Healthcare analytics, AI solutions for biological big data, providing an AI platform for the biotech, life sciences, medical and pharmaceutical industries, as well as for related technological approaches, i.e., curation and text analysis with machine learning and other activities related to AI applications to these industries.
New avenues for research in membrane biology reveals the mobility of protein at work
Curator and Reporter: Dr. Premalata Pati, Ph.D., Postdoc
Membrane proteins(MPs) are proteins that exist in the plasma membrane and conduct a variety of biological functions such as ion transport, substrate transport, and signal transduction. MPs undergo function-related conformational changes on time intervals spanning from nanoseconds to seconds. Many MP structures have been solved thanks to recent developments in structural biology, particularly in single-particle cryo-Electron Microscopy (cryo-EM). Obtaining time-resolved dynamic information on MPs in their membrane surroundings, on the other hand, remains a significant difficulty.
OmpG (Open state) in a fully hydrated dimyristoylphosphatidylcholine (DMPC) bilayer. The protein is shown in light green cartoon. Lipids units are depicted in yellow, while their phosphate and choline groups are illustrated as orange and green van der Waals spheres, respectively. Potassium and chloride counterions are shown in green and purple, respectively. A continuous and semi-transparent cyan representation is used for water. https://static-content.springer.com/esm/art%3A10.1038%2Fs41467-021-24660-1/MediaObjects/41467_2021_24660_MOESM1_ESM.pdf
Weill Cornell Medicine (WCM) researchers have found that they can record high-speed protein movements while linking them to function. The accomplishment should allow scientists to examine proteins in more depth than ever before, and in theory, it should allow for the development of drugs that work better by hitting their protein targets much more effectively.
The researchers utilized High-Speed Atomic Force Microscopy (HS-AFM) to record the rapid motions of a channel protein and published in a report in Nature Communications on July 16. Such proteins generally create channel or tube-like structures in cell membranes, which open to allow molecules to flow under particular conditions. The researchers were able to record the channel protein’s rapid openings and closings with the same temporal resolution as single channel recordings, a typical technique for recording the intermittent passage of charged molecules through the channel.
Senior author Simon Scheuring, professor of physiology and biophysics in anesthesiology at WCM, said,
There has been a significant need for a tool like this that achieves such a high bandwidth that it can ‘see’ the structural variations of molecules as they work.
Researchers can now produce incredibly detailed photographs of molecules using techniques like X-ray crystallography and electron microscopy, showing their structures down to the atomic scale. The average or dominant structural positionings, or conformations, of the molecules, are depicted in these “images,” which are often calculated from thousands of individual photos. In that way, they’re similar to the long-exposure still photos from the dawn of photography.
Many molecules, on the other hand, are flexible and always-moving machinery rather than fixed structures. Scientists need to generate videos, not still photos, to reveal how such molecules move as they work, to see how their motion translates to function to catch their critical functional conformations, which may only exist for a brief moment. Current techniques for dynamic structural imaging, on the other hand, have several drawbacks, one of which being the requirement for fluorescent tags to be inserted on the molecules being photographed in many cases.
Scheuring and his lab were early adopters of the tag-free HS-AFM approach for studying molecular dynamics. The technology, which can photograph molecules in a liquid solution similar to a genuine cellular environment, employs an extremely sensitive probe, similar to a record player’s stylus, to feel its way over a molecule and therefore build up a picture of its structure. Standard HS-AFM isn’t quick enough to capture the high-speed dynamics of many proteins, but Scheuring and colleagues have developed a modified version, HS-AFM height spectroscopy(HS-AFM-HS), that works much faster by collecting dynamic changes in only one dimension: height.
The researchers used HS-AFM-HS to record the opening and closing of a relatively simple channel protein, OmpG, found in bacteria and widely studied as a model channel protein in the new study, led by the first author Raghavendar Reddy Sanganna Gari, a postdoctoral research associate in Scheuring’s laboratory. They were able to monitor OmpG gating at an effective rate of roughly 20,000 data points per second, seeing how it transitioned from open to closed states or vice versa as the acidity of the surrounding fluid varied.
More significantly, they were able to correlate structural dynamics with functional dynamics in a membrane protein of this size for the first time in a partnership with Crina Nimigean, professor of physiology and biophysics in anesthesiology, and her group at WCM.
The demonstration opens the door for a wider application of this method in basic biology and drug development.
Sanganna Gari stated,
We’re now in an exciting period of HS-AFM technology, for example using this technique to study how some drugs modulate the structural dynamics of the channel proteins they target.
Main Source
Technique reveals proteins moving as they work. By Jim Schnabel in Cornell Chronicle, August 16, 2021.
Illustration depicting a biocell attached to a CMOS integrated circuit with a membrane containing sodium-potassium pumps in pores. Energy is stored chemically in ATP molecules. When the energy is released as charged ions (which are then converted to electrons to power the chip at the bottom of the experimental device), the ATP is converted to ADP + inorganic phosphate. (credit: Trevor Finney and Jared Roseman/Columbia Engineering)
Columbia Engineering researchers have combined biological and solid-state components for the first time, opening the door to creating entirely new artificial biosystems.
In this experiment, they used a biological cell to power a conventional solid-state complementary metal-oxide-semiconductor (CMOS) integrated circuit. An artificial lipid bilayer membrane containing adenosine triphosphate (ATP)-powered ion pumps (which provide energy for cells) was used as a source of ions (which were converted to electrons to power the chip).
The study, led by Ken Shepard, Lau Family Professor of Electrical Engineering and professor of biomedical engineering at Columbia Engineering, was published online today (Dec. 7, 2015) in an open-access paper in Nature Communications.
How to build a hybrid biochip
Living systems achieve this functionality with their own version of electronics based on lipid membranes and ion channels and pumps, which act as a kind of “biological transistor.” Charge in the form of ions carry energy and information, and ion channels control the flow of ions across cell membranes.
Solid-state systems, such as those in computers and communication devices, use electrons; their electronic signaling and power are controlled by field-effect transistors.
To build a prototype of their hybrid system, Shepard’s team packaged a CMOS integrated circuit (IC) with an ATP-harvesting “biocell.” In the presence of ATP, the system pumped ions across the membrane, producing an electrical potential (voltage)* that was harvested by the integrated circuit.
“We made a macroscale version of this system, at the scale of several millimeters, to see if it worked,” Shepard notes. “Our results provide new insight into a generalized circuit model, enabling us to determine the conditions to maximize the efficiency of harnessing chemical energy through the action of these ion pumps. We will now be looking at how to scale the system down.”
While other groups have harvested energy from living systems, Shepard and his team are exploring how to do this at the molecular level, isolating just the desired function and interfacing this with electronics. “We don’t need the whole cell,” he explains. “We just grab the component of the cell that’s doing what we want. For this project, we isolated the ATPases because they were the proteins that allowed us to extract energy from ATP.”
The capability of a bomb-sniffing dog, no Alpo required
Next, the researchers plan to go much further, such as recognizing specific molecules and giving chips the potential to taste and smell.
The ability to build a system that combines the power of solid-state electronics with the capabilities of biological components has great promise, they believe. “You need a bomb-sniffing dog now, but if you can take just the part of the dog that is useful — the molecules that are doing the sensing — we wouldn’t need the whole animal,” says Shepard.
The technology could also provide a power source for implanted electronic devices in ATP-rich environments such as inside living cells, the researchers suggest.
* “In general, integrated circuits, even when operated at the point of minimum energy in subthreshold, consume on the order of 10−2 W mm−2 (or assuming a typical silicon chip thickness of 250 μm, 4 × 10−2 W mm−3). Typical cells, in contrast, consume on the order of 4 × 10−6 W mm−3. In the experiment, a typical active power dissipation for the IC circuit was 92.3 nW, and the active average harvesting power was 71.4 fW for the biocell (the discrepancy is managed through duty-cycled operation of the IC).” — Jared M. Roseman et al./Nature Communications
There is enormous potential in combining the capabilities of the biological and the solid state to create hybrid engineered systems. While there have been recent efforts to harness power from naturally occurring potentials in living systems in plants and animals to power complementary metal-oxide-semiconductor integrated circuits, here we report the first successful effort to isolate the energetics of an electrogenic ion pump in an engineered in vitro environment to power such an artificial system. An integrated circuit is powered by adenosine triphosphate through the action of Na+/K+ adenosine triphosphatases in an integrated in vitro lipid bilayer membrane. The ion pumps (active in the membrane at numbers exceeding 2 × 106mm−2) are able to sustain a short-circuit current of 32.6pAmm−2 and an open-circuit voltage of 78mV, providing for a maximum power transfer of 1.27pWmm−2 from a single bilayer. Two series-stacked bilayers provide a voltage sufficient to operate an integrated circuit with a conversion efficiency of chemical to electrical energy of 14.9%.
(a) Illustration depicting biocell attached to CMOS integrated circuit. (b) Illustration of membrane in pore containing sodium–potassium pumps. (c) Circuit model of equivalent stacked membranes, =2.1pA, =98.6GΩ, =575GΩ and =75pF, Ag/AgCl electrode equivalent resistance RWE+RCE<20kΩ, energy-harvesting capacitor CSTOR=100nF combined with switch as an impedance transformation network (only one switch necessary due to small duty cycle), and CMOS IC voltage doubler and resistor representing digital switching load. RL represents the four independent ring oscillator loads. (d) Equivalent circuit detail of stacked biocell. (e) Switched-capacitor voltage doubler circuit schematic.
The energetics of living systems are based on electrochemical membrane potentials that are present in cell plasma membranes, the inner membrane of mitochondria, or the thylakoid membrane of chloroplasts1. In the latter two cases, the specific membrane potential is known as the proton-motive force and is used by proton adenosine triphosphate (ATP) synthases to produce ATP. In the former case, Na+/K+-ATPases hydrolyse ATP to maintain the resting potential in most cells.
While there have been recent efforts to harness power from some naturally occurring potentials in living systems that are the result of ion pump action both in plants2 and animals3, 4 to power complementary metal-oxide semiconductor (CMOS) integrated circuits (ICs), this work is the first successful effort to isolate the energetics of an electrogenic ion pump in an engineered in vitroenvironment to power such an artificial system. Prior efforts to harness power from in vitromembrane systems incorporating ion-pumping ATPases5, 6, 7, 8, 9 and light-activated bacteriorhodopsin9, 10, 11 have been limited by difficulty in incorporating these proteins in sufficient quantity to attain measurable current and in achieving sufficiently large membrane resistances to harness these currents. Both problems are solved in this effort to power an IC from ATP in an in vitro environment. The resulting measurements provide new insight into a generalized circuit model, which allows us to determine the conditions to maximize the efficiency of harnessing chemical energy through the action of electrogenic ion pumps.
ATP-powered IC
Figure 1a shows the complete hybrid integrated system, consisting of a CMOS IC packaged with an ATP-harvesting ‘biocell’. The biocell consists of two series-stacked ATPase bearing suspended lipid bilayers with a fluid chamber directly on top of the IC. Series stacking of two membranes is necessary to provide the required start-up voltage for IC and eliminates the need for an external energy source, which is typically required to start circuits from low-voltage supplies2, 3. As shown inFig. 1c, a matching network in the form of a switched capacitor allows the load resistance of the IC to be matched to that presented by the biocell. In principle, the switch S can be implicit. The biocell charges CSTOR until the self start-up voltage, Vstart, is reached. The chip then operates until the biocell voltage drops below the minimum supply voltage for operation, Vmin. Active current draw from the IC stops at this point, allowing the charge to build up again on CSTOR. In our case, however, the IC leakage current exceeds 13.5nA at Vstart, more than can be provided by the biocell. As a result, an explicit transistor switch and comparator (outside of the IC) are used for this function in the experimental results presented here, which are not powered by the biocell and not included in energy efficiency calculations (see Supplementary Discussion for additional details). The energy from the biocell is used to operate a voltage converter (voltage doubler) and some simple inverter-based ring oscillators in the IC, which receive power from no other sources.
…….. Prior to the addition of ATP, the membrane produces no electrical power and has an Rm of 280GΩ. A 1.7-pA short-circuit (SC) current (Fig. 2b) through the membrane is observed upon the addition of ATP (final concentration 3mM) to the cis chamber where functional, properly oriented enzymes generate a net electrogenic pump current. To perform these measurements, currents through each membrane of the biocell are measured using a voltage-clamp amplifier (inset of Fig. 2b) with a gain of 500GΩ with special efforts taken to compensate amplifier leakage currents. Each ATPase transports three Na+ ions from the cis chamber to the trans chamber and two K+ ions from thetrans chamber to the cis chamber (a net charge movement of one cation) for every molecule of ATP hydrolysed. At a rate of 100 hydrolysis events per second under zero electrical (SC) bias13, this results in an electrogenic current of ~16aA. The observed SC current corresponds to about 105 active ATPases in the membrane or a concentration of about 2 × 106mm−2, about 5% of the density of channels occurring naturally in mammalian nerve fibres14. It is expected that half of the channels inserted are inactive because they are oriented incorrectly.
(a)…Pre-ATP data linear fit (black line) slope yield Rm=280GΩ. Post ATP data fit to a Boltzmann curve, slope=0.02V (blue line). Post-ATP linear fit (red line) yields Ip=−1.8pA and Rp=61.6GΩ, which corresponds to a per-ATP source resistance of 6.16 × 1015. The current due to membrane leakage through R_{m} is subtracted in the post-ATP curve…. (b)…
Current–voltage characteristics of the ATPases
Figure 2a shows the complete measured current–voltage (I–V) characteristic of a single ATPase-bearing membrane in the presence of ATP. The current due to membrane leakage through Rm is subtracted in the post-ATP curve. The I–V characteristic fits a Boltzmann sigmoid curve, consistent with sodium–potassium pump currents measured on membrane patches at similar buffer conditions13, 15, 16. This nonlinear behaviour reflects the fact that the full ATPase transport cycle (three Na+ ions from cis to trans and two K+ ions from trans to cis) time increases (the turn-over rate, kATP, decreases) as the membrane potential increases16. No effect on pump current is expected from any ion concentration gradients produced by the action of the ATPases (seeSupplementary Discussion). Using this Boltzmann fit, we can model the biocell as a nonlinear voltage-controlled current source IATPase (inset Fig. 2a), in which the current produced by this source varies as a function of Vm. In the fourth quadrant, where the cell is producing electrical power, this model can be linearized as a Norton equivalent circuit, consisting of a DC current source (Ip) in parallel with a current-limiting resistor (Rp), which acts to limit the current delivered to the load at increasing bias (IATPase~Ip−Vm/Rp). Figure 2c shows the measured and simulated charging of Cm for a single membrane (open-circuited voltage). A custom amplifier with input resistance Rin>10TΩ was required for this measurement (see Electrical Measurement Methods).
Reconciling operating voltage differences
The electrical characteristics of biological systems and solid-state systems are mismatched in their operating voltages. The minimum operating voltage of solid-state systems is determined by the need for transistors to modulate a Maxwell–Boltzmann (MB) distribution of carriers by several orders of magnitude through the application of a potential that is several multiples of kT/q (where kis Boltzmann’s constant, T is the temperature in degrees Kelvin and q is the elementary charge). Biological systems, while operating under the same MB statistics, have no such constraints for operating ion channels since they are controlled by mechanical (or other conformational) processes rather than through modulation of a potential barrier. To bridge this operating voltage mismatch, the circuit includes a switched-capacitor voltage doubler (Fig. 1d) that is capable of self-startup from voltages as low Vstart=145mV (~5.5kT/q) and can be operated continuously from input voltages from as low as Vmin=110mV (see Supplementary Discussion)…..
Maximizing the efficiency of harvesting energy from ATP
Solid-state systems and biological systems are also mismatched in their operating impedances. In our case, the biocell presents a source impedance, =84.2GΩ, while the load impedance presented by the complete integrated circuit (including both the voltage converter and ring oscillator loads) is approximately RIC=200kΩ. (The load impedance, RL, of the ring oscillators alone is 305kΩ.) This mismatch in source and load impedance is manifest in large differences in power densities. In general, integrated circuits, even when operated at the point of minimum energy in subthreshold, consume on the order of 10−2Wmm−2 (or assuming a typical silicon chip thickness of 250μm, 4 × 10−2Wmm−3) (ref. 17). Typical cells, in contrast, consume on the order of 4 × 10−6Wmm−3 (ref. 18). In our case, a typical active power dissipation for our circuit is 92.3nW, and the active average harvesting power is 71.4fW for the biocell. This discrepancy is managed through duty-cycled operation of the IC in which the circuit is largely disabled for long periods of time (Tcharge), integrating up the power onto a storage capacitor (CSTOR), which is then expended in a very brief period of activity (Trun), as shown in Fig. 3a.
The overall efficiency of the system in converting chemical energy to the energy consumed in the load ring oscillator (η) is given by the product of the conversion efficiency of the voltage doubler (ηconverter) and the conversion efficiency of chemical energy to electrical energy in the biocell (ηbiocell), η=ηconverter × ηbiocell. ηconverter is relatively constant over the range of input voltages at ~59%, as determined by various loading test circuits included in the chip design (Supplementary Figs 1–6). ηbiocell, however, varies with transmembrane potential Vm. η is the efficiency in transferring power to the power ring oscillator loads from the ATP harvested by biocell.
…….
To first order, the energy made available to the Na+/K+-ATPase by the hydrolysis of ATP is independent of the chemical or electric potential of the membrane and is given by |ΔGATP|/(qNA), where ΔGATP is the Gibbs free energy change due to the ATP hydrolysis reaction per mole of ATP at given buffer conditions and NA is Avogadro’s number. Since every charge that passes through IATPase corresponds to a single hydrolysis event, we can use two voltage sources in series with IATPase to independently account for the energy expended by the pumps both in moving charge across the electric potential difference and in moving ions across the chemical potential difference. The dependent voltage source Vloss in this branch fixes the voltage across IATPase, and the total power produced by the pump current source is (|ΔGATP|/NA)(NkATP), which is the product of the energy released per molecule of ATP, the number of active ATPases and the ATP turnover rate. The power dissipated in voltage source Vchem models the work performed by the ATPases in transporting ions against a concentration gradient. In the case of the Na+/K+ ATPase,Vchem is given by . The power dissipated in this source is introduced back into the circuit in the power generated by the Nernst independent voltage sources, and . The power dissipated in the dependent voltage source Vloss models any additional power not used to perform chemical or electrical work. ……
Integration of ATP-harvesting ion pumps could provide a means to power future CMOS microsystems scaled to the level of individual cells22. In molecular diagnostics, the integration of pore-forming proteins such as alpha haemolysin23 or MspA porin24 with CMOS electronics is already finding application in DNA sequencing25. Exploiting the large diversity of function available in transmembrane proteins in these hybrid systems could, for example, lead to highly specific sensing platforms for airborne odorants or soluble molecular entities26, 27. Heavily multiplexed platforms could become high-throughput in vitro drug-screening platforms against this diversity of function. In addition, integration of transmembrane proteins with CMOS may become a convenient alternative to fluorescence for coupling to synthetic biological systems28.
Himes, C., Carlson, E., Ricchiuti, R. J., Otis, B. P. & Parviz, B. A.Ultralow voltage nanoelectronics powered directly, and solely, from a tree. IEEE Trans. Nanotechnol.9, 2–5(2010).
Mercier, P. P., Lysaght, A. C., Bandyopadhyay, S., Chandrakasan, A. P. & Stankovic, K. M.Energy extraction from the biologic battery in the inner ear. Nat. Biotechnol.30, 1240–1243(2012).
Curation, HealthCare System in the US, and Calcium Signaling Effects on Cardiac Contraction, Heart Failure, and Atrial Fibrillation, and the Relationship of Calcium Release at the Myoneural Junction to Beta Adrenergic Release
Curator and e-book Contributor: Larry H. Bernstein, MD, FCAP
Curator and BioMedicine e-Series Editor-in-Chief: Aviva Lev Ari, PhD, RN
This portion summarises what we have covered and is now familiar to the reader. There are three related topics, and an extension of this embraces other volumes and chapters before and after this reading. This approach to the document has advantages over the multiple authored textbooks that are and have been pervasive as a result of the traditional publication technology. It has been stated by the founder of ScoopIt, that amount of time involved is considerably less than required for the original publications used, but the organization and construction is a separate creative process. In these curations we amassed on average five articles in one curation, to which, two or three curators contributed their views. There were surprises, and there were unfulfilled answers along the way. The greatest problem that is being envisioned is the building a vision that bridges and unmasks the hidden “dark matter” between the now declared “OMICS”, to get a more real perspective on what is conjecture and what is actionable. This is in some respects unavoidable because the genome is an alphabet that is matched to the mino acid sequences of proteins, which themselves are three dimensional drivers of sequences of metabolic reactions that can be altered by the accumulation of substrates in critical placements, and in addition, the proteome has functional proteins whose activity is a regulatory function and not easily identified. In the end, we have to have a practical conception, recognizing the breadth of evolutionary change, and make sense of what we have, while searching for more.
We introduced the content as follows:
1. We introduce the concept of curation in the digital context, and it’s application to medicine and related scientific discovery.
Topics were chosen were used to illustrate this process in the form of a pattern, which is mostly curation, but is significantly creative, as it emerges in the context of this e-book.
Alternative solutions in Treatment of Heart Failure (HF), medical devices, biomarkers and agent efficacy is handled all in one chapter.
PCI for valves vs Open heart Valve replacement
PDA and Complications of Surgery — only curation could create the picture of this unique combination of debate, as exemplified of Endarterectomy (CEA) vs Stenting the Carotid Artery (CAS), ischemic leg, renal artery stenosis.
2. The etiology, or causes, of cardiovascular diseases consist of mechanistic explanations for dysfunction relating to the heart or vascular system. Every one of a long list of abnormalities has a path that explains the deviation from normal. With the completion of the analysis of the human genome, in principle all of the genetic basis for function and dysfunction are delineated. While all genes are identified, and the genes code for all the gene products that constitute body functions, there remains more unknown than known.
3. Human genome, and in combination with improved imaging methods, genomics offers great promise in changing the course of disease and aging.
4. If we tie together Part 1 and Part 2, there is ample room for considering clinical outcomes based on individual and organizational factors for best performance. This can really only be realized with considerable improvement in information infrastructure, which has miles to go.
Curation
Curation is an active filtering of the web’s and peer reviewed literature found by such means – immense amount of relevant and irrelevant content. As a result content may be disruptive. However, in doing good curation, one does more than simply assign value by presentation of creative work in any category. Great curators comment and share experience across content, authors and themes.
Great curators may see patterns others don’t, or may challenge or debate complex and apparently conflicting points of view. Answers to specifically focused questions comes from the hard work of many in laboratory settings creatively establishing answers to definitive questions, each a part of the larger knowledge-base of reference. There are those rare “Einstein’s” who imagine a whole universe, unlike the three blindmen of the Sufi tale. One held the tail, the other the trunk, the other the ear, and they all said this is an elephant!
In my reading, I learn that the optimal ratio of curation to creation may be as high as 90% curation to 10% creation. Creating content is expensive. Curation, by comparison, is much less expensive. The same source says “Scoop.it is my content marketing testing “sandbox”. In sharing, he says that comments provide the framework for what and how content is shared.
Healthcare and Affordable Care Act
We enter year 2014 with the Affordable Care Act off to a slow start because of the implementation of the internet signup requiring a major repair, which is, unfortunately, as expected for such as complex job across the US, and with many states unwilling to participate. But several states – California, Connecticut, and Kentucky – had very effective state designed signups, separate from the federal system. There has been a very large rush and an extension to sign up. There are many features that we can take note of:
1. The healthcare system needed changes because we have the most costly system, are endowed with advanced technology, and we have inexcusable outcomes in several domains of care, including, infant mortality, and prenatal care – but not in cardiology.
2. These changes that are notable are:
The disparities in outcome are magnified by a large disparity in highest to lowest income bracket.
This is also reflected in educational status, and which plays out in childhood school lunches, and is also affected by larger class size and cutbacks in school programs.
This is not helped by a large paralysis in the two party political system and the three legs of government unable to deal with work and distraction.
Unemployment is high, and the banking and home construction, home buying, and rental are in realignment, but interest rates are problematic.
3. The medical care system is affected by the issues above, but the complexity is not to be discounted.
The medical schools are unable at this time to provide the influx of new physicians needed, so we depend on a major influx of physicians from other countries
The technology for laboratories, proteomic and genomic as well as applied medical research is rejuvenating the practice in cardiology more rapidly than any other field.
In fields that are imaging related the life cycle of instruments is shorter than the actual lifetime use of the instruments, which introduces a shortening of ROI.
Hospitals are consolidating into large consortia in order to maintain a more viable system for referral of specialty cases, and also is centralizing all terms of business related to billing.
There is reduction in independent physician practices that are being incorporated into the hospital enterprise with Part B billing under the Physician Organization – as in Partners in Greater Boston, with the exception of “concierge” medical practices.
There is consolidation of specialty laboratory services within state, with only the most specialized testing going out of state (Quest, LabCorp, etc.)
Medicaid is expanded substantially under the new ACA.
The federal government as provider of services is reducing the number of contractors for – medical devices, diabetes self-testing, etc.
The current rearrangements seeks to provide a balance between capital expenses and fixed labor costs that it can control, reduce variable costs (reagents, pharmaceutical), and to take in more patients with less delay and better performance – defined by outside agencies.
Cardiology, Genomics, and calcium ion signaling and ion-channels in cardiomyocyte function in health and disease – including heart failure, rhythm abnormalities, and the myoneural release of neurotransmitter at the vesicle junction.
This portion is outlined as follows:
2.1 Human Genome: Congenital Etiological Sources of Cardiovascular Disease
2.2 The Role of Calcium in Health and Disease
2.3 Vasculature and Myocardium: Diagnosing the Conditions of Disease
Genomics & Genetics of Cardiovascular Disease Diagnoses
disruption of Ca2+ homeostasis cardiac & vascular smooth muscle
synaptotagmin as Ca2+ sensor & vesicles
atherosclerosis & ion channels
It is increasingly clear that there are mutations that underlie many human diseases, and this is true of the cardiovascular system. The mutations are mistakes in the insertion of a purine nucleotide, which may or may not have any consequence. This is why the associations that are being discovered in research require careful validation, and even require demonstration in “models” before pursuing the design of pharmacological “target therapy”. The genomics in cardiovascular disease involves very serious congenital disorders that are asserted early in life, but the effects of and development of atherosclerosis involving large and medium size arteries has a slow progression and is not dominated by genomic expression. This is characterized by loss of arterial elasticity. In addition there is the development of heart failure, which involves the cardiomyocyte specifically. The emergence of regenerative medical interventions, based on pleuripotent inducible stem cell therapy is developing rapidly as an intervention in this sector.
Finally, it is incumbent on me to call attention to the huge contribution that research on calcium (Ca2+) signaling has made toward the understanding of cardiac contraction and to the maintenance of the heart rhythm. The heart is a syncytium, different than skeletal and smooth muscle, and the innervation is by the vagus nerve, which has terminal endings at vesicles which discharge at the myocyte junction. The heart specifically has calmodulin kinase CaMK II, and it has been established that calmodulin is involved in the calcium spark that triggers contraction. That is only part of the story. Ion transport occurs into or out of the cell, the latter termed exostosis. Exostosis involves CaMK II and pyruvate kinase (PKC), and they have independent roles. This also involves K+-Na+-ATPase. The cytoskeleton is also discussed, but the role of aquaporin in water transport appears elsewhere, as the transport of water between cells. When we consider the Gibbs-Donnan equilibrium, which precedes the current work by a century, we recall that there is an essential balance between extracellular Na+ + Ca2+ and the intracellular K+ + Mg2+, and this has been superceded by an incompletely defined relationship between ions that are cytoplasmic and those that are mitochondrial. The glass is half full!








 . The power dissipated in this source is introduced back into the circuit in the power generated by the Nernst independent voltage sources,
. The power dissipated in this source is introduced back into the circuit in the power generated by the Nernst independent voltage sources,  and
and  . The power dissipated in the dependent voltage source Vloss models any additional power not used to perform chemical or electrical work. ……
. The power dissipated in the dependent voltage source Vloss models any additional power not used to perform chemical or electrical work. ……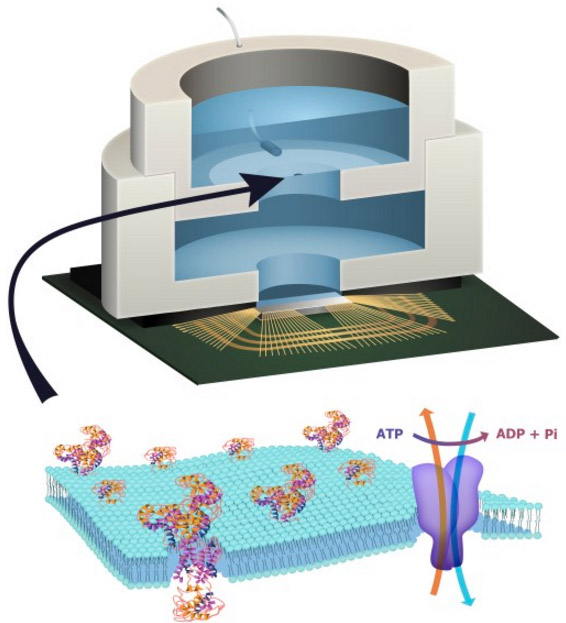{kind=link}
{kind=link}
{kind=link}