Lipids link to breast cancer
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Lipids Found Critical to Breast Cancer Cell Proliferation
http://www.genengnews.com/gen-news-highlights/lipids-found-critical-to-breast-cancer-cell-proliferation/81252572/

http://www.genengnews.com/Media/images/GENHighlight/112523_webApr6_2016_IRBBarcelona_BreastTumorsLIPD3119618425.jpg
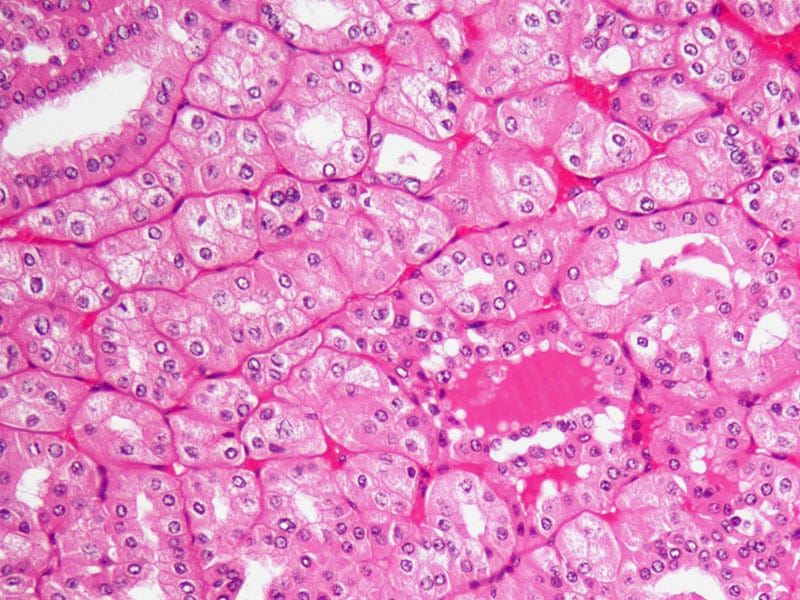
Scientists in Spain report finding that breast cancer cells need to take up lipids from the extracellular environment so that they can continue to proliferate. The main protein involved in this process is LIPG, an enzyme found in the cell membrane and without which tumor cell growth is arrested. Analyses of more than 500 clinical samples from patients with various kinds of breast tumors reveal that 85% have high levels of LIPG expression.
The research (“FoxA and LIPG Endothelial Lipase Control the Uptake of Extracellular Lipids for Breast Cancer Growth”) is published in Nature Communications.
In Spain, breast cancer is the most common tumor in women and the fourth most common type in both sexes (data from the Spanish Society of Medical Oncology, 2012), registering more than 25,000 new diagnoses each year. According to figures from the World Health Organization, every year 1.38 million new cases of breast cancer are diagnosed and 458,000 people die from this disease (International Agency for Research on Cancer Globocan, 2008).
It was already known that cancer cells require extracellular glucose to grow and that they reprogram their internal machinery to produce greater amounts of lipids. The relevance of this study is that it reveals for the first time that tumor cells must import extracellular lipids to grow.
“This new knowledge related to metabolism could be the Achilles heel of breast cancer,” explains ICREA researcher and Institute for Research in Biomedicine–Barcelona group leader Roger Gomis, Ph.D., co-leader of the study together with Joan J. Guinovart, Ph.D., director of IRB Barcelona and professor at the University of Barcelona. Using animal models and cancer cell cultures, the scientists have demonstrated that blocking of LIPG activity arrests tumor growth.
“What is promising about this new therapeutic target is that LIPG function does not appear to be indispensable for life, so its inhibition may have fewer side effects than other treatments,” explains the first author of the study, Felipe Slebe, a Ph.D. Fellow at IRB Barcelona.
According to Dr. Guinovart, “because LIPG is a membrane protein, it is potentially easier to design a pharmacological agent to block its activity.”
“If a drug were found to block its activity, it could be used to develop more efficient chemotherapy treatments that are less toxic than those currently available,” adds Dr. Gomis.
The scientists are now looking into international collaborations for developing LIPG inhibitors.
FoxA and LIPG endothelial lipase control the uptake of extracellular lipids for breast cancer growth
Felipe Slebe, Federico Rojo, Maria Vinaixa,…, Joan Albanell, Joan J. Guinovart & Roger R. Gomis
Nature Communications7,Article number:11199 http://dx.doi.org:/10.1038/ncomms11199
The mechanisms that allow breast cancer (BCa) cells to metabolically sustain rapid growth are poorly understood. Here we report that BCa cells are dependent on a mechanism to supply precursors for intracellular lipid production derived from extracellular sources and that the endothelial lipase (LIPG) fulfils this function. LIPG expression allows the import of lipid precursors, thereby contributing to BCa proliferation. LIPG stands out as an essential component of the lipid metabolic adaptations that BCa cells, and not normal tissue, must undergo to support high proliferation rates. LIPG is ubiquitously and highly expressed under the control of FoxA1 or FoxA2 in all BCa subtypes. The downregulation of either LIPG or FoxA in transformed cells results in decreased proliferation and impaired synthesis of intracellular lipids.
FoxA1 and FoxA2 in BCa growth
The importance of FoxA1 in BCa cells differentiation and its contribution to controlling the expression of metabolic genes in several other tissues makes this transcription factor a highly attractive target to explain the metabolic alterations reported in BCa. For these reason, we decided to ascertain the metabolic processes controlled by FoxA1 in BCa. We first confirmed the association between high FoxA1 expression (mRNA and protein) and luminal subtype (Fig. 1a). To this end, we used two cohorts of primary breast tumours with annotated clinical features and follow-up. The MSKCC/EMC BCa data set is based on gene expression profiles from an original series of 560 cases10, whereas the Spanish BCa data set (n=439) is a tissue microarray of formalin-fixed paraffin-embedded stage I–III breast tumour specimens11 (details provided in Methods Section). High FoxA1 gene expression significantly correlated with high expression of well-established luminal markers, such as GATA3 and ESR1, in primary tumours (Supplementary Fig. 1a). Next we explored FoxA1 expression beyond the luminal subtype. Lower FoxA1 expression was observed in non-luminal tumours (Fig. 1a,b); however, a subset also expressed higher FoxA1 levels (Supplementary Fig. 1b and Supplementary Table 1). Given that FoxA2, in conjunction with FoxA1, is also involved in the regulation of several metabolic pathways, we determined the expression of this factor in BCa samples. Unfortunately, no FoxA2 probes in the Affymetrix platform used in the MSKCC/EMC data set provided a reliable interpretation. To overcome this limitation, we used tissue arrays of early BCa samples (Spanish BCa set). Histological examination of FoxA2-stained tissue microarray slides from the Spanish BCa set revealed the expression of this factor in six non-luminal samples, which were scored as FoxA1− (examples in Fig. 1b and summarized inSupplementary Table 1). Collectively, the number of FoxA+ BCa samples detected by immunohistochemistry accounted for 81.3% of all samples in the Spanish BCa set (Supplementary Table 1), which represent a significant proportion of BCa and point to the participation of FoxA in this disease, beyond to its involvement in differentiation and control of hormonal responses.
Figure 1: FoxA1 and FoxA2 in BCa growth.
http://www.nature.com/ncomms/2016/160405/ncomms11199/images_article/ncomms11199-f1.jpg
(a, top) FoxA1 mRNA expression in the MSKCC/EMC set. BCa samples were stratified in Luminal A, Luminal B, Her2, triple negative and unknown subgroups. The unknown group represents specimens that were not classified in any group. (bottom) FoxA1 protein levels by IHC staining in Luminal, Her2 and triple negative samples in the Spanish BCa set (cohort of 439 BCa patients). Data is average±s.d. (b) FoxA1 and FoxA2 IHC staining in FFPE human specimens representative of the different BCa subtypes. Six independent cases are depicted. FoxA1 and FoxA2 are expressed mainly in the nuclei of tumour cells. Scale bar, 50 μm. (c) FoxA1 and FoxA2 mRNA expression analysis by qRT-PCR and protein expression by western blot in human BCa cell lines compared with HMECs. T-test was used. Data are average±s.e.m.; n= 3. Of note, MDA435 are of melanoma origin. (d) FoxA1 and FoxA2 expression in MCF7, MDA231 and their derivatives cells by qRT-PCR and western blot. FoxA1 and FoxA2 depletion was achieved with a doxycycline-inducible short hairpin vector. FoxA-depleted cells were rescued by expression of FoxA2 in MCF7 cells or FoxA1 in MDA231 cells. Cell populations were cultured in the presence or absence of doxycycline for 6 days. P value is the result of T-test. Data are average±s.e.m.;n=3. *P≤0.05, ***P≤0.001 (e, left) Schematic representation of MDA231 and MCF7 cells grown without doxycycline and inoculated in Balb/c nude mice treated with or without doxycycline to induce the expression of the indicated FoxA short hairpins. All tumour cell lines have GFP constitutive expression, and tRFP concomitantly with the short hairpin were expressed in doxycycline treated tumours. (right) Tumour growth of the indicated cell populations inoculated in Balb/c nude mice are determined at the indicated time points. P value is the result of T-test. Data are average±s.e.m.; n= 5–8 tumours. *P≤0.05,**P≤0.01, ***P≤0.001. FFPE, formalin-fixed paraffin-embedded.
Next, we extended our analysis to BCa cell lines for further mechanistic studies. We compared FoxA1 and FoxA2 mRNA expression in four estrogen receptor positive (ER+) (MCF7, T47D, BT474 and ZR75) and four estrogen receptor negative (ER−) (SKBR3, MDA468, BT20 and MDA231) BCa cell lines, a cell line of melanoma origin (MDA435), and human mammary epithelial cells (HMECs). Of note, two of the BCa lines tested were HER2+ (BT474 and SKBR3) (Fig. 1c). All ER+ BCa cells (MCF7, T47D, BT474 and ZR75), the ER−/HER2+ SKBR3 and both triple negative-like MDA468 and BT20 cell lines expressed FoxA1. Interestingly, MDA231 triple negative-like cells expressed high levels of FoxA2 but not FoxA1, and the non-tumour HMECs did not express these factors (Fig. 1c). No BCa cells co-expressed these two proteins (Fig. 1c). Our results suggest that the expression of FoxA transcription factors is a common feature of breast tumours, as well as of BCa cell lines. This notion implies that FoxA factors play a major role in BCa growth, independently of luminal fate specification.
To examine the molecular basis of the contribution of FoxA1 and FoxA2 to BCa growth, we engineered constitutive GFP-luciferase-expressing MCF7 and MDA231 cells with a doxycycline-inducible short-hairpin RNA (sh-RNA) vector targeting either FoxA1 or FoxA2. Doxycycline addition to the cell culture media decreased FoxA expression in both cell lines compared with control cells (ShControl (Dox+) and Sh FoxA1 or Sh FoxA2 (Dox−))(Fig. 1d), with the concomitant expression of tRFP (Supplementary Fig. 1c). Of note, there was no gain of expression of FoxA2 in FoxA1-depleted cells or vice versa (Fig. 1d). Interestingly, cancer cell proliferation was impaired in vitroupon depletion of either FoxA1 or FoxA2 in MCF7 and MDA231 cells, respectively (Supplementary Fig. 1d,e). Similarly, when Balb/c nude mice implanted with xenograft tumours from the above described cellular populations were treated with doxycycline and the short hairpins were induced, striking differences in tumour growth were observed. FoxA1-depleted MCF7 and FoxA2-depleted MDA231 tumour growth was blunted (Fig. 1e and additional controls in Supplementary Fig. 1f. Experimental details in the Supplementary Methods Section). Collectively, these observations confirm that FoxA1 or FoxA2 expression is required for BCa growth.
Previous studies indicate that FoxA1 and FoxA2 transcriptionally regulate common genes in the liver and pancreas that are central to development and metabolism. We therefore hypothesized that crossed expression of FoxA factors could rescue tumour growth by restoring the expression of essential metabolic genes. To this end, we engineered doxycycline-driven shFoxA1 MCF7 cells to express exogenous FoxA2 and doxycycline-driven shFoxA2 MDA231 cells to express exogenous FoxA1 (Fig. 1d). Interestingly, when these BCa modified cells were implanted in Balb/c nude mice and FoxA depletion was induced with doxycycline, the sustained expression of another FoxA factor (FoxA2 in MCF7 and FoxA1 in MDA231 cells) was sufficient for tumours to continuously grow (Fig. 1e and additional controls in Supplementary Fig. 1f). Quantitative real-time PCR (qRT-PCR) analysis confirmed FoxA expression in the distinct tumour populations ex-vivo (Supplementary Fig. 1g). These results showed that retention of minimal levels of FoxA1 or FoxA2 expression is necessary for BCa cell growth.
FoxA1- and FoxA2-regulated transcripts for BCa growth
Figure 2: A genomic approach to identify FoxA1- and FoxA2-regulated transcripts in MCF7 and MDA231 cells.
http://www.nature.com/ncomms/2016/160405/ncomms11199/images_article/ncomms11199-f2.jpg
(a) FACS profiling of MCF7 and MDA231 cells derived from tumours isolated from mice on the basis of the expression of GFP+ and RFP− (control group) or GFP+ and tRFP+ (knockdown and rescue groups). (b) Representation of the transcripts up- and downregulated by FoxA in MCF7 and MDA231 cells isolated from tumours. Up- and downregulated transcripts present a Bayesian false discovery rate below 5% and fold change >2.5. (c) LIPG, Bcl2 and Cdh11OB mRNA levels of the indicated genetically modified MCF7 and MDA231 tumour xenografts analysed by qRT-PCR. P value is the result of T-test. Data are average±s.e.m.; n= 5–8 tumours. *P≤0.05, ***P≤0.001. (d) LIPG protein expression in constitutive shFoxA1 MCF7 or shFoxA2 MDA231 cells. (e) Promoter reporter assay in HEK 293 cells. Cells were transfected with LIPG promoter reporter and FoxA1 or FoxA2 expressing vectors when indicated. P value is the result of T-test. Data are average±s.e.m.; n=3. ****P≤0.0001.
LIPG expression in BCa
Next, we showed that LIPG expression in primary tumours was specific to BCa tumour cells and not to other stroma cellular entities (Fig. 3a). Subsequently, we tested LIPG expression in normal breast epithelia and interrogated 20 samples from mammoplasty reductions. Normal breast epithelial cells showed a lower expression of LIPG than cells from tumour specimens (Fig. 3b). Similar results were obtained for LIPG protein levels in a panel from BCa lines compared with HMEC cells. Of the cellular populations tested, the eight BCa cell lines expressing FoxA1 or FoxA2 had very high levels of LIPG protein compared with the melanoma MDA435 cell line and the human epithelial cell (Fig. 3c). Consistent with this observation, 83.8% of BCa samples in the Spanish tumour cohort were LIPG+ (Fig. 3d and Supplementary Table 3), and LIPG expression correlated with FoxA expression (Spearman correlation; r=0.477, P=0.000001; Fig. 3e). Further analysis showed that LIPG expression levels in primary tumours do not have the capacity to stratify patients for differential risk of overall or disease-free survival (Supplementary Fig. 2a) and are not dependent on estrogen signalling (Supplementary Fig. 2b), thus reinforcing the notion that LIPG is essential for BCa growth.
Figure 3: LIPG contributes to BCa growth.
http://www.nature.com/ncomms/2016/160405/ncomms11199/images_article/ncomms11199-f3.jpg
a) Representative LIPG IHC staining on primary BCa tissues (cohort of 439 BCa patients). LIPG is expressed in the cytoplasm of tumour cells. Faint staining is also detected in the extracellular area. Scale bar, 50 μm. (b) Representative LIPG IHC staining in normal breast tissue from mammoplasty reductions. Weak LIPG expression occurs in epithelial cells from ducts and lobuli. Scale bar, 50 μm. (c) LIPG protein expression in human cancer cell lines compared with HMECs. Actin was used as loading control.*Unspecific band. Of note, MDA435 are of melanoma origin. (d) LIPG protein levels by IHC staining in Luminal, Her2, and triple negative samples in the Spanish BCa set (cohort of 439 BCa patients). Data is average±s.d. (e) Spearman correlation (P=0.000001) between FoxA and LIPG IHC staining intensities in Spanish BCa set (cohort of 439 BCa patients). (f) Left panel, in vitro proliferation curves of MCF7 and MDA231 cells transduced with a control or a LIPG short hairpin. Data are average±s.e.m.; n=3. (right) LIPG protein expression in shLIPG MCF7 and shLIPG MDA231 cells. The blot shown is representative of three independent experiments. P value is the result of T-test.**P≤0.01, ***P≤0.001. (g) Tumour growth of the indicated cell populations inoculated in Balb/c nude mice are determined at the indicated time points.P value is the result of T-test. Data are average±s.e.m.; n= 6–8 tumours. *P≤0.05.
LIPG is a phospholipase located in the cytosol and cellular membrane and has been shown to hydrolyse extracellular phospholipids from high-density lipoprotein that are afterwards incorporated into intracellular lipid species thus providing lipid precursors of cell metabolism17, 18. Thus we questioned whether LIPG regulates essential lipid intake in BCa and whether it is necessary for proliferation. To validate this hypothesis, we genetically downregulated the expression of this protein in MCF7 and MDA231 cells by means of sh-RNA (Fig. 3f and Supplementary Fig. 2c). LIPG depletion blunted BCa cell capacity to proliferate in vitro (Fig. 3f), as previously observed in FoxA-depleted cells (Supplementary Fig. 1d,e), and caused a reduction in invasion and self-renewal properties (Supplementary Fig. 3a–d). Similarly, LIPG-depleted cells were unable to grow tumours in vivo (Fig. 3g).
LIPG induces BCa cells lipid metabolic reprograming
Figure 4: LIPG regulates the uptake of lipids in BCa cells inducing a lipid metabolic reprograming.

http://www.nature.com/ncomms/2016/160405/ncomms11199/images_article/ncomms11199-f4.jpg
(a) Schematic representation of LIPG action. (b) Heat map representation of the downregulated (blue) lipids identified by MS/MS in the cell homogenates of MCF7 or MDA231 LIPG-depleted cells compared with shControl cells. Depicted lipids have a fold change >1.5 and P value<0.05 using the Welch’s t-testn=5. (c) Downregulated lipid species (previously identified in b) that are common to LIPG-depleted MCF7 and LIPG-depleted MDA231 cells. ShControl cells (red box), and shLIPG (blue box). P values are <0.05 and calculated using Welch’s t-test, n=5. Whiskers extend to a maximum of 1.5 × IQR beyond the box. (d) Heat map representation of the upregulated (red) lipids identified by MS/MS in the media of MCF7 or MDA231 LIPG-depleted cells compared with the corresponding shControl cells. Characterized lipids have a fold change >1.5 and P value<0.05 using the Welch’s t-test n=5. (e) Upregulated lipid species in the media (previously identified in d) that are common to LIPG-depleted MCF7 and LIPG-depleted MDA231 cells. ShControl cells (red box), and shLIPG cells (blue box). P values are <0.05 and calculated using Welch’s t-test, n=5. Whiskers extend to a maximum of 1.5 × IQR beyond the box. (f) Heat map representation of the MS/MS downregulated (blue) lipids in the cell media of MCF7/MDA231 LIPG-depleted or shControl cells (as described in d) compared with fresh medium (without cell incubation). Depicted lipid species have a log2 fold change>1.5 and P value<0.05 using the Welch’s t-test n=5. (g) MDA231 and MCF7 cell growth for 48 h in complete medium: medium containing 10% FBS 10%); lipoprotein-free medium: medium containing 10% free lipoprotein FBS; and LPC (18:0): medium containing 10% free lipoprotein FBS and 20 μM of LPC (18:0). P value is the result of T-test. Data are average±s.e.m.; n=3. **P≤0.01, ***P≤0.001, ****P≤0.0001. (h) Above, schematic representation of the experimental protocol used. (bottom) Tumour growth of the indicated cell populations inoculated in Balb/c nude mice treated with high-fat diet (HFD) are determined at the indicated time points. P value is the result of T-test. Data are average±s.e.m.; n= 6–8 tumours. *P≤0.05, **P≤0.01. Inside graph, plasma cholesterol levels of animals treated with standard diet (SD) or HFD. P value is the result of T-test. Data are average±s.e.m.; n= 4 animals per group. **P≤0.01, ***P≤0.001.
LIPG location has been shown to be functional on the outer face of the cellular membrane (Fig. 4a)18, thus we postulated the possibility that BCa cells are dependent on LIPG function to access extracellular lipids to support their growth needs. To test this notion, we profiled the media of control and LIPG-depleted MCF7 and MDA231 cells following the same liquid chromatography-mass spectrometry-based untargeted lipidomic approach as for cell homogenates. LIPG depletion prevented the absorption of particular lipids from the media (Supplementary Fig. 4a). The structural identification of the lipids by MS/MS confirms the absence of degradation of glycerophospholipids belonging to the LPC class in both MCF7 and MDA231 cells, which is depicted by higher levels in the media of these species in LIPG-depleted when compared with control cells (Fig. 4d,e). Interestingly when we analysed the LPCs species in the media of control and LIPG-depleted cells and compared with fresh media (without cells), all LPC species from control cell media were decreased. This reduction was weaker in the media of Sh LIPG cells, indicating that LIPG-depleted cells have a defect in processing and importing of pre-existing lipid species from the medium (Fig. 4f).
Finally, we evaluated which of the commonly identified potential substrates of LIPG sustains BCa cell proliferation. Initially, we confirmed that the growth of MCF7 and MDA231 cells is impaired when grown in vitro in lipoprotein-depleted media (Fig. 4g). Next we tested the capacity of LPC (18:0) to rescue BCa cell growth in the absence of lipoproteins and confirmed that this lysophosphatidylcholine was able to restore the cells’ capacity to proliferate (Fig. 4g). In accordance, this process was dependent on LIPG expression (Fig. 4g). Similarly, LIPG-depleted cells were not able to grow in vivo in animals fed with high-fat diet (Fig. 4h) indicating that LIPG is indispensable to process the extracellular lipids and mediate their uptake by the cells, irrespectively of the concentration of lipid substrates in circulation, a phenotype also observed in FoxA-depleted cells (Fig. 4h).
LIPG activity supports BCa growth
Figure 5: LIPG activity is essential for BCa growth.

http://www.nature.com/ncomms/2016/160405/ncomms11199/images_article/ncomms11199-f5.jpg
(a, top) Homology 3D structural model of LIPG (backbone coloured according to the QMEANlocal parameter values; red residues with low error). The heavy atoms of the three catalytic residues are shown explicitly and the residue mutated in this study is shown in green (Asp 193). (b) FoxA1, FoxA2 and LIPG protein expression in MCF7, MDA231 and their derivative cells determine by western blot. FoxA1 and FoxA2 depletion was achieved with a doxycycline-inducible short hairpin vector. FoxA-depleted cells were rescued by expression of a WT or Inactive LIPG. Cell populations were cultured in the presence or absence of doxycycline for 6 days. *blots represent different exposition times. (c) Tumour growth of the indicated cell populations inoculated in Balb/c nude mice are determined at the indicated time points. Pvalue is the result of T-test. Data are average±s.e.m.; n=5–8 tumours. *P≤0.05, **P≤0.01. (d) MDA231 and MCF7 cell growth for 48 h treated with DMSO (control), FAS inhibitor (C75) and/or lipase inhibitor (Orlistat). For MDA231 cells C75 was used at a final concentration of 10 μg ml−1 and for MCF7 cells 8 μg ml−1. Orlistat was used at a final concentration of 30 or 10 μg ml−1 in MCF7 or MD231 respectively. Pvalue is the result of T-test. Data are average±s.e.m.; n=3.*P≤0.05, **P≤0.01, ***P≤0.001. (e) Forty-eight hours cell growth of MDA231 or MCF7 cells overexpressing exogenous WT or Inactive LIPG. Cells were treated with DMSO (control) and FAS inhibitor (C75) at a final concentration of 20 μg ml−1. P value is the result of T-test. Data are average±s.e.m.; n=3.***P≤0.001, ****P≤0.0001 (f) Schematic representation showing how FoxA controls LIPG and lipid metabolism to support tumour growth.
As previous reports showed that de novo lipid metabolism is necessary for BCa growth3, 22, we next questioned whether this lipid synthesis was sufficient or, instead, whether exogenous sources are also required to support BCa cell growth and proliferation, as suggested by our experimental data. To this end, we inhibited the activity of fatty acid synthase (FAS) in BCa cells by means of the chemical inhibitor C75 (ref. 23). FAS activity is crucial for de novo lipid synthesis in cancer cells3,22. To test the complementarity of both de novo and/or exogenous lipid supplies, we used a C75 concentration causing a 50% reduction in BCa cell growth in vitro 48 h post incubation (Fig. 5d andSupplementary Fig. 5d). Similarly, we tested the contribution of LIPG inhibition by means of treatment with a lipase inhibitor, Orlistat21. A specific dose causing a 50% reduction in the growth of each BCa cell line was further used (Fig. 5d and Supplementary Fig. 5d). Interestingly, concomitant treatment with FAS and LIPG inhibitors caused an additive effect, blunting BCa cell growth (Fig. 5d). Next, we evaluated whether LIPG activity was sufficient to rescue the chemical inhibition of FAS. To this end, we overexpressed WT and inactive LIPG and grew MCF7 and MDA231 cells in the presence or absence of a high dose of C75 (20 mg ml−1), which blocks cell growth (Supplementary Fig. 5d). Complete blockade of FAS was not rescued by LIPG (Fig. 5e). Collectively, our results suggest that both exogenous lipid precursors provided by means of LIPG activity and de novo lipid synthesis mediated by FAS are necessary for BCa cell growth.
Here we reveal that FoxA factors provide a central metabolic growth function by specifically regulating LIPG expression, thereby allowing the acquisition of indispensable extracellular lipids for BCa tumour proliferation. FoxA family of transcription factors are expressed in the vast majority of BCa and FoxA1 is expressed across various BCa subtypes. Moreover we show that, in some cases, its absence is associated with the expression of FoxA2. Interestingly, in addition of FoxA1 contribution to luminal commitment24, 25, 26, 27 the factor may drive BCa growth by specifically regulating LIPG levels.
The catalytic activity of LIPG generates extracellular lipid precursors that are imported to fulfill the intracellular production of lipid species (Fig. 5f). LIPG downregulation blocks BCa cell growth, thereby indicating that the import of extracellular lipid precursors is important for the proliferation of these cells. This is a striking observation given that it is generally believed that de novo fatty acid synthesis is the main driver of tumour growth22. Indeed, our experimental data with LIPG-depleted BCa cells revealed a massive decrease of most intracellular glycerolipid intermediates in the synthesis of TG (PC, PE, PG and DG) and their derivatives (LPC and LPE). Accordingly, certain lipid species (LPC) in the media were not decreased in LIPG-depleted cells as much as in control cells, thus indicating that extracellular lipids are the substrates for intracellular lipid production. In particular, we demonstrate the relevance of extracellular LPC (18:0) for BCa cell proliferation in a lipoprotein-depleted medium, a process dependent on LIPG. In this context, a high-fat diet was shown to rescue the absence of a critical intracellular lipase, Monoacylglycerol lipase, for cancer pathogenesis given cancer cells ability to uptake lipids from the extracellular compartment was functional19. Herein, we showed that this rescue mechanism is not functional in BCa cells in the absence of FoxA2 or LIPG. In support of this notion, it is worth noting that extracellular LIPG activity releases fatty acids from high-density lipoprotein phospholipids and these acids are further employed for intracellular lipid production in the human hepatic cell line HepG2 (refs 28, 29).
In conclusion, BCa cells are dependent on a mechanism to supply precursors derived from extracellular sources for intracellular lipid production, and LIPG fulfills this function. Therefore, LIPG stands out as an important component of the lipid metabolic adaptations that BCa cells, and not normal tissue, must undergo to support high proliferation rates. Our results also suggest thatde novo lipid synthesis is necessary but not sufficient to support lipid production for BCa tumour growth. Accordingly, recent clinical studies demonstrate the association between lipids and lipoproteins in circulation and risk of BCa in women with extensive mammographic density. This observation implies that interventions aimed to reduce them may have effect on BCa risk30. All together, these observations make LIPG activity an Achilles heel of luminal and, more importantly, of triple negative/basal-like breast tumours, for which limited therapeutic options are currently available.
In normal cells, the glucose carbon flow is directed into a de novo lipogenic pathway that is regulated, in part, via phosphoinositide-3 kinase (PI-3K)-dependent activation of ATP citrate lyase (ACL), a key rate-limiting, enzyme in de novo lipogenesis. ACL is a cytosolic enzyme that catalyzes the generation of acetyl CoA from citrate. Inhibition of ACL results in a loss of B-cell growth and cell viability [10] .
The plasma membrane and its constituent phosphoinositides form the basis of the phosphatidylinositol 3-kinase (PI3-K) signaling pathway, which is crucial for cell proliferation and survival. Phosphatase and tensin-homolog deleted on chromosome 10 (PTEN) is a tumor-suppressor protein that regulates phosphatidylinositol 3-kinase (PI3-K) signaling by binding to the plasma membrane and hydrolyzing the 3′ phosphate from phosphatidylinositol (3,4,5)-trisphosphate (PI(3,4,5)P3) to form phosphatidylinositol (4,5)-bisphosphate (PI(4,5)P2). Several loss-of-function mutations in PTEN that impair lipid phosphatase activity and membrane binding are oncogenic, leading to the development of a variety of cancers. Of these three residues, R335 was observed to interact with the membrane to the greatest extent across all of the simulations. R335L, in common with several other germline mutations, has been associated with the inherited cancer [11] .
ACLY is up-regulated or activated in several types of cancers, and its inhibition is known to induce proliferation arrest in cancer cells both in vitro and in vivo. The last studies were showed that BCR-mediated signaling is regulated in part by the amount of membrane cholesterol. It was observed that statins (Lovostatin), the pharmacological inhibitors of cholesterol synthesis, induce apoptosis of CLL cells in vitro and in vivo. Also the ectopic expression of CD5 in a B-cell line stimulates the transcription of genes involved in the synthesis of cholesterol [12] .
[10] Zaidi N, Swinnen JV, Smans K. ATP-citrate lyase: a key player in cancer metabolism Cancer Res; 2012 (11): 3709-14.
[11] Craig N, Mark S.P. Sansom. Defining the Membrane-Associated State of the PTEN Tumor Suppressor Protein. Biophys J 2013; 5; 104(3: 613–21.
[12] Tomowiak C, Kennel A, Gary-Gouy, Hadife N. High Membrane Cholesterol in CLL B-Cells and Differential Expression of Cholesterol Synthesis Genes in IG GENE Unmutated vs Mutated Cells. British Journal of Medicine & Medical Research 2012; 2(3): 313-26.
Cancer’s Vanguard
Exosomes are emerging as key players in metastasis.
In 2005, David Lyden noticed something unexpected. He and his colleagues at Weill Cornell Medical College had been researching metastasis—the spread of cancer from one part of the body to another. The team had shown that bone marrow–derived cells (BMDCs) were recruited to future metastatic sites before the arrival of tumor cells, confirming that metastasis occurred after a habitable microenvironment, or “premetastatic niche,” had been prepared.1
But carefully studying images of this microenvironment in the lung tissue of mice, Lyden saw something else. Amongst the BMDCs, the micrographs showed tiny specks, far too small to be cells, gathering at the future site of metastasis. “I said, ‘What are these viruses doing here?’” recalls Lyden. “I had no idea about exosomes, microvesicles, and microparticles.”
Those specks, Lyden would come to realize, were in fact primary tumor–derived exosomes. These membrane-enclosed vesicles packed full of molecules are now attracting growing attention as important mediators of intercellular communication, particularly when it comes to cancer’s insidious capacity to spread from one organ to another.
Preparing the ground
Tumors require a community of support cells, including fibroblasts, BMDCs, and endothelial cells, to provide functional and structural assistance and to modulate immune system behavior. Bringing together the first members of this community before the arrival of tumor cells is all part of cancer’s survival strategy, says Joshua Hood, a cancer researcher at the University of Louisville.
“It wouldn’t be efficient for tumor cells to strike out on their own, and just say, ‘Oh, here we are!’” he says. “They would run the risk of being destroyed.” Preparing a “nest” in advance makes the process much safer. “Then the tumor can just efficiently come along and set up shop without ever having to fight much of a battle with the immune system.”
But although Lyden’s group had shown that this preparation was taking place, it remained unclear how such a process might be regulated. For the next few years, many cancer researchers believed that tumor cells must communicate with the premetastatic niche primarily through tumor-secreted signaling molecules such as cytokines.
Meanwhile, research into extracellular vesicles, previously considered biological garbage bags, was revealing new modes of intercellular communication. In 2007, a group of scientists in Sweden discovered that exosomes, tiny vesicles measuring just 30 nanometers to 100 nanometers across, transport mRNA and microRNAs intercellularly, with the potential to effect changes in protein synthesis in recipient cells.2 A new means for tumors to regulate distant cellular environments came into focus, and research on exosomes exploded. In 2011, Hood and his colleagues showed that exosomes facilitate melanoma metastasis through the lymphatic system.3 The following year, Lyden’s group demonstrated that tumor-derived exosomes can direct BMDCs to one of melanoma’s most common sites of metastasis, the lung.4 Exosomes, it seemed, had been underestimated.
Tiny terraformers
Armed with the knowledge that exosomes are involved in multiple stages of melanoma metastasis, Lyden’s lab went searching for the vesicles’ potential role in the metastasis of other cancers. Turning to pancreatic ductal adenocarcinoma (PDAC)—one of the most lethal cancers in humans—postdoctoral researcher Bruno Costa-Silva led a series of exhaustive in vitro and in vivo experiments in mouse models to detail the process of premetastatic niche formation in the liver, PDAC’s most common destination. The team’s results, published last May, reveal an intricate series of sequential steps—mediated by PDAC-derived exosomes (Nature Cell Biol, 17:816-26, 2015).
Using fluorescence labeling, Lyden’s group observed that PDAC-derived exosomes are taken up by Kupffer cells, specialized macrophages lining the outer walls of blood vessels in the liver. There, the exosomes trigger the cells’ secretion of transforming growth factor β (a type of cytokine involved in cell proliferation), plus the production of fibronectin by neighboring hepatic stellate cells, and the recruitment of BMDCs.
The researchers also showed that this cascade of events could be inhibited by depleting exosomal macrophage migratory inhibitory factor (MIF), an abundant protein in PDAC exosomes. “If you target the specific proteins of exosomes, you can reduce metastasis,” explains coauthor Héctor Peinado, leader of the microenvironment and metastasis group at the Spanish National Cancer Research Center.
For Hood, the findings add to a developing picture of exosomes’ vital role as “vanguard” in the progression of cancer. “It’s like the colonization of a new planet,” he says. “They’re terraforming the environment to make it hospitable.”
MOLECULAR & CELLULAR BIOLOGY
THE PAPERS
- B. Costa-Silva et al., “Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver,”Nature Cell Biol, 17:816-26, 2015.
- A. Hoshino et al., “Tumour exosome integrins determine organotropic metastasis,” Nature, 527:329-35, 2015.
- L. Zhang et al., “Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth,”Nature, 527:100-04, 2015.
Internal mail
Although research was revealing the steps involved in forming premetastatic sites, it was less clear how these sites were being selected. “This has always been a great mystery in cancer,” says Ayuko Hoshino, a research associate in Lyden’s lab. “Why do certain cancers metastasize to certain organs?”
One theory, proposed in 1928 by pathologist James Ewing, suggested that anatomical and mechanical factors explained organ specificity in metastasis. The premetastatic niche, then, might form wherever exosomes are likely to land. But this couldn’t be the whole story, says Hoshino. “For instance, there’s eye melanoma. Thinking about that site, you could imagine it metastasizing to the brain. But actually, it almost only metastasizes to the liver.”
Because exosomes arrive at metastatic sites before tumor cells, the team reasoned, perhaps the exosomes themselves were organotropic (i.e., attracted to particular organs or tissues). Sure enough, Lyden says, when Hoshino and Costa-Silva began injecting tumor-derived exosomes into mice, “their preliminary findings were that wherever they injected the exosomes, the pancreatic cancer ones were ending up in the liver and the breast metastasis exosomes would end up in the lung.”
Using mass spectrometry, the researchers analyzed the protein content of exosomes from lung-tropic, liver-tropic, and brain-tropic tumors. They found that the composition of exosomes’ integrins—membrane proteins involved in cell adhesion—was destination-specific (Nature, 527:329-35, 2015). Exosomes bearing integrin α6β4, for example, were directed to the lung, where they could prepare a premetastatic niche potent enough even for normally bone-tropic tumor cells to colonize. Integrin αvβ5, meanwhile, directed metastasis to the liver.
The researchers also showed that exosomal integrins didn’t necessarily correspond to the parent-cell proteins, making exosomes potentially better indicators of where a cancer will spread than the tumor cells themselves. “We can show that an integrin that’s high in the tumor cell might be completely absent in the tumor exosome or vice versa,” says Lyden, adding that, taken together, the results point to a role for exosomes in “dictating the future sites of metastasis.”
“It’s a beautiful story,” says Dihua Yu, a molecular and cellular oncologist at the University of Texas MD Anderson Cancer Center. “This is a very novel finding that gives really good indicators for potential strategies to intervene in metastasis.”
Metastatic crosstalk
In the same month that Lyden’s group published its work on organotropism, Yu’s own lab published a different exosome study—one that told another side of the story.
Yu and her colleagues had found that when tumor cells in mice metastasized to the brain, they downregulated expression of a tumor suppressor gene called PTEN, and became primed for growth at the metastatic site. When the tumor cells were taken out of the microenvironment and put in culture, however, they restored normal PTEN expression.
The researchers demonstrated that a microRNA from astrocytes—star-shape glial cells in the brain—reversibly downregulated the levels of PTEN transcripts in the tumor cells, but they couldn’t figure out how the microRNA was getting into the tumor. Blocking “obvious signaling pathways,” such as gap junctions, failed to have an effect, Yu says.
Scrutinizing astrocyte-conditioned media using electron microscopy, the researchers identified spherical vesicles between 30 nanometers and 100 nanometers in diameter—the defining size of exosomes. Exposing mouse tumor cells to these vesicles increased cell microRNA content and reduced PTENexpression (Nature, 527:100-04, 2015). The study revealed yet another role for exosomes in the communication between tumors and their microenvironment.
The findings were a surprise, says Yu, not least because they showed a different perspective from the bulk of recent research. “We’re talking about astrocytes in the brain secreting exosomes to give welcome help to the cancer cells,” she says.
“I find it an extremely interesting paper because it shows that the astrocytes can change the whole phenotype of the tumor in the brain,” says Lyden. He adds that the results underline the importance of studying the mutational status of tumors at various sites. “All this work in exosomes, it adds to the complexity,” he says. “We can’t just target tumor cells at the primary site. We’ll have to understand all the details of metastasis if we’re really going to tackle it.”
What’s next?
The discovery of multiple roles for exosomes in metastasis has generated excitement about the potential for their use in diagnostics and treatment. As protective containers of tumor-derived genetic material, exosomes could provide information about the status of cancer progression. And as mediators of premetastatic niche formation, they make obvious targets for inhibition. (See “Banking on Blood Tests,”here.)
Exosomes might even be useful as vehicles to deliver drugs because they’re patient-matched and “naturally designed to function in a biocompatible way with living systems,” says Hood. “You could take them out of people, and at some point down the road try to have patients be their own nanofactory, using their own particles for treatment purposes.”
Pancreatic cancer exosomes initiate pre-metastatic nihe formation in the liver
Bruno Costa-Silva, Nicole M. Aiello, Allyson J. Ocean, et al. Nature Cell Biology 2015; 17,816–826 http://dx.doi.org:/10.1038/ncb3169
Pancreatic ductal adenocarcinomas (PDACs) are highly metastatic with poor prognosis, mainly due to delayed detection. We hypothesized that intercellular communication is critical for metastatic progression. Here, we show that PDAC-derived exosomes induce liver pre-metastatic niche formation in naive mice and consequently increase liver metastatic burden. Uptake of PDAC-derived exosomes by Kupffer cells caused transforming growth factor β secretion and upregulation of fibronectin production by hepatic stellate cells. This fibrotic microenvironment enhanced recruitment of bone marrow-derived macrophages. We found that macrophage migration inhibitory factor (MIF) was highly expressed in PDAC-derived exosomes, and its blockade prevented liver pre-metastatic niche formation and metastasis. Compared with patients whose pancreatic tumours did not progress, MIF was markedly higher in exosomes from stage I PDAC patients who later developed liver metastasis. These findings suggest that exosomal MIF primes the liver for metastasis and may be a prognostic marker for the development of PDAC liver metastasis.
View full text
Tumour exosome integrins determine organotropic metastasis
Ayuko Hoshino, Bruno Costa-Silva, Tang-Long Shen, Goncalo Rodrigues, Ayako Hashimoto, Milica Tesic Mark, et al. Nature Nov 2015; 527,329–335 http://dx.doi.org:/10.1038/nature15756
Ever since Stephen Paget’s 1889 hypothesis, metastatic organotropism has remained one of cancer’s greatest mysteries. Here we demonstrate that exosomes from mouse and human lung-, liver- and brain-tropic tumour cells fuse preferentially with resident cells at their predicted destination, namely lung fibroblasts and epithelial cells, liver Kupffer cells and brain endothelial cells. We show that tumour-derived exosomes uptaken by organ-specific cells prepare the pre-metastatic niche. Treatment with exosomes from lung-tropic models redirected the metastasis of bone-tropic tumour cells. Exosome proteomics revealed distinct integrin expression patterns, in which the exosomal integrins α6β4 and α6β1 were associated with lung metastasis, while exosomal integrin αvβ5 was linked to liver metastasis. Targeting the integrins α6β4 and αvβ5 decreased exosome uptake, as well as lung and liver metastasis, respectively. We demonstrate that exosome integrin uptake by resident cells activates Src phosphorylation and pro-inflammatory S100 gene expression. Finally, our clinical data indicate that exosomal integrins could be used to predict organ-specific metastasis.
- Paget, S. The distribution of secondary growths in cancer of the breast. 1889. Cancer Metastasis Rev. 8, 98–101 (1989)
- Hart, I. R. & Fidler, I. J. Role of organ selectivity in the determination of metastatic patterns of B16 melanoma. Cancer Res. 40, 2281–2287 (1980)
- Müller, A. et al. Involvement of chemokine receptors in breast cancer metastasis. Nature410, 50–56 (2001)
- Weilbaecher, K. N., Guise, T. A. & McCauley, L. K. Cancer to bone: a fatal attraction. Nature Rev. Cancer 11, 411–425 (2011)
- Zhou, W. et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 25, 501–515 (2014)
- Chang, Q. et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis.Neoplasia 15, 848–862 (2013)
- Lu, X. & Kang, Y. Organotropism of breast cancer metastasis. J. Mammary Gland Biol. Neoplasia 12, 153–162 (2007)
…….
Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth
Lin Zhang, Siyuan Zhang, Jun Yao, Frank J. Lowery, Qingling Zhang, Wen-Chien Huang, et al. Nature Nov 2015; 527,100–104 http://dx.doi.org:/10.1038/nature15376
The development of life-threatening cancer metastases at distant organs requires disseminated tumour cells’ adaptation to, and co-evolution with, the drastically different microenvironments of metastatic sites1. Cancer cells of common origin manifest distinct gene expression patterns after metastasizing to different organs2. Clearly, the dynamic interaction between metastatic tumour cells and extrinsic signals at individual metastatic organ sites critically effects the subsequent metastatic outgrowth3, 4. Yet, it is unclear when and how disseminated tumour cells acquire the essential traits from the microenvironment of metastatic organs that prime their subsequent outgrowth. Here we show that both human and mouse tumour cells with normal expression of PTEN, an important tumour suppressor, lose PTEN expression after dissemination to the brain, but not to other organs. The PTEN level in PTEN-loss brain metastatic tumour cells is restored after leaving the brain microenvironment. This brain microenvironment-dependent, reversible PTEN messenger RNA and protein downregulation is epigenetically regulated by microRNAs from brain astrocytes. Mechanistically, astrocyte-derived exosomes mediate an intercellular transfer of PTEN-targeting microRNAs to metastatic tumour cells, while astrocyte-specific depletion of PTEN-targeting microRNAs or blockade of astrocyte exosome secretion rescues the PTEN loss and suppresses brain metastasis in vivo. Furthermore, this adaptive PTEN loss in brain metastatic tumour cells leads to an increased secretion of the chemokine CCL2, which recruits IBA1-expressing myeloid cells that reciprocally enhance the outgrowth of brain metastatic tumour cells via enhanced proliferation and reduced apoptosis. Our findings demonstrate a remarkable plasticity of PTEN expression in metastatic tumour cells in response to different organ microenvironments, underpinning an essential role of co-evolution between the metastatic cells and their microenvironment during the adaptive metastatic outgrowth. Our findings signify the dynamic and reciprocal cross-talk between tumour cells and the metastatic niche; importantly, they provide new opportunities for effective anti-metastasis therapies, especially of consequence for brain metastasis patients.
View full text
- Quail, D. F. & Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nature Med. 19, 1423–1437 (2013)
- Park, E. S. et al. Cross-species hybridization of microarrays for studying tumor transcriptome of brain metastasis. Proc. Natl Acad. Sci. USA 108, 17456–17461 (2011)
- Joyce, J. A. & Pollard, J. W. Microenvironmental regulation of metastasis. Nature Rev. Cancer 9, 239–252 (2009)
- Vanharanta, S. & Massagué, J. Origins of metastatic traits. Cancer Cell 24, 410–421 (2013)
- Gray, J. Cancer: genomics of metastasis. Nature 464, 989–990 (2010)
- Friedl, P. & Alexander, S. Cancer invasion and the microenvironment: plasticity and reciprocity. Cell 147, 992–1009 (2011)
Banking on Blood Tests
How close are liquid biopsies to replacing current diagnostics?
A tumor’s physical environment fuels its growth and causes treatment resistance.
Ahelium balloon tugs gently at the end of its string. The tension in the string resists the buoyant force of the helium, and the elastic nature of the balloon’s rubber contains the helium gas as it tries to expand. Cutting the string or poking the rubber with a pin reveals the precarious balance between the forces, upsets the equilibrium, and sets the system into motion.
Some biological tissues also exist in such a state of offsetting forces. The most familiar example is the balance between blood pressure and the elastic tension in the cardiovascular system that contains and conveys blood without bursting or collapsing. And in tumors, both solid and fluid forces are generated that make the cancerous tissue a lot like that helium balloon: cut a tumor with a scalpel and it rapidly swells and deforms as pent-up forces break free from structural elements that are severed.1
One force that is notably higher in tumors than in healthy tissues is fluid pressure, resulting from hyperpermeable, leaky blood vessels and a dearth of draining lymphatic vessels. Researchers have known since the 1950s that tumors exhibit elevated fluid pressure, but the implications for tumor progression and drug delivery were not realized until the late 1980s. That was when we (R.K.J. and colleagues) used a mathematical model to predict—and subsequently validate in animal and human tumors—that a precipitous drop in fluid pressure at the tumor–normal tissue interface causes interstitial fluid to ooze out of the tumor.2 This seeping fluid pushes drugs, growth factors, and cancer cells into the surrounding tissue and lymphatics, reducing drug delivery and facilitating local tumor invasion and distant metastasis.
Based on this insight, we suggested in 2001 that anti-angiogenic drugs could be used to lower a tumor’s fluid pressure and improve treatment outcome.3 This hypothesis changed the thinking about how existing anti-angiogenesis therapies actually work and spurred research into other physical forces acting in cancer.4 In the last 15 years, researchers have identified diverse sources of increased pressure in tumors, which may serve as possible targets for cancer therapy.5 For example, solid forces exerted by the extracellular matrix can be reduced by treatment with drugs approved by the US Food and Drug Administration (FDA) for controlling hypertension (angiotensin blockers) or diabetes (metformin). Retrospective clinical studies have found improved survival in cancer patients who were treated with these agents, which are now being tested in prospective trials for a variety of solid tumors.6,7
Tumors under pressure
In vitro experiments showing that cancer cells actively migrate in response to fluid flow have supported the hypothesis that fluid escaping from the boundary of a tumor may guide the invasive migration of cancer cells toward lymphatic or blood vessels, potentially encouraging metastasis. There remains controversy over how the fluid forces induce the migration; the cells may respond to chemical gradients created by the cells and distorted by the flowing fluid,8 or the fluid may activate cell mechanosensors.9 Because of the potential for new therapeutic interventions, the transduction of mechanical fluid forces into biochemical signals by cell mechanosensors is an active area of investigation. In a more direct manner, the fluid flow can physically carry cancer cells to lymph nodes.
Fluid forces may also promote tumor progression by recruiting blood vessels into the cancerous mass.10 Because tumor blood vessels are leaky, plasma can pass freely between vessels that have different pressures. When this happens at the periphery of a tumor, where angiogenic growth factors are prevalent, there can be synergistic induction of new vessel sprouts.

UNDER PRESSURE See full infographic: WEB | PDF© N.R.FULLER, SAYO-ART LLC
And fluid pressure is just one of the many forces in a tumor that can influence its development and progression. Tumors also develop increased solid pressure, as compared with normal tissue, stemming from the uncontrolled division of cancer cells and from the infiltration and proliferation of stromal and immune cells from the surrounding tissue and circulation. High-molecular-weight polysaccharides known as hydrogels found in the extracellular matrix (ECM) also add pressure on a tumor. The most well-studied of these hydrogels is hyaluronan; when the polysaccharide absorbs water, it swells, pressing on surrounding cells and structural elements of the tissue.
The ECM contains a highly interconnected network of collagen and other fibers and is normally very good at resisting and containing such tension. It also has support from infiltrating myofibroblasts, which detect areas where the ECM density or tension is not normal and initiate actomyosin-based contraction of collagen and elastin matrix structures to restore tensional homeostasis. But while this repair effort is typically effective in healthy tissues, uncooperative tumor cells interfere with these efforts, both by themselves generating pressure and by hyperactivating cancer-associated fibroblasts to produce more ECM and thus produce even more force.11
Because cell growth and ECM composition are not spatially uniform in cancer, tumors are subjected to multiple, dispersed sources of pressure associated with matrix “containers” of various sizes. This solid pressure from within the tumor deforms the surrounding normal tissue, potentially facilitating the metastatic escape of cancer cells. The physical forces also compress blood vessels and lymphatic vessels in the tumor and adjacent normal tissue,12 increasing the fluid pressure in the tumor13 and interrupting the delivery of nutrients, removal of waste, and entry of tumor-targeted drugs via the blood.4 Insufficient blood flow also results in poor oxygenation, which has been linked to immunosuppression, inflammation, invasion, and metastasis, as well as lowered efficacy of chemo-, radio-, and immunotherapies.4 These are all indirect consequences of solid stresses in and on tumors.
Such forces can also have direct effects on cancer cells, and may serve as independent triggers for tumor invasion. Mechanical forces are central to many of our sense systems, such as hearing, touch, and pain, and to tissue maintenance programs, such as bone regeneration and blood vessel remodeling. In these systems, mechanical forces are transduced by mechanosensors to activate downstream biochemical and genetic pathways. (See “Full Speed Ahead,” The Scientist, December 2009.) Cancer cells may similarly be able to sense and respond to dynamic forces in tumors. We have shown, for example, that metastatic cancer cells exposed to compressive stresses in a culture dish undergo a phenotypic transformation to become more invasive,14 and others have shown that compressive forces applied in vivo can also induce oncogenes in normal epithelium of the mouse colon.15
It is thus becoming quite clear that the physical environment can influence a tumor’s development and spread, and it may even be possible for physical forces to kick-start cancerous growth.
…..
Full Speed Ahead
Physical forces acting in and around cells are fast—and making waves in the world of molecular biology.
When it comes to survival, few things are more important than being able to respond quickly to a change of circumstances. And when it comes to fast-acting indicators, it turns out that signals induced by physical forces acting in and around cells, appropriately dubbed biomechanical signals, are the champions of the cellular world.
“If you look at this mechanical signaling, it’s about 30 meters per second—that’s very fast,” says bioengineer Ning Wang of the University of Illinois at Urbana-Champaign. That’s faster than most family-owned speedboats, and second only to electrical (e.g., nerve) impulses in biological signaling. By comparison, small chemicals moving by diffusion average a mere 2 micrometers per second—a speed even the slowest row boater could easily top.
Indeed, when the two signal types were pitted against each other in a cellular race last year, the mechanical signals left chemical signals in their wake, activating proteins at distant sites in the cytoplasm in just a fraction of a second, at least 40 times faster than their growth factor opponent.1 Mechanical signals are so fast, Wang adds, they are “beyond our resolution,” meaning that current imaging techniques cannot capture the very first cellular changes that result from mechanical stress, which occur within nanoseconds.
For centuries, scientists have scrutinized the molecular inner workings of the body, with little or no regard to the physical environment in which these biological reactions take place. But the growing realization that physical forces have a pervasive presence in physiology (operating in a variety of bodily systems in thebone, blood, kidney, and ear, for instance), and act with astonishing speed, has caused many to consider the possibility that mechanical signaling may be just as important as chemical communication in the life of a cell.
“Biologists have traditionally ignored the role of mechanics in biology,” says biomechanical engineer Mohammad Mofrad of the University of California, Berkley, “[but] biomechanics is becoming increasingly accepted, and people are recognizing its role in development, in disease, and in general cellular and tissue function.”
The wave within: Mechanical forces acting inside the cell
Once believed to be little more than sacks of chemically active goop, cells didn’t seem capable of transmitting physical forces into their depths, and researchers largely limited their search for molecules or structures that respond to physical forces, or mechanosensors, to the plasma membrane.
Mechanical signaling may be just as important as chemical communication in the life of a cell.
In the late 1990s, however, closer examination revealed that the cell’s interior is in fact a highly structured environment, composed of a network of filaments.2 Pull on one side of the cell, and these filaments will transmit the force all the way to other side, tugging on and bumping into a variety of cellular structures along the way—similar to how a boat’s wake sends a series of small waves lapping up on a distant and otherwise peaceful shoreline. Scientists are now realizing the potential of such intracellular jostling to induce molecular changes throughout the cell, and the search for mechanosensing molecules has escalated dramatically in scope, including, for example, several proteins of the nucleus.
It’s a search that will likely last a while, predicts cell biologist Donald Ingber, director of the Wyss Institute for Biologically Inspired Engineering at Harvard University. “To try to find out what’s the mechanosensor is kind of crazy at this point,” he says. As scientists are now learning, “the whole cell is the mechanosensor.”
A key player, most agree, is the cytoskeleton, which is comprised of a variety of microfilaments, including rigid actin filaments and active myosin motors—the two principle components of muscle. Activation of the so-called nonmuscle myosins causes the cytoskeleton to contract, much like an arm muscle does when it lifts a heavy object.
The first intimation that the cytoskeleton could go beyond its established inner-cell duties (molecule transport and cell movement and division) came in 1997, when Ingber did the logical (in hindsight, at least) experiment of pulling on the cells to see what happened inside.2 Using a tiny glass micropipette coated in ligands, Ingber and his team gently probed the surface proteins known as integrins, which secure the cell to the extracellular matrix. When they quickly pulled the micropipette away, they saw an immediate cellular makeover: cytoskeletal elements turned 90 degrees, the nucleus distorted, and the nucleolus—a small, dense structure within the nucleus that functions primarily in ribosome assembly—aligned itself with the direction of the applied force.
“That kind of blew people away,” Ingber recalls. “It revealed that cells have incredible levels of structure not only in the cytoplasm but in the nucleus as well.”
Wang (once a postdoc in Ingber’s lab at the Harvard School of Public Health) and other collaborators combined a similar technique with fluorescent imaging technology to visualize how these forces were channeled within the cell’s interior. Upping the resolution and further refining these techniques, Wang began mapping these intracellular forces as they made their way through the cell. In 2005, the maps confirmed the physical connection between the cell-surface integrins and the nucleus, and showed that these external forces follow a nonrandom path dictated by the tension of the cytoskeletal elements.3
“Biomechanics is becoming increasingly accepted, and people are recognizing its role in development, in disease, and in general cellular and tissue function.”
–Mohammad Mofrad
The end point of these mechanical pathways is likely a mechanosensitive protein, which changes shape in response to the force, thereby exposing new binding areas or otherwise changing the protein’s function. In mitochondria, for example, mechanical forces may trigger the release of reactive oxygen species and activation of signaling molecules that contribute to inflammation and atherosclerosis.
Similarly, proteins on the nuclear membrane may pass mechanical signals into the nucleus by way of a specialized structure known as LINC (linker of nucleoskeleton and cytoskeleton), which physically links the actin cytoskeleton to proteins important in nuclear organization and gene function. To determine if mechanical forces directly affect gene expression, last year scientists began exploiting the increasingly popular fluorescence resonance energy transfer (FRET) technology,1 in which energy emitted by one fluorescent molecule can stimulate another, resulting in a visible energy transfer that can track enzymatic activities in live cells. By combining FRET technology with the techniques that apply physical forces to specific cell membrane proteins, scientists can visualize entire mechanochemical transduction pathways, Wang says.
“The big issue right now in the field of mechanotransduction is whether the genes in the nucleus can be directly activated by forces applied to the cell surface,” Wang explains. While the physical maps of the cytoskeleton tentatively sketch out a path that supports this possibility, confirmatory data is lacking. This combination of new technologies will be “tremendously” helpful in answering that question, he says, and “push the field” towards a more complete understanding of how mechanical forces can influence cellular life.
An early start: Mechanical forces in development
In the world of developmental biology, the cytoskeleton’s role in biomechanics really comes into its own. As the embryo develops, the cells themselves are the force generators, and by contracting at critical times, the cytoskeleton can initiate many key developmental steps, from invagination and gastrulation to proliferation and differentiation, and overall cellular organization.
The idea that physical forces play a role in development is not a new one. In the early 20th century, back when Albert Einstein was first developing the molecular basis of viscosity and scientists were realizing molecules are distinct particles, biologist and mathematician D’Arcy Thompson of the University of Dundee in Scotland suggested that mechanical strain is a key player in morphogenesis. Now, nearly a century later, biologists are finally beginning to agree.
Because Thompson “couldn’t measure [the forces] at that time, that kind of thinking got pushed to the wayside as genetic thinking took over biology,” says bioengineer Christopher Chen of the University of Pennsylvania. That is, until 2003, when Emmanuel Farge of the Curie Institute in France squeezedDrosophila embryos to mimic the compression experienced during early development and activated twist—a critical gene in the formation of the digestive tract.4 These results gave weight to Thompson’s idea that stress in the embryo stimulates development and growth, and inspired developmental scientists to begin considering mechanical effects, Chen says. “Now we’re at the stage where there’s a lot of interest and willingness to consider the fact that mechanical forces are not only shaping the embryo, but are linked to the differentiation programs that are going on.”
Again, the cytoskeleton is a key player in this process. In fruit flies and frogs, for example, nonmuscle myosins contract the actin filaments to generate the compressive forces necessary for successful gastrulation—the first major shape-changing event of development. Myosins similarly influence proliferation in the development of the Drosophila egg chamber, with increased myosin activity resulting in increased cell division.
Cytoskeleton contractility also appears to direct stem cell differentiation. In 2006, Dennis Discher of the University of Pennsylvania demonstrated that the tension of the substrate on which cells are grown in culture is important for determining what type of tissue the cells will form.5 Cells grown on soft matrices that mimic brain tissue tended to grow into neural cells, while cells grown on stiffer matrices grew into muscle cell precursors, and hard matrices yielded bone. In this case, it seems that stiffer substrates increased the expression of nonmuscle myosin, generating greater tension in the actin cytoskeleton and affecting differentiation. (Altering or inhibiting myosin contraction can also affect differentiation.)
“To try to find out what’s the mechanosensor is kind of crazy at this point. As scientists are now learning, the whole cell is the mechanosensor.”
–Donald Ingber
……..
Shaping a tumor
In addition to the influence of physical forces on cancer growth and invasion, forces can alter a tumor’s mechanical properties, and vice versa. Tumors are more rigid, or stiffer, than surrounding tissues, usually because they contain excess collagen in the ECM,5 and this can contain and amplify local forces produced by proliferating cancer cells. On the other hand, tumor rigidity can be further enhanced if the cells exert tension on ECM collagen fibers by pulling on them, or by stretching them, as occurs when tumors grow uncontrollably. Fluid forces can also influence the assembly of collagen fibers within and around tumors,8potentially increasing stiffness.
Importantly, tumor stiffness tends to be associated with poor prognoses, though the reasons for this are not fully understood. Cells are known to differentiate into different lineages depending on the local rigidity;16 for example, stem cells differentiate into bone on stiff substrates, but make adipose (fat) cells on softer substrates. Similar mechanisms are thought to affect tumor progression when the ECM changes rigidity, inducing cancer cells to become more invasive as well as more likely to metastasize. Indeed, longer collagen fibers in the matrix are associated with increased invasion and metastasis, as well as reduced survival, in mice.17
In addition, the abnormal ECM in tumors can affect cancer progression by activating normal stromal cells, such as macrophages and fibroblasts, that accelerate tumor growth and treatment resistance. These activated stromal cells further strengthen and stretch the ECM, causing a snowball effect.
The biochemical composition and organization of the ECM also influences tumor biology. Dysregulation of normal matrix signals can lead to tumor progression, characterized by excessive cell proliferation, immortality, enhanced migration, changes in metabolism, and evasion of the immune response. More research is needed to dissect the relationships between the ECM’s mechanical properties, forces, and cell signaling pathways.
Targeting the ECM
Because unchecked proliferation of cancer cells increases solid stress in the tumor, anticancer therapies should decrease the compressive forces in tumors and reopen collapsed blood and lymphatic vessels.11 This is exactly what happens when tumors are treated with certain doses of paclitaxel or docetaxel, two widely used cancer drugs. Shrinking tumors increases blood flow and allows more efficient fluid movement through the extravascular space, lowering the tumor interstitial fluid pressure in mouse models and in patients with breast cancer.5 However, cancer cells invariably develop resistance to treatment and begin to regrow, increasing solid stress again. As a result, other targets for reducing solid stresses are needed.
Because of its role in containing and concentrating the forces in a tumor, the collagen matrix within and around the tumor is another potential target for relieving tumor-related stresses. Indeed, solid stress in tumors can be reduced by drugs that selectively reprogram activated fibroblasts or modify the assembly of matrix components such as collagen and hyaluronan. In rodent studies, targeting these force-altering components in the tumor microenvironment has been shown to decrease solid stress, improve blood perfusion and drug delivery, and improve tumor response to chemotherapy and animal survival.6 We have found, for example, that injecting tumors with a collagen-digesting enzyme increases the diffusion of antibodies and viral particles and improves drug penetration in the tumor. Similarly, treatments that target transforming growth factor–beta (TGF-β), which controls the production of collagen by myofibroblasts, increase perfusion, improve the delivery of drugs of all sizes in mammary tumors, and improve treatment outcomes in mice.5
A class of drugs that is widely used to control blood pressure in hypertensive patients also blocks the TGF-β pathway. These drugs, known as angiotensin receptor 1 blockers, can reduce collagen production in and around the tumor by reducing the activity of TGF-β, as well as by blocking the function of connective tissue growth factor (CTGF), which is involved in stabilizing collagen and inducing resistance to chemotherapy.6Losartan and other angiotensin inhibitors reduce levels of collagen in various experimental models of fibrosis, and decrease renal and cardiac fibrosis in hypertensive patients. When given to mice with one of four different types of tumors characterized by high levels of cancer-associated fibroblasts (CAFs) and excess extracellular matrix—pancreatic ductal adenocarcinoma, breast cancer, sarcoma, and melanoma—losartan treatment caused a decrease in collagen content in a dose-dependent manner, enhanced penetration of nanoparticles into the tumor, and improved efficacy of diverse anticancer drugs. This is supported by a number of retrospective studies in patients with pancreatic, lung, and kidney cancers.6Researchers at Massachusetts General Hospital are now running a Phase 1/2 clinical trial to test losartan in pancreatic cancer patients.
http://www.the-scientist.com/images/April2016/forces_cancer_2.jpg
THE TUMOR ENVIRONMENT: The extracellular matrix and stromal cells within a tumor’s microenvironment influence the physical forces a tumor experiences. Left: The immunofluorescent image shows stromal cells (red and green) surrounding tumor cells (red cluster with blue nuclei); the cells were isolated from a mouse model of lung adenocarcinoma. Right: In this immunofluorescent image of triple-negative breast cancer, tumor cells (blue) are in close contact with matrix collagen (purple). Immune cells are labeled in red and green.VASILENA GOCHEVA, JACKS LAB, KOCH INSTITUTE AT MIT; DONGMEI ZUO, LABORATORY DR. MORAG PARK
Another potential cancer treatment target is hyaluronan, which is abundant in 20 percent to 30 percent of human tumors, most notably breast, colon, and prostate cancers. In addition to its role as a pressure-creating gel, hyaluronan can sequester growth factors and inhibit interstitial fluid movement within the tumor. Hyaluronidase, an enzyme that digests hyaluronan, reduces mechanical stress in tumors grown in mice.1 And San Diego–based Halozyme Therapeutics’s PEGPH20, a formulation of hyaluronidase coated with polyethylene glycol to enhance bioavailability, can decompress blood vessels and improve treatment outcome in genetically engineered mouse models of pancreatic ductal adenocarcinoma. Based on these studies, Halozyme researchers are now testing PEGPH20 in a randomized clinical trial of pancreatic cancer patients. Another matrix-altering drug is the widely-prescribed antidiabetic drug metformin, which has been shown to decrease collagen and hyaluronan levels in pancreatic tumors in obese mice and patients.7 Metformin is currently being tested in more than 200 clinical trials worldwide as a treatment for different types of cancer.
Clearly, tumors should be studied not only in light of their biochemical processes and genetic underpinnings, but also for the specific physical forces and mechanical properties that may influence progression. Understanding the physical microenvironment of tumors, as well as its interplay with the biochemical environment, is necessary to improve cancer detection, prevention, and treatment.
- T. Stylianopoulos et al., “Causes, consequences, and remedies for growth-induced solid stress in murine and human tumors,” PNAS, 109:15101-08, 2012.
- R.K. Jain, L.T. Baxter, “Mechanisms of heterogeneous distribution of monoclonal antibodies and other macromolecules in tumors: Significance of elevated interstitial pressure,” Cancer Res, 48:7022-32, 1988.
- R.K. Jain, “Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy,”Science, 307:58-62, 2005.
- R.K. Jain, “Antiangiogenesis strategies revisited: From starving tumors to alleviating hypoxia,”Cancer Cell, 26:605-22, 2014.
- R.K. Jain et al., “The role of mechanical forces in tumor growth and therapy,” Annu Rev Biomed Eng, 16:321-46, 2014.
- V.P. Chauhan et al., “Angiotensin inhibition enhances drug delivery and potentiates chemotherapy by decompressing tumour blood vessels,” Nat Commun, 4:2516, 2013.
- J. Incio et al., “Metformin reduces desmoplasia in pancreatic cancer by reprogramming stellate cells and tumor-associated macrophages,” PLOS ONE, 10:e0141392, 2015.
- M.A. Swartz, A.W. Lund, “Lymphatic and interstitial flow in the tumour microenvironment: linking mechanobiology with immunity,” Nat Rev Cancer, 12:210-19, 2012.
- H. Qazi et al., “Cancer cell glycocalyx mediates mechanotransduction and flow-regulated invasion,”Integr Biol, 5:1334-43, 2013.
- J.W. Song, L.L. Munn, “Fluid forces control endothelial sprouting,” PNAS, 108:15342-47, 2011.
Read Full Post »














































































 7). (e) Left: skin flap preparation after injection of Patent Blue V dye into the primary tumour (PT) showing the dye taken up into the tumour-draining LV and into the tumour-draining axillary lymph node (AxLN). The LV is adjacent to a blood vessel (BV). Right top panel: epifluorescence image of mCherry-tagged MDA-MB-231 tumour cells (TCs, red) that had spontaneously disseminated from orthotopic PT and were present in the tumour-draining LV that contained microspheres (green) and was adjacent to an autofluorescent BV. Right lower panel: corresponding maximum projection of multiphoton image. Scale bar, 100
7). (e) Left: skin flap preparation after injection of Patent Blue V dye into the primary tumour (PT) showing the dye taken up into the tumour-draining LV and into the tumour-draining axillary lymph node (AxLN). The LV is adjacent to a blood vessel (BV). Right top panel: epifluorescence image of mCherry-tagged MDA-MB-231 tumour cells (TCs, red) that had spontaneously disseminated from orthotopic PT and were present in the tumour-draining LV that contained microspheres (green) and was adjacent to an autofluorescent BV. Right lower panel: corresponding maximum projection of multiphoton image. Scale bar, 100

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}