functional Impairment in the Progression of Dementia: The Cache County Dementia Progression Study1
Nutritional status may be a modifiable factor in the progression of dementia. We examined the association of nutritional status and rate of cognitive and functional decline in a U.S. population-based sample. Study design was an observational longitudinal study with annual follow-ups up to 6 years of 292 persons with dementia (72% Alzheimer’s disease, 56% female) in Cache County, UT using the Mini-Mental State Exam (MMSE), Clinical Dementia Rating Sum of Boxes (CDR-sb), and modified Mini Nutritional Assessment (mMNA). mMNA scores declined by approximately 0.50 points/year, suggesting increasing risk for malnutrition. Lower mMNA score predicted faster rate of decline on the MMSE at earlier follow-up times, but slower decline at later follow-up times, whereas higher mMNA scores had the opposite pattern (mMNA by time β= 0.22, p = 0.017; mMNA by time2 β= –0.04, p = 0.04). Lower mMNA score was associated with greater impairment on the CDR-sb over the course of dementia (β= 0.35, p < 0.001). Assessment of malnutrition may be useful in predicting rates of progression in dementia and may provide a target for clinical intervention.
Shared Genetic Risk Factors for Late-Life Depression and Alzheimer’s Disease
Background: Considerable evidence has been reported for the comorbidity between late-life depression (LLD) and Alzheimer’s disease (AD), both of which are very common in the general elderly population and represent a large burden on the health of the elderly. The pathophysiological mechanisms underlying the link between LLD and AD are poorly understood. Because both LLD and AD can be heritable and are influenced by multiple risk genes, shared genetic risk factors between LLD and AD may exist. Objective: The objective is to review the existing evidence for genetic risk factors that are common to LLD and AD and to outline the biological substrates proposed to mediate this association. Methods: A literature review was performed. Results: Genetic polymorphisms of brain-derived neurotrophic factor, apolipoprotein E, interleukin 1-beta, and methylenetetrahydrofolate reductase have been demonstrated to confer increased risk to both LLD and AD by studies examining either LLD or AD patients. These results contribute to the understanding of pathophysiological mechanisms that are common to both of these disorders, including deficits in nerve growth factors, inflammatory changes, and dysregulation mechanisms involving lipoprotein and folate. Other conflicting results have also been reviewed, and few studies have investigated the effects of the described polymorphisms on both LLD and AD. Conclusion: The findings suggest that common genetic pathways may underlie LLD and AD comorbidity. Studies to evaluate the genetic relationship between LLD and AD may provide insights into the molecular mechanisms that trigger disease progression as the population ages.
Association of Vitamin B12, Folate, and Sulfur Amino Acids With Brain Magnetic Resonance Imaging Measures in Older Adults: A Longitudinal Population-Based Study
Importance Vitamin B12, folate, and sulfur amino acids may be modifiable risk factors for structural brain changes that precede clinical dementia.
Objective To investigate the association of circulating levels of vitamin B12, red blood cell folate, and sulfur amino acids with the rate of total brain volume loss and the change in white matter hyperintensity volume as measured by fluid-attenuated inversion recovery in older adults.
Design, Setting, and Participants The magnetic resonance imaging subsample of the Swedish National Study on Aging and Care in Kungsholmen, a population-based longitudinal study in Stockholm, Sweden, was conducted in 501 participants aged 60 years or older who were free of dementia at baseline. A total of 299 participants underwent repeated structural brain magnetic resonance imaging scans from September 17, 2001, to December 17, 2009.
Main Outcomes and Measures The rate of brain tissue volume loss and the progression of total white matter hyperintensity volume.
Results In the multi-adjusted linear mixed models, among 501 participants (300 women [59.9%]; mean [SD] age, 70.9 [9.1] years), higher baseline vitamin B12 and holotranscobalamin levels were associated with a decreased rate of total brain volume loss during the study period: for each increase of 1 SD, β (SE) was 0.048 (0.013) for vitamin B12 (P < .001) and 0.040 (0.013) for holotranscobalamin (P = .002). Increased total homocysteine levels were associated with faster rates of total brain volume loss in the whole sample (β [SE] per 1-SD increase, –0.035 [0.015]; P = .02) and with the progression of white matter hyperintensity among participants with systolic blood pressure greater than 140 mm Hg (β [SE] per 1-SD increase, 0.000019 [0.00001]; P = .047). No longitudinal associations were found for red blood cell folate and other sulfur amino acids.
Conclusions and Relevance This study suggests that both vitamin B12 and total homocysteine concentrations may be related to accelerated aging of the brain. Randomized clinical trials are needed to determine the importance of vitamin B12supplementation on slowing brain aging in older adults.
Notes from Kurzweill
This vitamin stops the aging process in organs, say Swiss researchers
A potential breakthrough for regenerative medicine, pending further studies
Improved muscle stem cell numbers and muscle function in NR-treated aged mice: Newly regenerated muscle fibers 7 days after muscle damage in aged mice (left: control group; right: fed NR). (Scale bar = 50 μm). (credit: Hongbo Zhang et al./Science) http://www.kurzweilai.net/images/improved-muscle-fibers.png
EPFL researchers have restored the ability of mice organs to regenerate and extend life by simply administering nicotinamide riboside (NR) to them.
NR has been shown in previous studies to be effective in boosting metabolism and treating a number of degenerative diseases. Now, an article by PhD student Hongbo Zhang published in Science also describes the restorative effects of NR on the functioning of stem cells for regenerating organs.
As in all mammals, as mice age, the regenerative capacity of certain organs (such as the liver and kidneys) and muscles (including the heart) diminishes. Their ability to repair them following an injury is also affected. This leads to many of the disorders typical of aging.
Mitochondria —> stem cells —> organs
To understand how the regeneration process deteriorates with age, Zhang teamed up with colleagues from ETH Zurich, the University of Zurich, and universities in Canada and Brazil. By using several biomarkers, they were able to identify the molecular chain that regulates how mitochondria — the “powerhouse” of the cell — function and how they change with age. “We were able to show for the first time that their ability to function properly was important for stem cells,” said Auwerx.
Under normal conditions, these stem cells, reacting to signals sent by the body, regenerate damaged organs by producing new specific cells. At least in young bodies. “We demonstrated that fatigue in stem cells was one of the main causes of poor regeneration or even degeneration in certain tissues or organs,” said Zhang.
How to revitalize stem cells
Which is why the researchers wanted to “revitalize” stem cells in the muscles of elderly mice. And they did so by precisely targeting the molecules that help the mitochondria to function properly. “We gave nicotinamide riboside to 2-year-old mice, which is an advanced age for them,” said Zhang.
“This substance, which is close to vitamin B3, is a precursor of NAD+, a molecule that plays a key role in mitochondrial activity. And our results are extremely promising: muscular regeneration is much better in mice that received NR, and they lived longer than the mice that didn’t get it.”
Parallel studies have revealed a comparable effect on stem cells of the brain and skin. “This work could have very important implications in the field of regenerative medicine,” said Auwerx. This work on the aging process also has potential for treating diseases that can affect — and be fatal — in young people, like muscular dystrophy (myopathy).
So far, no negative side effects have been observed following the use of NR, even at high doses. But while it appears to boost the functioning of all cells, it could include pathological ones, so further in-depth studies are required.
Abstract of NAD+ repletion improves mitochondrial and stem cell function and enhances life span in mice
Adult stem cells (SCs) are essential for tissue maintenance and regeneration yet are susceptible to senescence during aging. We demonstrate the importance of the amount of the oxidized form of cellular nicotinamide adenine dinucleotide (NAD+) and its impact on mitochondrial activity as a pivotal switch to modulate muscle SC (MuSC) senescence. Treatment with the NAD+ precursor nicotinamide riboside (NR) induced the mitochondrial unfolded protein response (UPRmt) and synthesis of prohibitin proteins, and this rejuvenated MuSCs in aged mice. NR also prevented MuSC senescence in the Mdx mouse model of muscular dystrophy. We furthermore demonstrate that NR delays senescence of neural SCs (NSCs) and melanocyte SCs (McSCs), and increased mouse lifespan. Strategies that conserve cellular NAD+ may reprogram dysfunctional SCs and improve lifespan in mammals.
Discriminating the gene target of a distal regulatory element from other nearby transcribed genes is a challenging problem with the potential to illuminate the causal underpinnings of complex diseases. We present TargetFinder, a computational method that reconstructs regulatory landscapes from diverse features along the genome. The resulting models accurately predict individual enhancer–promoter interactions across multiple cell lines with a false discovery rate up to 15 times smaller than that obtained using the closest gene. By evaluating the genomic features driving this accuracy, we uncover interactions between structural proteins, transcription factors, epigenetic modifications, and transcription that together distinguish interacting from non-interacting enhancer–promoter pairs. Most of this signature is not proximal to the enhancers and promoters but instead decorates the looping DNA. We conclude that complex but consistent combinations of marks on the one-dimensional genome encode the three-dimensional structure of fine-scale regulatory interactions.
Cyclic Dinucleotides and Histone deacetylase inhibitors
Curators: Larry H. Bernsten, MD, FCAP and Aviva Lev-Ari, PhD, RN
LPBI
New Class of Immune System Stimulants: Cyclic Di-Nucleotides (CDN): Shrink Tumors and bolster Vaccines, re-arm the Immune System’s Natural Killer Cells, which attack Cancer Cells and Virus-infected Cells
Reporter: Aviva Lev-Ari, PhD, RN
The Immunotherapeutics and Vaccine Research Initiative (IVRI), a UC Berkeley effort funded by Aduro Biotech, Inc.
A new class of immune system stimulants called cyclic di-nucleotides have shown promise in shrinking tumors and bolstering vaccines against tuberculosis, and research that could help re-arm the immune system’s natural killer cells, which normally attack cancer cells and virus-infected cells, to better fight tumors.
Much of the excitement around combining these two areas — the immunology of cancer and the immunology of infectious disease — comes from the amazing success of immunotherapy against cancer, which started with the work of James Allison when he was a professor of immunology at UC Berkeley and director of the Cancer Research Laboratory from 1985 to 2004. Allison, now at the University of Texas MD Anderson Cancer Center, discovered a way to release a brake on the body’s immune response to cancer that has proved highly successful at unleashing the immune system to attack melanoma and is being tried against other types of cancer. Allison’s technique uses an antibody that blocks an immune suppressor called CTLA4, antibodies that block another immune suppressor, PD1. This has been successful in treating melanoma, renal cancer and a type of lung cancer. Both CTLA4 and PD1 antibodies are now FDA-approved as cancer therapies.
Another promising avenue involves a protein in cells that responds to foreign DNA to launch an innate immune response — the first response of the body’s immune system. The protein, dubbed STING, is triggered by small molecules called cyclic di-nucleotides (CDN), and makes immune cells release interferon and other cytokines that activate disease-fighting T cells and stimulate the production of antibodies that together kill invading pathogens and destroy cancer cells. Listeria bacteria, for example, secrete a CDN directly into infected cells that activates STING.
Russell Vance, a UC Berkeley professor of molecular and cell biology and current head of the Cancer Research Laboratory, discovered several years ago that the chemical structure of these di-nucleotides is critical to their ability to work in humans. Aduro has since developed a CDN designed to work in humans and found that injecting it directly into a tumor in mice caused the tumor to shrink.
Sarah Stanley, an assistant professor of public health, has found evidence that CDNs can help improve the imperfect vaccines we have today against tuberculosis.
Researchers at UC Berkeley will have access to Aduro’s novel technology platforms for research use, including its STING pathway activators, proprietary monoclonal antibodies and the engineered listeria bacteria, referred to as LADD (listeria attenuated double-deleted). David Raulet, professor of molecular and cell biology and director of IVRI has contributed to making these cells a new focus of cancer research. As tumors advance, NK cells inside the tumors appear to become desensitized, he said. Recent research shows that some cytokines and other immune mediators Raulet discovered are able to “wake them up” and get them to resume their elimination of cancer cells.
Histone deacetylase inhibitors: molecular mechanisms of action
This review focuses on the mechanisms of action of histone deacetylase (HDAC) inhibitors (HDACi), a group of recently discovered ‘targeted’ anticancer agents. There are 18 HDACs, which are generally divided into four classes, based on sequence homology to yeast counterparts. Classical HDACi such as the hydroxamic acid-based vorinostat (also known as SAHA and Zolinza) inhibits classes I, II and IV, but not the NAD+-dependent class III enzymes. In clinical trials, vorinostat has activity against hematologic and solid cancers at doses well tolerated by patients. In addition to histones, HDACs have many other protein substrates involved in regulation of gene expression, cell proliferation and cell death. Inhibition of HDACs causes accumulation of acetylated forms of these proteins, altering their function. Thus, HDACs are more properly called ‘lysine deacetylases.’ HDACi induces different phenotypes in various transformed cells, including growth arrest, activation of the extrinsic and/or intrinsic apoptotic pathways, autophagic cell death, reactive oxygen species (ROS)-induced cell death, mitotic cell death and senescence. In comparison, normal cells are relatively more resistant to HDACi-induced cell death. The plurality of mechanisms of HDACi-induced cell death reflects both the multiple substrates of HDACs and the heterogeneous patterns of molecular alterations present in different cancer cells.
Acetylation and deacetylation of histones play an important role in transcription regulation of eukaryotic cells (Lehrmann et al., 2002;Mai et al., 2005). The acetylation status of histones and non-histone proteins is determined by histone deacetylases (HDACs) and histone acetyl-transferases (HATs). HATs add acetyl groups to lysine residues, while HDACs remove the acetyl groups. In general, acetylation of histone promotes a more relaxed chromatin structure, allowing transcriptional activation. HDACs can act as transcription repressors, due to histone deacetylation, and consequently promote chromatin condensation. HDAC inhibitors (HDACi) selectively alter gene transcription, in part, by chromatin remodeling and by changes in the structure of proteins in transcription factor complexes (Gui et al., 2004). Further, the HDACs have many non-histone proteins substrates such as hormone receptors, chaperone proteins and cytoskeleton proteins, which regulate cell proliferation and cell death (Table 1). Thus, HDACi-induced transformed cell death involves transcription-dependent and transcription-independent mechanisms (Marks and Dokmanovic, 2005; Rosato and Grant, 2005; Bolden et al., 2006;Minucci and Pelicci, 2006).
In humans, 18 HDAC enzymes have been identified and classified, based on homology to yeast HDACs (Blander and Guarente, 2004;Bhalla, 2005; Marks and Dokmanovic, 2005). Class I HDACs include HDAC1, 2, 3 and 8, which are related to yeast RPD3 deacetylase and have high homology in their catalytic sites. Recent phylogenetic analyses suggest that this class can be divided into classes Ia (HDAC1 and -2), Ib (HDAC3) and Ic (HDAC8) (Gregoretti et al., 2004). Class II HDACs are related to yeast Hda1 (histone deacetylase 1) and include HDAC4, -5, -6, -7, -9 and -10 (Bhalla, 2005; Marks and Dokmanovic, 2005). This class is divided into class IIa, consisting of HDAC4, -5, -7 and -9, and class IIb, consisting of HDAC6 and -10, which contain two catalytic sites. All class I and II HDACs are zinc-dependent enzymes. Members of a third class, sirtuins, require NAD+ for their enzymatic activity (Blander and Guarente, 2004) (see review by E Verdin, in this issue). Among them, SIRT1 is orthologous to yeast silent information regulator 2. The enzymatic activity of class III HDACs is not inhibited by compounds such as vorinostat or trichostatin A (TSA), that inhibit class I and II HDACs. Class IV HDAC is represented by HDAC11, which, like yeast Hda 1 similar 3, has conserved residues in the catalytic core region shared by both class I and II enzymes (Gao et al., 2002).
HDACs are not redundant in function (Marks and Dokmanovic, 2005; Rosato and Grant, 2005; Bolden et al., 2006). Class I HDACs are primarily nuclear in localization and ubiquitously expressed, while class II HDACs can be primarily cytoplasmic and/or migrate between the cytoplasm and nucleus and are tissue-restricted in expression.
The structural details of the HDAC–HDACi interaction has been elucidated in studies of a histone deacetylase-like protein from an anerobic bacterium with TSA and vorinostat (Finnin et al., 1999). More recently, the crystal structure of HDAC8–hydroxamate interaction has been solved (Somoza et al., 2004; Vannini et al., 2004). These studies provide an insight into the mechanism of deacetylation of acetylated substrates. The hydroxamic acid moiety of the inhibitor directly interacts with the zinc ion at the base of the catalytic pocket.
This review focuses on the molecular mechanisms triggered by inhibitors of zinc-dependent HDACs in tumor cells that explain in part: (I) the effects of these compounds in inducing transformed cell death and (II) the relative resistance of normal and certain cancer cells to HDACi induced cell death. HDACi, for example, the hydroxamic acid-based vorinostat (SAHA, Zolinza), are promising drugs for cancer treatment (Richon et al., 1998; Marks and Breslow, 2007). Several HDACi are in phase I and II clinical trials, being tested in different tumor types, such as cutaneous T-cell lymphoma, acute myeloid leukemia, cervical cancer, etc (Bug et al., 2005; Chavez-Blanco et al., 2005; Kelly and Marks, 2005;Duvic and Zhang, 2006) (Table 2). Although considerable progress has been made in elucidating the role of HDACs and the effects of HDACi, these areas are still in early stages of discovery.
Recent phylogenetic analyses of bacterial HDACs suggest that all four HDAC classes preceded the evolution of histone proteins (Gregoretti et al., 2004). This suggests that the primary activity of HDACs may be directed against non-histone substrates. At least 50 non-histone proteins of known biological function have been identified, which may be acetylated and substrates of HDACs (Table 1) (Glozak et al., 2005; Marks and Dokmanovic, 2005;Rosato and Grant, 2005; Bolden et al., 2006; Minucci and Pelicci, 2006; Zhao et al., 2006). In addition, two recent proteomic studies identified many lysine-acetylated substrates (Iwabata et al., 2005; Kim et al., 2006). In view of all these findings, HDACs may be better called ‘N-epsilon-lysine deacetylase’. This designation would also distinguish them from the enzymes that catalyse other types of deacetylation in biological reactions, such as acylases that catalyse the deacetylation of a range of N-acetyl amino acids (Anders and Dekant, 1994).
Non-histone protein targets of HDACs include transcription factors, transcription regulators, signal transduction mediators, DNA repair enzymes, nuclear import regulators, chaperone proteins, structural proteins, inflammation mediators and viral proteins (Table 1). Acetylation can either increase or decrease the function or stability of the proteins, or protein–protein interaction (Glozak et al., 2005). These HDAC substrates are directly or indirectly involved in many biological processes, such as gene expression and regulation of pathways of proliferation, differentiation and cell death. These data suggest that HDACi could have multiple mechanisms of inducing cell growth arrest and cell death (Figure 1).
HDACi have been discovered with different structural characteristics, including hydroximates, cyclic peptides, aliphatic acids and benzamides (Table 2) (Miller et al., 2003; Yoshida et al., 2003; Marks and Breslow, 2007). Certain HDACi may selectively inhibit different HDACs. For example, MS-275 preferentially inhibits HDAC1 with IC50, at 0.3 m, compared to HDAC3 with an IC50 of about 8 m, and has little or no inhibitory effect against HDAC6 and HDAC8 (Hu et al., 2003). Two novel synthetic compounds, SK7041 and SK7068, preferentially target HDAC1 and 2 and exhibit growth inhibitory effects in human cancer cell lines and tumor xenograft models (Kim et al., 2003a). A small molecule, tubacin, selectively inhibits HDAC6 activity and causes an accumulation of acetylated -tubulin, but does not affect acetylation of histones, and does not inhibit cell cycle progression (Haggarty et al., 2003). No other HDACi for a specific HDAC has been reported.
HDACi selectively alters gene expression
HDACi-induced antitumor pathways
HDACi induces cell cycle arrest
HDACi activates the extrinsic apoptotic pathways
HDACi activates the intrinsic apoptotic pathways
HDACi induces mitotic cell death
HDACi induces autophagic cell death and senescence
ROS, thioredoxin and Trx binding protein 2 in HDACi-induced cell death
Antitumor effects of HDAC6 inhibition
Activation of protein phosphatase 1
Disruption of the function of chaperonin HSP90
Disruption of the aggresome pathway
HDACi inhibits angiogenesis
HDACi can block tumor angiogenesis by inhibition of hypoxia inducible factors (HIF) (Liang et al., 2006). HIF-1 and HIF-2 are transcription factors for angiogenic genes (Brown and Wilson, 2004). The oxygen level can control HIF activity through two mechanisms. First, under normoxic conditions, HIF-1 binds to von Hippel–Lindau protein (pVHL) and is degraded by the ubiquitination–proteasome system. Second, HIF activity depends on its transactivation potential (TAP), which is affected by the interaction with the coactivator p300/CBP among others. This complex can be disrupted by Factor Inhibiting HIF (FIH). Hypoxic conditions activate HIF through repression of the hydroxylases responsible for HIF degradation and loss of function.
Combination of HDACi with other antitumor agents
The HDACi have shown synergistic or additive antitumor effects with a wide range of antitumor reagents, including chemotherapeutic drugs, new targeted therapeutic reagents and radiation, by various mechanisms, some unique for particular combinations (Rosato and Grant, 2004; Bhalla, 2005; Marks and Dokmanovic, 2005; Bolden et al., 2006).
Clinical development of HDACi
At least 14 different HDACi are in some phase of clinical trials as monotherapy or in combination with retinoids, taxols, gemcitabine, radiation, etc, in patients with hematologic and solid tumors, including cancer of lung, breast, pancreas, renal and bladder, melanoma, glioblastoma, leukemias, lymphomas, multiple myeloma (see National Cancer Institute website for CTEP clinical trials, ctep.cancer.gov or clinicaltrials.gov, and website of companies developing HDACi; Table 2).
The resistance to HDACi
Conclusions and perspectives
HDACs have multiple substrates involved in many biological processes, including proliferation, differentiation, apoptosis and other forms of cell death. Indeed, the fact that HDACs have histone and multiple nonhistone protein substrates suggests these enzymes should be referred to as ‘lysine deacetylases’. HDACi can cause transformed cells to undergo growth arrest, differentiation and/or cell death. Normal cells are relatively resistant to HDACi. HDACi are selective in altering gene expression, which may reflect, in part, the proteins composing the transcription factor complex to which HDACs are recruited. Both altered gene expression and changes in non-histone proteins caused by HDACi-induced acetylation play a role in the antitumor activity of HDACi. This is reflected in the different inducer-activated antitumor pathways in transformed cells (Figure 1). The functions of HDACs are not redundant. Thus, a pan-HDAC inhibitor such as vorinostat may activate more antitumor pathways and have therapeutic advantages compared to HDAC isotype-specific inhibitors.
Almost all cancers have multiple defects in the expression and/or structure of proteins that regulate cell proliferation and death. Compared to other antitumor reagents, the plurality of action of HDACi potentially confers efficacy in a wide spectrum of cancers, which have heterogeneity and multiple defects, both among different types of cancer and within different individual tumors of the same type. The multiple defects in a cancer cell may be the reason for transformed cells being more sensitive than normal cells to HDACi. Thus, given the relatively rapid reversibility of vorinostat inhibition of HDACs, normal cells may be able to compensate for HDACi-induced changes more effectively than cancer cells.
HDACi have synergistic or additive antitumor effects with many other antitumor reagents – suggesting that combination of HDACi and other anticancer agents may be very attractive therapeutic strategies for using these agents. Complete understanding of the mechanisms underlying the resistance and sensitivity to HDACi has obvious therapeutic importance. Targeting resistant factors will enhance the antitumor efficacy of HDACi. Identifying markers that can predict response to HDACi is a high priority for expanding the efficacy of these novel anticancer agents.
The complexity of cancer has been known for almost a century, in large part from the seminal work of Otto Warburg in the 1920s using manometry, and following the work of Louis Pasteur 60 years earlier with fungi.
The volume of work and our unlocking of mitotic activity, apoptosis, mitochondria, and the cytoskeleton has taken us further into the cell interior, cell function, metabolic regulation, and pathophysiology. Despite the enormous contributions to our knowledge of genomics, there is a large body of work in pathways of cell function that resides in no small part in activity of protein catalysts and enzymes.
The work that has been described covers only cyclic dinucleotides and HDACi’s. Some of the activities described have relevance to microorganisms as well as cancer. As we have seen, blocking HDACs boosts the activity of regulatory T-cells. There are many specific functional alterations elucidated above.
The first presentation is of an antibody that blocks an immune suppressor called CTLA4, antibodies that block another immune suppressor, PD1. This also involves a protein in cells that responds to foreign DNA to launch an innate immune response — the first response of the body’s immune system. The protein, dubbed STING, is triggered by small molecules called cyclic di-nucleotides (CDN), and makes immune cells release interferon and other cytokines that activate disease-fighting T cells and stimulate the production of antibodies that together kill invading pathogens and destroy cancer cells. Listeria bacteria, for example, secrete a CDN directly into infected cells that activates STING.
The second is resident in acetylation status of histones and non-histone proteins is determined by histone deacetylases (HDACs) and histone acetyl-transferases (HATs). HATs add acetyl groups to lysine residues, while HDACs remove the acetyl groups. In general, acetylation of histone promotes a more relaxed chromatin structure, allowing transcriptional activation. HDACs can act as transcription repressors, due to histone deacetylation, and consequently promote chromatin condensation. HDAC inhibitors (HDACi) selectively alter gene transcription, in part, by chromatin remodeling and by changes in the structure of proteins in transcription factor complexes (Gui et al., 2004). The description focuses on the molecular mechanisms triggered by inhibitors of zinc-dependent HDACs in tumor cells that explain in part: (I) the effects of these compounds in inducing transformed cell death and (II) the relative resistance of normal and certain cancer cells to HDACi induced cell death.
HDACs have multiple substrates involved in many biological processes, including proliferation, differentiation, apoptosis and other forms of cell death. Indeed, the fact that HDACs have histone and multiple nonhistone protein substrates suggests these enzymes should be referred to as ‘lysine deacetylases’. HDACi can cause transformed cells to undergo growth arrest, differentiation and/or cell death. Normal cells are relatively resistant to HDACi. HDACi are selective in altering gene expression, which may reflect, in part, the proteins composing the transcription factor complex to which HDACs are recruited. Both altered gene expression and changes in non-histone proteins caused by HDACi-induced acetylation play a role in the antitumor activity of HDACi.
A little adversity builds character, or so the saying goes. True or not, the saying does seem an apt description of a developmental phenomenon that shapes gene expression. While it knows nothing of character, the gene expression apparatus appears to respond well to short-term mitochondrial stress that occurs early in development. In fact, transient stress seems to result in lasting benefits. These benefits, which include improved metabolic function and increased longevity, have been observed in both worms and mice, and may even occur—or be made to occur—in humans.
Gene expression is known to be subject to reprogramming by epigenetic modifiers, but such modifiers generally affect metabolism or lifespan, not both. A new set of epigenetic modifiers, however, has been found to trigger changes that do just that—both improve metabolism and extend lifespan.
Scientists based at the University of California, Berkeley, and the École Polytechnique Fédérale de Lausanne (EPFL) have discovered enzymes that are ramped up after mild stress during early development and continue to affect the expression of genes throughout the animal’s life. When the scientists looked at strains of inbred mice that have radically different lifespans, those with the longest lifespans had significantly higher expression of these enzymes than did the short-lived mice.
“Two of the enzymes we discovered are highly, highly correlated with lifespan; it is the biggest genetic correlation that has ever been found for lifespan in mice, and they’re both naturally occurring variants,” said Andrew Dillin, a UC Berkeley professor of molecular and cell biology. “Based on what we see in worms, boosting these enzymes could reprogram your metabolism to create better health, with a possible side effect of altering lifespan.”
Details of the work, which appeared online April 29 in the journal Cell, are presented in a pair of papers. One paper (“Two Conserved Histone Demethylases Regulate Mitochondrial Stress-Induced Longevity”) resulted from an effort led by Dillin and the EPFL’s Johan Auwerx. The other paper (“Mitochondrial Stress Induces Chromatin Reorganization to Promote Longevity and UPRmt”) resulted from an effort led by Dillin and his UC Berkeley colleague Barbara Meyer.
According to these papers, mitochondrial stress activates enzymes in the brain that affect DNA folding, exposing a segment of DNA that contains the 1500 genes involved in the work of the mitochondria. A second set of enzymes then tags these genes, affecting their activation for much or all of the lifetime of the animal and causing permanent changes in how the mitochondria generates energy.
The first set of enzymes—methylases, in particular LIN-65—add methyl groups to the DNA, which can silence promoters and thus suppress gene expression. By also opening up the mitochondrial genes, these methylases set the stage for the second set of enzymes—demethylases, in this case jmjd-1.2 and jmjd-3.1—to ramp up transcription of the mitochondrial genes. When the researchers artificially increased production of the demethylases in worms, all the worms lived longer, a result identical to what is observed after mitochondrial stress.
“By changing the epigenetic state, these enzymes are able to switch genes on and off,” Dillin noted. This happens only in the brain of the worm, however, in areas that sense hunger or satiety. “These genes are expressed in neurons that are sensing the nutritional status of the animal, and these signals emanate out to the periphery to change peripheral metabolism,” he continued.
When the scientists profiled enzymes in short- and long-lived mice, they found upregulation of these genes in the brains of long-lived mice, but not in other tissues or in the brains of short-lived mice. “These genes are expressed in the hypothalamus, exactly where, when you eat, the signals are generated that tell you that you are full. And when you are hungry, signals in that region tell you to go and eat,” Dillin explained said. “These genes are all involved in peripheral feedback.”
Among the mitochondrial genes activated by these enzymes are those involved in the body’s response to proteins that unfold, which is a sign of stress. Increased activity of the proteins that refold other proteins is another hallmark of longer life.
These observations suggest that the reversal of aging by epigenetic enzymes could also take place in humans.
“It seems that, while extreme metabolic stress can lead to problems later in life, mild stress early in development says to the body, ‘Whoa, things are a little bit off-kilter here, let’s try to repair this and make it better.’ These epigenetic switches keep this up for the rest of the animal’s life,” Dillin stated.
Two Conserved Histone Demethylases Regulate Mitochondrial Stress-Induced Longevity
Carsten Merkwirth6, Virginija Jovaisaite6, Jenni Durieux,…., Reuben J. Shaw, Johan Auwerx, Andrew Dillin
•H3K27 demethylases jmjd-1.2 and jmjd-3.1 are required for ETC-mediated longevity
•jmjd-1.2 and jmjd-3.1 extend lifespan and are sufficient for UPRmt activation
•UPRmt is required for increased lifespan due to jmjd-1.2 or jmjd-3.1 overexpression
•JMJD expression is correlated with UPRmt and murine lifespan in inbred BXD lines
Across eukaryotic species, mild mitochondrial stress can have beneficial effects on the lifespan of organisms. Mitochondrial dysfunction activates an unfolded protein response (UPRmt), a stress signaling mechanism designed to ensure mitochondrial homeostasis. Perturbation of mitochondria during larval development in C. elegans not only delays aging but also maintains UPRmt signaling, suggesting an epigenetic mechanism that modulates both longevity and mitochondrial proteostasis throughout life. We identify the conserved histone lysine demethylases jmjd-1.2/PHF8 and jmjd-3.1/JMJD3 as positive regulators of lifespan in response to mitochondrial dysfunction across species. Reduction of function of the demethylases potently suppresses longevity and UPRmt induction, while gain of function is sufficient to extend lifespan in a UPRmt-dependent manner. A systems genetics approach in the BXD mouse reference population further indicates conserved roles of the mammalian orthologs in longevity and UPRmt signaling. These findings illustrate an evolutionary conserved epigenetic mechanism that determines the rate of aging downstream of mitochondrial perturbations.
Mitochondrial Stress Induces Chromatin Reorganization to Promote Longevity and UPRmt
Ye Tian, Gilberto Garcia, Qian Bian, Kristan K. Steffen, Larry Joe, Suzanne Wolff, Barbara J. Meyer, Andrew Dillin
•LIN-65 accumulates in the nucleus in response to mitochondrial stress
•Mitochondrial stress-induced chromatin changes depend on MET-2 and LIN-65
•LIN-65 and DVE-1 exhibit interdependence in nuclear accumulation
•met-2 and atfs-1 act in parallel to affect mitochondrial stress-induced longevity
Organisms respond to mitochondrial stress through the upregulation of an array of protective genes, often perpetuating an early response to metabolic dysfunction across a lifetime. We find that mitochondrial stress causes widespread changes in chromatin structure through histone H3K9 di-methylation marks traditionally associated with gene silencing. Mitochondrial stress response activation requires the di-methylation of histone H3K9 through the activity of the histone methyltransferase met-2 and the nuclear co-factor lin-65. While globally the chromatin becomes silenced by these marks, remaining portions of the chromatin open up, at which point the binding of canonical stress responsive factors such as DVE-1 occurs. Thus, a metabolic stress response is established and propagated into adulthood of animals through specific epigenetic modifications that allow for selective gene expression and lifespan extension
Siddharta Mukherjee’s Writing Career Just Got Dealt a Sucker Punch
Siddharha Mukherjee won the 2011 Pulitzer Prize in non-fiction for his book, The Emperor of All Maladies. The book has received widespread acclaim among lay audience, physicians, and scientists alike. Last year the book was turned into a special PBS series. But, according to a slew of scientists, we should all be skeptical of his next book scheduled to hit book shelves this month, The Gene, An Intimate History.
Publishing an article on epigenetics in the New Yorker this week–perhaps a selection from his new book–Mukherjee has waltzed into one of the most active scientific debates in all of biology: that of gene regulation, or epigenetics.
Jerry Coyne, the evolutionary biologist known for keeping journalists honest, has published a two part critique of Mukherjee’s New Yorker piece. The first part–wildly tweeted yesterday–is a list of quotes from Coyne’s colleagues and those who have written in to the New Yorker, including two Nobel prize winners, Wally Gilbert and Sidney Altman, offering some very unfriendly sentences.
Wally Gilbert: “The New Yorker article is so wildly wrong that it defies rational analysis.”
Sidney Altman: “I am not aware that there is such a thing as an epigenetic code. It is unfortunate to inflict this article, without proper scientific review, on the audience of the New Yorker.”
The second part is a thorough scientific rebuttal of the Mukherjee piece. It all serves as a great drama about one of the most contested ideas in biology and also as a cautionary tale to journalists, even experienced writers such as Mukherjee, about the dangers of wading into scientific arguments. Readers may remember that a few years ago, science writer, David Dobbs, similarly skated into the same topic with his piece, Die, Selfish Gene, Die, and which raised a similar shitstorm, much of it from Coyne.
Mukherjee’s mistake is in giving credence to only one side of a very fierce debate–that the environment causes changes in the genome which can be passed on; another kind of evolution–as though it were settled science. Either Mukherjee, a physicisan coming off from a successful book and PBS miniseries on cancer, is setting himself up as a scientist, or he has been a truly naive science reporter. If he got this chapter so wrong, what does it mean about an entire book on the gene?
Coyne quotes one of his colleagues who raised some questions about the New Yorker’s science reporting, one particular question we’ve been asking here at Mendelspod. How do we know what we know? Does science now have an edge on any other discipline for being able to create knowledge?
Coyne’s colleague is troubled by science coverage in the New Yorker, and goes so far as to write that the New Yorker has been waging a “war on behalf of cultural critics and literary intellectuals against scientists and technologists.”
From my experience, it’s not quite that tidy. First of all, the New Yorker is the best writing I read each week. Period. Second, I haven’t found their science writing to have the slant claimed in the quote above. For example, most other mainstream outlets–including the New York Times with the Amy Harmon pieces–have given the anti-GMO crowd an equal say in the mistaken search for a “balance” on whether GMOs are harmful. (Remember John Stewart’s criticism of Fox News? That they give a false equivalent between two sides even when there is no equivalent on the other side?)
But the New Yorker has not fallen into this trap on GMOs and most of their pieces on the topic–mainly by Michael Specter–have been decidedly pro science and therefore decided pro GMO.
So what led Mukherjee to play scientist as well as journalist? There’s no question about whether I enjoy his prose. His writing beautifully whisks me away so that I don’t feel that I’m really working to understand. There is a poetic complexity that constantly brings different threads effortlessly together, weaving them into the same light. At one point he uses the metaphor of a web for the genome, with the epigenome being the stuff that sticks to the web. He borrows the metaphor from the Hindu notion of “being”, or jaal.
“Genes form the threads of the web; the detritus that adheres to it transforms every web into a singular being.”
There have been a few writers on Twitter defending Mukherjee’s piece. Tech Review’s Antonio Regalado called Coyne and his colleagues “tedious literalists” who have an “issue with epigenetic poetry.”
At his best, Mukherjee can take us down the sweet alleys of his metaphors and family stories with a new curiosity for the scientific truth. He can hold a mirror up to scientists, or put the spotlight on their work. At their worst, Coyne and his scientific colleagues can reek of a fear of language and therefore metaphor. The always outspoken scientist and author, Richard Dawkins, who made his name by personifying the gene, was quick to personify epigentics in a tweet: “It’s high time the 15 minutes of underserved fame for “epigenetics” came to an overdue end.” Dawkins is that rare scientist who has consistently been as comfortable with rhetoric and language as he is with data.
Hats off to Coyne who reminds us that a metaphor–however lovely–does not some science make. If Mukherjee wants to play scientist, let him create and gather data. If it’s the role of science journalist he wants, let him collect all the science he can before he begins to pour it into his poetry.
Same but Different
How epigenetics can blur the line between nature and nurture.
The author’s mother (right) and her twin are a study in difference and identity. CREDIT: PHOTOGRAPH BY DAYANITA SINGH FOR THE NEW YORKER
October 6, 1942, my mother was born twice in Delhi. Bulu, her identical twin, came first, placid and beautiful. My mother, Tulu, emerged several minutes later, squirming and squalling. The midwife must have known enough about infants to recognize that the beautiful are often the damned: the quiet twin, on the edge of listlessness, was severely undernourished and had to be swaddled in blankets and revived.
The first few days of my aunt’s life were the most tenuous. She could not suckle at the breast, the story runs, and there were no infant bottles to be found in Delhi in the forties, so she was fed through a cotton wick dipped in milk, and then from a cowrie shell shaped like a spoon. When the breast milk began to run dry, at seven months, my mother was quickly weaned so that her sister could have the last remnants.
Tulu and Bulu grew up looking strikingly similar: they had the same freckled skin, almond-shaped face, and high cheekbones, unusual among Bengalis, and a slight downward tilt of the outer edge of the eye, something that Italian painters used to make Madonnas exude a mysterious empathy. They shared an inner language, as so often happens with twins; they had jokes that only the other twin understood. They even smelled the same: when I was four or five and Bulu came to visit us, my mother, in a bait-and-switch trick that amused her endlessly, would send her sister to put me to bed; eventually, searching in the half-light for identity and difference—for the precise map of freckles on her face—I would realize that I had been fooled.
But the differences were striking, too. My mother was boisterous. She had a mercurial temper that rose fast and died suddenly, like a gust of wind in a tunnel. Bulu was physically timid yet intellectually more adventurous. Her mind was more agile, her tongue sharper, her wit more lancing. Tulu was gregarious. She made friends easily. She was impervious to insults. Bulu was reserved, quieter, and more brittle. Tulu liked theatre and dancing. Bulu was a poet, a writer, a dreamer.
….. more
Why are identical twins alike? In the late nineteen-seventies, a team of scientists in Minnesota set out to determine how much these similarities arose from genes, rather than environments—from “nature,” rather than “nurture.” Scouring thousands of adoption records and news clips, the researchers gleaned a rare cohort of fifty-six identical twins who had been separated at birth. Reared in different families and different cities, often in vastly dissimilar circumstances, these twins shared only their genomes. Yet on tests designed to measure personality, attitudes, temperaments, and anxieties, they converged astonishingly. Social and political attitudes were powerfully correlated: liberals clustered with liberals, and orthodoxy was twinned with orthodoxy. The same went for religiosity (or its absence), even for the ability to be transported by an aesthetic experience. Two brothers, separated by geographic and economic continents, might be brought to tears by the same Chopin nocturne, as if responding to some subtle, common chord struck by their genomes.
One pair of twins both suffered crippling migraines, owned dogs that they had named Toy, married women named Linda, and had sons named James Allan (although one spelled the middle name with a single “l”). Another pair—one brought up Jewish, in Trinidad, and the other Catholic, in Nazi Germany, where he joined the Hitler Youth—wore blue shirts with epaulets and four pockets, and shared peculiar obsessive behaviors, such as flushing the toilet before using it. Both had invented fake sneezes to diffuse tense moments. Two sisters—separated long before the development of language—had invented the same word to describe the way they scrunched up their noses: “squidging.” Another pair confessed that they had been haunted by nightmares of being suffocated by various metallic objects—doorknobs, fishhooks, and the like.
The Minnesota twin study raised questions about the depth and pervasiveness of qualities specified by genes: Where in the genome, exactly, might one find the locus of recurrent nightmares or of fake sneezes? Yet it provoked an equally puzzling converse question: Why are identical twins different? Because, you might answer, fate impinges differently on their bodies. One twin falls down the crumbling stairs of her Calcutta house and breaks her ankle; the other scalds her thigh on a tipped cup of coffee in a European station. Each acquires the wounds, calluses, and memories of chance and fate. But how are these changes recorded, so that they persist over the years? We know that the genome can manufacture identity; the trickier question is how it gives rise to difference.
….. more
But what turns those genes on and off, and keeps them turned on or off? Why doesn’t a liver cell wake up one morning and find itself transformed into a neuron? Allis unpacked the problem further: suppose he could find an organism with two distinct sets of genes—an active set and an inactive set—between which it regularly toggled. If he could identify the molecular switches that maintain one state, or toggle between the two states, he might be able to identify the mechanism responsible for cellular memory. “What I really needed, then, was a cell with these properties,” he recalled when we spoke at his office a few weeks ago. “Two sets of genes, turned ‘on’ or ‘off’ by some signal.”
more…
“Histones had been known as part of the inner scaffold for DNA for decades,” Allis went on. “But most biologists thought of these proteins merely as packaging, or stuffing, for genes.” When Allis gave scientific seminars in the early nineties, he recalled, skeptics asked him why he was so obsessed with the packing material, the stuff in between the DNA. …. A skein of silk tangled into a ball has very different properties from that same skein extended; might the coiling or uncoiling of DNA change the activity of genes?
In 1996, Allis and his research group deepened this theory with a seminal discovery. “We became interested in the process of histone modification,” he said. “What is the signal that changes the structure of the histone so that DNA can be packed into such radically different states? We finally found a protein that makes a specific chemical change in the histone, possibly forcing the DNA coil to open. And when we studied the properties of this protein it became quite clear that it was also changing the activity of genes.” The coils of DNA seemed to open and close in response to histone modifications—inhaling, exhaling, inhaling, like life.
Allis walked me to his lab, a fluorescent-lit space overlooking the East River, divided by wide, polished-stone benches. A mechanical stirrer, whirring in a corner, clinked on the edge of a glass beaker. “Two features of histone modifications are notable,” Allis said. “First, changing histones can change the activity of a gene without affecting the sequence of the DNA.” It is, in short, formally epi-genetic, just as Waddington had imagined. “And, second, the histone modifications are passed from a parent cell to its daughter cells when cells divide. A cell can thus record ‘memory,’ and not just for itself but for all its daughter cells.”
…..
The New Yorker screws up big time with science: researchers criticize the Mukherjee piece on epigenetics
Abstract: This is a two part-post about a science piece on gene regulation that just appeared in the New Yorker. Today I give quotes from scientists criticizing that piece; tomorrow I’ll present a semi-formal critique of the piece by two experts in the field.
esterday I gave readers an assignment: read the new New Yorkerpiece by Siddhartha Mukherjee about epigenetics. The piece, called “Same but different” (subtitle: “How epigenetics can blur the line between nature and nurture”) was brought to my attention by two readers, both of whom praised it. Mukherjee, a physician, is well known for writing the Pulitzer-Prize-winning book (2011) The Emperor of All Maladies: A Biography of Cancer. (I haven’t read it yet, but it’s on my list.) Mukherjee has a new book that will be published in May: The Gene: An Intimate History. As I haven’t seen it, the New Yorker piece may be an excerpt from this book.
Everyone I know who has read The Emperor of All Maladies gives it high praise. I wish I could say the same for Mukherjee’s New Yorker piece. When I read it at the behest of the two readers, I found his analysis of gene regulation incomplete and superficial. Although I’m not an expert in that area, I knew that there was a lot of evidence that regulatory proteins called “transcription factors”, and not “epigenetic markers” (see discussion of this term tomorrow) or modified histones—the factors emphasized by Mukherjee—played hugely important roles in gene regulation. The speculations at the end of the piece about “Lamarckian evolution” via environmentally induced epigenetic changes in the genome were also unfounded, for we have no evidence for that kind of adaptive evolution. Mukherjee does, however, mention that lack of evidence, though I wish he’d done so more strongly given that environmental modification of DNA bases is constantly touted as an important and neglected factor in evolution.
Unbeknownst to me, there was a bit of a kerfuffle going on in the community of scientists who study gene regulation, with many of them finding serious mistakes and omissions in Mukherjee’s piece. There appears to have been some back-and-forth emailing among them, and several wrote letters to the New Yorker, urging them to correct the misconceptions, omissions, and scientific errors in “Same but different.” As I understand it, both Mukherjee and the New Yorker simply batted these criticisms away, and, as far as I know, will not publish any corrections. So today and tomorrow I’ll present the criticisms here, just so they’ll be on the record.
Because Mukherjee writes very well, and because even educated laypeople won’t know the story of gene regulation revealed over the last few decades, they may not see the big lacunae in his piece. It is, then, important to set matters straight, for at least we should know what science has told us about how genes are turned on and off. The criticism of Mukherjee’s piece, coming from scientists who really are experts in gene regulation, shows a lack of care on the part of Mukherjee and theNew Yorker: both a superficial and misleading treatment of the state of the science, and a failure of the magazine to properly vet this piece (I have no idea whether they had it “refereed” not just by editors but by scientists not mentioned in the piece).
Let me add one thing about science and the New Yorker. I believe I’ve said this before, but the way the New Yorker treats science is symptomatic of the “two cultures” problem. This is summarized in an email sent me a while back by a colleague, which I quote with permission:
The New Yorker is fine with science that either serves a literary purpose (doctors’ portraits of interesting patients) or a political purpose (environmental writing with its implicit critique of modern technology and capitalism). But the subtext of most of its coverage (there are exceptions) is that scientists are just a self-interested tribe with their own narrative and no claim to finding the truth, and that science must concede the supremacy of literary culture when it comes to anything human, and never try to submit human affairs to quantification or consilience with biology. Because the magazine is undoubtedly sophisticated in its writing and editing they don’t flaunt their postmodernism or their literary-intellectual proprietariness, but once you notice it you can make sense of a lot of their material.
. . . Obviously there are exceptions – Atul Gawande is consistently superb – but as soon as you notice it, their guild war on behalf of cultural critics and literary intellectuals against scientists, technologists, and analytic scholars becomes apparent.
…. more
Researchers criticize the Mukherjee piece on epigenetics: Part 2
Trigger warning: Long science post!
Yesterday I provided a bunch of scientists’ reactions—and these were big names in the field of gene regulation—to Siddhartha Mukherjee’s ill-informed piece in The New Yorker, “Same but different” (subtitle: “How epigenetics can blur the line between nature and nurture”). Today, in part 2, I provide a sentence-by-sentence analysis and reaction by two renowned researchers in that area. We’ll start with a set of definitions (provided by the authors) that we need to understand the debate, and then proceed to the critique.
Let me add one thing to avoid confusion: everything below the line, including the definition (except for my one comment at the end) was written by Ptashne and Greally.
by Mark Ptashne and John Greally
Introduction
Ptashne is The Ludwig Professor of Molecular Biology at the Memorial Sloan Kettering Cancer Center in New York. He wrote A Genetic Switch, now in its third edition, which describes the principles of gene regulation and the workings of a ‘switch’; and, with Alex Gann, Genes and Signals, which extends these principles and ideas to higher organisms and to other cellular processes as well. John Greally is the Director of the Center for Epigenomics at the Albert Einstein College of Medicine in New York.
The New Yorker (May 2, 2016) published an article entitled “Same But Different” written by Siddhartha Mukherjee. As readers will have gathered from the letters posted yesterday, there is a concern that the article is misleading, especially for a non-scientific audience. The issue concerns our current understanding of “gene regulation” and how that understanding has been arrived at.
First some definitions/concepts:
Gene regulation refers to the “turning on and off of genes”. The primary event in turning a gene “on” is to transcribe (copy) it into messenger RNA (mRNA). That mRNA is then decoded, usually, into a specific protein. Genes are transcribed by the enzyme called RNA polymerase.
Development: the process in which a fertilized egg (e.g., a human egg) divides many times and eventually forms an organism. During this process, many of the roughly 23,000 genes of a human are turned “on” or “off” in different combinations, at different times and places in the developing organism. The process produces many different cell types in different organs (e.g. liver and brain), but all retain the original set of genes.
Transcription factors: proteins that bind to specific DNA sequences near specific genes and turn transcription of those genes on and off. A transcriptional ‘activator’, for example, bears two surfaces: one binds a specific sequence in DNA, and the other binds to, and thereby recruits to the gene, protein complexes that include RNA polymerase. It is widely acknowledged that the identity of a cell in the body depends on the array of transcription factors present in the cell, and the cell’s history. RNA molecules can also recognize specific genomic sequences, and they too sometimes work as regulators. Neither transcription factors nor these kinds of RNA molecules – the fundamental regulators of gene expression and development – are mentioned in the New Yorker article.
Signals: these come in many forms (small molecules like estrogen, larger molecules (often proteins such as cytokines) that determine the ability of transcription factors to work. For example, estrogen binds directly to a transcription factor (the estrogen receptor) and, by changing its shape, permits it to bind DNA and activate transcription.
“Memory”: a dividing cell can (often does) produce daughters that are identical, and that express identical genes as does the mother cell. This occurs because the transcription factors present in the mother cell are passively transmitted to the daughters as the cell divides, and they go to work in their new contexts as before. To make two different daughters, the cell must distribute its transcription factors asymmetrically.
PositiveFeedback: An activator can maintain its own expression by positive feedback. This requires, simply, that a copy of the DNA sequence to which the activator binds is present near its own gene. Expression of the activator then becomes self-perpetuating. The activator (of which there now are many copies in the cell) activates other target genes as it maintains its own expression. This kind of ‘memory circuit’, first described in bacteria, is found in higher organisms as well. Positive feedback can explain how a fully differentiated cell (that is, a cell that has reached its developmental endpoint) maintains its identity.
Nucleosomes: DNA in higher organisms (eukaryotes) is wrapped, like beads on a string, around certain proteins (called histones), to form nucleosomes. The histones are subject to enzymatic modifications: e.g., acetyl, methyl, phosphate, etc. groups can be added to these structures. In bacteria there are no nucleosomes, and the DNA is more or less ‘naked’.
“Epigenetic modifications”: please don’t worry about the word ”epigenetic”; it is misused in any case. What Mukherjee refers to by this term are the histone modifications mentioned above, and a modification to DNA itself: the addition of methyl groups. Keep in mind that the organisms that have taught us the most about development – flies (Drosophila) and worms (C. elegans)—do not have the enzymes required for DNA methylation. That does not mean that DNA methylation cannot do interesting things in humans, for example, but it is obviously not at the heart of gene regulation.
Specificity Development requires the highly specific sequential turning on and off of sets of genes. Transcription factors and RNA supply this specificity, but enzymes that impart modifications to histones cannot: every nucleosome (and hence every gene) appears the same to the enzyme. Thus such enzymes cannot pick out particular nucleosomes associated with particular genes to modify. Histone modifications might be imagined to convey ‘memory’ as cells divide – but there are no convincing indications that this happens, nor are there molecular models that might explain why they would have the imputed effects.
Analysis and critique of Mukherjee’s article
The picture we have just sketched has taken the combined efforts of many scientists over 50 years to develop. So what, then, is the problem with the New Yorker article?
There are two: first, the picture we have just sketched, emphasizing the primary role of transcription factors and RNA, is absent. Second, that picture is replaced by highly dubious speculations, some of which don’t make sense, and none of which has been shown to work as imagined in the article.
(Quotes from the Mukherjee article are indented and in plain text; they are followed by comments, flush left and in bold, by Ptashne and Greally.)
In 1978, having obtained a Ph.D. in biology at Indiana University, Allis began to tackle a problem that had long troubled geneticists and cell biologists: if all the cells in the body have the same genome, how does one become a nerve cell, say, and another a blood cell, which looks and functions very differently?
The problems referred to were recognized long before 1978. In fact, these were exactly the problems that the great French scientists François Jacob and Jacques Monod took on in the 1950s-60s. In a series of brilliant experiments, Jacob and Monod showed that in bacteria, certain genes encode products that regulate (turn on and off) specific other genes. Those regulatory molecules turned out to be proteins, some of which respond to signals from the environment. Much of the story of modern biology has been figuring out how these proteins – in bacteria and in higher organisms – bind to and regulate specific genes. Of note is that in higher organisms, the regulatory proteins look and act like those in bacteria, despite the fact that eukaryotic DNA is wrapped in nucleosomes whereas bacterial DNA is not. We have also learned that certain RNA molecules can play a regulatory role, a phenomenon made possible by the fact that RNA molecules, like regulatory proteins, can recognize specific genomic sequences.
In the nineteen-forties, Conrad Waddington, an English embryologist, had proposed an ingenious answer: cells acquired their identities just as humans do—by letting nurture (environmental signals) modify nature (genes). For that to happen, Waddington concluded, an additional layer of information must exist within a cell—a layer that hovered, ghostlike, above the genome. This layer would carry the “memory” of the cell, recording its past and establishing its future, marking its identity and its destiny but permitting that identity to be changed, if needed. He termed the phenomenon “epigenetics”—“above genetics.”
This description greatly misrepresents the original concept. Waddington argued that development proceeds not by the loss (or gain) of genes, which would be a “genetic” process, but rather that some genes would be selectively expressed in specific and complex cellular patterns as development proceeds. He referred to this intersection of embryology (then called “epigenesis”) and genetics as “epigenetic”.We now understand that regulatory proteins work in combinations to turn on and off genes, including their own genes, and that sometimes the regulatory proteins respond to signals sent by other cells. It should be emphasized that Waddington never proposed any “ghost-like” layer of additional information hovering above the gene. This is a later misinterpretation of a literal translation of the term epigenetics, with “epi-“ meaning “above/upon” the genetic information encoded in DNA sequence. Unfortunately, this new and pervasive definition encompasses all of transcriptional regulation and is of no practical value.
…..more
By 2000, Allis and his colleagues around the world had identified a gamut of proteins that could modify histones, and so modulate the activity of genes. Other systems, too, that could scratch different kinds of code on the genome were identified (some of these discoveries predating the identification of histone modifications). One involved the addition of a chemical side chain, called a methyl group, to DNA. The methyl groups hang off the DNA string like Christmas ornaments, and specific proteins add and remove the ornaments, in effect “decorating” the genome. The most heavily methylated parts of the genome tend to be dampened in their activity.
It is true that enzymes that modify histones have been found—lots of them. A striking problem is that, after all this time, it is not at all clear what the vast majority of these modifications do. When these enzymatic activities are eliminated by mutation of their active sites (a task substantially easier to accomplish in yeast than in higher organisms) they mostly have little or no effect on transcription. It is not even clear that histones are the biologically relevant substrates of most of these enzymes.
In the ensuing decade, Allis wrote enormous, magisterial papers in which a rich cast of histone-modifying proteins appear and reappear through various roles, mapping out a hatchwork of complexity. . . These protein systems, overlaying information on the genome, interacted with one another, reinforcing or attenuating their signals. Together, they generated the bewildering intricacy necessary for a cell to build a constellation of other cells out of the same genes, and for the cells to add “memories” to their genomes and transmit these memories to their progeny. “There’s an epigenetic code, just like there’s a genetic code,” Allis said. “There are codes to make parts of the genome more active, and codes to make them inactive.”
By ‘epigenetic code’ the author seems to mean specific arrays of nucleosome modifications, imparted over time and cell divisions, marking genes for expression. This idea has been tested in many experiments and has been found not to hold.
….. and more
Larry H. Bernstein, MD, FCAP
I hope that this piece brings greater clarity to the discussion. I have heard the use of the term “epigenetics” for over a decade. The term was never so clear. I think that the New Yorker article was a reasonable article for the intended audience. It was not intended to clarify debates about a mechanism for epigenetic based changes in evolutionary science. I think it actually punctures the “classic model” of the cell depending only on double stranded DNA and transcription, which deflates our concept of the living cell. The concept of epigenetics was never really formulated as far as I have seen, and I have done serious work in enzymology and proteins at a time that we did not have the technology that exists today. I have considered with the critics that protein folding, protein misfolding, protein interactions with proximity of polar and nonpolar groups, and the regulatory role of microRNAs that are not involved in translation, and the evolving concept of what is “dark (noncoding) DNA” lend credence to the complexity of this discussion. Even more interesting is the fact that enzymes (and isoforms of enzymes) have a huge role in cellular metabolic differences and in the function of metabolic pathways. What is less understood is the extremely fast reactions involved in these cellular reactions. These reactions are in my view critical drivers. This is brought out by Erwin Schroedinger in the book What is Life? which infers that there can be no mathematical expression of life processes.








 Article Info
Article Info






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
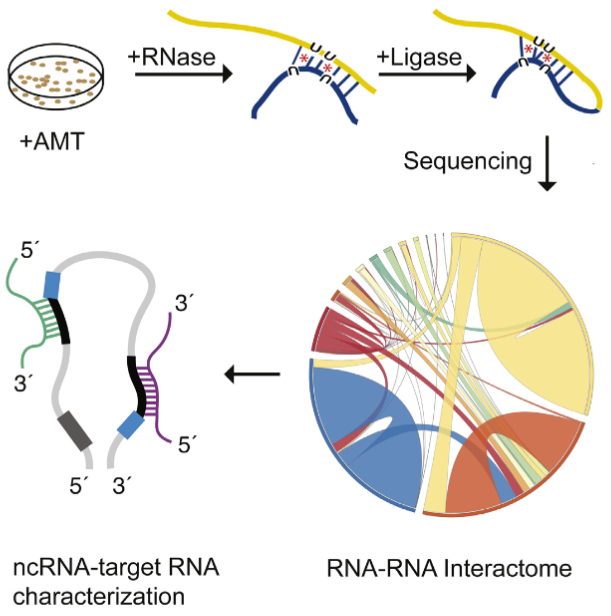{kind=link}
{kind=link}
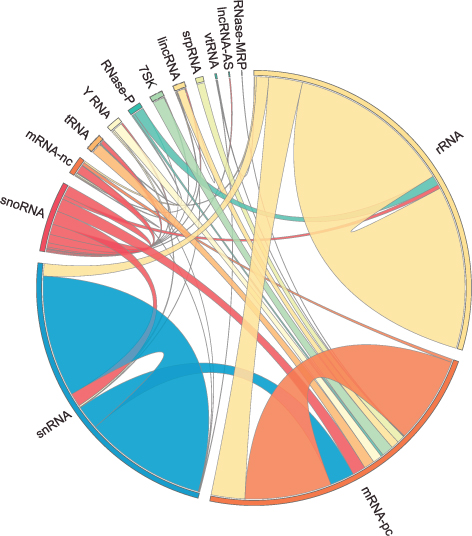{kind=link}
{kind=link}
{kind=link}
{kind=link}
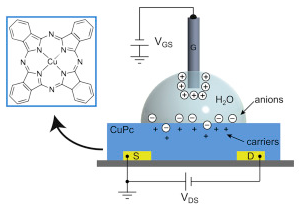{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}