Sex Hormones
Author: Larry H Bernstein, MD, FCAP
A steroid hormone is a steroid that acts as a hormone. Steroid hormones can be
grouped into five groups by the receptors to which they bind:
- glucocorticoids,
- mineralocorticoids,
- androgens,
- estrogens, and
- progestogens.
- Vitamin D derivatives, are a sixth closely related hormone system with homologous receptors. They have some of the characteristics of true steroids as receptor ligands.
Steroid hormones help control metabolism, inflammation, immune functions, salt
and water balance, development of sexual characteristics, and the ability to withstand
illness and injury. The term steroid describes both hormones produced by the body
and artificially produced medications that duplicate the action for the naturally occurring steroids
The natural steroid hormones are generally synthesized from cholesterol in the gonads and adrenal glands. These forms of hormones are lipids. They can pass through the cell membrane as they are fat-soluble,[4] and then bind to steroid hormone receptors (which may be nuclear or cytosolic depending on the steroid hormone) to bring about changes within the cell. Steroid hormones are generally carried in the blood, bound to specific carrier proteins such as sex hormone-binding globulin or corticosteroid-binding globulin. Further conversions and catabolism
occurs in the liver, in other “peripheral” tissues, and in the target tissues.
Synthetic steroids and sterols
A variety of synthetic steroids and sterols have also been contrived. Most are
steroids, but some non-steroidal molecules can interact with the steroid receptors
because of a similarity of shape. Some synthetic steroids are weaker or stronger
than the natural steroids whose receptors they activate.
Some examples of synthetic steroid hormones:
Glucocorticoids: alclometasone, prednisone, dexamethasone, triamcinolone
Mineralocorticoid: fludrocortisone
Vitamin D: dihydrotachysterol
Androgens: apoptone, oxandrolone, oxabolone, testosterone, nandrolone (also
known as anabolic steroids)
Estrogens: diethylstilbestrol (DES)
Progestins: danazol, norethindrone, medroxyprogesterone acetate,
17-Hydroxyprogesterone caproate.
Some steroid antagonists:
Androgen: cyproterone acetate
Progestins: mifepristone, gestrinone
http://www.en.wikipedia.org/wiki/Steroid

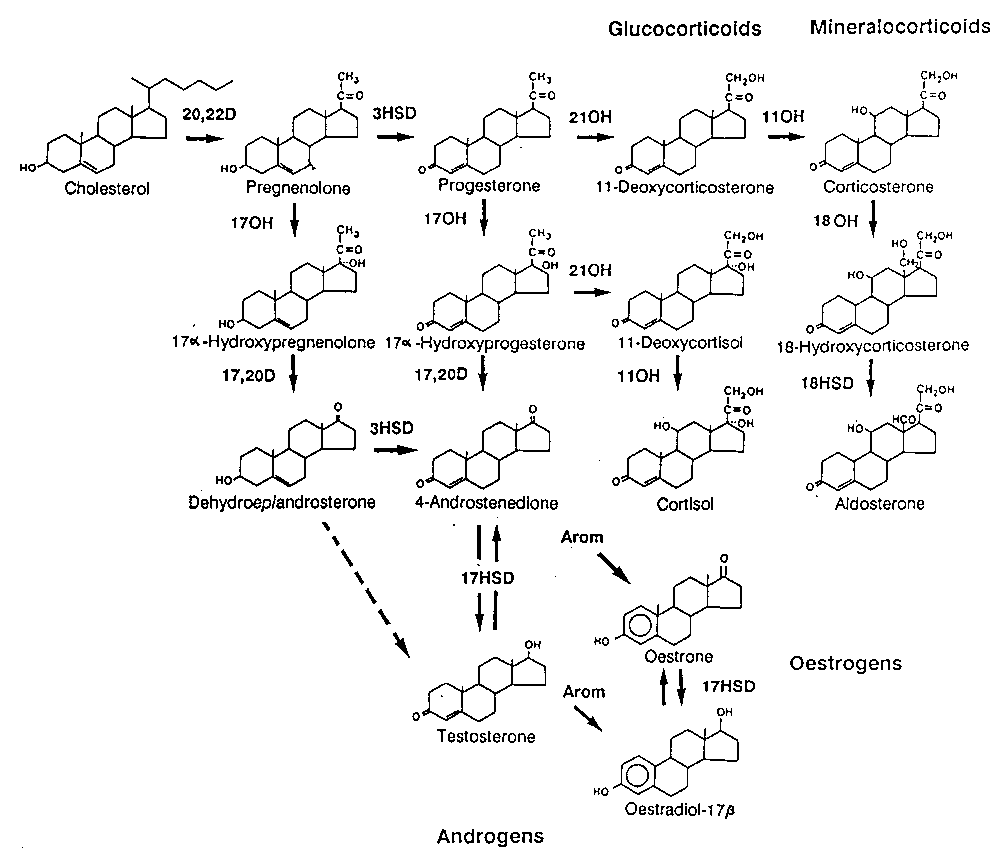
Steroid-Hormone-Synthesis

Steroidogenesis
http://www.gfmer.ch/Books/Reproductive_health/Image171.gif
The regulation of spermatogenesis by androgens
Lee B. Smith, William H. Walker
Seminars in Cell & Developmental Biology 30 (2014) 2–13
http://dx.doi.org/10.1016/j.semcdb.2014.02.012
Testosterone is essential for maintaining spermatogenesis and male fertility.
However, the molecular mechanisms by which testosterone acts have not
begun to be revealed until recently. With the advances obtained from the use
of transgenic mice lacking or overexpressing the androgen receptor, the cell
specific targets of testosterone action as well as the genes and signaling pathways
that are regulated by testosterone are being identified. In this review, the critical
steps of spermatogenesis that are regulated by testosterone are discussed as well
as the intracellular signaling pathways by which testosterone acts. We also review
the functional information that has been obtained from the knock out of the androgen
receptor from specific cell types in the testis and the genes found to be regulated
after altering testosterone levels or androgen receptor expression.
The essence of female–male physiological dimorphism: Differential Ca2+-homeostasis
enabled by the interplay between farnesol-like endogenous sesquiterpenoids and
sex-steroids? The Calcigender paradigm
Arnold De Loof
General and Comparative Endocrinology 211 (2015) 131–146
http://dx.doi.org/10.1016/j.ygcen.2014.12.003
Ca2+ is the most omnipresent pollutant on earth, in higher concentrations a real
threat to all living cells. When [Ca2+]i rises above 100 nM (=resting level), excess
Ca2+ needs to be confined in the SER and mitochondria, or extruded by the different
Ca2+-ATPases. The evolutionary origin of eggs and sperm cells has a crucial, yet
often overlooked link with Ca2+-homeostasis. Because there is no goal whatsoever
in evolution, gametes did neither originate ‘‘with the purpose’’ of generating a progeny
nor of increasing fitness by introducing meiosis. The explanation may simply be that
females ‘‘invented the trick’’ to extrude eggs from their body as an escape strategy
for getting rid of toxic excess Ca2+ resulting from a sex-hormone driven increased
influx into particular cells and tissues.
The production of Ca2+-rich milk, seminal fluid in males and all secreted proteins
by eukaryotic cells may be similarly explained. This view necessitates an upgrade
of the role of the RER-Golgi system in extruding Ca2+. In the context of insect
metamorphosis, it has recently been (re)discovered that (some isoforms of) Ca2+-
ATPases act as membrane receptors for some types of lipophilic ligands, in
particular for endogenous farnesol-like sesquiterpenoids (FLS) and, perhaps, for
some steroid hormones as well.
A novel paradigm, tentatively named ‘‘Calcigender’’ emerges. Its essence is: gender-
specific physiotypes ensue from differential Ca2+-homeostasis enabled by genetic
differences, farnesol/FLS and sex hormones. Apparently the body of reproducing
females gets temporarily more poisoned by Ca2+ than the male one, a selective
benefit rather than a disadvantage.
Sex differences in the expression of estrogen receptor alpha within noradrenergic
neurons in the sheep brain stem
J.L. Rose, A.S. Hamlin, C.J. Scott
Domestic Animal Endocrinology 49 (2014) 6–13
http://dx.doi.org/10.1016/j.domaniend.2014.04.003
In female sheep, high levels of estrogen exert a positive feedback action
on gonadotropin releasing hormone (GnRH) secretion to stimulate a surge in
luteinizing hormone (LH) secretion. Part of this action appears to be via brain
stem noradrenergic neurons. By contrast, estrogen action in male sheep has
a negative feedback action to inhibit GnRH and LH secretion. To investigate
whether part of this sex difference is due to differences in estrogen action in
the brain stem, we tested the hypothesis that the distribution of estrogen
receptor a (ERα) within noradrenergic neurons in the brain stem differs
between rams and ewes. To determine the distribution of ERα, we used
double-label fluorescence immunohistochemistry for dopamine b-Hydroxylase,
as a marker for noradrenergic and adrenergic cells, and ERα. In the ventro-
lateral medulla (A1 region), most ERα-immunoreactive (-ir) cells were
located in the caudal part of the nucleus. Overall, there were more ERα-ir
cells in rams than ewes, but the proportion of double-labeled cells was did
not differ between sexes. Much greater numbers of ERα–ir cells were
found in the nucleus of the solitary tract (A2 region), but <10% were double
labeled and there were no sex differences. The majority of ERα-labeled cells
in this nucleus was located in the more rostral areas. Erα labeled cells were
found in several rostral brain stem regions but none of these were double
labeled and so were not quantified. Because there was no sex difference
in the number of ERα-ir cells in the brain stem that were noradrenergic,
the sex difference in the action of estrogen on gonadotropin secretion in
sheep is unlikely to involve actions on brain stem noradrenergic cells.
Androgens, estrogens, and second messengers
William Rosner, DJ Hryb, MS Khan, AM Nakhla, and NA Romas
Steroids 1998; 63:278 –281 PII S0039-128X(98)00017-8
Over the course of the last four decades, a detailed understanding of the
molecular mechanisms by which steroid hormones exert their effects has
evolved, and continues to evolve. The major focus of research in this area
has been on the manner in which steroid receptors activate transcription.
Pathways of steroid action other than by direct interaction with intracellular
receptors have received relatively little attention. However, there is a growing
body of evidence that steroid hormones exert effects through mechanisms
in addition to those involving their classic intracellular receptors. One such
mechanism is based on the observation that a number of cells have receptors
on their plasma membranes for the plasma protein, sex hormone binding
globulin (SHBG). It is the purpose of this review to briefly describe our current
knowledge of this system.
SHBG binds to a receptor (RSHBG) on cell membranes cAMP and the steroid-SHBG-RSHBG system
Biology of the SHBG-RSHBG system
Relation between the affinity of steroid for SHBG and its potency in inhibiting
the binding of SHBG to RSHBG.
KA (SHBG) = Association constant for SHBG and the indicated steroid.
Ki SHBG-RSHBG = The inhibition constant for the indicated steroid on the
binding of SHBG to RSHBG.
PSA secretion was stimulated by DHT. Although estradiol alone had no effect
on PSA secretion, it caused an increase equal to that seen with DHT if the
prostate tissue was first loaded with SHBG, e.g., if RSHBG was occupied by
SHBG. Because estradiol-SHBG increases intracellular cAMP, we ascertained
whether other compounds that raise cAMP (forskolin), or cAMP itself, could
increase PSA secretion. Such was the case. cAMP begins its signal cascade
by activating protein kinase A (PKA) so that if estradiol-SHBG increases PSA
secretion by a mechanism involving cAMP, inhibition of PKA should block
estradiol-SHBG-initiated PSA secretion. Estradiol-SHBG failed to stimulate
PSA when PKA was inhibited with PKI. On the other hand, DHT-stimulated
PSA secretion, which does not involve PKA, was not inhibited by PKI. That
the effect of estradiol-SHBG was independent of the estrogen receptor was
shown by the lack of inhibition of estrogen-stimulated PSA secretion by two
anti-estrogens, tamoxifen and ICI 164,284. The promoter of the PSA gene
has an androgen response element, and both PSA secretion and the
expression of PSA mRNA are androgen-regulated. We investigated the
effect of hydroxyflutamide and cyproterone acetate. Both potent anti-
androgens, on the E2-SHBG-mediated increase in PSA secretion. secretion.
They also blocked the effect of E2-SHBG on PSA secretion. Since E2 is
not exerting its effect by binding to the AR, e.g., it is not its cognate ligand,
the E2-induced secretion of PSA observed in this study reflects ligand-
independent activation of the AR.26 Thus, estradiol activates a typical
AR-mediated event, PSA synthesis and secretion, by activating SHBG-
RSHBG. These data make clear the fact that there is cross-talk between a
steroid hormone-engendered event at the cell membrane and a classic
intracellular steroid hormone receptor.
Abbreviations: PSA, prostate specific antigen; DHT, dihydrotestosterone;
E2, estradiol; PKI, inhibitor of protein kinase A; ICI 164,384 (a pure anti-
estrogen); 2MeOE2, 2 methoxyestradiol; Cypro, cyproterone acetate,
OHFlut, hydroxyflutamide.
Role of G protein-coupled estrogen receptor 1, GPER, in inhibition of oocyte
maturation by endogenous estrogens in zebrafish
Yefei Pang, Peter Thomas
Developmental Biology 342 (2010) 194–206
http://dx.doi.org:/10.1016/j.ydbio.2010.03.027
Estrogen inhibition of oocyte maturation (OM) and the role of GPER (formerly
known as GPR30) were investigated in zebrafish. Estradiol-17β (E2) and G-1,
a GPER-selective agonist, bound to zebrafish oocyte membranes suggesting
the presence of GPER which was confirmed by immunocytochemistry using
a specific GPER antibody. Incubation of follicle-enclosed oocytes with an
aromatase inhibitor, ATD, and enzymatic and manual removal of the ovarian
follicle cell layers significantly increased spontaneous OM which was partially
reversed by co-treatment with either 100 nM E2 or G-1. Incubation of
denuded oocytes with the GPER antibody blocked the inhibitory effects of
estrogens on OM, whereas microinjection of estrogen receptor alpha (ERα)
antisense oligonucleotides into the oocytes was ineffective. The results
suggest that endogenous estrogens produced by the follicle cells inhibit or
delay spontaneous maturation of zebrafish oocytes and that this estrogen
action is mediated through GPER. Treatment with E2 and G-1 also attenuated
the stimulatory effect of the teleost maturation-inducing steroid, 17,20 β-
dihyroxy-4-pregnen-3-one (DHP), on OM. Moreover, E2 and G-1 down-
regulated the expression of membrane progestin receptor alpha (mPRα),
the intermediary in DHP induction of OM. Conversely DHP treatment caused
a N50% decline in GPER mRNA levels. The results suggest that estrogens
and GPER are critical components of the endocrine system controlling
the onset of OM in zebrafish. A model is proposed for the dual control of the
onset of oocyte maturation in teleosts by estrogens and progestins acting
through GPER and mPRα, respectively, at different stages of oocyte
development.
Reprint of ’’GPR30 mediates estrogen rapid signaling and neuroprotection’’
Hui Tang, Q Zhang, L Yang, Y Dong, M Khan, F Yang, DW Brann, R Wang
Molecular and Cellular Endocrinology 389 (2014) 92–98
http://dx.doi.org/10.1016/j.mce.2014.01.024
http://dx.doi.org/10.1016/j.mce.2014.05.005
G-protein-coupled estrogen receptor-30 (GPR30), also known as G-protein
estrogen receptor-1 (GPER1), is a putative extranuclear estrogen receptor
whose precise functions in the brain are poorly understood. Studies using
exogenous administration of the GPR30 agonist, G1 suggests that GPR30
may have a neuroprotective role in cerebral ischemia. However, the
physiological role of GPR30 in mediating estrogen (E2)-induced neuro-
protection in cerebral ischemia remains unclear. Also unclear is whether
GPR30 has a role in mediating rapid signaling by E2 after cerebral ischemia,
which is thought to underlie its neuroprotective actions. To address these
deficits in our knowledge, the current study examined the effect of antisense
oligonucleotide (AS) knockdown of GPR30 in the hippocampal CA1 region
upon E2-BSA induced neuroprotection and rapid kinase signaling in a rat
model of global cerebral ischemia (GCI). Immunohistochemistry demonstrated
that GPR30 is strongly expressed in the hippocampal CA1 region and
dentate gyrus, with less expression in the CA3 region. E2-BSA exerted
robust neuroprotection of hippocampal CA1 neurons against GCI, an effect
abrogated by AS knockdown of GPR30. Missense control oligonucleotides had
no effect upon E2-BSA-induced neuroprotection, indicating specificity of the
effect. The GPR30 agonist, G1 also exerted significant neuroprotection against
GCI. E2-BSA and G1 also rapidly enhanced activation of the prosurvival
kinases, Akt and ERK, while decreasing proapototic JNK activation. Importantly,
AS knockdown of GPR30 markedly attenuated these rapid kinase signaling
effects of E2-BSA. As a whole, the studies provide evidence of an important
role of GPR30 in mediating the rapid signaling and neuroprotective actions
of E2 in the hippocampus.
Regulation of brain microglia by female gonadal steroids
Pardes Habib, Cordian Beyer
Journal of Steroid Biochemistry & Molecular Biology 2015; 146: 3–14
http://dx.doi.org/10.1016/j.jsbmb.2014.02.018
Microglial cells are the primary mediators of the CNS immune defense system
and crucial for shaping inflammatory responses. They represent a highly
dynamic cell population which is constantly moving and surveying their
environment. Acute brain damage causes a local attraction and activation of
this immune cell type which involves neuron-to-glia and glia-to-glia interactions.
The prevailing view attributes microglia a “negative” role such as defense and
debris elimination. More topical studies also suggest a protective and “positive”
regulatory function. Estrogens and progestins exert anti-inflammatory and
neuroprotective effects in the CNS in acute and chronic brain diseases.
Recent work revealed that microglial cells express subsets of classical and
non-classical estrogen and progesterone receptors in a highly dynamic way.
In this review article, we would like to stress the importance of microglia for
the spreading of neural damage during hypoxia, their susceptibility to functional
modulation by sex steroids, the potency of sex hormones to switch microglia
from a pro-inflammatory M1 to neuroprotective M2 phenotype, and the
regulation of pro-and anti-inflammatory properties including the inflammasome.
We will further discuss the possibility that the neuroprotective action of sex
steroids in the brain involves an early and direct modulation of local microglia
cell function. Neuroprotection by gonadal steroid hormones in acute brain
damage requires cooperation with astroglia and microglia
Sonja Johann, Cordian Beyer
http://dx.doi.org/10.1016/j.jsbmb.2012.11.006
The neuroactive steroids 17β-estradiol and progesterone control a broad
spectrum of neural functions. Besides their roles in the regulation of classical
neuroendocrine loops, they strongly influence motor and cognitive systems,
behavior, and modulate brain performance at almost every level. Such a
statement is underpinned by the widespread and lifelong expression pattern
of all types of classical and non-classical estrogen and progesterone receptors
in the CNS. The life-sustaining power of neurosteroids for tattered or seriously
damaged neurons aroused interest in the scientific community in the past years
to study their ability for therapeutic use under neuropathological challenges.
Documented by excellent studies either performed in vitro or in adequate animal
models mimicking acute toxic or chronic neuro-degenerative brain disorders,
both hormones revealed a high potency to protect neurons from damage
and saved neural systems from collapse. Unfortunately, neurons, astroglia,
microglia, and oligodendrocytes are comparably target cells for both steroid
hormones. This hampers the precise assignment and understanding of
neuroprotective cellular mechanisms activated by both steroids. In this article,
we strive for a better comprehension of the mutual reaction between these
steroid hormones and the two major glial cell types involved in the maintenance
of brain homeostasis, astroglia and microglia, during acute traumatic brain
injuries such as stroke and hypoxia. In particular, we attempt to summarize
steroid-activated cellular signaling pathways and molecular responses in these
cells and their contribution to dampening neuroinflammation and neural
destruction.
Photoperiod influences the ontogenetic expression of aromatase
and estrogen receptor α in the developing tilapia brain.
Li-Hsueh Wang, Ching-Lin Tsai
General and Comparative Endocrinology 2006; 145: 62–66
http://dx.doi.org:/10.1016/j.ygcen.2005.07.004
Neural development is determined not only by genetic regulation, but also
by environmental cues. Central estrogen-forming/estrogen-sensitive systems
play an important role in the neural development of the brain. In the present
study, the quantitative reverse transcription-polymerase chain reaction method
was used to investigate the effects of photoperiod on the ontogenetic
expression of aromatase and estrogen receptor a (ERα) in the developing
tilapia brain. Before day 5 post-hatch, brain aromatase mRNA expression was
significantly decreased by constant light but not influenced by constant darkness.
During this period, brain ERα mRNA expression was significantly increased
under both constant light and constant darkness. Between days 5 and 10, and
between days 10 and 15, neither brain aromatase nor brain ERα expression
was altered under constant darkness and constant light. These results indicate
that the ontogenetic expression of brain aromatase and brain ERα is not via a
light-inducing process but influenced by a light-entraining signal during the
very early period of development.
Orphanin FQ-ORL-1 Regulation of Reproduction and Reproductive Behavior in
the Female
Kevin Sinchak, Lauren Dalhousay, Nayna Sanathara
Vitamins and Hormones 187-220. http://dx.doi.org/10.1016/bs.vh.2014.11.002
Orphanin FQ (OFQ/N) and its receptor, opioid receptor-like receptor-1 (ORL-1),
are expressed throughout steroid-responsive limbic and hypothalamic circuits
that regulate female ovarian hormone feedback and reproductive behavior
circuits. The arcuate nucleus of the hypothalamus (ARH) is a brain region
that expresses OFQ/N and ORL-1 important for both sexual behavior and
modulating estradiol feedback loops. Within the ARH, the activation of the
OFQ/N-ORL-1 system facilitates sexual receptivity (lordosis) through the
inhibition of β-endorphin neuronal activity. Estradiol initially activates ARH
β-endorphin neurons to inhibit lordosis. Simultaneously, estradiol upregulates
coexpression of OFQ/N and progesterone receptors and ORL-1 in ARH
β-endorphin neurons. Ovarian hormones regulate pre- and postsynaptic
coupling of ORL-1 to its G protein-coupled signaling pathways. When the
steroid-primed rat is nonreceptive, estradiol acts pre- and postsynaptically
to decrease the ability of the OFQ/N-ORL-1 system to inhibit ARH β-endorphin
neurotransmission. Conversely, when sexually receptive, ORL-1 signaling is
restored to inhibit β-endorphin neurotransmission. Although steroid signaling
that facilitates lordosis converges to deactivate ARH.
Estradiol Activates the Prostate Androgen Receptor and Prostate specific Antigen
Secretion through the Intermediacy of Sex Hormone-binding Globulin
Atif M. Nakhla, Nicholas A. Romas, and William Rosner
J Biol Chem Mar 14, 1997; 272(11): 6838–6841 http://www-jbc.stanford.edu/jbc/
These experiments were designed to examine the relationship between the
effects of steroid hormones mediated by classic intracellular steroid hormone
receptors and those mediated by a signaling system subserved at the plasma
membrane by a receptor for sex hormone binding globulin. It is known that
unliganded sex hormone-binding globulin (SHBG) binds to a receptor (RSHBG)
on prostate membranes. The RSHBG*SHBG complex is rapidly activated by
estradiol to stimulate adenylate cyclase, with a resultant increase in intracellular
cAMP. In this paper we examine the effect of this system on a prostate gene
product known to be activated by androgens, prostate-specific antigen.
We have shown previously that estradiol (E2) participates in a signaling
system that originates, not within the cell, but at the plasma membrane.
Through the intermediacy of the plasma protein, sex hormone-binding
globulin (SHBG), it causes the generation of cAMP. In brief, unliganded
SHBG binds to a receptor (RSHBG) on certain cell surfaces and the
RSHBG*SHBG complex is rapidly activated by E2 to stimulate adenylate cyclase,
with a resultant increase in intracellular cAMP. There is a paucity of information
on events subsequent to the generation of cAMP by this system. In this paper
we examine the effect of E2-SHBG-RSHBG on an androgen responsive gene.
The gene for prostate-specific antigen (PSA) contains an androgen response
element. After binding its cognate ligand, the androgen receptor (AR) interacts
with this response element to initiate PSA mRNA transcription and secretion.
We show that, in the absence of androgens, E2 in concert with SHBG*RSHBG,
acts at the cell membrane to cause secretion of PSA and that this effect is
blocked by anti-androgens. This observation provides a first functional link
between a classic steroid hormone receptor and a cell membrane-mediated
steroidal effect. In serum-free organ culture of human prostates,
dihydrotestosterone caused an increase in prostate specific antigen secretion.
This event was blocked by the anti-androgens cyproterone acetate and
hydroxyflutamide. In the absence of androgens, estradiol added to prostate
tissue, whose RSHBG was occupied by SHBG, reproduced the results seen
with dihydrotestosterone. Neither estradiol alone nor SHBG alone duplicated
these effects. The estradiol*SHBG-induced increase in prostate-specific
antigen was not blocked by anti-estrogens, but was blocked both by anti-
androgens and a steroid (2-methoxyestradiol) that prevents the binding of
estradiol to SHBG. Furthermore, an inhibitor of protein kinase A prevented
the estradiol*SHBG-induced increase in prostate-specific antigen but not
that which followed dihydrotestosterone. These data indicate that there is a
signaling system that amalgamates steroid-initiated intracellular events
with steroid-dependent occurrences generated at the cell membrane and
that the latter signaling system proceeds by a pathway that involves protein
kinase A.
Mechanisms of crosstalk between endocrine systems: Regulation of sex steroid
hormone synthesis and action by thyroid hormones
Paula Duarte-Guterman, Laia Navarro-Martín, Vance L. Trudeau
General and Comparative Endocrinology 203 (2014) 69–85
http://dx.doi.org/10.1016/j.ygcen.2014.03.015
Thyroid hormones (THs) are well-known regulators of development and
metabolism in vertebrates. There is increasing evidence that THs are also
involved in gonadal differentiation and reproductive function. Changes in TH
status affect sex ratios in developing fish and frogs and reproduction
(e.g., fertility), hormone levels, and gonad morphology in adults of species of
different vertebrates. In this review, we have summarized and compared the
evidence for cross-talk between the steroid hormone and thyroid axes and
present a comparative model. We gave special attention to TH regulation of
sex steroid synthesis and action in both the brain and gonad, since these are
important for gonad development and brain sexual differentiation and have
been studied in many species. We also reviewed research showing that
there is a TH system, including receptors and enzymes, in the brains and
gonads in developing and adult vertebrates. Our analysis shows that THs
influences sex steroid hormone synthesis in vertebrates, ranging from fish
to pigs. This concept of crosstalk and conserved hormone interaction has
implications for our understanding of the role of THs in reproduction, and
how these processes may be dysregulated by environmental endocrine
disruptors.
Inverse relationship between hSHBG affinity for testosterone and hSHBG
concentration revealed by surface plasmon resonance
Laurence Heinrich-Balard, Wael Zeinyeh, Henri Déchaud, Pascaline Rivory, et al.
Molecular and Cellular Endocrinology 399 (2015) 201–207
http://dx.doi.org/10.1016/j.mce.2014.10.002
A wide range of human sex hormone-binding globulin (hSHBG) affinity constants
for testosterone (KA_hSHBG) has been reported in literature. To bring new insight
on the KA_hSHBG value, we implemented a study of the molecular interactions
occurring between testosterone and its plasma transport proteins by using
surface plasmon resonance. The immobilization on the sensor-chip of a
testosterone derivative was performed by an oligoethylene glycol linker.
For different plasmas with hSHBG concentrations, an assessment of the
KA_hSHBG was obtained from a set of sensor-grams and curve-fitting these
data.We observed that KA_hSHBG decreased, from at least two decades,
when the plasma hSHBG concentration increased from 4.4 to 680 nmol/L.
Our study shows a wide biological variability of KA_hSHBG that is related
to the hSHBG concentration.
These unexpected results may have a physiological significance and question
the validity of current methods that are recommended for calculating free
testosterone concentrations to evaluate androgen disorders in humans.
Intracrinology in action: Importance of extragonadal sex steroid biosynthesis
and inactivation in peripheral tissues in both women and men.
Editorial
Journal of Steroid Biochemistry & Molecular Biology 145 (2015) 131–132
http://dx.doi.org/10.1016/j.jsbmb.2014.09.012
It seems appropriate, as introduction, to summarize the mechanisms at the
basis of the new paradigm of steroid biosynthesis in the human peripheral
tissues, namely intracrinology. While the first clinical proof of the role of
extragonadal sex steroid biosynthesis was obtained with combined androgen
blockade in men treated for prostate cancer, the first demonstration of the
efficacy of DHEA replacement therapy was on the symptoms of vulvovaginal
atrophy in postmenopausal women; (Archer, this issue).
DHEA is transformed specifically in each cell of each peripheral tissue into
the proper amounts of estrogens and/or androgens, depending upon the
local expression of the appropriate steroid forming enzymes; (Labrie, this issue).
Most importantly, the sex steroids synthesized and acting intracellularly in
peripheral tissues are also inactivated locally before being released in the
extracellular space, thus maintaining the serum levels of estradiol and
testosterone at biologically inactive concentrations, thus avoiding systemic
exposure to sex steroids during menopause as well illustrated by atrophy
of the endometrium.
As mentioned above, that extragonadal androgen biosynthesis is clinically
important became obvious in 1982 when the addition of the antiandrogen
flutamide to castration provided very exciting and unexpected beneficial results
(Labrie, this issue). In fact, combining a pure anti-androgen to castration has
been the first treatment shown to prolong life in prostate cancer and very clearly
confirmed by the prolongation of life of 2.2–4.8 months observed following
addition of MDV-3100 or abiraterone to castration resistant prostate cancer
patients (Grist et al., this issue). (Mizokami et al., this issue) very competently
complement the mechanisms potentially involved in extragonadal steroid
biosynthesis. A repeated observation is the association between serum DHEA
levels and increased longevity, a subject reviewed by Ohlsson et al., this issue.
Most importantly, a subject which remains to be supported by long-term clinical
trials but which shows very promising preclinical data is the possibility of a
beneficial effect of DHEA on brain functions, especially cognition, memory
and delayed development of mild cognitive impairment and Alzheimer’s
disease (see Starka et al.; Soma et al; Pluchino et al; Maggio et al.; Hill et al.,
this issue). The information summarized in the very up-to-date manuscripts
of this special JSBMB issue has the potential of opening the way to a prodrug
replacement therapy already well illustrated on the symptoms and signs of
vulvovaginal atrophy and sexual dysfunction (Archer, this issue). The
administration to sex steroid deficient women of an appropriate amount of
DHEA able to correct the symptoms of vulvovaginal atrophy (mostly estrogen-
sensitive) and sexual dysfunction (androgen-sensitive), and potentially, in the
future, other problems of menopause, does permit to the sex steroid-deficient
women to benefit from a normal/sufficient level of sex steroids in specific tissues
using the enzymes developed over 500 million years to permit a better quality
of life during the menopausal years.
Inactivation of androgens by UDP-glucuronosyltransferase enzymes in humans
Alain Belanger, Georges Pelletier, Fernand Labrie, Olivier Barbier and Sarah Chouinard
TRENDS in Endocrinology and Metabolism 2003; 14(10):473-78
http://dx.doi.org:/10.1016/j.tem.2003.10.005
In humans, 3b-hydroxysteroid dehydrogenase (3β-HSD), 17β-HSD and
5α-reductase activities in androgen target tissues, such as the prostate and
skin, convert dehydroepiandrosterone, androstenedione and testosterone into
the most potent natural androgen dihydrotestosterone (DHT). This androgen
is converted mainly in situ into two phase I metabolites, androsterone (ADT)
and androstane-3α,17β-diol (3α-DIOL), which might be back converted to DHT.
Here, we discuss the recent findings regarding the characterization of specific
UDP glucuronosyltransferases (UGTs), UGT2B7, B15 and B17, responsible for
the glucuronidation of these metabolites. The tissue distribution and cellular
localization of the UGT2B transcripts and proteins in humans clearly indicate
that these enzymes are synthesized in androgen-sensitive tissues. It is
postulated that the conjugating activity of UGT enzymes is the main mechanism
for modulating the action of steroids and protecting the androgen-sensitive
tissues from deleteriously high concentrations of DHT, ADT and 3α-DIOL.
Synthesis and Evaluation of Potential Radioligands for the Progesterone Receptor
R.M. Hoyte, W. Rosner, I.S. Johnson, J. Zielinski, and R. B. Hochberg
J. Med. Chem. 1985; 28: 1695-1699
Several steroidal analogues were synthesized as potential y-emitting radioligands
for the progesterone receptor. Each of these compounds was tested as an inhibitor
of the specific binding of [3H]-17α,21-dimethyl-19-nor-4,9-pregnadiene-3,20-dione
(R5020) to the progesterone receptor in rabbit uterine cytosol. R5020 is a well-
known progestin with high affinity for the receptor. Of the compounds synthesized,
aromatic N-substituted (2-17 steroidal carboxamides inhibited the binding only
poorly. Three compounds, 16α-iodo-4-estren-17β-ol-3-one, 17α-[2(E)-iodovinyl]
-4-estren-17β-ol-3-one, and 17α-[2(Z)-iodovinyl]-4-estren-l7β-ol-3-one are
excellent competitors, each having a Ki less than or equal to that of the natural
progestin, progesterone. Since similar iodinated analogues of estrogens
have been shown to be extremely stable both in vivo and in vitro, these compounds
are potentially useful ligands for the progesterone receptor.
Estradiol concentration and the expression of estrogen receptors in the testes of
the domestic goose (Anser anser f. domestica) during the annual reproductive cycle
Leska, J. Kiezun, B. Kaminska, L. Dusza
Domestic Animal Endocrinology 51 (2015) 96–104
http://dx.doi.org/10.1016/j.domaniend.2014.12.002
Seasonal fluctuations in the activity of bird testes are regulated by a complex mechanism
where androgens play a key role. Until recently, the role played by estrogens in males has
been significantly underestimated. However, there is growing evidence that the proper
functioning of the testes is associated with optimal estradiol (E2) concentration
in both the plasma and testes of many mammalian species. Estrogens are
gradually emerging as very important players in hormonal regulation of
reproductive processes in male mammals. Despite the previously mentioned,
it should be noted that estrogenic action is limited by the availability of
specific receptors – estrogen receptor alpha (ERα) and estrogen receptor beta
(ERβ). Interestingly, there is a general scarcity of information concerning the
estrogen responsive system in the testes of male birds, which is of particular
interest in exploring the phenomenon of seasonality of reproduction. To address
this question, we have investigated for the first time the simultaneous
expression of testicular ERα and ERβ genes and proteins with the
accompanying plasma and testicular E2 concentrations during the annual
reproductive cycle of male bird. The research model was the domestic
goose (Anser anser f. domestica), a species whose annual reproductive
cycle can be divided into 3 distinct phases characterized by changes
in testicular activity. It has been revealed that the stable plasma E2 profile
did not correspond to changing intratesticular E2 profile throughout the
experiment. The expression of ERα and ERβ genes and proteins was detected
in gander testes and it fluctuated on a seasonal basis with lower level in
breeding and sexual reactivation stages and higher level during the
nonbreeding stage. Our results demonstrated changes in testicular sensitivity
to estrogens in male domestic goose during the annual reproductive cycle.
The seasonal pattern of estrogen receptors (ERs) expression was analyzed
against the hormonal background and a potential mechanism of ERs regulation
in bird testes was proposed. The present study revealed seasonal variations
in the estrogen responsive system, but further research is needed to fully
explore the role of estrogens in the reproductive tract of male birds.
Effects of 5α-dihydrotestosterone on expression of genes related to steroidogenesis
and spermatogenesis during the sex determination and differentiation periods of
the pejerrey, Odontesthes bonariensis
Anelisa González, Juan I. Fernandino, Gustavo M. Somoza
Comparative Biochemistry and Physiology, Part A 182 (2015) 1–7
http://dx.doi.org/10.1016/j.cbpa.2014.12.003
Sex steroid hormones are important players in the control of sex differentiation
by regulating gonadal development in teleosts. Although estrogens are clearly
associated with the ovarian differentiation in teleosts, the effects of androgens
on early gonadal development are still a matter of debate. Traditionally,
11-ketotestosterone (11-KT) is considered themajor androgen in fish; however,
5α-dihydrotestosterone (5α-DHT), the most potent androgen in tetrapods, was
recently found in fish testis and plasma, but its physiological role is still unknown.
In this context, the expression of genes associated with steroidogenesis and
spermatogenesis, body growth and sex differentiation were assessed in
Odontesthes bonariensis larvae fed with food supplemented with two doses of
5α-DHT (0.1 and 10 μg/g of food) from hatching to 6 weeks of age. At the lowest
dose, 5α-DHT treated larvae showed an estrogenic gene expression pattern, with
low hsd11β2 and high cyp19α1α and er2 expression levels with no differences
in sex ratio. At the highest dose, 5α-DHT produced a male-shifted sex ratio and
the larvae exhibited a gene expression profile characteristic of an advancement
of spermatogenesis, with inhibition of amh and stimulation of ndrg3. No
differences were observed in somatic growth. These results suggest that in
this species, 5α-DHT could have a role on sex differentiation and its effects
can differ according to the dose.
Do androgens link morphology and behavior to produce phenotype-specific
behavioral strategies?
Douglas G. Barron, Michael S. Webster, Hubert Schwabl
Animal Behaviour 100 (2015) 116e124
http://dx.doi.org/10.1016/j.anbehav.2014.11.016
Morphological and behavioral traits often covary with each other, and the links
between them may arise from shared physiological mechanisms. In particular,
androgens such as testosterone have emerged as prime candidates for linking
behaviour and morphology due to the environmental sensitivity and pleiotropic
effects of these hormones. In this study we investigated the hypothesis that
androgens simultaneously relate to morphological and behavioral variation,
thereby producing the integrated reproductive phenotypes of male red-backed
fairy-wrens, Malurus melanocephalus. Males of this species can adopt one of
three discrete breeding phenotypes: breeding in red/black plumage, breeding
in brown plumage, or remaining as nonbreeding brown natal auxiliaries. Although
the expression of morphological traits in this species is regulated by androgens
and phenotypes differ in baseline androgen levels (red/black breeder > brown
breeder > auxiliary), injection with GnRH failed to expose phenotype specific
constraints on androgen production. Observations of territoriality, nestling
feeding and extraterritorial forays revealed phenotype-specific patterns of mating
and parental effort, yet these were largely related to age and were not correlated
with baseline or GnRH-induced androgen levels, or the androgen change between
these points. While these findings support the idea that morphological and
behavioral traits are linked via phenotypic correlations, they do not support
the hypothesis that behavioral differences arise from variation in circulating
androgens or the capacity to produce them.
Effects of sex steroids on expression of genes regulating growth-related
mechanisms in rainbow trout (Oncorhynchus mykiss)
Beth M. Cleveland, Gregory M. Weber
General and Comparative Endocrinology xxx (2015) xxx–xxx
http://dx.doi.org/10.1016/j.ygcen.2014.11.018
Effects of a single injection of 17b-estradiol (E2), testosterone (T), or
5b-dihydrotestosterone (DHT) on expression of genes central to the
growth hormone (GH)/insulin-like growth factor (IGF) axis, muscle
regulatory factors, transforming growth factor-beta (TGFβ) superfamily
signaling cascade, and estrogen receptors were determined in rainbow
trout (Oncorhynchus mykiss) liver and white muscle tissue. In liver in
addition to regulating GH sensitivity and IGF production, sex
steroids also affected expression of IGF binding proteins, as E2, T,
and DHT increased expression of igfbp2β and E2 also increased
expression of igfbp2 and igfbp4. Regulation of this system also occurred
in white muscle in which E2 increased expression of igf1, igf2, and
igfbp5β1, suggesting anabolic capacity may be maintained in white
muscle in the presence of E2. In contrast, DHT decreased expression
of igfbp5β1. DHT and T decreased expression of myogenin, while other
muscle regulatory factors were either not affected or responded similarly
for all steroid treatments. Genes within the TGFβ superfamily signaling
cascade responded to steroid treatment in both liver and muscle,
suggesting a regulatory role for sex steroids in the ability to transmit
signals initiated by TGFβ superfamily ligands, with a greater number
of genes responding in liver than in muscle. Estrogen receptors were
also regulated by sex steroids, with era1 expression increasing for all
treatments in muscle, but only E2- and T-treatment in liver. E2 reduced
expression of erb2 in liver. Collectively, these data identify how
physiological mechanisms are regulated by sex steroids in a manner
that promotes the disparate effects of androgens and estrogens on
growth in salmonids.
Distribution and function of 3′,5′-Cyclic-AMP phosphodiesterases in the human ovary
T.S. Petersen, S.G. Kristensen, J.V. Jeppesen, .., K.T. Macklon, C.Y. Andersen
Molecular and Cellular Endocrinology 403 (2015) 10–20
http://dx.doi.org/10.1016/j.mce.2015.01.004
The concentration of the important second messenger cAMP is regulated by
phosphodiesterases (PDEs) and hence an attractive drug target. However,
limited human data are available about the PDEs in the ovary. The aim of the
present study was to describe and characterise the PDEs in the human ovary.
Results were obtained by analysis of mRNA microarray data from follicles and
granulosa cells (GCs), combined RT-PCR and enzymatic activity analysis in GCs,
immunohisto-chemical analysis of ovarian sections and by studying the effect
of PDE inhibitors on progesterone production from cultured GCs. We found that
PDE3, PDE4, PDE7 and PDE8 are the major families present while PDE11A
was not detected. PDE8B was differentially expressed during folliculogenesis.
In cultured GCs, inhibition of PDE7 and PDE8 increased basal progesterone
secretion while PDE4 inhibition increased forskolin-stimulated progesterone
secretion. In conclusion, we identified PDE3, PDE4, PDE7 and PDE8 as
the major PDEs in the human ovary.
Diethylstilbestrol can effectively accelerate estradiol-17-O-glucuronidation, while
potently inhibiting estradiol-3-O-glucuronidation
Liangliang Zhu, Ling Xiao, Yangliu Xia, .., Yan Wu, Ganlin Wu, Ling Yang
Toxicology and Applied Pharmacology 283 (2015) 109–116
http://dx.doi.org/10.1016/j.taap.2015.01.003
This in vitro study investigates the effects of diethylstilbestrol (DES), a widely
used toxic synthetic estrogen, on estradiol-3- and 17-O- (E2-3/17-O)
glucuronidation, via culturing human liver microsomes (HLMs) or
recombinant UDP-glucuronosyl-transferases (UGTs) with DES and E2.
DES can potently inhibit E2-3-O-glucuronid-ation in HLM, a probe reaction
for UGT1A1. Kinetic assays indicate that the inhibition follows a competitive
inhibition mechanism, with the Ki value of 2.1 ± 0.3 μM, which is less than
the possible in vivo level. In contrast to the inhibition on E2-3-O-glucuronidation,
the acceleration is observed on E2-17-O-glucuronidation in HLM, in which
cholestatic E2-17-O-glucuronide is generated. In the presence of DES
(0–6.25 μM), Km values for E2-17-Oglucuronidation are located in the
range of 7.2–7.4 μM, while Vmax values range from 0.38 to 1.54 nmol/min/mg.
The mechanism behind the activation in HLM is further demonstrated by
the fact that DES can efficiently elevate the activity of UGT1A4 in catalyzing
E2-17-O-glucuronidation. The presence of DES (2 μM) can elevate Vmax from
0.016 to 0.81 nmol/min/mg, while lifting Km in a much lesser extent from 4.4 to
11 μM. Activation of E2-17-O-glucuronidation is well described by a two binding
site model, with KA, α, and β values of 0.077 ± 0.18 μM, 3.3 ± 1.1 and 104 ± 56,
respectively. However, diverse effects of DES towards E2-3/17-O-glucuronidation
are not observed in liver microsomes from several common experimental animals.
In summary, this study issues new potential toxic mechanisms for DES: potently
inhibiting the activity of UGT1A1 and powerfully accelerating the formation of
cholestatic E2-17-O-glucuronide by UGT1A4.
Dehydroepiandrosterone: A neuroactive steroid
Luboslav Stárka, Michaela Dusková, Martin Hill
Journal of Steroid Biochemistry & Molecular Biology 145 (2015) 254–260
http://dx.doi.org/10.1016/j.jsbmb.2014.03.008
Dehydroepiandrosterone (DHEA) and its sulfate bound form (DHEAS) are important
steroids of mainly adrenal origin. They are produced also in gonads and in the brain.
Dehydroepiandrosterone easily crosses the brain–blood barrier and in part is also
produced locally in the brain tissue. In the brain, DHEA exerts its effects after
conversion to either testosterone and dihydrotestosterone or estradiol via androgen
and estrogen receptors present in the most parts of the human brain, through
mainly non-genomic mechanisms, or eventually indirectly via the effects of its
metabolites formed locally in the brain. As a neuroactive hormone, DHEA in
cooperation with other hormones and transmitters significantly affects some
aspects of human mood, and modifies some features of human emotions and
behavior. It has been reported that its administration can increase feelings of well-
being and is useful in ameliorating atypical depressive disorders. It has
neuroprotective and antiglucocorticoid activity and modifies immune reactions,
and some authors have also reported its role in degenerative brain diseases.
Here we present a short overview of the possible actions of dehydroepiandrosterone
and its sulfate in the brain, calling attention to various mechanisms of their action
as neurosteroids and to prospects for the knowledge of their role in brain disorders.
Androgens and mammalian male reproductive tract development
Aki Murashima, Satoshi Kishigami, Axel Thomson, Gen Yamada
Biochimica et Biophysica Acta 1849 (2015) 163–170
http://dx.doi.org/10.1016/j.bbagrm.2014.05.020
One of the main functions of androgen is in the sexually dimorphic development of
the male reproductive tissues. During embryogenesis, androgen determines the
morphogenesis of male specific organs, such as the epididymis, seminal vesicle,
prostate and penis. Despite the critical function of androgens in masculinization,
the downstream molecular mechanisms of androgen signaling are poorly
understood. Tissue recombination experiments and tissue specific androgen
receptor (AR) knockout mouse studies have revealed epithelial or mesenchymal
specific androgen-AR signaling functions. These findings also indicate that
epithelial–mesenchymal interactions are a key feature of AR specific activity,
and paracrine growth factor action may mediate some of the effects of androgens.
This review focuses on mouse models showing the interactions of androgen and
growth factor pathways that promote the sexual differentiation of reproductive organs.
Recent studies investigating context dependent AR target genes are also discussed.
This article is part of a Special Issue entitled: Nuclear receptors in animal development.
All sex steroids are made intracellularly in peripheral tissues by the mechanisms of
intracrinology after menopause
Fernand Labrie
Journal of Steroid Biochemistry & Molecular Biology 145 (2015) 133–138
http://dx.doi.org/10.1016/j.jsbmb.2014.06.001
Following the arrest of estradiol secretion by the ovaries at menopause, all estrogens
and all androgens in postmenopausal women are made locally in peripheral target
tissues according to the physiological mechanisms of intracrinology. The locally
made sex steroids exert their action and are inactivated intracellularly without
biologically significant release of the active sex steroids in the circulation.The
level of expression of the steroid-forming and steroid-inactivating enzymes is
specific to each cell type in each tissue, thus permitting to each cell/tissue to
synthesize a small amount of androgens and/or estrogens in order to meet the
local physiological needs without affecting the other tissues of the organism.
Achieved after 500 million years of evolution, combination of the arrest of ovarian
estrogen secretion, the availability of high circulating levels of DHEA and the
expression of the peripheral sex steroid-forming enzymes have permitted the
appearance of menopause with a continuing access to intra-tissular sex steroids
for the individual cells/tissues without systemic exposure to circulating estradiol.
In fact, one essential condition of menopause is to maintain serum estradiol at
biologically inactive (subthreshold) concentrations, thus avoiding stimulation of the
endometrium and risk of endometrial cancer. Measurement of the low levels of
serum estrogens and androgens in postmenopausal women absolutely requires
the use of MS/MS-based technology in order to obtain reliable accurate, specific
and precise assays. While the activity of the series of steroidogenic enzymes can
vary, the serum levels of DHEA show large individual variations going from barely
detectable to practically normal “premenopausal” values, thus explaining the
absence of menopausal symptoms in about 25% of women. It should be added
that the intracrine system has no feedback elements to adjust the serum levels
of DHEA, thus meaning that women with low DHEA activity will not be improved
without external supplementation. Exogenous DHEA, however, follows the same
intracrine rules as described for endogenous DHEA, thus maintaining serum
estrogen levels at subthreshold or biologically inactive concentrations. Such blood
concentrations are not different from those observed in normal postmenopausal
women having high serum DHEA concentrations. Androgens, on the other hand,
are practically all made intracellularly from DHEA by the mechanisms of intracrinology
and are always maintained at very low levels in the blood in both pre- and
postmenopausal women. Proof of the importance of intracrinology is also provided,
among others, by the well-recognized benefits of aromatase inhibitors and
anti-estrogens used successfully for the treatment of breast cancer in
postmenopausal women where all estrogens are made locally. Each medical
indication for the use of DHEA, however, requires clinical trials performed
according to the FDA guidelines and the best rules of clinical medicine.
A multi-step, dynamic allosteric model of testosterone’s binding to sex hormone
binding globulin
Mikhail N. Zakharov, Shalender Bhasin, Thomas G. Travison, Ran Xue, et al.
Molecular and Cellular Endocrinology 399 (2015) 190–200
http://dx.doi.org/10.1016/j.mce.2014.09.001
Purpose: Circulating free testosterone (FT) levels have been used widely in the
diagnosis and treatment of hypogonadism in men. Due to experimental
complexities in FT measurements, the Endocrine Society has recommended
the use of calculated FT (cFT) as an appropriate approach for estimating FT.
We show here that the prevailing model of testosterone’s binding to SHBG,
which assumes that each SHBG dimer binds two testosterone molecules
and that the two binding sites on SHBG have similar binding affinity is
erroneous and provides FT values that differ substantially from those
obtained using equilibrium dialysis.
Methods: We characterized testosterone’s binding to SHBG using
binding isotherms, ligand depletion curves, and isothermal titration
calorimetry (ITC). We derived a new model of testosterone’s binding to
SHBG from these experimental data and used this model to determine
FT concentrations and compare these values with those derived from
equilibrium dialysis.
Results: Experimental data on testosterone’s association with SHBG
generated using binding isotherms including equilibrium binding, ligand
depletion experiments, and ITC provide evidence of a multi-step dynamic
process, encompassing at least two inter-converting microstates in unliganded
SHBG, readjustment of equilibria between unliganded states upon binding
of the first ligand molecule, and allosteric interaction between two binding
sites of SHBG dimer. FT concentrations in men determined using the new
multistep dynamic model with complex allostery did not differ from those
measured using equilibrium dialysis. Systematic error in calculated FT
vales in females using Vermeulen’s model was also significantly reduced.
In European Male Aging Study, the men deemed to have low FT (<2.5th
percentile) by the new model were at increased risk of sexual symptoms
and elevated LH.
Conclusion: Testosterone’s binding to SHBG is a multi-step dynamic
process that involves complex allostery within SHBG dimer. FT values
obtained using the new model have close correspondence with those
measured using equilibrium dialysis.
Cohesin modulates transcription of estrogen-responsive genes
Jisha Antony, Tanushree Dasgupta, Jenny M. Rhodes, Miranda V. McEwan, et al.
Biochimica et Biophysica Acta 1849 (2015) 257–269
http://dx.doi.org/10.1016/j.bbagrm.2014.12.011
The cohesin complex has essential roles in cell division, DNA damage repair
and gene transcription. The transcriptional function of cohesin is thought to
derive from its ability to connect distant regulatory elements with gene promoters.
Genome-wide binding of cohesin in breast cancer cells frequently coincides
with estrogen receptor alpha (ERα), leading to the hypothesis that cohesin
facilitates estrogen-dependent gene transcription. We found that cohesin
modulates the expression of only a subset of genes in the ER transcription
program, either activating or repressing transcription depending on the gene
target. Estrogen-responsive genes most significantly influenced by cohesin
were enriched in pathways associated with breast cancer progression such
as PI3K and ErbB1. In MCF7 breast cancer cells, cohesin depletion enhanced
transcription of TFF1 and TFF2, and was associated with increased ER binding
and increased interaction between TFF1 and its distal enhancer situated
within TMPRSS3. In contrast, cohesin depletion reduced c-MYC mRNA and
was accompanied by reduced interaction between a distal enhancer of c-MYC
and its promoters. Our data indicates that cohesin is not a universal facilitator
of ER-induced transcription and can even restrict enhancer–promoter communication.
We propose that cohesion modulates transcription of estrogen-dependent genes
to achieve appropriate directionality and amplitude of expression.
Angiogenesis in Breast Cancer and its Correlation with Estrogen, Progesterone
Receptors and other Prognostic Factors
Jyotsna Naresh Bharti, Poonam Rani, Vinay Kamal, Prem Narayan Agarwal
Journal of Clinical and Diagnostic Research. 2015 Jan, Vol-9(1): EC05-EC07
http://dx.doi.org:/10.7860/JCDR/2015/10591.5447
Purpose: The aim of study is to evaluate angiogenesis using CD34, in
estrogen, progesterone positive and negative breast cancer and to correlate
the microvessel density with known histological prognostic factors,
morphological type of breast carcinoma and lymph node metastasis.
Materials and Methods: Twenty eight untreated cases of breast cancer were
included in the study and paraffin embedded sections were obtained
from representative mastectomy specimen of breast cancer patient. The sections
were stained with hematoxylin and eosin stain and immunohistochemistry was
performed using CD34, estrogen, progesterone, cytokeratin and epithelial
membrane antigen antibody. Angiogenesis was analyzed using CD 34 antibody.
For statistical analysis, cases were grouped into estrogen, progesterone positive
and negative receptors.
Results: Mean microvessel density in ER-/PR-, ER-/ PR+, ER+/PR-, ER+/PR+
was 15.45, 14.83, 11, 10.89 respectively. A significant correlation was found
between ER receptors and mean vascular density with p-value (< 0.05).
A significant difference was observed in mean vascular density between
the four groups comprising (p-value < 0.05). Infiltrating duct carcinoma
(NOS) grade III has got the highest mean microvessel density (14.17)
followed by grade II (12.93) and grade I (12.33).
Conclusion: Information about prognostic factors in breast cancer
patients may lead to better ways to identify those patients at high risk
who might benefit from adjuvant therapies.
Combined blockade of testicular and locally made androgens in prostate cancer:
A highly significant medical progress based upon intracrinology
Fernand Labrie
Journal of Steroid Biochemistry & Molecular Biology 145 (2015) 144–156
http://dx.doi.org/10.1016/j.jsbmb.2014.05.012
Recently two drugs, namely the antiandrogen MDV-3100 and the inhibitor
of 17β-hydroxylase abiraterone have been accepted by the FDA for the
treatment of castration-resistant prostate cancer (CRPC) with or without
previous chemotherapy, with a prolongation of overall survival of 2.2–4.8months.
While medical (GnRH agonist) or surgical castration reduces the serum levels
of testosterone by about 97%, an important concentration of testosterone and
dihydrotestosterone remains in the prostate and activates the androgen receptor
(AR), thus offering an explanation for the positive data obtained in CRPC. In fact,
explanation of the response observed with MDV-3100 or enzalutamide in CRPC
is essentially a blockade of the action or formation of intraprostatic androgens.
In addition to the inhibition of the action or formation of androgens made locally
by the mechanisms of intracrinology, increased AR levels and AR mutations can
be involved, especially in very advanced disease.





 Prostate cancer is the second most common cancer in American men, killing nearly 30,000 per year. In 2004, I attended a conference where one of the nation’s leading researchers in the field declared that the gold-standard test for this disease was not successful at identifying dangerous invasive tumors. That triggered my interest in how to address the challenge of developing a blood test to detect the deadly form of prostate cancer.
Prostate cancer is the second most common cancer in American men, killing nearly 30,000 per year. In 2004, I attended a conference where one of the nation’s leading researchers in the field declared that the gold-standard test for this disease was not successful at identifying dangerous invasive tumors. That triggered my interest in how to address the challenge of developing a blood test to detect the deadly form of prostate cancer.




")















{kind=link}
{kind=link}