Vitamin D debates
Larry H. Bernstein, MD, FCAP, Curator
LPBI

Article ID #196: Vitamin D debates. Published on 11/7/2015
WordCloud Image Produced by Adam Tubman
Vitamin D: Time for Rational Decision-Making
JoAnn E. Manson, MD, DrPH
Hello. This is Dr JoAnn Manson, professor of medicine at Harvard Medical School and Brigham and Women’s Hospital. I would like to talk with you about the vitamin D dilemma. The question of whether to screen routinely for vitamin D deficiency or to recommend high-dose vitamin D supplementation for our patients continues to be one of the most perplexing and vexing issues in clinical practice, and many clinicians are seeking guidance on these issues.
There appears to be a growing disconnect between the observational studies and the randomized clinical trials of vitamin D. For example, the observational studies are showing a fairly consistent relationship between low blood levels of vitamin D and an increased risk for heart disease, cancer, diabetes, and many other chronic diseases. Yet, the randomized clinical trials of vitamin D supplementation to date have been generally disappointing. This includes several randomized trials published over the past few months, including a meta-analysis[1] of randomized trials of vitamin D supplementation showing minimal, if any, benefit in terms of lowering blood pressure; a trial[2] of high-dose vitamin D supplementation showing no clear benefit for muscle strength, bone mineral density, or even the risk for falls; and, most recently, a randomized trial[3] of vitamin D supplementation with and without calcium showing no clear benefit in reducing the risk for colorectal adenomas. The latter trial was very recently published in the New England Journal of Medicine.
The Institute of Medicine (IOM)[4] and the US Preventive Services Task Force[5] do not endorse routine universal screening for vitamin D deficiency. They also recommend more moderate intakes [of vitamin D]. For example, the IOM recommends 600-800 IU a day for adults and also recommends avoiding daily intakes above 4000 IU, which has been set as the tolerable upper intake level.
However, it is important to keep in mind that these are public health population guidelines for a generally healthy population, and they by no means preclude individual decision-making by the clinician in the context of a patient who may have health conditions or risk factors that would indicate a benefit from targeted screening for vitamin D deficiency or higher-dose supplementation. For example, some patients may have higher vitamin D requirements. This may include patients with bone health problems (osteoporosis, osteomalacia) or poor diets, those who spend minimal time outdoors, those with malabsorption syndromes, or those who take medications that may interfere with vitamin D metabolism (glucocorticoids, anticonvulsant medications, and antituberculosis drugs). Therefore, overall, there is a role for individualized decision-making, in terms of screening for vitamin D deficiency in patients who have bone health problems or special risk factors, and even treating with higher doses of vitamin D, which may go above 4000 IU a day in patients who have higher requirements.
In the next several years, large-scale, randomized trials of vitamin D supplementation, including high-dose vitamin D supplementation, will be completed—and these results will be published. They will help to inform clinical decision-making, so stay tuned for those results.
Thank you so much for your attention. This is JoAnn Manson.
References
- Beveridge LA, Struthers AD, Khan F, et al. D-PRESSURE Collaboration. Effect of vitamin D supplementation on blood pressure: a systematic review and meta-analysis incorporating individual patient data. JAMA Intern Med. 2015;175:745-754. Abstract
- Hansen KE, Johnson RE, Chambers KR, et al. Treatment of vitamin D insufficiency in postmenopausal women: a randomized clinical trial. JAMA Intern Med. 2015;175:1612-1621. Abstract
- Baron JA, Barry EL, Mott LA, et al. A trial of calcium and vitamin D for the prevention of colorectal adenomas. N Engl J Med. 2015;373:1519-1530. Abstract
- Institute of Medicine. Dietary Reference Intakes for Calcium and Vitamin D. Washington, DC: National Academies Press; 2011. http://iom.nationalacademies.org/Reports/2010/Dietary-Reference-Intakes-for-Calcium-and-Vitamin-D.aspx Accessed October 28, 2015.
- US Preventive Services Task Force. Final Recommendation Statement: Vitamin D Deficiency: Screening, 2014.http://www.uspreventiveservicestaskforce.org/Page/Document/RecommendationStatementFinal/vitamin-d-deficiency-screening Accessed October 28, 2015.
Isn’t there much more to this than the debates entail?
The vitamin D hormone and its nuclear receptor: molecular actions and disease states.
Vitamin D plays a major role in bone mineral homeostasis by promoting the transport of calcium and phosphate to ensure that the blood levels of these ions are sufficient for the normal mineralization of type I collagen matrix in the skeleton. In contrast to classic vitamin D-deficiency rickets, a number of vitamin D-resistant rachitic syndromes are caused by acquired and hereditary defects in the metabolic activation of the vitamin to its hormonal form, 1,25-dihydroxyvitamin D3 (1,25(OH)2D3), or in the subsequent functions of the hormone in target cells. The actions of 1,25(OH)2D3 are mediated by the nuclear vitamin D receptor (VDR), a phosphoprotein which binds the hormone with-high affinity and regulates the expression of genes via zinc finger-mediated DNA binding and protein-protein interactions. In hereditary hypocalcemic vitamin D-resistant rickets (HVDRR), natural mutations in human VDR that confer patients with tissue insensitivity to 1,25(OH)2D3 are particularly instructive in revealing VDR structure function relationships. These mutations fall into three categories: (i) DNA binding/nuclear localization, (ii) hormone binding and (iii) heterodimerization with retinoid X receptors (RXRs). That all three classes of VDR mutations generate the HVDRR phenotype is consistent with a basic model of the active receptor as a DNA-bound, 1,25(OH)2D3-liganded heterodimer of VDR and RXR. Vitamin D responsive elements (VDREs) consisting of direct hexanucleotide repeats with a spacer of three nucleotides have been identified in the promoter regions of positively controlled genes expressed in bone, such as osteocalcin, osteopontin, beta 3-integrin and vitamin D 24-OHase. The 1,25(OH)2D3 ligand promotes VDR-RXR heterodimerization and specific, high affinity VDRE binding, whereas the ligand for RXR, 9-cis retinoic acid (9-cis RA), is capable of suppressing 1,25(OH)2D3-stimulated transcription by diverting RXR to form homodimers. However, initial 1,25(OH)2D3 liganding of a VDR monomer renders it competent not only to recruit RXR into a heterodimer but also to conformationally silence the ability of its RXR partner to bind 9-cis RA and dissociate the heterodimer. Additional probing of protein-protein interactions has revealed that VDR also binds to basal transcription factor IIB (TFIIB) and, in the presence of 1,25(OH)2D3, an RXR-VDR-TFIIB ternary complex can be created in solution. Moreover, for transcriptional activation by 1,25(OH)2D3, both VDR and RXR require an intact short amphipathic alpha-helix, known as AF-2, positioned at their extreme C-termini. Because the AF-2 domains participate neither in VDR-RXR heterodimerization nor in TFIIB association, it is hypothesized that they contact, in a ligand-dependent fashion, transcriptional coactivators such as those of the steroid receptor coactivator family, constituting yet a third protein-protein interaction for VDR. Therefore, in VDR-mediated transcriptional activation, 1,25(OH)2D3 binding to VDR alters the conformation of the ligand binding domain such that it: (i) engages in strong heterodimerization with RXR to facilitate VDRE binding, (ii) influences the RXR ligand binding domain such that it is resistant to the binding of 9-cis RA but active in recruiting coactivator to its AF-2 and (iii) presents the AF-2 region in VDR for coactivator association. The above events, including bridging by coactivators to the TATA binding protein and associated factors, may position VDR such that it is able to attract TFIIB and the balance of the RNA polymerase II transcription machinery, culminating in repeated transcriptional initiation of VDRE-containing, vitamin D target genes. Such a model would explain the action of 1,25(OH)2D3 to elicit bone remodeling by stimulating osteoblast and osteoclast precursor gene expression, while concomitantly triggering the termination of its hormonal signal by inducing the 24-OHase catabolizing enzyme.
Classic nutritional rickets is caused by the simultaneous deprivation of sunlight exposure and dietary vitamin D. As depicted in Fig. 1, the pathways comprising the metabolic activation of the vitamin to its hormonal form and subsequent functions in target tissues present a number of additional steps where defects elicit vitamin D-resistant rachitic syndromes. Two of these disorders involve the inadequate bioactivation of 25-hydroxy¬ vitamin D3 (25(OH)D3) to 1,25-dihydroxyvitamin D3 (l,25(OH)2D3) by the kidney as catalyzed by the 1-OHase enzyme (Fig. 1).
Figure 1 Bioactivation of vitamin D3 and actions of the 1,25(OH)2D3 hormonal metabolite on intestine, bone and kidney, along with related rachitic syndromes. The production of 1,25(OH)2D3 is depicted in the lowet portion and its functions on mineral ttansport in target cells ate pictured in the upper portion. Defects eliciting rachitic syndromes ate boxed, with the televant mutated gene and chromosomal location denoted where appropriate
Acquired chronic renal failure results in renal rickets and secondary hyperparathyroidism (renal osteodystrophy) when the compromising of renal mass reduces 1-OHase activity (Haussier oc McCain 1977). The etiology of pseudo-vitamin D-deficiency rickets (PDDR) apparently involves a hereditary defect in the gene coding for the 1-OHase enzyme (Labuda et al. 1992). Interestingly, the PDDR locus is resolvable from that of the vitamin D receptor (VDR) but maps very close to it on chromosome 12 in the 12ql3—14 region (Labuda et al. 1992). Recently, a cDNA was cloned for the rat 1-OHase (St-Arnaud et al. 1996) and it is expected that the human renal 1-OHase gene will soon be cloned and its chromosomal location determined. The likelihood that both the gene encoding the enzyme that generates the l,25(OH)2D3 hormone and the cognate hormone receptor gene lie in close proximity on chromosome 12 invites speculation about the evolution of the vitamin D ligand receptor system. The traditional actions of vitamin D, via its l,25(OH)2D3 hormonal metabolite, are to effect calcium and phosphate homeostasis to ensure the deposition of bone mineral on type I collagen matrix (summarized in Fig. 1).
Figure 1 Bioactivation of vitamin D3 and actions of the 1,25(OH)2D3 hormonal metabolite on intestine, bone and kidney, along with related rachitic syndromes. The production of 1,25(OH)2D3 is depicted in the lowet portion and its functions on mineral ttansport in target cells ate pictured in the upper portion. Defects eliciting rachitic syndromes ate boxed, with the televant mutated gene and chromosomal location denoted where appropriate
l,25(OH)2D3 stimulates intestinal calcium and phosphate absorption, bone calcium and phosphate résorption, and renal calcium and phosphate reabsorption, all resulting in a sufficient CaP04 ion product to precipitate hydroxyapatite. Failure to achieve normal bone mineral accretion by these mechanisms leads to rachitic syndromes. Recently, a breakthrough has occurred in our understand¬ ing of what was originally known as hypophosphatemic vitamin D-resistant rickets, a familial disorder of renal phosphate wasting more appropriately referred to as dominant X-linked hypophosphatemic (HYP) rickets (Fig. 1). The gene defect responsible for HYP rickets has been fine mapped in the Xp22T region, harboring a gene identified as PEX, or phosphate regulating gene with homologies to endopeptidases located on the X-chromosome (Francis et al. 1995). One hypothesis is that PEX codes for an endopeptidase that apparently correctly processes a peptide precursor to yield a novel, as yet unidentified, phosphate retaining hormone. The normal function of this hormone may be to oppose the action of parathyroid hormone (PTH) and stimulate phosphate reabsorption by the renal tubule by inducing the Na -phosphate cotransporter. However, the existence of tumor-induced osteomalacia, an acquired disorder that closely resembles the phosphate wasting of HYP rickets and is characterized by low circulating l,25(OH)2D3 (Parker et al. 1981), combined with renal cross-transplantation (Nesbitt et al. 1992) and parabiosis (Meyer et al. 1989) studies in normal and hyp mice, indicates strongly that the HYP phenotype is caused by excessive amounts of a phosphaturic hormone in the circulation. This humoral peptide is distinct from PTH and has been named phosphatonin (Cai et al. 199A, Econs & Drezner 1994). Thus, instead of PEX mutations result¬ ing in insufficient generation of a novel phosphate retaining peptide, they may instead elicit the appearance of abnormally high circulating levels of phosphatonin, with the normal role of the PEX gene product postulated to be the proteolytic inactivation of this phosphaturic principle. Most germane to the vitamin D endocrine system is the fact that serum l,25(OH)2D3 levels are inappropriately low for the prevailing phosphate concentrations in HYP rickets and patients can be cured with a therapeutic combination of phosphate and l,25(OH)2D3 (Harrel et al. 1985). Because it is well known that hypophosphatemia stimulates l,25(OH)2D3 production (Hughes et al. 1975), the PEX/phosphatonin system might constitute yet another regulatory loop in maintaining normal phosphate homeostasis. One could hypothesize that under hypo- phosphatemic conditions, when l,25(OH)2D3 levels are elevated, the sterol hormone not only increases intestinal phosphate absorption (Fig. 1) and suppresses PTH synthesis (DeMay et al. 1992) to conserve phosphate, but also induces the PEX gene product (Rowe et al. 1996) to cleave phosphatonin and further promote renal phosphate reclamation. l,25(OH)2D3 is primarily recognized as a calcémic hormone, perhaps due to the abundance of dietary phosphate, or because calcium homeostasis is more vitamin D-dependent than the regulation of extracellular phos¬ phate. Regardless of the mechanism, traditional vitamin D-deficiency and clinically significant defects in the vitamin D receptor lead invariably to hypocalcemia and secondary hyperparathyroidism, with phosphate being somewhat less affected. As illustrated in Fig. 1, target tissue insensitivity to l,25(OH)2D3 is known as hereditary hypocalcémie vitamin D-resistant rickets (HVDRR) and is caused by defects in the gene on chromosome 12 coding for the VDR. A review of the etiology of HVDRR and the natural mutations in the VDR that confer tissue insensitivity and clinical resistance to l,25(OH)2D3 is particularly instructive in illuminating the physiologic relevance of the l,25(OH)-,D3-VDR hormone-receptor complex as well as structure/function relationships in the receptor itself.
Natural mutations in the nuclear vitamin D receptor Clinically significant hereditary hypocalcémie vitamin D-resistant rickets is an autosomal recessive disorder resulting in a phenotype characterized by severe bowing of the lower extremities, short stature and, often, alopecia (Rut et al. 199A). The serum chemistry in HVDRR includes frank hypocalcemia, secondary hyperpara¬ thyroidism, elevated alkaline phosphatase, variable hypophosphatemia and markedly increased l,25(OH)2D3. The symptoms of HVDRR, with the exception of alopecia, mimic classic vitamin D-deficiency rickets, suggesting that VDR not only mediates the bone mineral homeostatic actions of vitamin D but may also participate in the differentiation of hair follicles in utero. Recently, VDR knockout mice have been created (Yoshizawa et al. 1996), revealing apparently normal hétérozygotes but severely affected homozygotes (VDR-/-), 90% ofwhich die within 8—10 weeks. Surviving mice lose their hair and possess low bone mass, hypocalcemia, hypophosphatemia and 10-fold elevated l,25(OH)2D3 coincident with extremely low 24,25(OH)2D3. All of these parameters in the VDR knockout mouse mimic the phenotype of patients with HVDRR, confirming that VDR normally mediates all of the bone mineral regulating functions of vitamin D. Interestingly, although natural point mutations in other receptors related to VDR, such as thyroid hormone receptor ß (TRß) (Collingwood et al. 1994), are charac¬ terized by dominant negative receptors that generate the thyroid hormone resistant phenotype in the heterozygotic context, no natural, dominant negative mutations have yet been identified in HVDRR patients (Whitfield et al. 1996). Thus, all HVDRR cases studied to date are homozygous for the particular VDR mutation.
Figure 2 Natural mutations in the human vitamin D receptor leading to 1,25(OH)2D¡ hormone resistance. See text for details and citations. N37, K91 and E92 are not sites of VDR natural mutations, but are so designated because they ate heterodimerization contacts that lie within the DNA binding domain (Hsieh et al. 1995, Rastinejad et al. 1995). The eight cysteine residues (C) that tetrahedrally coordinate two zinc atoms in the finger sttucture are also denoted.
Figure 2 illustrates a number of point mutations in VDR that have been detected in HVDRR patients (reviewed in Rut et al. 199A, Haussler et al. 1995). Three of these genetic alterations result in nonsense mutations that introduce stop codons in VDR (K73stop, Q152sfo|> and Y295stop), creating truncated VDRs that lack both hormone- and DNA-binding (heterodimerization) capacities and are associated with unstable mRNAs. More revealing are the series of missense mutations (Fig. 2) that can be classified according to three of the basic molecular functions of VDR: (i) DNA binding/nuclear localization by the N-terminal zinc finger region, (ii) l,25(OH)2D3 hormone binding by the C-terminal domain and (iii) heterodimerization with retinoid X receptors (RXRs) through subregions of the C-terminal domain. As depicted schematically in Fig. 2 and discussed in detail later, VDR is a ligand-dependent transcription factor that controls gene expression by heterodimerizing with RXR and associating specifically with vitamin D responsive elements (VDREs) in target genes. Since VDR is a member of the steroid, retinoid, thyroid hormone receptor superfamily, and belongs to the VDR/retinoic acid receptor (RAR)/TR subfamily of RXR heterodimerizing species (Haussler et al. 1991), it is reasonable to draw from data on RAR and TR for comparison with VDR.
The greatest number of VDR natural mutations char¬ acterized to date are localized to the DNA binding, zinc finger region (Fig. 2). The first two discovered, G33D and R73Q (Hughes et al. 1988), reside at the ‘tips’ of the fingers and affect charge—charge interactions between VDR and the phosphate backbone of DNA. When viewed in toto, the zinc finger region mutations in HVDRR (Fig. 2) have the following two general prop¬ erties: (i) they occur in residues conserved across the entire nuclear receptor superfamily and (ii) most lie within -helices on the C-terminal side of the first and second fingers which are intimately involved in DNA base recognition and phosphate backbone contacts respectively (Rastinejad et al. 1995). These observations suggest that many of the clinically significant mutations in VDR which are still compatible with life may not greatly perturb the fundamental structure of the DNA binding domain of the receptor, but instead compromise its ability to recog¬ nize DNA with specificity and high affinity. Whether HVDRR cases with mutations in zinc finger region residues unique to VDR will be uncovered depends upon the properties of such alterations, which could range from innocuous to lethal.
Mutations located within the hormone binding domain of VDR also elicit the HVDRR phenotype (Fig. 2), including R274L (Kristjansson et al. 1993) and H305Q (Malloy et al 1995). Transcriptional activation by R274L and H305G VDR is attenuated as a result of inefficient l,25(OH)2D3 binding, ranging from severe in the case of R274L to a modest increase in Kd for H305Q. In both instances, transcriptional activation is restored when the dose of l,25(OH)2D3 is raised to pharmacologie levels (10 m) in transfection experiments (Kristjansson et al. 1993, Malloy et al. 1995). Our laboratory has recently characterized two novel VDR hormone binding domain mutations in HVDRR patients, I314S and R391C, that significantly affect the heterodimerization of VDR with RXR (Whitfield et al. 1996). Both of these C-terminal replacements (Fig. 2), however, do display some degree of what may be a hormone binding deficit, a phenomenon not observable in typical in vitro ligand binding kinetic assays at 4 °C. Thus, only at 37 °C in intact cells do R391C and I314S exhibit apparent slight and significant impairment of l,25(OH)2D3 high affinity retention respectively (Whitfield et al. 1996). Further, the two mutations in question are situated in or adjacent to heptad repeats (Fig. 2), hypothetical coiled-coil-like structures that were originally proposed to participate in the heterodimerization of VDR, RAR, and TR with RXR (Forman & Samuels 1990, Nakajima et al. 1994). Consist¬ ent with this concept, both R391C and I314S VDRs do not bind RXR with normal affinity when assayed in vitro, with the greatest impairment of heterodimerization occur¬ ring with R391C (affinity reduced by one order of magnitude) (Whitfield et al. 1996). Additional evidence supporting blunted RXR heterodimerization by these two mutant VDRs is provided by transfection experiments in restored to that of normal fibroblasts when fibroblasts from patients harboring either the R391C or the I314S mutation are cotransfected with exogenous RXR. Yet this apparent RXR rescue of the mutated VDRs requires approximately 10-fold elevated l,25(OH)2D3 doses com¬ pared with the response to hormone in normal fibroblasts (Whitfield et al. 1996). This latter observation reveals that the hormone binding and heterodimerization functions of VDR are not entirely separable, an aspect which is also apparent from fundamental biochemical analysis of the hormone dependency of VDR-RXR heterodimer binding to VDREs as discussed in detail below.
Understanding the molecular properties of natural VDR mutations in HVDRR allows us to comprehend why the patients respond differentially to therapy with massive doses of l,25(OH)2D3, or suitable analogs. For example, cases with zinc finger region aberrations are unresponsive to the hormone because DNA binding is precluded by the absence of structural complemen¬ tarity between VDR and the VDRE, regardless of the l,25(OH)2D3 liganding or heterodimerization of the receptor in solution. Conversely, patients harboring mutations in the hormone binding/heterodimerization domain can be responsive to pharmacologie doses of l,25(OH)2D3 or analogs, even though the hormone already is increased in the circulation because of the hypocalcemia caused by tissue insensitivity. For example, patient I314S was essentially cured by excess vitamin D metabolite, indicating that compensating for the hormone binding deficit was able to override the milder heterodimerization defect and allow sufficient VDRE binding by the VDR-RXR heterodimer. Conversely, patient R391C responded only modestly to treatment with excess l,25(OH)2D3 analog, presumably because the fundamental heterodimerization defect could not be overcome and therefore normal VDRE binding could not be achieved (Whitfield et al. 1996).
The final insights gained from the natural VDR mutations summarized in Fig. 2 are structural in nature. We have discussed previously that the zinc finger mutations are confined to absolutely conserved residues. In the crystal structure of the DNA binding domain heterodimers of RXRa and TRß (Rastinejad et al. 1995), the lysine and arginine residues corresponding to K45 and R50 in human VDR (hVDR) make direct base contacts with DNA, while the arginines corresponding to R73 and R80 in hVDR make direct DNA phosphate backbone contacts. That mutations in these four residues are clinically important in the etiology of HVDRR argues for structural congruity between the VDR finger region and that of TR. Rastinejad et al. (1995) have extended this assumption to include a modeling of RXR-TR vs RXRVDR bound to DNA which accommodates the fact that TR binds as a heterodimer to a direct hexanucleotide repeat spaced by four nucleotides (DR+4), while VDR binds as a heterodimer to a similar set of half elements spaced by three nucleotides (DR+3). In addition to verifying the common protein-DNA interfaces, their modeling predicts that hVDR residues N37 in the first finger and K91/E92 C-terminal of the second finger (see Fig. 2) engage in heterodimeric contacts with residues in the second zinc finger ofRXR to form effectively a stable, DNA-supported heterodimer. Indeed, recent site-directed mutational studies (Hsieh et al. 1995) indicate that the alteration ofK91 and E92 in hVDR in fact grossly reduces transactivation while moderately attenuating hetero¬ dimerization and DNA binding, thus confirming the importance of K91 and E92. An additional surprising finding was that the K91/E92 double mutant manifested dominant negative characteristics (Hsieh et al. 1995), distinguishing it from the natural HVDRR replacements discussed above. Apparently, the K91/E92 mutant VDR is able to bind DNA sufficiently through its native zinc finger and strong heterodimerization function in the ligand binding domain such that it can block binding by wild type receptor, but is rendered inactive in stimulating transcription because of a presumed conformational per¬ turbation initiated by unstable or improper alignment of the heterodimer on the VDRE.
Based upon recently reported X-ray crystal structures of the ligand binding domains of ligand-occupied hRARy (Renaud et al. 1995), agonist-occupied rat TRa, (Wagner et al. 1995) and unoccupied, but dimeric hRXRa (Bourguet et al. 1995), it is also possible to incorporate the HVDRR mutations in the hormone binding domain (Fig. 2) into a hypothetical structural context. Figure 3 constitutes a schematic compilation of the existing crystallographic data and compares them with natural and artificially generated mutations in hVDR. At the top of Fig. 3, the residue numbers for VDR in the ligand binding domain appear in relation to the older heptad repeat nomenclature (heptads 1—9, dotted boxes). At least some of these heptads, particularly heptads 4 and 9, are thought to facilitate heterodimerization (Nakajima et al. 1994). The El region is a highly conserved area that supports heterodimerization (Whitfield et al. 1995è). The helices depicted schematically in Fig. 3 (open boxes) are those determined for hRARy; this general pattern of -helices and ß-strands (solid boxes) appears to be well conserved across the TR, RAR and RXR members of the subfamily crystallized thus far (Bourguet et al. 1995, Renaud et al. 1995, Wagner et al. 1995). Although the heterodimerization domains have yet to be elucidated by structural analysis, the homodimerization domain of RXR is comprised of helices 7, 9 and 10 (Fig. 3 and Bourguet et al. 1995). Flanking the dimerization region are clusters of ligand binding contacts, shown for RAR and TR in Fig. 3, which paint a picture of hormone binding involving helices 3, 5, 11 and 12 plus portions of helices 6 and 7 along with their intervening loop, as well as the loop between ß-strands 1 and 2.
Figure 3 Hormone binding (R274L and H305Q) and heterodimerization (I314S and R391C) natural mutations in VDR that confer the HVDRR phenotype are positioned in the context of retinoid and thyroid hormone receptor subfamily ligand binding domain structures. See text for details and citations.
As summarized in Fig. 3 and discussed by Whitfield et al. (1995a, 1996), a number of artificially generated mutants in hVDR support the con¬ cept that the dimerization and honnone binding regions in VDR are well aligned with those in RXR, RAR and TR. Of even greater interest and relevance to the present monograph, the four clinically important hVDR mutants under consideration correspond to pertinent locations in the known structures of the retinoid and thyroid hormone receptor ligand binding domains. We postulate that this general structural organization represents that of the VDR ligand binding domain. As shown in Fig. 3, the pure hormone binding mutant hVDRs, namely R274L and H305Q, are located precisely within ligand clusters in helix 5 and in the loop between helix 6 and 7 respectively. I314S, which endows hVDR with combined defects in hormone retention and heterodimerization, lies within helix 7 at a presumed interface of ligand binding and dimerization activities of the receptor (Fig. 3). Finally, R391C is positioned well within the helix 10 dimerization surface, but not far removed from C-terminal ligand binding contacts that are likely influenced by replacement of this amino acid in hVDR. Thus, at least within the context of the assumed structural organization of VDR derived from that of other subfamily members, the I314S and R391C mutations are situated precisely where they would be predicted to lie, given the biological properties of the mutant receptors and the phenotype of the patients. These results not only have profound implications con¬ cerning the putative structure of VDR in relation to its closest relatives, but prove unequivocally that the calcémic actions of l,25(OH)2D3 are mediated by the vitamin D receptor, existing as a l,25(OH)2D3-liganded heterodimer with RXR that is bound to DNA.
Physiology and cellular actions of l,25(OH)2D3
In order to delineate the physiologic roles for the vitamin D hormone, it is appropriate first to place the VDR mediator into the context of vitamin D metabolism and cellular actions. Figure 4 summarizes the integration of vitamin D metabolism and cellular actions introduced in Fig. 1, with physiologic regulatory events now super¬ imposed on the metabolic pathway and the inclusion of an expanded list of physiologic actions for the 1,25( )2 4 hormone. The conversion of vitamin D3 to 25(OH)D3 by the liver is a constitutive metabolic step, followed by the 1-hydroxylation of25(OH)D3 to l,25(OH)2D3, a reaction under exquisite control (Haussler & McCain 1977). When blood calcium is low, activation of this latter step occurs, either as a result of the hypocalcémie state per se, or in response to elevated PTH, each of which serves indepen¬ dently to enhance renal 1-OHase activity. Low phosphate is also capable of separately upregulating the 1-OHase enzyme. To limit activation, the hormonal product, l,25(OH)2D3, effects an ultra-short feedback loop to suppress its own biosynthesis in the kidney and also represses PTH synthesis to remove the peptide hormone stimulus of the 1-OHase via a longer feedback loop (Fig. 4). However, the dominant negative feedback controls of 1-OHase activity appear to result from the concerted actions of l,25(OH)2D3 to stimulate bone mineral résorption and to promote intestinal calcium and phosphate absorption, which together elicit an increase in blood calcium and phosphate levels, each of which down-regulates the 1-OHase.
Figure 4 Vitamin D metabolism and cellulat actions, mediated by the VDR-RXR heterodimer binding to a VDRE
The process by which l,25(OH)2D3 causes bone remodeling is complex, involving stimulation of osteoclast differentiation and osteoblastic production of osteopontin, both of which activate résorption in part through the recognition of bone matrix osteopontin by osteoclast surface avß3-integrin. The résorption effect is supported by l,25(OH)2D3-elicited suppression of bone formation via the induction of osteocalcin and the repression of type I collagen. This latter insight that the normal function of osteocalcin is to curtail bone matrix formation arises from the creation of osteocalcin knockout mice (Ducy et al. 1996). In addition to stimulating the transcription of bone-related genes such as osteopontin and osteocalcin, the l,25(OH)2D3 hormone also induces its own eatab¬ olism in kidney as well as other target tissues like bone by enhancing the expression of the vitamin D-24-OHase enzyme. 24-Hydroxylation of l,25(OH)2D3 is the first step in deactivating the hormone, which is eventually metabolized by side chain cleavage to calcitróle acid (Haussler 1986). Thus, the synthesis of l,25(OH),D3 is not only governed by feedback mechanisms that sense l,25(OH)2D3, calcium, PTH and phosphate concentrations, but the hormone induces the termination of its own signal in target tissues, qualifying l,25(OH)2D3 as a bonafide hormone by any definition.
Figure 4 Vitamin D metabolism and cellulat actions, mediated by the VDR-RXR heterodimer binding to a VDRE
As introduced in the section on HVDRR, mediation of the cellular functions of l,25(OH)2D3 requires that VDR bind the hormonal ligand specifically and with high affinity (Fig. 4). Upon such binding, VDR becomes hyperphosphorylated (Jurutka et al. 1993, Haussler et al. 1994) and recruits RXR into a hetero¬ dimeric complex that binds strongly to DNA (Fig. 4). The l,25(OH)2D3-hganded RXR-VDR heterocomplex selectively recognizes VDREs in the promoter regions of positively controlled genes such as osteocalcin (MacDonald et al. 1991), osteopontin (Noda et al. 1990), vitamin D-24-OHase (Ohyama et al. 199A) and ß3-integrin (Cao et al. 1993). Negative VDREs (Haussler et al. 1995) exist in the 5′-regions of the genes for type I collagen (Pavlin et al. 199A), bone sialoprotein (Li & Sodek 1993), PTH (DeMay et al. 1992) and PTH-related peptide (Falzon 1996, Kremer et al. 1996). The mechanisms whereby VDR accomplishes positive and negative control of DNA transcription after VDRE association are not well under¬ stood, although substantial progress has been made in comprehending the stimulation of transcription as detailed in later sections of this article. Moreover, as summarized in Fig. 5, a number of VDREs have been definitively characterized. The prototypical VDBJS is found in the osteocalcin gene, consisting of an imperfect direct repeat of hexanucleotide estrogen responsive element (ERE)-like, half-sites with a spacer of three nucleotides (DR+3). Classic EREs possess a central GT core at positions 3 and 4 of the hexanucleotide, but this feature is only partially conserved in the six natural positive VDREs listed in Fig. 5. There is, however, absolute conservation of the A in position 6 of the 5′ half-element and of the G at position 2 of the 3′ half-element. A preliminary working consensus for the positive VDRE can be derived from these natural VDREs (see boxed sequence in Fig. 5). This generaliz¬ ation is supported, in part, by PCR experiments that were designed to select, from random oligonucleotides, the highest affinity DNA ligand for the RXR-VDR heterodimer (Nishikawa et al. 1994, Colnot et al. 1995).
Figure 5 Natural vitamin D responsive elements (DR+3s) in genes positively tegulated by l,25(OH)2D3. The consensus VDREs are based on either sequence comparisons (boxed) or a selection of random sequences (at bottom).
The random selection process yields an identical VDRE 5′ half-element of GGGTCA (Fig. 5, bottom), which is also a preferred RXR target when RXR homodimers bind to DNA (Yang et al. 1995). This observation is in concert with the conclusion (Jin & Pike 1996) that, with respect to association ofRXR-VDR with VDREs, RXR lies on the 5′ half-element whereas VDR is situated on the 3′ half-element. Examination of both consensus sequences suggests that the G at position 3 of the spacer is important in VDR binding, a deduction consistent with the finding (MacDonald et al. 1991) that this base is partially protected by RXR-VDR in methylation interference assays. How¬ ever, interesting differences arise when one compares the most frequently encountered 3′ half-element bases in natural VDREs, namely the GGGGCA composite which actually occurs in human osteocalcin, with the GGTTCA random consensus selection for the 3′ half-element (Fig. 5). Clearly, GGTTCA represents a potent VDR binding site, a supposition that is bolstered by the fact that osteopontin, which possesses a perfect DR+3 of GGTTCA, is the highest affinity VDRE we have tested (data not shown). Intriguingly, Ts at positions 3 and 4 in the 3′ VDR half-site occur infrequently in the balance of natural VDREs (Fig. 5). The paucity of Ts in the 3′ half-element could be related to a need for varying potency of VDREs in regulated genes, or may even provide for a repertoire of different VDR conformations that could be induced by contact with distinct 3′ half-site core sequences. This postulated range of VDR conforma¬ tions might endow the receptor with the ability to recruit a variety of different coactivators and corepressors, or even to favor the binding of one vitamin D metabolite ligand over another. Irrespective of the above considerations, it is evident that the primary VDRE is a DR+3 recognition site in DNA that directs the VDR to the promoter region of l,25(OH)2D3 regulated genes, ultimately altering the functions of target cells as a result of transcriptional control of gene expression.
Significance of lipophilic ligands in the association of RXR-VDR with DNA
Dimeric complexes are a feature commonly employed in the regulation of eukaryotic transcriptional systems. This process of protein dimerization often will generate novel heterodimeric complexes which display highly cooperative binding to DNA as well as an altered target sequence specificity (Glass 1994). Among the classical steroid hormone receptors, dimerization results in the formation of symmetrical homodimeric protein complexes on palindromic DNA half sites. Dimerization has been shown to be mediated in part by residues within the DNA binding domain of the receptor (Luisi et al. 1991) and is enhanced by residues within the ligand binding domain (Falwell et al. 1990). The other subfamily of nuclear hormone receptors, including VDR, TR and RAR, apparently binds with highest affinity to direct repeat elements either as homodimers or, more commonly, as heterodimers with RXR (Kliewer et al. 1992). In both subgroups of nuclear receptors, protein-protein interactions serve to align the DNA binding domains so that they are optimally positioned to bind to their specific DNA target sequences (Kurokawa et al. 1993, Perlmann et al. 1993, Rastinejad et al. 1995). The ligand binding region of these receptors is multifunctional, in that this domain not only binds the cognate ligand, but also it possesses a dimerization surface as well as the ligand-dependent transactivation function, AF-2 (Gronemeyer 1991, Chambón 1994). The dimerization surface consists of packed helices which are stabilized by hydrophobic heptad repeats interspersed throughout the structure. Ligand apparently can influence different functional components, including the dimerization interface, and the activating AF-2 domain (Renaud et al. 1995, Wagner et al. 1995). Therefore, a likely role for ligand is to regulate the association and dissociation of dimeric protein complexes and hence regulate specific binding to DNA target sequences.
In this regard the following three questions remain regarding l,25(OH)2D3-mediated control of positively regulated genes: (i) does VDR bind as a homodimer (Freedman et al. 1994, Nishikawa et al. 1994) as well as a heterodimer to DR+3 VDREs? (ii) What is the effect of the l,25(OH)2D3 ligand on VDR or VDR-RXR binding to VDREs? (iii) What role does 9-cis retinoic acid, the RXR ligand, play m RXR-VDR binding to VDREs and enhanced transcription of l,25(OH)2D3-responsive genes? It is generally accepted that TR forms homodimers as well as heterodimers with RXR on thyroid hormone responsive elements (TREs), although recent data suggest that the TR homodimer, when unoccupied by thyroid hormone, operates as a repressor of transcription (Chin & Yen 1996, Schulman et al. 1996). Thyroid hormone is proposed to dissociate TR homodimers to facilitate TRRXR heterodimerization on the TRE and stimulate transcription. In contrast, RAR does not appear to be capable of forming homodimers on DR+5 retinoic acid responsive elements (RAREs) (Perlmann et al. 1996), instead cooperating exclusively with RXR in RARE association and vitamin A metabolite-responsive transcrip¬ tion. When present in excess in gel mobility shift DNA binding assays in vitro, both TR and RAR display RXR heterodimeric association with their respective hormone responsive elements (HREs) in the absence of added lipophilic ligand. These in vitro studies are consistent with immunocytochemical data indicating that, unlike classic steroid honnone receptors that reside in the cytoplasm complexed with Hsp-90 and other proteins in their unoccupied state, unliganded TR, RAR and VDR (Clemens et al. 1988) exist in the nucleus in general association with DNA. These findings have led to the dogma that ligand is not required for TR, RAR and VDR to associate with target HREs. Indeed, we have observed that addition of 260 ng baculovirus-expressed hVDR to a gel shift reaction generates weak homodimeric VDR as well as strong VDR-RXR-heterodimeric binding to a rat osteocalcin VDRE probe, both of which are independent of the presence of l,25(OH)2D3 (Nakajima et al. 1994). However, in vivo footprinting experiments (Blanco et al. 1996, Chen et al. 1996) have led to the conclusion that, at least in the case of RAR-RXR heterodimers, RAR ligands are required for RARE binding. We, therefore, sought to devise an in vitro gel shift assay that would more accurately reflect the in vivo situation, primarily consisting of the use of physiologic salt (0-15 m KCl) concentrations and limited amounts of partially purified, baculovirusexpressed VDR and RXRs (Thompson et al. 1997). Utilizing this assay, we have addressed the three questions regarding VDR/RXR listed above, namely heterodimer versus homodimer, the potential role of l,25(OH)2D3 and the effect of 9-cis retinoic acid (9-cis RA).
When 20 ng VDR (~ 10 nM) or 20 ng VDR plus 20 ng RXR are incubated with either the rat osteocalcin or mouse osteopontin VDREs (see Fig. 5), no DNA-bound homodimeric VDR species is apparent, but a VDRE complexed VDR-RXR heterodimer occurs that is strik¬ ingly dependent upon the presence of the l,25(OH)2D3 ligand (Thompson et al. 1997). Thus, at receptor levels approaching that in a typical target cell, a VDR liganddependent heterodimer with RXR is the preferred VDRE binding species. Only when VDR or VDR plus RXR levels are raised to 100 ng of each receptor with the mouse osteopontin VDRE (Thompson et al. 1997), or 260 ng with the weaker rat osteocalcin VDRE (Nakajima et al. 1994), can faint homodimers of VDR bound to the probe be visualized. In addition, at these greater amounts ofreceptors, neither the VDR homodimer nor the VDRRXR heterocomplexes are modulated significantly by inclusion of l,25(OH)2D3 in the incubation (Thompson et al. 1997). We, therefore, conclude that higher receptor levels in vitro generate artifactual VDR homodimers as well as attenuate the normal physiological ligand dependence of VDR-RXR binding to the VDRE. To explain seemingly ligand-independent VDR-RXR association with the VDRE, we postulate the existence of a subpopulation of VDR that is unstably activated in the absence of l,25(OH)2D3 (Schulman et al. 1996) and therefore capable of heterodimerization to generate a positive gel mobility shift under conditions of vast receptor excess. In contrast, our physiologically relevant gel shift assay at <10nM receptor levels and 0-15 m KCl reflects the presumed in vivo events of ligand triggered heterodimerization (Blanco et al. 1996, Chen et al. 1996), and extends earlier in vitro data showing that l,25(OH)2D3 enhances VDRRXR complex formation (Sone et al. 1991, MacDonald et al. 1993, Ohyama et al. 1994).
Next, we tested the effect of 9-cis RA in this gel shift assay. A spectrum of data exists on the role of 9-cis RA in l,25(OH)2D3-stimulated transcription, including demon¬ stration of synergistic action with l,25(OH)2D3 (Carlberg et al. 1993, Schrader et al. 1994, Kato et al. 1995, Sasaki et al. 1995), negligible action (Ferrara et al. 1994), or an inhibitory effect (MacDonald et al. 1993, Jin & Pike 1994, Lemon & Freedman 1996). These marked differences likely result from varying transfection and ligand addition protocols, as well as cell and species specificity. Employing the physiological gel shift procedure with biochemically defined components, we obtained clear evidence that 9-cis RA is a potent inhibitor of l,25(OH)2D3-enhanced, VDR-RXR binding to VDREs such as osteocalcin, with dramatic attenuation by the retinoid occurring at concentrations as low as 10 m (Thompson et al. 1997). Previous gel shift data had also hinted at 9-cis RA inhibition (MacDonald et al. 1993, Cheskis & Freedman 1994), even though higher concentrations of 9-cis RA were utilized in these earlier studies. One somewhat puzzling finding, however, was that the suppressive effect of 9-cis RA seemed more pronounced in vitro than in transfected cells, where retinoid inhibition of l,25(OH)2D3-stimulated transcription is significant, but 50% or less in magnitude (MacDonald et al. 1993). This suggested that multiple pathways may exist for the assembly of the RXR-VDR heterocomplex in vivo. To probe for distinct routes of assembly, we varied the order of addition ofVDR, RXR, l,25(OH)2D3 and 9-cis RA in the gel shift assay for VDRE binding (Thompson et al. 1997). The results showed that 9-cis RA is a potent inhibitor of VDR-RXR heterodimerization on the VDRE in all situations except when VDR alone is preincubated with l,25(OH)2D3 followed by addition of RXR (Thompson et al. 1997). To explain these data, we have developed the model depicted in Fig. 6, which hypothesizes two alternative allosteric pathways for the interaction ofVDR-RXR with the VDRE.
Figure 6 Model of two different allosteric pathways for VDR-RXR-1,25(OH)2D3 binding to DNA.
In pathway A (Fig. 6), l,25(OH)2D3 occupies monomeric VDR, altering the conformation of the ligand binding domain such that it recruits RXR for heterodimeric binding to DNA and subsequent VDRE recognition. Importantly, we pos¬ tulate that previously occupied VDR conformationally influences RXR in the resulting heterodimer such that it is incapable of being liganded by 9-cis RA (pathway A, Fig. 6). This action to abolish RXR ligand responsiveness both silences the ability of 9-cis RA spuriously to trigger vitamin D hormone signal transduction, and prevents 9-cis RA from dissociating the RXR-VDR complex in order to divert RXR for retinoid signal transduction. On the other hand, as illustrated in pathway (Fig. 6), we propose that RXR exists in a different, 9-cis RA-receptive, allosteric state in most other circumstances, such as when present as a monomer, in an apoheterodimer with VDR, or even when the apoheterodimer of RXR and VDR is subsequently liganded with l,25(OH)2D3. This latter species of RXR-VDR-l,25(OH)2D3 (pathway B) is hypothesized to be fully competent in VDRE recognition, but the 9-cis RA binding function of the RXR partner has not been conformationally repressed, rendering this form sensitive to dissociation by 9-cis RA, which would then favor the formation of retinoid-occupied RXR homo¬ dimers. Therefore, unless VDR monomers are first occu¬ pied by l,25(OH)2D3 (pathway A), 9-cis RA can operate to divert or dissociate RXR and direct it to form RXR homodimers (pathway B). It is tempting to speculate that the l,25(OH),D3-liganded heterodimer in pathway A is more potent in transcriptional stimulation than the analogous species in pathway B, perhaps because the AF-2 function of the RXR partner is allosterically activated only in the former instance. The l,25(OH)2D3-occupied VDR-RXR in pathway has the advantage of flexible regulation because it is effectively a two-ligand switch. It likely occurs in vivo because, as stated above, the fact that 9-cis RA blunting significant but incomplete suggests that at least two populations of RXR-VDR heterodimers exist. Finally, when our model (Fig. 6) is compared with those for RXR-RAR and RXR-TR (Forman et al. 1995), it is evident that VDR is closer in mechanism of action to the TR, where 9-cis RA inhibits TR signal transduction by diversion of BJÍR (Lehmann et al. 1993). Also analogous is the fact that thyroid hormone occupation of the TR partner abolishes 9-cis RA binding to the RXR counter¬ part (Forman et al. 1995). Finally, the action of RXRPJ\R heterodimers seems to be fundamentally different from that of RXR-VDR in that RAR liganding by a retinoid facilitates RXR occupation by its retinoid ligand, resulting in cooperative stimulation of gene transcription by the repertoire of vitamin A metabolites.
VDR protein-protein interactions that effect gene transcription
Although we now have at least a rudimentary understand¬ ing of ligand-induced VDR binding to a VDRE, the next logical question is how does VDR regulate the machinery for gene transcription? In the basal state ofDNA transcrip¬ tion, the TATA-box binding protein (TBP) and its associated factors (TAFs) are bound to the TATA box at approximately position — 20 in the 5′ region of controlled genes, but the frequency of transcriptional initiations is very low because the RNA polymerase II-basal transcription factor IIB (TFIIB) enzyme complex is not stably associated with TBP-TAFs. The recruitment of the TFIIB-RNA polymerase II complex appears to be the rate limiting step in preinitiation complex formation, and is stimulated dramatically when a transacting factor or factors bind to upstream enhancers. In a process involving DNA looping, transactivators are thought to attract TFIIB and also interact with TAFs, forming a stable preinitiation complex that executes repeated rounds of productive transcription. Recent data indicate that the activation function in the hormone binding domain of the estrogen receptor, AF-2, associates specifically with a TAF known as TAFn30 (Jacq et al. 1994) and that the estrogen receptor (ER) binds to TFIIB in vitro (lng et al. 1992). In collaboration with Ozato and associates and Tsai and O’Malley, we have observed that hVDR also specifically associates with hTFIIB (Blanco et al. 1995). In this work, Blanco et al. (1995) showed that VDR binds to a TFIIB-glutathione S transferase fusion protein linked to glutathione-laden beads. Additionally, it was observed that both TRa and RARa interact with hTFIIB (Blanco et al. 1995), but that RXR does so only very weakly (P W Jurutka, L S Remus and M R Haussler, unpublished results). This last result suggests that, while the ligand binding partners in the VDR/TR/RAR subfamily provide a hard-wired connection to the assembly and en¬ hancement of the transcription machinery, the RXR partner is not primarily engaged in TFIIB contact.
Independent data obtained by MacDonald et al. (1995) using the powerful yeast two-hybrid system to detect protein-protein interactions also revealed that hVDR binds efficiently to TFIIB. Moreover, MacDonald et al. (1995) further exploited the yeast two-hybrid system to prove that, while hVDR and RXR interact, no homodimeric association occurs for hVDR alone, providing further evidence against the existence of physiologically significant VDR homodimers. Utilizing fusion protein technology, they also showed that VDR interacts directly with RXR to form a heterodimer in solution in the absence of DNA and, further, that this process was enhanced 8-fold by the presence of l,25(OH)2D3 hor¬ mone (MacDonald et al. 1995). Because hVDR-TFIIB association is not dependent upon the l,25(OH)2D, ligand (Blanco et al. 1995, MacDonald et al. 1995), the role of l,25(OH)2D3 can now be further resolved to an early participation in conforming VDR such that it attracts RXR followed by the targeting of the resulting RXR-VDR heterodimer to VDREs (see Fig. 6).
Figure 6 Model of two different allosteric pathways for VDR-RXR-1,25(OH)2D3 binding to DNA.
Interestingly, the presence of BJCR further facilitates VDR-TFIIB association, especially in the presence of l,25(OH)2D3 (PW Jurutka, LS Remus and MR Haussler, unpublished results). In fact, because of its capacity to enhance VDR-RXR heterodimerization, the l,25(OH)2D3 ligand is capable ofgenerating high levels of an RXR-VDR-TFIIB ternary complex in solution, sig¬ nificantly in excess ofthat occurring with either RXR and TFIIB or even with VDR and TFIIB (P W Jurutka, L S Remus and M R Haussler, unpublished results). These data not only reaffirm the interaction ofVDR with TFIIB, but also they imply that the l,25(OH)2D3-liganded VDR-RXR complex is the most efficient binder of TFIIB. This latter effect may be the result of positive conformational influences of RXR on liganded VDR, since VDR is the primary attachment moiety for TFIIB.
Because VDR-TFIIB interactions have been detected either in vitro or in the yeast system where certain mammalian cell restrictions may be relaxed, it was import¬ ant to confirm the relevance ofVDR-TFIIB association in mammalian cells. Blanco et al. (1995) have reported functional studies which, for the first time, show the interaction ofTFIIB with a member ofthe steroid receptor superfamily in ligand-dependent activation oftranscription in intact cells. In pluripotent PI9 mouse embryonal carcinoma cells, transfection of hVDR or hTFIIB alone produced no better than a 2-fold induction of VDREluciferase reporter expression by l,25(OH)2D3. However, when transfected together, hVDR and hTFIIB mediated a synergistic transcriptional response of approximately 30-fold when l,25(OH)2D3 was added, an effect which was absolutely dependent on the presence of the VDRE in the luciferase construct. It should be noted that the VDR-TFIIB positive cooperation appears to be cellspecific because similar experiments in contact-inhibited NIH/3T3 Swiss mouse embryo cells resulted in squelching of transcription by TFIIB. Therefore, in more differentiated cells, perhaps including osteoblasts or fibro¬ blasts, accessory coactivators may be present to modulate TFIIB or bridge between VDR and TFIIB.
In summary, VDR and TFIIB are hypothesized to exist in a multi-subunit transcription complex which also con¬ tains TAFs and/or coactivators that may be promoter- or tissue-specific. Further characterization of this complex will require the discovery of cell type and promoterspecific components via transfection and biochemical interaction studies. Ultimately, an in vitro transcription system must be devised which utilizes defined components to replicate faithfully l,25(OH)2D3-stimulated gene expression.
One subdomain of VDR that likely interacts with coactivators and/or basal transcription factors is the extreme C-terminus. We have previously shown that 403 hVDR, a truncated receptor that lacks the C-terminal 25 amino acids, binds l,25(OH),D3 ligand with reasonable affinity and heterodimerizes normally with RXR, but is devoid of transcriptional activity (Nakajima et al. 1994). These data suggest that VDR contains a transcriptional activation domain near its C-terminus.
Figure 7 The extreme C-terminal amino acid sequence compared across the nuclear receptor superfamily: VDR appears to share the ligand-dependent transcription activation function (AF-2). AR, androgen receptor; CR, glucocorticoid receptor; PR, progesterone receptor.
Indeed, as illustrated in Fig. 7, the region of VDR from residues 416 to 422 possesses a high degree of similarity to the analogous sequences in the entire nuclear receptor superfamily. One hallmark of this conserved sequence is the glutamic acid residue at position 420 of hVDR (Fig. 7) included in a consensus of (where cp=a hydrophobic amino acid) for this domain (Renaud et al. 1995, Wagner et al. 1995). Allowing for conservative replace¬ ments, it seems virtually certain that hVDR forms an amphipathic helix (corresponding to helix 12 in the other receptors) surrounding glutamic acid-420 that is analogous to the ligand-dependent activation function (AF-2) char¬ acterized for TR (Barettino et al. 199A), RAR (Renaud et al. 1995), RXR (Leng et al. 1995) and ER (Danielian et al. 1992). Although this AF-2 domain is capable of autonomously activating transcription (Leng et al. 1995), that such activity is modest may be because of the fact that the AF-2 region is proposed to operate in a liganddependent fashion, involving a structural rearrangement to reposition the AF-2 for both intramolecular and intermolecular protein—protein interactions. Specifically, based upon the crystal structure of unoccupied RXR (Bourguet et al. 1995) and liganded RAR (Renaud et al. 1995) and TR (Wagner et al. 1995), helix 12/AF-2 appears to protrude outward from the more globular ensemble of helices 1—11 in the absence ofligand, such that it is unable to interact efficiently with coactivator/transcription factor. Upon liganding, a conformational signal is then transmit¬ ted to helix 12 that causes it to fold back on helix 11 and attach to the face of the globular ligand binding domain. The pivoting of helix 12 seemingly accomplishes two feats that mediate ligand-activated transcription by the receptor: (i) closing of a ‘door’ on the channel through which the lipophilic ligand enters the internal binding pocket of the receptor, and (ii) locking helix 12 into a stable confor¬ mation that facilitates its interaction with coactivator/ transcription factor. Ligand binding contacts on or near helix 12 (see Fig. 3) probably are significant in maintaining this active positioning of helix 12, essentially trapping ligand in the binding pocket to effect more sustained transactivation events.
Figure 7 The extreme C-terminal amino acid sequence compared across the nuclear receptor superfamily: VDR appears to share the ligand-dependent transcription activation function (AF-2). AR, androgen receptor; CR, glucocorticoid receptor; PR, progesterone receptor.
In order to evaluate the relevance of the above proposed mechanism for VDR action, we (Jurutka et al. 1997) have altered E-420 and L-417 (see Fig. 7) individually to alanine residues, which preserves the putative -helical character of this region. The altered VDRs bind ligand near-normally, with only a mild increase (about 3-fold) in the Kd for the E420A receptor. Both E420A and L417A hVDRs also heterodimerize efficiently with RXR and associate with VDREs similarly to wild-type hVDR, yet their capacity for l,25(OH)2D3-stimulated transcription is abolished, even at high doses ofligand (Jurutka et al. 1997). These point mutations, therefore, identify a C-terminal AF-2 in VDR which corresponds to similar activation domains in other nuclear receptor superfamily members. Because VDR interacts with TFIIB, one of the first questions we asked was whether the VDR AF-2 consti¬ tutes a contact site for this basal transcription factor. Although some very preliminary evidence existed for an association between TFIIB and the C-terminus of hVDR (MacDonald et al. 1995), we observed that neither the E420A nor the L417A mutant VDRs are impaired in their interaction with TFIIB as probed with glutathione-S transferase—TFIIB fusion protein binding technology (Jurutka et al. 1997). Thus, the domain(s) of VDR that interfaces with TFIIB apparently lies elsewhere in the receptor, possibly in the N-terminal portion of the ligand-binding region (Blanco et al. 1995), in the hinge (MacDonald et al. 1995), or in the vicinity of the DNA-binding zinc fingers.
The present experiments with VDR are in concert with recent insight into the function of AF-2 in other nuclear receptors, which is to recruit coactivators of the type of steroid receptor coactivator-1 (SRC-1) (Oñate et al. 1995). A number of candidate coactivators have been isolated in addition to SRC-1 (Halachmi et al. 199A, Baniahmad et al. 1995, CavaiUes et at. 1995, Lee et al. 1995, Hong et al. 1996) and, in several cases, interaction with nuclear receptors requires intact AF-2 core regions (Baniahmad et al. 1995, CavaiUes et al. 1995). Moreover, AF-2 mutations act as dominant negative receptors, for example in the case of hRARy (Renaud et al. 1995). Indeed, we have observed that VDR AF-2 mutants E420A and L417A exhibit dominant negative properties with respect to transcriptional activation (Jurutka et al. 1997). Such AF-2 altered receptors are inactive transcriptionally, but can bind l,25(OH)2D3 ligand and heterodimerize normally on VDREs, the consequence being competition with wild-type VDR-RXR heterodimers for VDRE binding. These data argue that the AF-2 of the primary VDR partner in an RXR-VDR heterodimer is absolutely required for the mediation of l,25(OH)2D3-activated transcription, not only for its intrinsic activation potential, but also because of its presumed role in stabilizing the retention of l,25(OH)2D3 ligand in the VDR binding pocket.
Figure 6 Model of two different allosteric pathways for VDR-RXR-1,25(OH)2D3 binding to DNA.
What part, if any, is played by the AF-2 domain (Fig. 7) of the RXR ‘silent’ partner in the RXR-VDRl,25(OH)2D3 signal transduction pathway? To investigate this phenomenon, AF-2 truncated mutants of RXRa or RXRß were created and tested for their ability to function as dominant negative modulators of l,25(OH)2D3- stimulated transcription (Blanco et al. 1996). Because previous data with RXR-RAR control of gene expression seemed to indicate that the RXR AF-2 was dispensable (Durand et al. 1994), we were surprised to find that AF-2 truncated RXRs were potent dominant negative effectors of l,25(OH)2D3 action in transfected cells (Blanco et al. 1996). We, therefore, conclude that although the RXR ‘silent’ partner in VDR signaling apparently is not occupied by retinoid ligand (see Fig. 6), its AF-2 does play an active role in transcriptional stimulation. A similar conclusion has also been reached recently by two other groups studying RXR-RAR action (Chen et al. 1996, Schulman et al. 1996), with the use of RAR-specific ligands precluding ligand binding by the RXR partner. However, Schulman et al. (1996) have introduced a caveat to the above theory as they point out that AF-2-truncated RXRs in heterodimers become strong, constitutive binders of corepressors like the silencing mediators of retinoid and thyroid hormone receptors (SMRTs). Thus, an alternative explanation to an active coactivator-binding role for RXR AF-2 in heterodimers is that it plays a more passive role in excluding corepressors. In this latter scenario, truncation or point mutation (Schulman et al. 1996) of RXR AF-2 generates spurious corepressor binding rather than compromising coactivator contact. Only additional research into coactivator and corepressor associations of VDR-RXR heterodimers will resolve this issue.
General mechanism for vitamin D hormone action on transcription
In order to provide a working hypothesis for l,25(OH)2D3 action at the molecular level, we have developed the model illustrated in Fig. 8. It is based primarily on data from our laboratory and others studying 1,25(OH)2D3 and VDR, and it relies on the assumed similarities between VDR action and that of TR and RAR. VDR is proposed to exist in target cell nuclei, perhaps very weakly associ¬ ated with DNA, in a monomeric, inactive conformation with the C-terminal AF-2 domain extended away from the hormone binding cavity. Upon liganding with l,25(OH)2D3, VDR assumes an active conformation, with the AF-2 pivoted into correct position for both ligand retention and coactivator contact. In addition, the hormone facilitates interaction of VDR and RXR through a stabilized heterodimerization interface. In turn, 1,25(OH)2D3-occupied VDR may itselffunction as a kind of allosteric regulator of RXR, perhaps by conveying a confonnational signal through the juxtapositioned dimer¬ ization domains to induce the AF-2 ofRXR into an active conformation for coactivator binding. As discussed above (see Fig. 6), the joining of preliganded VDR and unliganded RXR apparently renders the RXR partner unresponsive to binding and either activation or dissocia¬ tion by 9-cis RA. Alternatively, if 9-cis RA encounters RXR monomer first (Fig. 8), or binds to RXR that is complexed with VDR in an apoheterodimer (Fig. 6), the retinoid is able to divert the RXR to generate homo¬ dimers and effectively blunt l,25(OH)2D3-driven transcription In the primary activation pathway pictured in Fig. 8, the RXR-VDR-l,25(OH)2D3 complex recognizes and targets the genes to be controlled through high affinity association with the VDRE in a gene promoter region. Coactivators that are presumed to bind to VDR and RXR AF-2 s are then postulated to link with TAFs/TBP, thereby looping out DNA 5′ of the TATA box. This series of events positions VDR such that it can independently recruit TFIIB to the promoter complex, a process that initiates the assembly of the RNA polymerase II holoenzyme into the preinitiation complex. Precedents exist for transcription factors independently attracting TFIIB, such as hepatocyte nuclear factor-4 (Malik & Karathanasis 1996), as well as for a sequential, two-step pathway for activator-stimulated transcriptional initiation (Struhl 1996, Stargell & Struhl 1996). Using the latter model as an analogy, the VDR activator would contact both TBP/ TAFs (via – coactivator bridges) and TFIIB in order to initiate RNA polymerase II holoenzyme assembly. The order of attachment of these two ‘arms’ of activation has not been determined but, at least, in the case of acidic activators, recruitment to the TATA element precedes interaction with components of the initiation complex (Stargell & Struhl 1996). It is of interest that the mechan¬ ism of l,25(OH)2D3 action depicted in Fig. 8 is not only essential for induction of bone remodeling and other vitamin D functions, but is also self-limiting via 24-OHase induction. In addition, these actions of l,25(OH),D, would be blunted under conditions within a cell where 9-cis RA concentrations dominate over those of l,25(OH)2D3.
Figure 8 Model for transcriptional activation by 1,25(OH)2D3 on the promoter of a target gene
The above described molecular mechanism whereby the vitamin D hormone controls gene expression requires further experimental evaluation. To advance our under¬ standing of the structure/function relationships in VDR, a physical characterization of the structure of VDR via X-ray crystallography will be required. Furthermore, in order to comprehend the genomic action of vitamin D in calcium homeostatic and other target cells, it will be necessary to elucidate the detailed involvement of various RXR isoforms, specific TAFs and novel coactivators/ corepressors that might influence the regulation of differ¬ ent vitamin D-controlled promoters. This information in its entirety should assist in determining the potential role for VDR and l,25(OH)2D3 in the pathophysiology of osteoporosis and other endocrine-related bone diseases.
References
Baniahmad C, Nawaz Z, Baniahmad A, Gleeson MAG, Tsai M-J & O’Malley BW 1995 Enhancement of human estrogen receptor activity by SPT6: a potential coactivator. Molecular Endocrinology 9 34-43.
Barettino D, Ruiz MdMV & Stunnenberg HG 1994 Characterization of the ligand-dependent transactivation domain of thyroid hormone receptor. EMBOJournal 13 3039-3049.
Blanco JCG, Wang I-M, Tsai SY, Tsai MJ, O’Malley BW, Jurutka PW, Haussler MR & Ozato 1995 Transcription factor TFIIB and the vitamin D receptor cooperatively activate ligand-dependent transcription. Proceedings of the National Academy of Sciences of the USA 92 1535-1539.
Blanco JCG, Dey A, Leid M, Minucci S, Park B-K, Jurutka PW, Haussler MR & Ozato 1996 Inhibition of ligand induced promoter occupancy in vivo by a dominant negative RXR. Genes to Cells 1 209-221.
Bourguet W, Ruff M, Chambón , Gronemeyer & Moras D 1995 Crystal structure of the ligand-binding domain of the human nuclear receptor RXR-a. Nature 375 377-382.
Cai Q, Hodgson SF, Kao PC, Lennon VA, Klee GG, Zinmeister AR & Kumar R 1994 Inhibition of renal phosphate transport by a tumor product in a patient with oncogeneic osteomalacia. New England Journal of Medicine 330 1645-1649.
Cao X, Ross FP, Zhang L, MacDonald PN, Chappel J & Teitelbaum SL 1993 Cloning of the promoter for the avian integrin ß3 subunit gene and its regulation by 1,25-dihydroxyvitamin D3. Journal of Biological Chemistry 268 27371-27380.
Carlberg C, Bendik I, Wyss A, Meier E, Sturzenbecker LJ, Grippo JF & Hunziker W 1993 Two nuclear signalling pathways for vitamin D. Nature 361 657-660.
more…
The structure of the nuclear hormone receptors.
The functions of the group of proteins known as nuclear receptors will be understood fully only when their working three-dimensional structures are known. These ligand-activated transcription factors belong to the steroid-thyroid-retinoid receptor superfamily, which include the receptors for steroids, thyroid hormone, vitamins A- and D-derived hormones, and certain fatty acids. The majority of family members are homologous proteins for which no ligand has been identified (the orphan receptors). Molecular cloning and structure/function analyses have revealed that the members of the superfamily have a common functional domain structure. This includes a variable N-terminal domain, often important for transactivation of transcription; a well conserved DNA-binding domain, crucial for recognition of specific DNA sequences and protein:protein interactions; and at the C-terminal end, a ligand-binding domain, important for hormone binding, protein: protein interactions, and additional transactivation activity. Although the structure of some independently expressed single domains of a few of these receptors have been solved, no holoreceptor structure or structure of any two domains together is yet available. Thus, the three-dimensional structure of the DNA-binding domains of the glucocorticoid, estrogen, retinoic acid-beta, and retinoid X receptors, and of the ligand-binding domains of the thyroid, retinoic acid-gamma, retinoid X, estrogen, progesterone, and peroxisome proliferator activated-gamma receptors have been solved. The secondary structure of the glucocorticoid receptor N-terminal domain, in particular the taul transcription activation region, has also been studied. The structural studies available not only provide a beginning stereochemical knowledge of these receptors, but also a basis for understanding some of the topological details of the interaction of the receptor complexes with coactivators, corepressors, and other components of the transcriptional machinery. In this review, we summarize and discuss the current information on structures of the steroid-thyroid-retinoid receptors.
Cellular retinoid-binding proteins.
A number of specific carrier proteins for members of the vitamin A family have been discovered. Two of these proteins bind all-trans-retinol and are found within cells important in vitamin A metabolism or function. These two proteins have considerable sequence homology and have been named cellular retinol-binding protein (CRBP) and cellular retinol-binding protein, type II (CRBP [II]). A third intracellular protein, cellular retinoic acid-binding protein (CRABP) also is structurally similar but binds only retinoic acid. Although retinol appears to be bound quite similarly by the two retinol-binding proteins, subtle differences are apparent that appear to be related to the different functions of the two proteins. That, coupled with the specific cellular locations of the two proteins, suggests their roles. Cellular retinol-binding protein appears to have several roles, including (1) delivering retinol to specific binding sites within the nucleus and (2) participating in the transepithelial movement of retinol across certain blood-organ barriers. In contrast, CRBP (II) appears to be involved in the intestinal absorption of vitamin A and, in particular, may direct retinol to a specific esterifying enzyme, resulting in the production of fatty acyl esters of retinol that are incorporated into chylomicrons for release to the lymph. Like CRBP, CRABP can deliver its ligand retinoic acid to specific binding sites within the nucleus, sites different from those for retinol. The nuclear binding of retinol and retinoic acid may be part of the mechanism by which vitamin A directs the state of differentiation of epithelial tissue.
Interaction of the Retinol/Cellular Retinol-binding Protein Complex with Isolated Nuclei and Nuclear Components
GENE LIAU, DAVID E. ONG, and FRANK CHYTIL
Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, Tennessee 37232
http://jcb.rupress.org/content/91/1/63.full.pdf
Retinol (vitamin A alcohol) is involved in the proper differentiation of epithelia. The mechanism of this involvement is unknown. We have previously reported that purified cellular retinol-binding protein (CRBP) will mediate specific binding of retinol to nuclei isolated from rat liver. We now report that pure CRBP delivers retinol to the specific nuclear binding sites without itself remaining bound. Triton X-100-treated nuclei retain the majority of these binding sites. CRBP is also capable of delivering retinol specifically to isolated chromatin with no apparent loss of binding sites, as compared to whole nuclei . CRBP again does not remain bound after transferring retinol to the chromatin binding sites. When isolated nuclei are incubated with [ 3H]retinol-CRBP, sectioned, and autoradiographed, specifically bound retinol is found distributed throughout the nuclei . Thus, CRBP delivers retinol to the interior of the nucleus, to specific binding sites which are primarily, if not solely, on the chromatin . The binding of retinol to these sites may affect gene expression.
Early histological studies have clearly shown that when animals become vitamin A deficient various epithelial tissues of these animals lose the ability to maintain proper differentiation (1) . However, providing retinol (vitamin A alcohol) to the animal permits tissue repair, with improperly differentiated cells rapidly replaced by normal cells (2) . This indicates that vitamin A has an essential role in cellular differentiation . The action of retinol appears to be mediated by a specific intracellular protein called cellular retinol-binding protein (CRBP). CRBP binds retinol with great avidity and specificity and has been detected in a number oftissues (3, 4) . Recently, CRBP was purified and partially characterized (5, 6) . It is distinct from the well-known serum retinol transport protein called retinol-binding protein (5, 7) . That CRBP plays an important role in the action of vitamin A is suggested by the following observations: It is found complexed with retinol in vivo (4, 8). It binds cis-isomers of retinol with a specificity that parallels the in vivo activity of these isomers (9), Finally, if retinol is first complexed with CRBP, the retinol can bind to the nucleus in a specific and saturable manner (10) . In this study we compare the interaction of the CRBP-retinol complex with isolated nuclei to its interaction with isolated chromatin and follow the fate of both the protein and the ligand . The nuclear binding sites for retinol were localized using autoradiography .
……
The experiments described here were designed to gain insight concerning the still unknown molecular mechanisms by which retinol exerts its effects on the differentiation of epithelia. Alterations in genomic expression appear to be induced in animals fed a retinol-deficient diet, as shown by changes in nuclear RNA synthesis observed in vivo (27-30) as well as in vitro (13) . A working hypothesis has been used that retinol, being a small molecule, might exert its action in a way similar to the accepted model for the mode of action of steroid hormones in differentiation . This model involves binding of the steroid hormone inside the target cell to a specific binding protein called a receptor . The resulting cytoplasmic ligand receptor complex, after undergoing a not fully understood conformational change, translocates to the nucleus . The receptor protein can then be detected in nuclear extracts by its ability to bind specifically the steroid hormone. The receptor steroid complex has been shown to interact with chromatin. Such interaction is believed to lead to an altered expression of the genome, which is the basis for the steroid hormone-induced differentiation (31) .
The steroid hormone model has been used profitably to investigate the mode of retinol action . Indeed, a specific binding protein for retinol, CRBP, was discovered to be present in many tissues (3) . Moreover, after purifying this protein to homogeneity, it was demonstrated that CRBP is able to deliver retinol to the nucleus in a specific manner (10) .
However, we report here a unique feature which appears to be distinct from the steroid hormone model. Using retinol CRBP complex in which the radioactive label is on the protein, we find that CRBP delivers retinol in a specific manner to the nucleus; the retinol associates with chromatin, but the protein itself does not remain bound. This conclusion is based on the observation that the radioactively-labeled protein is still able to deliver retinol inside the nucleus, but it cannot be recovered with the nucleus, in contrast to steroid hormone receptors.
The interaction of the specifically delivered retinol appears to be primarily with chromatin. The outer nuclear envelope is apparently not significantly involved in the interaction as Triton X-100-treated nuclei retain 70% of the retinol binding sites found in intact nuclei. It is still possible that the isolated chromatin and the Triton-treated nuclei contain some of the nuclear matrix and that it is actually the matrix which contains the specific binding sites for retinol. However, preliminary evidence indicates that the specific binding sites remain with a soluble chromatin preparation prepared by mild nuclease digest of nuclei rather than with the nuclear matrix. That the CRBP is necessary for delivering retinol to the nucleus is clearly documented by autoradiography. Free retinol, not bound to CRBP, binds nonspecifically to the nuclei, and to chromatin, and autoradiography shows indiscriminate localization of retinol in the lipid-rich nuclear membrane areas.
The data presented here invite the proposal that the retinolCRBP complex enters the nucleus in some manner which is apparently not dependent on the nuclear membrane. The complex then recognizes a limited number (generally an order of magnitude greater than for steroid hormones) of specific sites on the chromatin where the transfer of retinol from CRBP to these sites takes place. The sites were not detectable and may not be accessible if the retinol is free from CRBP. After the transfer CRBP does not remain associated with the specific sites . The functional significance of the specific interaction between retinol and chromatin remains to be demonstrated .
Inhibition of vitamin D receptor-retinoid X receptor-vitamin D response element complex formation by nuclear extracts of vitamin D-resistant New World primate cells.
Most New World primate (NWP) genera evolved to require high circulating levels of steroid hormones and vitamin D. We hypothesized that an intracellular vitamin D binding protein (IDBP), present in both nuclear and cytoplasmic fractions of NWP cells, or another protein(s) may cause or contribute to the steroid hormone-resistant state in NWP by disruption of the receptor dimerization process and/or by interference of receptor complex binding to the consensus response elements present in the enhancer regions of steroid-responsive genes. We employed electromobility shift assay (EMSA) to screen for the presence of proteins capable of binding to the vitamin D response element (VDRE). Nuclear and post-nuclear extracts were prepared from two B-lymphoblastoid cell lines known to be representative of the vitamin D-resistant and wild type phenotypes, respectively. The extracts were compared for their ability to retard the migration of radiolabeled double stranded oligomers representative of the VDREs of the human osteocalcin and the mouse osteopontin gene promoters. A specific, retarded band containing VDR-RXR was identified when wild type cell but not when vitamin D-resistant cell nuclear extract was used in the binding reaction with either probe. In addition, vitamin D-resistant cell nuclear extract contained a protein(s) which was bound specifically to the VDRE and was capable of completely inhibiting VDR-RXR-VDRE complex formation; these effects were not demonstrated with nuclear extract from the wild type cell line or with the post-nuclear extract of the vitamin D-resistant cell line. We conclude that a VDRE-binding protein(s), distinct from IDBP and present in nuclear extract of cells from a prototypical vitamin D-resistant NWP, is capable of inhibiting normal VDR-RXR heterodimer binding to the VDRE.
Reversing Bacteria-Induced Vitamin D Receptor Dysfunction to Treat Chronic Disease: Why Vitamin D Supplementation Can Be Immunosuppressive, Potentially Leading to Pathogen Increase
by J.C. Waterhouse, PhD
Recent attempts to increase vitamin D supplementation to prevent and treat chronic disease have arisen primarily out of observations of low vitamin D levels (25-D) being associated with a variety of diseases. However, new research indicates that these low vitamin D levels are often the result rather than the cause of the disease process, just as in the autoimmune disease, sarcoidosis. Trevor Marshall, PhD, recently summarized this alternative perspective on vitamin D, in a session he co-chaired at the 6th International Congress on Autoimmunity. He and his colleagues presented in silico* and clinical data from the last eight years, indicating that intraphagocytic bacteria are able to block the vitamin D receptor (VDR), and this leads to abnormally low measured vitamin D levels. A second consequence of the bacteria-induced VDR blockage is inhibition of innate immunity. By blocking the VDR, bacteria are able to cause persistent infection and inflammation and thus cause many chronic diseases. Short-term symptom reduction observed from vitamin D supplementation appears to be due to immune suppression by precursor forms of vitamin D that add to the bacterial blockage of the VDR. In silico data also indicates that high levels of vitamin D metabolites suppress antimicrobial peptide production by binding to other nuclear receptors (e.g., thyroid-alpha-1, glucocorticoid). Increasingly, epidemiological, geographical and clinical data are lending support to this model of disease. Studies using more advanced cell culture and molecular techniques are confirming the presence of previously undetected bacteria, including biofilm and cell wall deficient bacteria, as well as “persisters.” A greater understanding of how bacteria resist standard antibiotic approaches is also being gained. A protocol has been developed that is successfully restoring VDR and innate immune function with a VDR agonist and eliminating pathogens with low-dose, pulsed combinations of antibiotics. Immunopathological reactions (a.k.a., Jarisch-Herxheimer reactions) occur due to increased pro-inflammatory cytokines resulting from bacterial killing. The result is an exacerbation of symptoms with each dose of antibiotic, but improvement occurs over the long-term. Remission is being achieved in numerous chronic conditions, including many autoimmune diseases and fibromyalgia, as well as many diseases of aging. Although vitamin D ingestion is avoided as part of this protocol, the evidence indicates that the net result of the protocol is improved vitamin D receptor activation.
Introduction
Vitamin D is a topic of increasing interest and has been implicated in many physiological processes beyond its initially recognized role in calcium absorption and metabolism.1 Vitamin D is found in supplements and a few foods (e.g., fish, liver, egg yolk, fortified products). The majority of vitamin D is produced in the skin when exposed to UV radiation from sunlight. But some have begun advocating consumption of levels of vitamin D above the RDA, and some advocate very high levels, ranging from 1,000 to 5,000 IU or more daily.2 Vitamin D is a secosteroid, with a close resemblance in structure to immunosuppressive steroids. Levels of the various vitamin D metabolites are the result of complex feedback mechanisms involving multiple enzymes and receptors, indicating that it is regulated more like a steroid than a nutrient.1
Short-term symptom reduction has sometimes been observed through increases in sun exposure 3,4 or vitamin D supplementation.5 However, this appears to be due to the anti-inflammatory effect arising from immune suppression, analogous to the effect of a steroid, such as prednisone. If one were to assume that the inflammation is purely pathological, this might be considered beneficial, but evidence that has been accumulating over many decades indicates that inflammation in most chronic diseases is occurring in response to undetected chronic bacterial infection (see below). Since immune suppression can promote the increase of pathogens, the effect of vitamin D supplementation is not likely to be harmless in this situation, but appears to have long-term effects associated with increased levels of bacterial pathogens. The role of this microbiota in producing the inflammation and oxidative stress observed in so many diseases will be discussed near the end of this article.6-8
Vitamin D from food or sun is first converted to 25-D (25-hydroxyvitamin-D) and then converted in a second step to the active 1,25D form (1,25-dihydroxyvitamin-D) that is able to activate the vitamin D receptor (VDR). The type of vitamin D usually measured in the blood is the precursor form, 25-D, rather than 1,25-D, the form that activates the receptor. Activation of the vitamin D receptor is extremely important, as it has numerous effects, including effects on the immune system1 and cancer.9,10 However, recent research indicates that increasing vitamin D via supplementation or sun exposure is not the way to achieve more VDR activation in chronic disease, due to blockage of the VDR by bacterial products.6 This insight has been put to use in a new model of chronic disease and a new protocol.6,8,11-14
A New Perspective on Vitamin D and a New Treatment Approach
Trevor Marshall, PhD, (Murdoch University, Australia) has developed a model of chronic autoimmune and inflammatory diseases in which intraphagocytic bacteria cause disease by producing a substance that binds to and blocks the VDR.1 One such substance has been already identified providing proof of principle.1
- The VDR is important for adequate innate immune function, including the production of numerous antimicrobial peptides.15
These include
- cathelicidin and
- beta-defensin,
two of the body’s own arsenal of internally produced antibiotics.
Thus, VDR blockage would seem to be an excellent bacterial strategy, as it would lead to poor innate immune system function and further growth of bacteria and other pathogens. A functioning VDR also appears to be important in controlling cell growth and metastasis, so as to help prevent and control cancerous growths.9,10
A protocol based on this model of disease has been achieving a high rate of improvement/remissions in a wide array of conditions.6,11-14,16-18 It involves the use of
- a VDR agonist, olmesartan, which is able to activate the VDR effectively and safely.
In addition, low dosages of combinations of select pulsed antibiotics are used to eliminate the bacteria, which also helps restore VDR functioning. The protocol also involves avoidance of vitamin D supplementation. When faced with VDR dysfunction, the evidence indicates that
- attempting to increase 25-D only adds to the dysregulation of the vitamin D metabolites without being able to adequately overcome the bacteria-induced VDR blockage.6,8
Too much vitamin D can be harmful in two ways, according to Marshall’s work.1,6
- In silico data from highly sophisticated molecular modeling shows that high vitamin D levels can block the VDR and thus block innate immune function.18 In addition,
- high levels of various vitamin D metabolites can affect thyroid-alpha-1, glucorticoid, and androgen receptors and disrupt hormonal control and further affect innate immune function.1
Thus, any short-term symptom reduction from high levels of vitamin D that may occur is probably occurring at the cost of long-term pathogen increase. This has been supported by observations of patient’s responses over time. In the short-term, even for ten years or more in some cases, the person may feel better with high vitamin D intake. But in the long-term, the chronic infection progresses, because the high 25-D is only adding to the bacterial blockage of the VDR and the suppression of bacterial killing.18
Symptoms increase when the immune system is better able to kill the pathogens, due to the high levels of inflammatory cytokine levels that occur. This is called the immunopathological reaction or Jarisch-Herxheimer reaction.6,11 The symptoms range from pain and fatigue to cognitive impairment and depression, but include numerous other symptoms characteristic of the underlying inflammatory condition.6,11 By suppressing the immune response, vitamin D supplementation may suppress these symptoms in the short-term and may even result in a sort of dependence on vitamin D supplementation or sun exposure to keep the symptoms at bay.
The long-term efficacy of the protocol (sometimes called the Marshall Protocol or MP) in activating the VDR is also supported by improved or stabilized bone density, which is typical in patients on the protocol, if the RDA of calcium is consumed. The protocol replaces vitamin D supplementation with use of the VDR agonist olmesartan (120 to 160 mg in divided doses) and reduces the level of bacteria blocking the VDR with antibiotics and, in this way, is apparently effective in activating the VDR.6,12
Marshall proposes that vitamin D receptor blockage results in the low levels of 25-D that have been observed in numerous diseases. The precursor, 25-D form is the form that is most frequently measured. The VDR blockage typically leads to dysregulation of metabolite levels, and one effect is down-regulation of the conversion of vitamin D to 25-D.1 Thus, according to this perspective, low 25-D levels are the result, not a cause, of the disease process. It follows that a low serum 25-D is not indicative of a true vitamin D deficiency in this situation. Both laboratory19 and clinical findings20 have supported the existence of an apparently similar type of down-regulation of conversion to 25-D.
At the same time that low 25-D is observed, high 1,25-D levels are also usually observed. In fact, elevated 1,25-D has been shown to be a good indicator of inflammatory and autoimmune disease.13,16 When interpreting the results, however, it should be remembered that samples must be frozen until analyzed for accurate 1,25-D results. And occasionally, in cases of quite advanced disease or elderly patients, 1,25-D will be low as well, yet still be consistent with VDR blockage and inflammatory disease.21
Marshall’s protocol was first used to treat sarcoidosis. It is well established that a dysregulation of vitamin D levels, often with very high 1,25-D and low 25-D, occurs in this condition.22 Marshall’s and other’s work has confirmed that this dysregulation also occurs in a wide range of other diseases.12,13,23,24 This pattern of high 1,25-D and low 25-D also exists in VDR knockout mice.25 These mice are genetically engineered to lack a VDR, a situation analogous to a bacteria-blocked VDR.
The very complex relationships among genes, metabolites, enzymes, and receptors that Marshall recently summarized1,6 show that vitamin D is not a mere nutrient. In fact, the active form is a secosteroid transcriptional factor. It is part of a highly regulated and complex system influencing many aspects of metabolism and immune function. There are several feedback and feedforward pathways that influence the levels of various vitamin D forms that Marshall reviewed in depth.1
Marshall was recently invited to co-chair a session on vitamin D at the 6th Annual International Autoimmunity Conference, and he gave one of the keynote presentations of the session.6 Several other presentations were given that support the protocol and model. For example, Perez presented data on treatment response in 20 autoimmune conditions that support Marshall’s model.11 The autoimmune diseases successfully treated in this open-label trial include rheumatoid arthritis, systemic lupus erythematosis, diabetes type 1 and 2, psoriasis, Hashimoto’s thyroiditis, Sjogren’s syndrome, scleroderma, uveitis, myasthenia gravis, and ankylosing spondylitis. Chronic fatigue syndrome and fibromyalgia were shown to respond to the protocol in another presentation.17 And another study indicated that dysregulation of nuclear receptors in the endometrium by vitamin D, along with chronic bacterial infection, can help explain the higher prevalence of some autoimmune diseases in women.26
Epidemiological and Short-Term Clinical and Experimental Data
The in silico and clinical data discussed above provide strong evidence for Marshall’s model, and some might argue it is more reliable than epidemiological and short-term evidence. It is widely recognized that there are many limitations inherent in epidemiological and short-term experimental data due to difficulties in obtaining relevant and accurate results. Confounding factors and the inability to assess the effects of long-term immune suppression from high levels of vitamin D make the results less reliable.13,21 Experiments using animal models have the problem of genetic differences and different disease causation methods.1,13Studies of supplementation are often not randomized and thus are subject to unknown confounding factors that may affect the choice to take vitamin D supplements.13 Furthermore, sun exposure is hard to quantify and is often left out of the analyses. Any of the above can lead to invalid conclusions.
Despite this, a number of recent studies that may be relevant will be discussed here to show that there is much independent support for Marshall’s model among these types of studies. In addition, some lesser-known aspects of some of the studies used to support a high vitamin D intake will be reviewed, which cast doubt on some of their conclusions.
Cancer and All-Cause Mortality
In the case of cancer prevention, a recent randomized controlled trial of calcium and vitamin D by Lappe et al.27 is used to support vitamin D supplementation. However, it has a number of serious limitations. One problem is the assumption that removing the data from the first year is justified. If one looks at Figure 1, in the article by Lappe et al,27 in which the data from the first year was included, there is very little difference between calcium and vitamin D vs. calcium alone throughout the study period. No group of patients was given vitamin D alone. Also, there is not yet long-term data on incidence, since the study lasted only four years. Any reduced incidence may reflect delay in diagnosis. In addition, long-term survival may not ultimately improve. In fact, patients taking vitamin D might even die sooner (see below). In addition to the above critique, a number of published comments have also taken issue with this trial, pointing to other problems and limitations.9,28
Another recent study29 reported finding barely significant lower cancer rates in premenopausal women (95% confidence interval, 0.42-1.0) who consumed more vitamin D. However, they found a marginally significant higher rate of moderately differentiated tumors in postmenopausal women who had higher vitamin D intake. And since postmenopausal women make up a much higher proportion of breast cancer cases, this is particularly concerning. This is just one example of the rather inconclusive, mixed data on vitamin D supplementation that becomes apparent when the vitamin D studies are looked at as a whole (see Discussion section in ref. 29). Even the benefit for premenopausal women is questionable. Bertone-Johnson et al.30 pointed out a quite plausible rationale for the existence of a bias toward low estrogen in those who choose to take vitamin D supplements.
A number of limitations found in the other studies are used as a basis for supporting vitamin D supplementation. For instance, the data is rarely long-term enough and rarely covers all the effects possible. Although there may be an appearance of benefit in the short-term or for subsets of the populations studied, a large, long-term prospective study showed no effect of 25-D on the overall cancer mortality rate in the long-term.31 Freedman et al.31 even showed a suggestion of a negative effect of higher vitamin D levels. There was a non-significant increase in overall mortality in the two groups with 25-D at higher levels (80 to <100 nmol/L: Risk Ratio = 1.21, 95% CI =0.83 to 1.78; =100 nmol/L: Risk Ratio = 1.35; 95% CI = 0.78 to 2.31, where 100 nmol/L corresponds to about 40 ng/ml).
This is in accord with a study in prostate cancer32 (also see discussion in ref. 21) and one in pancreatic cancer33 that found higher cancer rates when 25-D was high. Cancer rates increased among patients with a 25-D level above approximately 32 ng/ml. Evidence regarding solar radiation and geographical/latitudinal analyses are also used as evidence, yet solar radiation has many other effects besides raising 25-D.34,35 Many other relevant factors, such as pathogen distributions, climate effects on pathogen spread36,37 and host susceptibility,38 diet, and pollution levels also vary with geographical location.
It was recently pointed out in the Bulletin of the World Health Organization that high 25-D has been found to be associated with greater cancer risk in some studies.39 Studies mentioned, included one that found that there was a higher rate of many internal cancers in patients who have a type of skin cancer that is considered to be the best indicator of long-term sun exposure.40 Another study discussed failed to find a geographical pattern that would support a protective effect of increased 25-D.41 On the whole, in these epidemiological studies, the data is mixed and inconsistent, which is to be expected when there are so many unknown confounding factors affecting 25-D levels and disease incidence that may bias the results.13 In addition, a recent large prospective study presented evidence suggesting that circulating 25-D concentrations may be associated with increased risk of aggressive prostate cancer.42 For all types of prostate cancer, the data failed to support the hypothesis that higher vitamin D decreases prostate cancer risk.42
Studies looking at overall mortality benefits of vitamin D are sometimes misleading at first glance. In the large meta-analysis done recently on the effect of vitamin D and calcium on mortality rates,43 the abstract attributes reduced mortality to vitamin D, yet the only statistically significant results were for calcium together with vitamin D. Another serious problem is that most of the studies analyzed in the meta-analysis were only a few years in duration, so long-term effects on mortality and morbidity could not be accurately assessed.
Bone Density, Parathyroid Hormone
Another area that should be re-evaluated is the negative association between parathyroid hormone and 25-D levels. This association is often used to assert that high levels of 25-D (e.g., 40 –50 ng/ml or more) are optimal. Aloia et al.44 has pointed out that the studies that conclude these high levels of vitamin D are needed fail to require adequate calcium intake, and that is why such high levels are suggested. It should also be considered whether both low 25-D and high PTH are due to the disease process rather than the low 25-D causing the elevated PTH. In addition, only a small percentage of patients with low 25-D have elevated PTH. The low 25-D may be indicating a systemic chronic bacterial infection, and the abnormally high PTH levels in a small percentage of patients may merely be pointing to those cases in which bacteria have infected the parathyroid gland to a greater degree.
In a study comparing vitamin D supplementation with calcium supplementation,45“the effect of calcium on bone loss was blunted in subjects with the highest levels of serum 25OH vitamin D [25-D].” This last finding is supportive of Marshall’s in silico work indicating that high 25-D actually blocks the VDR.6,18 The largest meta-analysis so far clearly showed benefit from calcium supplementation; however, benefit for vitamin D was much less clear.46 No significant benefit for fracture risk was found when comparing vitamin D and calcium to calcium alone, though some differences were found between vitamin D levels.
Another factor that needs to be considered is whether immune suppression is the cause of bone density improvement when high vitamin D levels are used. Immunosuppressive drugs that lower TNF-alpha using antibodies can improve bone density by reducing inflammation.47 High levels of vitamin D supplementation can also lower TNF-alpha48 and suppress the immune response. Thus, it is possible that an increase in bone density from vitamin D supplementation could be the result of immune suppression via TNF reduction, rather than correction of a vitamin D deficiency. TNF-lowering drugs such as infliximab (Remicade) increase risk of cancer and tuberculosis. Thus, the desirability of improving bone density through immune suppression is questionable. This immunosuppressive effect of vitamin D may even explain what seems to be a beneficial effect on falls and muscle strength of elevating vitamin D through supplementation.21 This may be only a symptom reduction in the short-term and may be harmful in the long-term due to the immune suppression.
Autoimmune Disease
In the area of autoimmune disease, the data is equally mixed, and sometimes the larger, more recent studies fail to show any effect of vitamin D levels. For example, a recent large study failed to find an association between serum 25-D levels and the incidence of systemic lupus erythematosis and rheumatoid arthritis.49 Research has found that the average age at which patients acquired rheumatoid arthritis is 12 years earlier in Mexico than in Canada and pointed to the possible role of infectious agents in causing the disease.50 And clearly this study does not support the idea that sun exposure is beneficial for rheumatoid arthritis, since Mexico gets far more sun than Canada.
Although some studies in type 2 diabetes have indicated vitamin D supplementation may be preventive,51 these studies were not randomized and thus are subject to many known and unknown confounding factors affecting a parent’s decision to give a child supplemental vitamin D.13 And even if it were clearly established that vitamin D supplementation reduced the incidence of diabetes in infants and small children, that would not mean that it would help in established disease or older patients, nor would it necessarily mean it is the optimal way to achieve diabetes prevention and long-term health. The positive response of both type 1 and type 2 diabetes patients to the Marshall Protocol11 indicates research on the role of bacteria in diabetes should be a priority.
Influenza and Colds
It has been proposed that vitamin D levels’ decline in winter best accounts for the seasonality of colds and influenza52 and that this potentially supports the need for increased supplementation.52,53 However, new evidence indicates that changes in the viral coat properties can account for the seasonal outbreaks at higher latitudes.36,37 Effects on the airways in dry, cold climates also appear to increase susceptibility to viral and bacterial infections in winter and could contribute to higher winter prevalence of respiratory infections in cold climates.38
Another important point is that the patients being followed on the Marshall Protocol include a number of individuals who report that during the worst period of their chronic illness, they had few or no colds or flu-like illnesses, sometimes for many years at a time. And sometimes this low rate of colds was apparent even years before their illness. This has also been reported in Parkinson’s disease, with the decrease in viral respiratory infections also occurring several years before the disease was diagnosed.54 Thus, even if future research were to establish that vitamin D supplementation reduced colds and influenza, this is by no means an adequate argument for its use. The above observations in chronically ill patients indicate that observing a reduction in respiratory viral infections is not always a sign of good overall health.
Indications of Long-Term Negative Effects of Vitamin D Supplementation
Brannon et al.55 pointed out in a recent report from a roundtable discussion of vitamin D data needs that many studies so far have not yet adequately investigated potential negative consequences such as soft tissue calcification. Vitamin D has been implicated in arterial calcification in the past56 as well as other negative effects.13 The report by the roundtable of vitamin D experts expressed concern that many studies may be shortsighted with regard to adverse outcomes.55
A disturbing new study showed a highly significant correlation (p=0.007) between increased vitamin D intake from food and supplements and the volume of brain lesions shown by MRI in elderly adults.57 In the multivariable regression model, vitamin D intake retained its significant correlation with brain lesion volume even after the effects of calcium were statistically removed. However, calcium did not retain a significant independent correlation with the lesions when the study controlled for vitamin D. Thus, the analysis points to vitamin D supplementation as the key factor in higher lesion volume in this study. These types of brain lesions have been linked to adverse effects in many studies, e.g., stroke,58 psychiatric disorders,59,60 brain atrophy,61 and earlier death.62 Interestingly, the levels of vitamin D intake were not particularly high by some standards, with the highest intake estimated at 1015 mg daily (mean of 341 mg), about half coming from supplements and the rest from food.
The correlation between vitamin D intake and brain lesions seems to lend further support to Marshall’s work. In another study, the finding that over a three-year period, a small percentage of patients were found to have a slight regression of their brain lesions,63 leaves room for hope that the lesions are potentially reversible. Reversibility would be in accord with the improvement of depression and cognitive deficits and other neurological symptoms reported in patients on the Marshall Protocol.6,64
Elusive Bacterial Pathogens Are Detected with Improved Methods
Over many decades, researchers have reported evidence that hard-to-detect bacterial infections are the cause of many diseases,65,66 including autoimmune disease,65-68 cardiovascular disease,69-71 and even cancer.72-77 Some have noted the recent trend toward finding more infectious causes of disease and suggested this is likely to increase in the coming years.6,71,77-80
Recently, Barry Marshall received the Nobel Prize for discovering that the bacteria Helicobacter pylori causes ulcers. And it is now known that H. pylori is a causal factor in stomach cancer.77
New techniques using 16s ribosomal RNA shotgun sequencing,81,82 as well as more advanced culturing and observational techniques65,66,80,83-85 are suggesting that, up until now, most microbiologists have failed to detect a large percentage of potential disease-causing agents. “Persister” cells have been identified that escape antibiotic treatment.86 Cell wall deficient organisms have long been studied,65-66and just recently, advances have been made in understanding their structure and in culturing techniques.80 Research is also indicating that a bacterial biofilm-like microbiota of multiple species even exists within human cells.6,8
Bacteria that grow on a surface in a multi-species community, protected by both a biofilm and the combined effect of their individual resistance strategies, have been a growing area of research.79 Bacterial biofilms have been found to cause the non-healing ulcers in diabetics and may be successfully treated using novel approaches, thus reducing the need for limb amputation.88
Other examples of studies detecting unexpected bacterial pathogens include work linking pathogens in amniotic fluid to pre-term birth89 and research showing numerous previously undetected species in the biofilms that coat prosthetic hip joints.82 Many species of bacteria have been in wounds that were previously undetected using older techniques.81 Macfarlane et al.90 used a combination of more advanced techniques to study bacteria in biofilm communities in patients with Barrett’s esophagus, a pre-cancerous condition. Their methods revealed significant differences between patients and controls in the types and numbers of bacterial species, differences that were previously undetected using older techniques.
Increasingly, inflammation is observed in chronic diseases ranging from depression to cardiovascular disease and cancer.87 The above trends, when combined with observations of bacteria in numerous diseases6,13,65,66,71,91 and the success of the anti-bacterial protocol developed by Marshall6,8,11,13 suggest an extensive role for previously unidentified chronic bacterial infections.
Research is also supporting the ineffectiveness of most standard antibiotic protocols against these bacteria70 and suggesting why other approaches may work better. For instance, some antibiotics target cell walls, and this actually promotes the production of cell wall deficient forms of bacteria that resist many antibiotics.80 Furthermore, many antibiotics are known to inhibit phagocytosis and other aspects of the immune response when taken at high, constant dosages.92
The ability of bacteriostatic antibiotics such as clindamycin to be effective at low doses has been documented.93,94 The survival of “persister” cells mean that pulsed antibiotics are likely to be more effective.86 And fascinating investigations of biofilm communities have revealed many ways in which bacteria can resist antibiotics when used in traditional ways.95 The existence of communities of many bacterial species means that combinations of antibiotics are probably needed to be effective against all the species present. Thus, there is increasing support for the use of pulsed, low dosages of combinations of bacteriostatic antibiotics as used in the anti-bacterial protocol discussed here.
What is particularly encouraging is that the effectiveness of Marshall’s protocol in many systemic chronic disease indicates that these elusive pathogens do respond to select currently available bacteriostatic antibiotics when innate immune function is restored through restoring vitamin D receptor function.6,11 Not only do the bacterial infections appear to resolve, the evidence so far suggests that the improved immune response leads to reduced viral, fungal, and protozoal infections as well.

Conclusions
In silico and clinical data indicate that it is likely that associations between low vitamin D levels and chronic diseases are not evidence of deficiency, but result from a bacteria-induced blockage of the vitamin D receptor, leading to down-regulation of 25-D levels.1,6 According to this model of chronic disease, the short-term benefits sometimes perceived with high vitamin D levels are not due to correction of a vitamin D deficiency but due to suppression of bacterial killing and the immunopathological reaction that accompanies it. Data on reversal of a range of inflammatory and autoimmune diseases through an anti-bacterial protocol that includes vitamin D avoidance and a VDR agonist support this view.6,11
As discussed in detail above, it appears that increasing vitamin D supplementation is not the answer to these chronic diseases and is likely to be counter-productive. Other researchers have also raised concerns regarding vitamin D supplementation’s potential adverse effects. Potential dangers include increased aortic calcification55,56 and brain lesions shown by MRI57 (also see above). In addition, some studies have even found evidence of increased danger from cancer in association with higher levels of vitamin D.32,33,39,40,42
Many have been attracted to the area of vitamin D research, recognizing interesting patterns and responses to supplementation that at first seemed to indicate widespread deficiency and, at the very least, indicate that vitamin D plays a powerful role in physiological processes. Great strides have been made in the last 30 years by scientists with a range of perspectives, and this has led to great excitement and a laudable commitment to use that knowledge to help patients.
However, new genomic and molecular research and the positive response to a new anti-bacterial protocol that involves the avoidance of vitamin D indicate the need for a reappraisal of the data gathered so far. It appears that attempting to raise 25-D through vitamin D supplementation or sun exposure is not the right approach to many, if not most, common chronic diseases. Instead, as discussed above, the evidence supports the effectiveness of a new protocol in restoring vitamin D receptor function, which appears to be a crucial factor in recovery.
One of the most commendable attributes of a truly objective scientist is the willingness to be open to changing long-held positions in the light of new evidence. It will be interesting to see how many have this all-too-rare quality, as research and discussion of vitamin D and the VDR continues. It is to be hoped that the tremendous healing potential likely to be available from eliminating the pathogens that cause chronic disease will inspire an especially high level of open-minded discussion and cooperation.
Caution: The immunopathological reactions from killing the high levels of bacteria that have accumulated in chronically ill patients can be severe and even life-threatening, and thus the Marshall Protocol must be done very carefully and slowly, according to the guidelines.7,96 For the sake of safety, antibiotics must be started at quite low dosages, starting with only one antibiotic. Health care providers are responsible for the use of this information. Neither Autoimmunity Research, Inc., nor the author assume responsibility for the use or misuse of this protocol.
Note: Neither the author, Prof. Marshall, nor the non-profit Autoimmunity Research, Inc. have any financial connection with any product or lab mentioned with regard to the Marshall Protocol. The information needed to implement the Marshall Protocol is available free of charge fromwww.AutoimmunityResearch.org.
Vitamin D3 and Its Nuclear Receptor Increase the Expression and Activity of the Human Proton-Coupled Folate Transporter
Folates are essential for nucleic acid synthesis and are particularly required in rapidly proliferating tissues, such as intestinal epithelium and hemopoietic cells. Availability of dietary folates is determined by their absorption across the intestinal epithelium, mediated by the proton-coupled folate transporter (PCFT) at the apical enterocyte membranes. Whereas transport properties of PCFT are well characterized, regulation of PCFT gene expression remains less elucidated. We have studied the mechanisms that regulate PCFT promoter activity and expression in intestine-derived cells. PCFT mRNA levels are increased in Caco-2 cells treated with 1,25-dihydroxyvitamin D3 (vitamin D3) in a dose-dependent fashion, and the duodenal rat Pcft mRNA expression is induced by vitamin D3 ex vivo. The PCFTpromoter region is transactivated by the vitamin D receptor (VDR) and its heterodimeric partner retinoid X receptor-α (RXRα) in the presence of vitamin D3. In silico analyses predicted a VDR response element (VDRE) in the PCFT promoter region −1694/−1680. DNA binding assays showed direct and specific binding of the VDR:RXRα heterodimer to the PCFT(−1694/−1680), and chromatin immunoprecipitations verified that this interaction occurs within living cells. Mutational promoter analyses confirmed that the PCFT(−1694/−1680) motif mediates a transcriptional response to vitamin D3. In functional support of this regulatory mechanism, treatment with vitamin D3 significantly increased the uptake of [3H]folic acid into Caco-2 cells at pH 5.5. In conclusion, vitamin D3 and VDR increase intestinal PCFT expression, resulting in enhanced cellular folate uptake. Pharmacological treatment of patients with vitamin D3 may have the added therapeutic benefit of enhancing the intestinal absorption of folates.
Folates are water-soluble B vitamins that act as one-carbon donors required for purine biosynthesis and for cellular methylation reactions. They are essential for de novo synthesis of nucleic acids, and thus for production and maintenance of new cells, particularly in rapidly dividing tissues such as bone marrow and intestinal epithelium (Kamen, 1997). Adequate dietary folate availability is especially important during periods of rapid cell division, such as during pregnancy and infancy. Folate deficiency has been associated with reduced erythropoiesis, which can lead to megaloblastic anemia in both children and adults (Ifergan and Assaraf, 2008). Deficiency of folate availability in pregnant women has been linked to neural tube defects, such as spina bifida, in children (Pitkin, 2007). This has prompted the application of folate supplementation schemes either as pills or via fortification of grain products with folates (Eichholzer et al., 2006). Folates have also been proposed to act as protective agents against colorectal neoplasia, although contradictory results have also been reported (Sanderson et al., 2007).
The availability of diet-derived folates is primarily determined by the rate of their uptake into the epithelial cells of the intestine, mediated by the proton-coupled folate transporter (PCFT, gene symbol SLC46A1), localized at the apical brush-border membranes of enterocytes (Subramanian et al., 2008a). PCFT is an electrogenic transporter that functions optimally at a low pH (Qiu et al., 2006;Umapathy et al., 2007). Despite being abundantly expressed in enterocytes, the second folate transporter, termed reduced folate carrier (RFC, gene symbolSLC19A1), has recently been shown not to play an important role in intestinal folate absorption (Zhao et al., 2004; Wang et al., 2005).
The human PCFT gene resides on chromosome 17, contains 5 exons, and is expressed as two prominent mRNA isoforms of 2.1 and 2.7 kilobase pairs (Qiu et al., 2006). Mutations in the PCFT gene have been associated with hereditary folate malabsorption, a rare autosomal recessive disorder (Qiu et al., 2006; Zhao et al., 2007). The PCFT protein is predicted to have a structure harboring 12 transmembrane domains (Qiu et al., 2007; Subramanian et al., 2008a). Although the transport function of PCFT has been studied extensively, relatively little is known about the regulation of PCFT gene expression. PCFT promoter activity has been shown possibly to be epigenetically regulated by its methylation status in human tumor cell lines (Gonen et al., 2008). Furthermore, both the PCFT mRNA expression levels and PCFT promoter activity positively correlate with the level of differentiation of colon-derived Caco-2 cells (Subramanian et al., 2008b).
In addition to its well known roles in regulating calcium homeostasis and bone mineralization, 1,25-dihydroxyvitamin D3 (vitamin D3), the biologically active metabolite of vitamin D, executes many other important functions, particularly in the intestine. For example, vitamin D3 promotes the integrity of mucosal tight junctions (Kong et al., 2008). Many effects of vitamin D3 are mediated via its action as a ligand for the vitamin D receptor (VDR; gene symbol NR1I1), a member of the nuclear receptor family of transcription factors (Dusso et al., 2005). VDR typically regulates gene expression by directly interacting with so-called direct repeat-3 (DR-3; a direct repeat of AGGTCA-like hexamers separated by three nucleotides) motifs within the target promoters, as a heterodimer with another nuclear receptor, retinoid X receptor-α (RXRα; gene symbol NR2B1) (Haussler et al., 1997). Genetic variants of VDR have been associated with inflammatory bowel disease (Simmons et al., 2000; Naderi et al., 2008). Similarly to folates, both VDR and its ligand vitamin D3 have been proposed to be protective against intestinal neoplasia (Ali and Vaidya, 2007). Dietary folate intake has been suggested to regulate gene expression of the components of the vitamin D system, possibly via epigenetic control through the function of folates as methyl donors (Cross et al., 2006). Several intestinally expressed transporter genes, such as those encoding the multidrug resistance protein 1 and multidrug resistance-associated protein 2, have recently been shown to be induced by vitamin D3 (Fan et al., 2009). We investigated whether vitamin D3 regulates the expression of the PCFT gene, encoding a transporter crucial for intestinal folate absorption. The human well polarized enterocyte-derived Caco-2 cells exhibit many of the characteristics associated with mature enterocytes and were used here to investigate the effects of vitamin D3 on PCFT gene expression and folate transport activity.
……..
Vitamin D3 regulates the expression of its target genes primarily by acting as an agonistic ligand for its DNA-binding nuclear receptor VDR, although nongenomic actions by vitamin D3 have also been described previously (Christakos et al., 2003;Dusso et al., 2005). VDR, an important regulator of differentiation and proliferation of enterocytes, typically activates gene expression by heterodimerizing with its nuclear receptor partner RXRα. VDR:RXRα heterodimers then directly bind to DR-3-like elements on the target genes. It should be noted that other modes of VDR-mediated regulation, either via direct interaction with other DNA-binding factors or through nongenomic actions, have also been reported (Dusso et al., 2005).
Here we demonstrate that VDR is a ligand-dependent transactivator of the humanPCFT gene, coding for a vital transporter for intestinal absorption of dietary folates. PCFT mRNA is also abundantly expressed in the liver (Qiu et al., 2006). However, VDR is expressed at very low levels in primary human hepatocytes or hepatocyte-derived cell lines (Gascon-Barre et al., 2003; data not shown), suggesting that VDR-mediated regulation of the PCFT gene may not occur in hepatocytes.
Endogenous PCFT mRNA levels were induced by vitamin D3 in a dose-dependent manner in Caco-2 cells (Fig. 1A). This increase was not further enhanced by cotreatment of cells with the RXRα ligand 9-cis retinoic acid (data not shown), consistent with a previous report that VDR:RXRα heterodimers, at least in some promoter contexts, may not respond to RXRα ligands (Forman et al., 1995). Alternatively, saturating levels of RXRα ligands may already be endogenously present in cells in these experimental conditions. In transient transfection assays, the PCFT promoter fragment −2231/+96 exhibited significant response to exogenous expression of VDR alone in the presence of its ligand (Fig. 2), most probably supported by endogenously expressed RXRα in Caco-2 cells.
Supporting the importance of the VDR:RXRα heterodimer formation for PCFTpromoter regulation, the luciferase values were further significantly elevated upon exogenous expression of RXRα. Exogenous expression of VDR in the absence of vitamin D3 did not notably influence the activity of the PCFT(−2231/+96) promoter, indicating ligand-dependence of VDR action. In deletional transfection analysis, the strongest induction in response to VDR and RXRα in the presence of their ligands was achieved with the PCFT(−2231/+96) promoter fragment (Fig. 3A). Induction of the shortest deletion variant tested [PCFT(−843/+96)luc] was approximately 50% of that achieved for the PCFT(−2231/+96), indicating that this more proximal region is likely to contain further DNA elements mediating a response to vitamin D3. However, in our current study, we focused on the distal region between the nucleotides −2231 and −1674 upstream of the transcriptional start site of the human PCFT gene, which confers maximal response to vitamin D3. In our computational analysis, we identified a putative VDRE within the PCFTpromoter region between nucleotides −1694 and −1680. We have not so far been successful in identifying further binding sites for the VDR:RXRα heterodimer in the more proximal region of the PCFT promoter. It may be that, in addition to direct DNA-binding to the PCFT(−1694/−1680) element identified here, VDR may also affect PCFT promoter activity indirectly, via interactions with other DNA-binding factors. For example, it has been proposed that the p27Kip1 gene is regulated by VDR via response elements for unrelated DNA-binding transcription factors Sp1 and NF-Y (Huang et al., 2004).
Both endogenously expressed and recombinant VDR and RXRα bound to thePCFT(−1694/−1680) element specifically and as obligate heterodimers (Fig. 4). The interaction between VDR and this region of the PCFT promoter within living cells treated with VDR and RXRα ligands was confirmed by chromatin immunoprecipitation tests (Fig. 5). Heterologous promoter assays proved that thePCFT(−1694/−1680) element can function as an independent VDR response element. The significant decrease in VDR:RXRα-mediated induction upon mutagenesis of the PCFT(−1694/−1680) element confirmed that it is an important functional mediator of the effect (Fig. 6, A and B).
Although we observed vitamin D3-mediated increase of rat Pcft mRNA expression ex vivo (Fig. 1C), the rat Pcft promoter (chromosome 10; GenBank accession number NW_047336) exhibits no significant overall homology with the humanPCFT promoter over the proximal 3000-bp regions. This suggests that despite the divergence of the promoter sequences between human and rodent PCFT/Pcftgenes, the functional response to vitamin D3 is conserved.
The activation of PCFT gene transcription by VDR also translates into an increase in PCFT protein function. Vitamin D3 treatment of Caco-2 cells led to significantly increased uptake of folate across the apical membrane, in a dose-dependent manner (Fig. 7). In keeping with the fact that PCFT strongly prefers an acidic milieu for its transport function (Qiu et al., 2006; Nakai et al., 2007; Unal et al., 2009), we only observed vitamin D3-stimulated transport activity at pH5.5, but not at neutral pH. These data strongly suggest that vitamin D3-mediated transcriptional activation of PCFT gene expression leads to an increase of PCFT transport function. Consistent with our model, mRNA expression of the other known folate carrier expressed in Caco-2 cells, RFC, which functions efficiently at neutral pH (Ganapathy et al., 2004; Wang et al., 2004), was not affected by vitamin D3treatment (Fig. 1B). It has been reported that vitamin D3-induced gene expression increases as Caco-2 cells differentiate (Cui et al., 2009). Thus, our current findings on VDR-mediated regulation of PCFT expression provide a possible molecular mechanism for a prior observation that folate uptake into Caco-2 cells is enhanced upon confluence-associated differentiation (Subramanian et al., 2008b).
Our results suggest that intestinal folate absorption may be enhanced by an increase in dietary vitamin D3 intake. Food products are often supplemented with folates, because of their proposed beneficial health effects. Based on our current study, supplementation of vitamin D3 may enhance the intestinal absorption of folates. PCFT also transports the antifolate drug methotrexate (MTX) (Inoue et al., 2008; Yuasa et al., 2009) widely used in the treatment of autoimmune diseases and cancer. MTX interferes with folate metabolism by competitively inhibiting the enzyme dihydrofolate reductase. Our results may further suggest a potential mechanism to increase intestinal absorption of MTX via simultaneous treatment with vitamin D3, thereby affecting the bioavailability of MTX. Patients suffering from inflammatory bowel disease are frequently on long-term treatment with calcium and vitamin D3 as a prophylaxis against osteopenia and osteoporosis (Lichtenstein et al., 2006). This patient group is frequently treated with folates (in the case of folate deficiency) or MTX (as a second-line immunosuppressant) (Rizzello et al., 2002). MTX therapy per se requires prophylactic administration of folates, and these patients often receive additional calcium/vitamin D3. Our current results may warrant a closer investigation into potential drug-drug interactions between pharmacologically administered vitamin D3, MTX, and folates. Taking into account the previous report that folates regulate the expression of genes involved in vitamin D3 metabolism, it may be that folate and vitamin D3 homeostasis are closely interlinked through such mutual regulatory interactions.
Innate immune response and Th1 inflammation
http://mpkb.org/home/pathogenesis/innate_immunity
The innate immune response is the body’s first line of defense against and non-specific way for responding to bacterial pathogens.1 Located in the nucleus of a variety of cells, the Vitamin D nuclear receptor (VDR) plays a crucial, often under-appreciated, role in the innate immune response.
When functioning properly, the VDR transcribes between hundreds2 and thousands of genes3including those for the proteins known as the antimicrobial peptides. Antimicrobial peptides are “the body’s natural antibiotics,” crucial for both prevention and clearance of infection.4The VDR also expresses the TLR2 receptor, which is expressed on the surface of certain cells and recognizes foreign substances.
The body controls activity of the VDR through regulation of the vitamin D metabolites. 25-hydroxyvitamin D (25-D) antagonizes or inactivates the Receptor while 1,25-dihydroxyvitamin D (1,25-D) agonizes or activates the Receptor.
Greater than 36 types of tissue have been identified as having a Vitamin D Receptor.5
Another component of the innate immune response is the release of inflammatory cytokines. The result is what medicine calls inflammation, which generally leads to an increase in symptoms.
Before the Human Microbiome Project, scientists couldn’t link bacteria to inflammatory diseases. But with the advent of DNA sequencing technology, scientists have detected many of the bacteria capable of generating an inflammatory response. All diseases of unknown etiology are inflammatory diseases.
Nuclear receptors and ligands
Nuclear receptors are a class of proteins found within the interior of cells that are responsible for sensing the presence of hormones and certain other molecules. A unique property of nuclear receptors which differentiate them from other classes of receptors is their ability to directly interact with and control the expression of genomic DNA. Some of the molecules (or ligands) which bind the nuclear receptor activate (agonize) it and some inactivate (antagonize) it.
It is commonly accepted that most ligands, approximately 95% to 98%, inactivate the nuclear receptors. Since the nuclear receptors play a significant role in the immune response, this factor alone may explain why so many drugs and substances found in food and drink are immunosuppressive.
Because the expression of a large number of genes is regulated by nuclear receptors, ligands that activate these receptors can have profound effects on the organism. Many of these regulated genes are associated with various diseases which explains why the molecular targets of approximately 13% of FDA approved drugs are nuclear receptors.6
Different cell types have different nuclear receptors. One of the nuclear receptors seen in immune cells is the Vitamin D Receptor (VDR). The VDR has two endogenous or “native” ligands, which are also the two main forms of vitamin D in the human body: 25-hydroxyvitamin D (25-D) and 1,25-dihydroxyvitamin D (1,25-D). Non-native or exogenous ligands can also inactivate or activate a nuclear receptor, depending on its molecular structure.
Ligands compete to dock at nuclear receptors. When is a given kind of ligand such as 25-D as opposed to 1,25-D more likely to bind to the VDR? It depends. 1,25-D tends to be much less common than 25-D – by a factor of 1,000 or more – so it binds to the receptor much more infrequently. A greater concentration of a given molecule can displace competing molecules off the nuclear receptor. Affinity occurs in logarithmic fashion, which is to say that it operates on the basis of a sliding scale. In short, an increase in 1,25-D and a decrease in 25-D can tilt the odds in favor of 1,25-D, and vise versa.
Affinity as well as the question of whether a ligand inactivates or activates a nuclear receptor can all be validated using in silicomodeling. Although less precise, it is also possible to measure these properties in vitro.

Activated by 1,25-D and inactivated by 25-D, the Vitamin D nuclear receptor (VDR) transcribes a number of genes crucial to the function of the innate immune response.
Role of Vitamin D Receptor in innate immunity
Vitamin D/VDR have multiple critical functions in regulating the response to intestinal homeostasis, tight junctions, pathogen invasion, commensal bacterial colonization, antimicrobe peptide secretion, and mucosal defense…. The involvement of Vitamin D/VDR in anti-inflammation and anti-infection represents a newly identified and highly significant activity for VDR.
Jun Sun 7
When activated by 1,25-D, the Vitamin D Receptor (also called the calcitriol receptor) transcribes thousands of genes.8 It is commonly known that the VDR functions in regulating calcium metabolism.9 It is becoming increasingly clear, however, that the clinically accepted role of the Vitamin D metabolites, that of regulating calcium homeostasis, is just a small subset of the functions actually performed by these hormones.
Transcription of antimicrobial peptides
One of the VDR’s key functions is the transcription of antimicrobial peptides.10 11 See below.
Other antimicrobial activity of the VDR
Additionally, when the VDR is activated, TLR2 is expressed.12 TLR2 is a receptor, which is expressed on the surface of certain cells and recognizes native or foreign substances, and passes on appropriate signals to the cell and/or the nervous system.
When activated TLR2 allows the immune system to recognize gram-positive bacteria, including Staphylococcus aureus13 14Chlamydia pneumoniae15 and Mycoplasma pneumoniae.16 TLR2 also protects from intracellular infections such as Mycobacteria tuberculosis.17
Antimicrobial peptides
The antimicrobial peptides (AMPs), of which there are hundreds, are families of proteins, which have been called “the body’s natural antibiotics,” crucial for both prevention and clearance of infection. AMPs are broad-spectrum, responding to pathogens in a non-specific manner.18
For example, consider cathelicidin, a protein transcribed the VDR, which not unlike a Swiss Army knife, has many different functions. Because it can be differentially spliced, the cathelicidin protein itself can respond to a range of very different microbial challenges. In humans, the cathelicidin antimicrobial peptide gene encodes an inactive precursor protein (hCAP18) that is processed to release a 37amino-acid peptide (LL-37) from the C-terminus. LL-37 is susceptible to proteolitic processing by a variety of enzymes, generating many different cathelicidin-derived peptides, each of which has specific targets. For example, LL-37 is generated in response toStaphylococcus aureus, yet LL-37 represents 20% of the cathelicidin-derived peptides, with the smaller peptides being much more abundant and able to target even more diverse microbial forms.19
AMPs have been documented to kill bacteria and disrupt their function through the following modes of action:
-
interfering with metabolism
-
targeting cytoplasmic components
-
disrupting membranes
-
act as chemokines and/or induce chemokine production, which directs traffic of bacteria
Also, AMPs aid in recovery from infection by:
-
promoting wound healing
-
inhibiting inflammation
In many cases, the exact mechanism by which antimicrobial peptides kill bacteria is unknown. In contrast to many conventional antibiotics including those used by the Marshall Protocol, AMPs appear to be bacteriocidal (a killer of bacteria) instead of bacteriostatic (an inhibitor of bacterial growth).
Two of the more significant families of AMPs are cathelicidin and the beta-defensins. Of these two families, cathelicidin is the most common.
The full extent by which microbes interfere with AMP expression is the subject of a rapidly growing body of research.20 21 22
Antimicrobial peptides target fungi and viruses
The antimicrobial peptides play a role in mitigating the virulence of the virome and other non-bacterial infectious agents. In addition to its antibacterial activity, alpha-defensin human neutrophil peptide-1 inhibits HIV and influenza virus entry into target cells.23 It diminishes HIV replication and can inactivate cytomegalovirus, herpes simplex virus, vesicular stomatitis virus and adenovirus.24 In addition to killing both gram positive and gram-negative bacteria, human beta-defensins HBD-1, HDB-2, and HBD-3 have also been shown to kill the opportunistic yeast species Candida albicans.25 Cathelicidin also possesses antiviral and antifungal activity.26 27
In other words, there is a reason why this group of proteins are named antimicrobial peptides rather than antibacterial peptides.
Unexpected antimicrobial peptides
There are now several examples of substances believed to cause disease, which have since been proven to be part of host defense.
Evolutionarily conserved
The TLR2/1 and cathelicidin-vitamin D pathway has long played a “powerful force” in protecting the body against infection. This is evidenced by the fact that the Alu short interspersed element (SINE), which transcribes the vitamin D receptor binding element (VDRE), has been evolutionarily conserved for 55-60 million years, but not prior.30 The differences in this pathway between humans/primates and other mammals call into question animal models that try to emulate the vitamin D system and indeed the immune system.
Inflammation
Another component of the innate immune response is inflammation, the universal initial response of the organism to any injurious agent.31 Inflammation is a systemic physiological process fundamental for survival.32 The identification of bacteria and other pathogens triggers the release of inflammatory cytokines. These cytokines include interferon-gamma, tumor necrosis factor-alpha (TNF-alpha), and Nuclear Factor-kappa B (NF-kappaB). Cytokines are regulatory proteins, such as the interleukins and lymphokines, that are released by cells of the immune system and act as intercellular mediators in the generation of an immune response. The result is what medicine calls inflammation, which generally leads to an increase in symptoms.
Th1/Th17 inflammation
One key type of inflammation is the Th1/Th17 (T-helper) inflammatory response. In the interests of concision, the Th1/Th17, on this site and others, the Th1/Th17 response is referred to as the Th1 response. This reaction occurs in response to intracellular pathogens, which according to the Marshall Pathogenesis, play a driving force in chronic disease.
All Th1 diseases are marked by an inflammatory response
Before the Human Microbiome Project, scientists couldn’t consistently link bacteria to inflammatory diseases. But with the advent of DNA sequencing technology, scientists have detected many of the bacteria capable of generating an inflammatory response. All diseases of unknown etiology are inflammatory diseases.
An inflammatory immune response—one of the body’s primary means to protect against infection—defines multiple established infectious causes of chronic diseases, including some cancers. Inflammation also drives many chronic conditions that are still classified as (noninfectious) autoimmune or immune-mediated (e.g., systemic lupus erythematosus, rheumatoid arthritis, Crohn’s disease). Both [the innate and adaptive immune systems] play critical roles in the pathogenesis of these inflammatory syndromes. Therefore, inflammation is a clear potential link between infectious agents and chronic diseases.
Siobhán M. O’Connor et al. 33
Th2 inflammation
According to the Marshall Pathogenesis, generally speaking, any activity of the Th2 cytokines in chronic disease is a result of the primary Th1-inducing pathogens.
Many palliative therapies interfere with inflammation
While inflammation is associated with disease, inflammation often serves an invaluable role as the immune system fights off chronic pathogens. Numerous medications artificially suppress inflammation including anti-TNF drugs, interferon, corticosteroids, antifungals, and anti-pyreutics. While interfering with the inflammatory response typically reduces immunopathology and makes a patient feel less symptomatic in the near term, doing so allows the bacteria which cause chronic disease to proliferate.
The release of cytokines appears to be essential for recovery after an infection. One study found that the cytokine TNF-alpha – which is blocked by anti-TNF drugs – is necessary for the proper expression of acquired specific resistance following infection withMycobacterium tuberculosis.34 35 36 Another effect of the use of TNF blockers is to break or reduce the formation of granuloma, one of the body’s mechanisms to control bacterial pathogens.37
Commensal microbes
The host innate immune defense system is highly active in healthy tissue.38 Commensal bacteria can activate innate immune responses.39 40
References
Role of Dihydroxyvitamin D3 and Its Nuclear Receptor in Novel Directed Therapies for Cancer
S. Ondková, D. Macejová and J. Brtko
Gen. Physiol. Biophys. (2006), 25, 339—353 http://www.gpb.sav.sk/2006_04_339.pdf
Dihydroxyvitamin D3 is known to affect broad spectrum of various biochemical and molecular biological reactions in organisms. Research on the role and function of nuclear vitamin D receptors (VDR) playing a role as dihydroxyvitamin D3 inducible transcription factor belongs to dynamically developing branches of molecular endocrinology. In higher organisms, full functionality of VDR in the form of heterodimer with nuclear 9-cis retinoic acid receptor is essential for biological effects of dihydroxyvitamin D3. This article summarizes selected effects of biologically active vitamin D3 acting through their cognate nuclear receptors, and also its potential use in therapy and prevention of various types of cancer.
……
Vitamin D family consists of 9,10-secosteroids which differ in their side-chain structures. They are classified into five forms: D2, ergocalciferol; D3, cholecalciferol; D4, 22,23-dihydroergocalciferol; D5, sitosterol (24-ethylcholecalciferol) and D6, stigmasterol (Napoli et al. 1979). The main forms are vitamin D2 (ergocalciferol: plant origin) and vitamin D3 (cholecalciferol: animal origin). Both 25-hydroxyvitamin D2 and 1α,25-dihydroxyvitamin D2 have been evaluated for their biological functions. Vitamin D itself is a prohormone that is metabolically converted to the biologically active metabolite, 1,25-dihydroxyvitamin D3 in kidney. This vitamin D3, currently considered a steroid hormone, activates its cognate nuclear receptor (vitamin D receptor or VDR) which alter transcription rates of the target genes responsible for its biological responses. In general, vitamin D is essential for mineral homeostasis, for absorption and utilization of both calcium and phosphate and it aids in the mobilization of bone calcium and maintenance of serum calcium concentrations. Through these function, it plays an important role in ensuring proper functioning of muscles, nerves, blood clotting, cell growth and energy utilization. It has been proposed that vitamin D is also important for insulin and prolactin secretion, immune and stress responses, melanin synthesis and for differentiation of skin and blood cells (Lips 2006). Vitamin D metabolites also play a role in the prevention of auto-immune diseases and cancer (Pinette et al. 2003; Dusso et al. 2005). The steroid hormone 1α,25-dihydroxyvitamin D3 (calcitriol) exerts biological responses by interaction with both the well-characterized nuclear receptor (VDRnuc) responsible for activation gene transcription and not fully characterized membrane-associated protein/receptor (VDRmem) involved in generating a variety of rapid, non-genotropic responses (Evans 1988; Norman et al. 2002).
Vitamin D metabolism
Vitamin D, the “sunshine” vitamin, is synthesized under the influence of ultraviolet light in the skin. Many mammals have provitamin D (7-dehydrocholesterol) which is converted to provitamin D3 in their skin. When human skin is exposed to sunlight, the UV-B photons (wavelengths 290–315 nm) interact with 7-dehydrocholesterol causing photolysis and cleavage of the B-ring of the steroid structure, which upon thermoisomerization yields a secosteroid. Thus, provitamin D3 which is inherently unstable rapidly converts by a temperature-dependent process to vitamin D3 (MacLaughlin et al. 1982; Holick 1994). Vitamin D3 enters the blood circulation and binds to vitamin D binding protein (DBP) (Haddad et al. 1993) which carries vitamin D3 to liver and kidney for bioactivation (Wikvall 2001). In the first activation step, vitamin D3 is hydroxylated by the enzyme 25-hydroxylase to 25- hydroxyvitamin D3 mainly in the liver. This metabolite is present in the circulation at the concentration of more than 0.05 µmol/l (20 ng/ml). In the second step, the biologically active hormone 1α,25-dihydroxyvitamin D3 is generated by hydroxylation of 25-hydroxyvitamin D3 at 1α-position in kidney. The enzyme 1α-hydroxylase has been shown to be also present in keratinocytes and prostate epithelial cells, suggesting that those organs may also be able to generate 1α,25-dihydroxyvitamin D3 from 25-dihydroxyvitamin D3 (Schwartz et al. 1998). The activity of 1α-hydroxylase in the kidney serves as the major control point in production of the active hormone. The active metabolite 1α,25-dihydroxyvitamin D3 is present in human plasma at the concentration ranging from 0.05 to 0.15 nmol/l (20–60 pg/ml) (Hartwell et al. 1987; Gross et al. 1996). In general, 90 to 100% of the most human being vitamin D requirement comes from exposure to sunlight (Holick 2003) and the rest of the vitamin D3 content is obtained from diet (Malloy and Feldman 1999). The catabolism of vitamin D occurs by further hydroxylation of 25-dihydroxyvitamin D3 by 24-hydroxylase to yield 24,25-dihydroxyvitamin D3. The 24-hydroxylase is ubiquitous enzyme and is expressed in all the cells expressing VDR. This enzyme is regulated by parathyroid hormone and 1α,25-dihydroxyvitamin D3. The major significance of 24-hydroxylation is inactivation of vitamin D (Nishimura et al. 1994; Brenza and DeLuca 2000). The combinations of 1,25-dihydroxyvitamin D3 with inhibitors of 24-hydroxylase such as ketoconazole or liarozole may enhance its antitumour effects in prostate cancer therapy.
Vitamin D3 receptor
More than 2000 synthetic analogues of the biological active form of vitamin D, 1α,25-dihydroxyvitamin D3, are presently known. Basically, all of them interfere with the molecular switch of nuclear 1α,25-dihydroxyvitamin D3 signalling, which is the complex of the VDR, the retinoid X receptor (RXR), and a 1α,25-dihydroxyvitamin D3 response element (VDRE) (Carlberg 2003).
VDR is the only nuclear protein that binds the biologically most active vitamin D metabolite, 1α,25-dihydroxyvitamin D3, with high affinity (Kd = 0.1 nmol/l). This classifies the VDR into the classical endocrine receptor subgroup of the nuclear receptor superfamily, which also contains the nuclear receptors for hormones as retinoic acid, thyroid hormone, estradiol, progesterone, testosterone, cortisol, and aldosterol (Carlberg 1995). Similarly, like other biologically active ligand for nuclear hormone receptors, 1,25-dihydroxyvitamin D3 can modulate expression of selected ion transport protein genes (Van Baal et al. 1996; Hudecova et al. 2004).
The VDR was first isolated after trancfection of COS-1 cells with cloned sequences of complementary DNA that was isolated from human intestine (Baker et al. 1988). VDR has been found in more than 30 tissues including intestine, colon, breast, lung, ovary, bone, kidney, parathyroid gland, pancreatic β-cells, monocytes, keratinocytes, and many cancer cells, suggesting that the vitamin D endocrine system may also be involved in regulating the immune systems, cellular growth, differentiation and apoptosis (Jones et al. 1998). The active form of vitamin D binds to intracellular receptors that then function as transcription factors to modulate gene expression. Like the receptors for other steroid hormones and thyroid hormones, the VDR has specific hormone-binding and DNA-binding domains. It contains two zinc finger structures forming a characteristic DNA-binding domain (DBD) of 66 amino acids and a carboxy-terminal ligand-binding domain (LBD) of approximately 300 amino acids, which is formed by 12 α-helices. Ligand binding causes a conformational change within the LBD, in which helix 12, the most carboxy-terminal α-helix, closes the ligand-binding pocket via a “mouse-trap like” intramolecular folding (Moras and Gronemeyer 1998). Moreover, the LBD is involved in a variety of interactions with nuclear proteins, such as other nuclear receptors, corepressor and coactivator proteins. These ligand-triggered protein-protein interactions are the central molecular event of nuclear 1α,25-dihydroxyvitamin D3 signalling.
….
Role of vitamin D3 in cancer
Some of biologically active ligands for nuclear receptors exert tumour-suppressive activity, and they have therapeutical exploitation due to their antiproliferative and apoptosis-inducing effects (Brtko and Thalhamer 2003).
…..
During the last decade, evidence for vitamin D3 effects has been accumulating not only for prostate cancer (Feldman et al. 1995; Ma et al. 2004) but also for colon cancer (Cross et al. 1997; Bischof et al. 1998). 1α-hydroxylase was found to be Vitamin D and Cancer Treatment 347 expressed and active in colorectal cancer (Bareis et al. 2001; Cross et al. 2001; Tangpricha et al. 2001; Ogunkolade et al. 2002) and ovarian cancer (Miettinen et al. 2004). In both colon and also lung tumours, CYP24A1 mRNA was significantly up-regulated, while VDR mRNA was generally down-regulated when compared to respective normal tissues. When the level of VDR in 12 malignant colonic tumours was compared with that of adjacent normal tissue, in 9 cases out of 12, expression of VDR in tumours was decreased. However, in that study, the expression of CYP24A1 was not assessed. It has also been shown that, at least in human colon cancer cell lines, the level of VDR correlates with the degree of cell differentiation (Shabahang et al. 1993; Anderson et al. 2006).
Recently, it has been suggested that actually 20–30% of colorectal cancer incidence might be due to insufficient exposure to sunlight. This fact was strengthen by correlation between reduced colorectal cancer incidence and sunlight exposure, low skin pigmentation, nutritional vitamin D intake and high serum levels of 25- hydroxyvitamin D3 (Grant and Garland 2003). In the colon at least, CYP27B1 and VDR expression was described to be actually elevated during early tumour progression and that described dual positivity was found in many, but not all the tumour cells. In human colon tumours, CYP24 mRNA is quite highly expressed and the studies also demonstrated that with the exception of differentiated Caco-2 cells, CYP24 activity is constitutively present or can be induced by 1α,25- dihydroxyvitamin D3. During tumour progression in the colon, not only VDR but CYP27B1 and CYP24 expression were found to be increased in tumour tissues (Bareis et al. 2001; Bises et al. 2004).
Androgens, retinoids, glucocorticoids, estrogens and agonists of peroxisome proliferator-activated receptor directly or indirectly have reasonable impact on vitamin D signalling pathways, and vice versa. It was proposed that sex hormones might reduce colorectal cancer risk (McMichael and Potter 1980). The studies suggested that current and long-term use of estrogens is associated with a substantial decrease in risk of fatal colon cancer. The mechanism, however, by which estrogens could inhibit colonic tumour growth, remains an enigma. There are at least two distinct estrogen receptors in the human body: ERα and ERβ. In the normal human colon, ERβ is widely regarded to be the predominant subtype (CampbellThompson et al. 2001). In a recently terminated pilot study together with Strang Cancer Prevention Centre at Rockefeller University (NY, USA), tissues from postmenopausal women receiving 17β-estradiol for expression of CYP27B1 by real time RT-PCR were examined. CYP27B1 was found to be elevated significantly in all subjects after receiving 17β-estradiol for 4 weeks.
Amplification of chromosomal region 20q12q13 containing the CYP24A1 gene has been reported in ovarian cancer, as well (Tanner et al. 2000). Although inhibition of ovarian cancer cell growth by 1α,25-dihydroxyvitamin D3 has been reported (Saunders et al. 1992, 1995), a clinical trial testing the efficacy of 1α,25- dihydroxyvitamin D3 combined with isotretinoin in treating 22 epithelial ovarian cancer patients for 74 weeks has not produced positive results (Rustin et al. 1996).
Read Full Post »



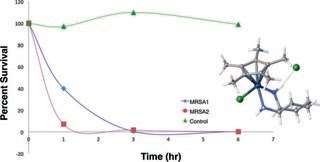




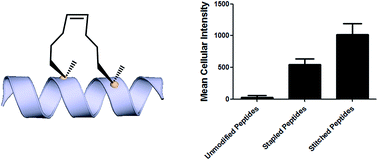
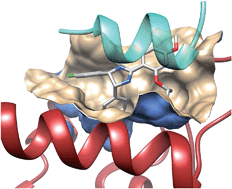

{kind=link}
{kind=link}