Pharmacological Action of Steroid Hormones
Curator: Larry H. Bernstein, MD, FCAP
Hormone Receptors
Steroid hormone receptors are found on the plasma membrane, in the cytosol and also in the nucleus of target cells. They are generally intracellular receptors (typically cytoplasmic) and initiate signal transduction for steroid hormones which lead to changes in gene expression over a time period of hours to days. The best studied steroid hormone receptors are members of the nuclear receptor subfamily 3 (NR3) that include receptors for estrogen (group NR3A)[1] and 3-ketosteroids (group NR3C).[2] In addition to nuclear receptors, several G protein-coupled receptors and ion channels act as cell surface receptors for certain steroid hormones.
Steroid Hormone Receptors and their Response Elements
Steroid hormone receptors are proteins that have a binding site for a particular steroid molecule. Their response elements are DNA sequences that are bound by the complex of the steroid bound to its Steroid receptor.
The response element is part of the promoter of a gene. Binding by the receptor activates or represses, as the case may be, the gene controlled by that promoter.
It is through this mechanism that steroid hormones turn genes on (or off).

steroid hormone receptor
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/S/Sigler.jpg
{kind=link}
This image (courtesy of P. B. Sigler) shows a stereoscopic view of the glucocorticoid response element (DNA, the double helix shown in yellow at the left of each panel) with the glucocorticoid receptor (a protein homodimer, right portion of each panel) bound to it.
The DNA sequence of the glucocorticoid response element is
- 5′ AGAACAnnnTGTTCT 3′
- 3′ TCTTGTnnnACAAGA 5′
where n represents any nucleotide. (Note the inverted repeats.)
The glucocorticoid receptor, like all steroid hormone receptors, is a zinc-finger transcription factor; the zinc atoms are the four yellow spheres. Each is attached to four cysteines.
For a steroid hormone to regulate (turn on or off) gene transcription, its receptor must:
- bind to the hormone (cortisol in the case of the glucocorticoid receptor)
- bind to a second copy of itself to form a homodimer
- be in the nucleus, moving from the cytosol if necessary
- bind to its response element
- bind to other protein cofactors
Each of these functions depend upon a particular region of the protein (e.g., the zinc fingers for binding DNA). Mutations in any one region may upset the function of that region without necessarily interfering with other functions of the receptor.
Positive and Negative Response Elements
Some of the hundreds of glucocorticoid response elements in the human genome activate gene transcription when bound by the hormone/receptor complex. Others inhibit gene transcription when bound by the hormone/receptor complex.
Example: When the stress hormone cortisol — bound to its receptor — enters the nucleus of a liver cell, the complex binds to
the positive response elements of the many genes needed for gluconeogenesis — the conversion of protein and fat into glucose resulting in a rise in the level of blood sugar.
the negative response element of the insulin receptor gene thus diminishing the ability of the cells to remove glucose from the blood. (This hyperglycemic effect is enhanced by the binding of the cortisol/receptor complex to a negative response element in the beta cells of the pancreas thus reducing the production of insulin.)
Note that every type of cell in the body contains the same response elements in its genome. What determines if a given cell responds to the arrival of a hormone depends on the presence of the hormone’s receptor in the cell.
Visual Evidence of Hormone Binding
This autoradiograph (courtesy of Madhabananda Sar and Walter E. Stumpf) shows the endometrial cells from the uterus of a guinea pig 15 minutes after an injection of radioactive progesterone. The radioactivity has concentrated within the nuclei of the endometrial cells as shown by the dark grains superimposed on the images of the nuclei. The same effect is seen when radioactive estrogens are administered.
The cells of the endometrium are target cells for both progesterone and estrogens, preparing the uterus for possible pregnancy. [Link to discussion]

Endometrium
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/E/Endometrium.jpg
{kind=link}
Nontarget cells (e.g. liver cells or lymphocytes) show no accumulation of female sex hormones. Although their DNA contains the response elements, their cells do not have the protein receptors needed.
The Nuclear Receptor Superfamily

Retinoids
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/R/Retinoids.png
{kind=link}
The zinc-finger proteins that serve as receptors for glucocorticoids and progesterone are members of a large family of similar proteins that serve as receptors for a variety of small, hydrophobic molecules. These include:
- other steroid hormones like
- the mineralocorticoid aldosterone
- estrogens
- the thyroid hormone, T3
- calcitriol, the active form of vitamin D
- retinoids: vitamin A (retinol) and its relatives
- retinal
- retinoic acid (tretinoin — also available as the drug Retin-A®); and its isomer
- isotretinoin (sold as Accutane® for the treatment of acne).
- bile acids
- fatty acids.

The three dimensional crystal structure of holo-retinol binding protein (RBP–ROH)


Chemical structures of vitamin A (retinol)

vit D and receptor complex
These bind members of the superfamily called peroxisome-proliferator-activated receptors (PPARs). They got their name from their initial discovery as the receptors for
- drugs that increase the number and size of peroxisomes in cells.
In every case, the receptors consist of at least
- three functional modules or domains.
From N-terminal to C-terminal, these are:
- a domain needed
- the zinc-finger domain needed for DNA binding (to the response element)
- the domain responsible for binding the particular hormone as well as the second unit of the dimer.
- for the receptor to activate the promoters of the genes being controlled
The Steroid Hormone Receptors
Klinge, C, Rao, C, Glob. libr. women’s med.,
(ISSN: 1756-2228) 2008;
http://dx.doi.org:/10.3843/GLOWM.10281
Structure of The Steroid Hormone Receptor Protein
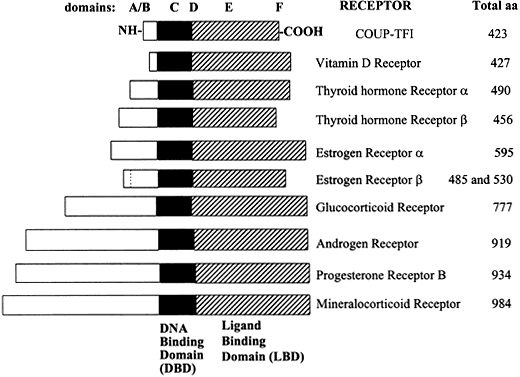
In order to understand how steroid hormone receptors regulate gene function, it is important to know the structure of the receptor proteins as well as the identity and cellular function of the genes that they regulate. Members of the steroid receptor superfamily share direct amino acid homology and a common structure (Fig. 1).
Fig. 1 Relative lengths of several members of the steroid/nuclear hormone receptor superfamily, shown schematically as linearized proteins with common structural and functional domains. Variability between members of the steroid hormone receptor family is due primarily to differences in the length and amino acid sequence of the amino (N)-terminal domain. Adapted from Wahli W, Martinez E. Superfamily of steroid nuclear receptors: Positive and negative regulators of gene expression. FASEB J 1991;5:2243-2249.

lengths of steroid hormone receptor superfamily
http://resources.ama.uk.com/glowm_www/graphics/figures/v5/0040/001f.gif
{kind=link}
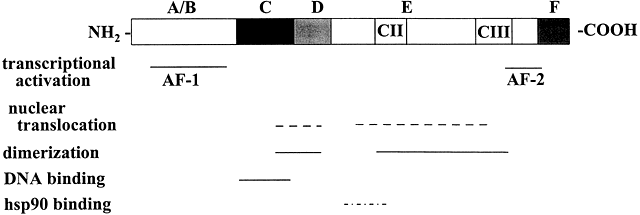
Molecular cloning of the complementary DNA (cDNA) for each of the major steroid receptors has greatly enhanced our understanding of the structure–function relationships for these molecules. The receptor proteins have five or six domains called A–F from N- to C-terminus, encoded by 8–9 exons. The receptors contain three major functional domains that have been shown experimentally to operate as independent “cassettes”,13 unrestricted as to position within the molecule. The three major functional domains (Fig. 2) of the receptor are:
- A variable N-terminus (domains A and B) that confers immunogenicity and modulates transcription in a gene and cell-specific manner through its N-terminal Activation Function-1 (AF-1);
- A central DNA-binding domain (DBD, consisting of the C domain), comprised of two functionally distinct zinc fingers through which the receptor physically interacts directly with the DNA helix;
- The ligand-binding domain (LBD, domains E and in some receptors F) that contains Activation Function-2 (AF-2).
Fig. 2 Schematic representation of the common structural and functional domains of the steroid hormone receptors. The horizontal lines indicate the domains of the receptor. Adapted from Wahli W, Martinez E. Superfamily of steroid nuclear receptors: Positive and negative regulators of gene expression. FASEB J 1991;5:2243-2249.
http://resources.ama.uk.com/glowm_www/graphics/figures/v5/0040/002f.gif
{kind=link}
The F domain is thought to play a role in distinguishing estrogen agonists from antagonists, perhaps through interaction with cell-specific factors. Domain-swapping experiments in which the DBD of estrogen receptor α (ERα) was switched with that of the glucocorticoid receptor (GR), yielded a chimeric receptor that bound to specific DNA sequences bound by GR, but up-regulated transcription of glucocorticoid-responsive target genes when treated with estrogen, thus demonstrating the specificity of the DNA-binding domain in target gene regulation.
The amino (N)-terminal domain is hypervariable (less than 15% homology among steroid receptors) in both size and amino acid sequence, ranging in length from 25 amino acids to 603 amino acids and constituting the major source of size differences between receptors. The AF-1 domain in this region is involved in activation of gene transcription, but does not depend on ligand binding. In rat GR, the AF-1 region is called tau 1 or enh2 and constitutes aa 108–317. Tau 1 is necessary for transcriptional activation and repression. Deletion of the C-terminal LBD of GR yields constitutive (hormone-independent) transcriptional activation, implying that the N-terminal regions harbor autonomous transcriptional activation functions.
Some steroid receptors exist as isoforms, encoded by the same gene, but differing in their N-terminus. The progesterone and androgen receptors (PR and AR) exist in two distinct forms, A and B, synthesized from the same mRNA by alternate splicing. The two PR receptor isoforms differ by 128 amino acids in the N-terminal region, yielding PR-A = 90 kDa and PR-B = 120 kDa, that have strikingly differing capacities to regulate transcription. In contrast, AR-A and AR-B isoforms show minimal differences in activation of a reporter gene in response to androgen agonists or antagonists in transiently transfected cells.
Receptors in this superfamily contain several key structural elements which enable them to bind to their respective ligands with high affinity and specificity, recognize and bind to discrete response elements within the DNA sequence of target genes with high affinity and specificity, and regulate gene transcription.
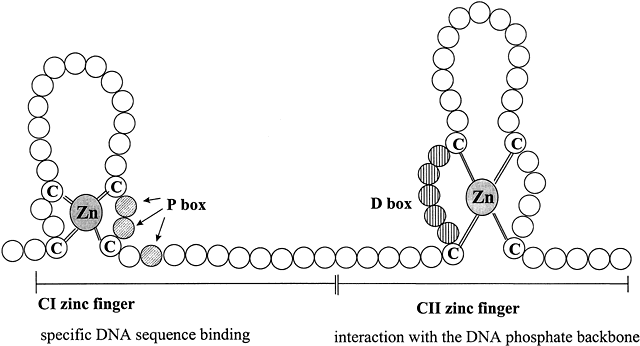
The central core or DNA-binding domain (DBD) is highly conserved and shows 60–95% homology among steroid receptors.1 The DBD varies in size from 66 to 70 amino acids, and is hydrophilic due to its high content of basic amino acids. The major function of this region is to bind to specific hormone response elements (HREs) of the target gene. DNA-binding is achieved through the tetrahedral coordination of zinc (Zn) by four cysteine residues in each of two extensions, that form two structurally distinct “Zn fingers” (Fig. 3). Zn fingers are common among gene regulatory proteins. Specificity of HRE binding is determined by the more highly conserved hydrophilic first Zn finger (C1), while the second Zn finger (C2) is involved in dimerization and stabilizing DNA binding by ionic interactions with the phosphate backbone of the DNA.18 The D box is involved in HRE half-site spacing recognition. The highly conserved DBD shared by AR, GR, mineralocorticoid receptor (MR), and PR enables them to bind to the same HRE, called the glucocorticoid response element (GRE). The more C-terminal part of the C2 Zn finger and amino acids in the hinge region are involved in receptor dimerization in coordination with amino acids in the hinge region and the LBD.
Fig. 3 Schematic diagram of type II zinc finger proteins characteristic of the DNA-binding domain structure of members of the steroid hormone receptor superfamily. Zinc fingers are common features of many transcription factors, allowing proteins to bind to DNA. Each circle represents one amino acid. The CI zinc finger interacts specifically with five base pairs of DNA and determines the DNA sequence recognized by the particular steroid receptor. The three shaded amino acids indicated by the arrows in the knuckle of the CI zinc finger are in the “P box” that allows HRE sequence discrimination between the GR and ERα. The vertically striped aa within the knuckle of the CII zinc finger constitutes the “D box” that is important for dimerization and contacts with the DNA phosphate backbone. Adapted from Tsai M-J, O’Malley BW. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem 1994;63:451-483; Gronemeyer H. Transcription activation by estrogen and progesterone receptors. Annu Rev Genet 1991;25:89-123.

type II zinc finger proteins
http://resources.ama.uk.com/glowm_www/graphics/figures/v5/0040/003f.gif
{kind=link}
The hinge region or D domain is a 40–50 amino acid sequence separating the DNA-binding and ligand-binding domains that contains sequences for receptor dimerization and ligand-dependent and independent nuclear localization sequences (NLSs). The hinge region interacts with nuclear corepressor proteins, and with L7/SPA, a 27 kDa protein that increases the partial agonist activity of certain antagonist-liganded steroid hormone receptors, i.e., tamoxifen-liganded ERα, RU486-occupied PR, or RU486-occupied GR. ….
The carboxy (C)-terminal or ligand-binding domain (LBD) is poorly conserved, ranging in size from 218 to 264 amino acids and is hydrophobic. This region contains the ligand-binding site and dictates hormone binding specificity.
Two human GR isoforms, GRα and GRβ, derived from the same gene by differential splicing at the C-terminus, have been reported. While GRα and GRβ share the first eight exons, they differ in their last two exons, i.e., exons 9α or 9β, spliced into the respective mRNA.40 GRβ was reported to localize in the cell nucleus in the absence of ligand and to block hGRα activity. …
Sequences within the LBD form the binding site for hsp90 that blocks the DBD in the cytosolic, nonliganded GR.40 The CII and CIII regions (Fig. 2) show homology among members of the steroid/nuclear receptor superfamily and are important in forming the ligand binding pocket. …
The Bifunctional Role of Steroid Hormones: Implications for Therapy in Prostate Cancer
Paul Mathew, MD
Review Article | May 15, 2014 | Oncology Journal, Genitourinary Cancers, Prostate Cancer
In a biomarker-driven study reported in 1941, Drs. Huggins and Hodges of the University of Chicago demonstrated reduction in elevated levels of serum acid phosphatase in five men with metastatic prostate cancer treated with estrogens and orchiectomy, whereas three men who received testosterone injections after orchiectomy exhibited increased serum levels of the enzyme. Hitherto, serum elevations of acid phosphatase had been associated strictly with prostate cancer, and Huggins and Hodges thus concluded that androgens activated prostate cancer. Nevertheless, in the years that followed, several investigators experimented with testosterone injections in prostate cancer. Pearson[3] of the Sloan-Kettering Institute reviewed the inconsistent biochemical and clinical responses to testosterone injections associated with these studies and puzzled over two case studies of his own, one of a hormone-naive patient, another of a castration-resistant patient, both of whom had responded to testosterone injection: “These observations invite the development of new concepts to explain the response of these prostatic cancers to alterations in the endocrine environment.”
Table 1: Sex Steroids as Tumor Suppressors (not shown)
ABSTRACT: Ablation of the androgen-signaling axis is currently a dominant theme in developmental therapeutics in prostate cancer. Highly potent inhibitors of androgen biosynthesis and androgen receptor (AR) function have formally improved survival in castration-resistant metastatic disease. Resistance to androgen-ablative strategies arises through diverse mechanisms. Strategies to preserve and extend the success of hormonal therapy while mitigating the emergence of resistance have long been of interest. In preclinical models, intermittent hormonal ablative strategies delay the emergence of resistant stem-cell–driven phenotypes, but clinical studies in hormone-naive disease have not observed more than noninferiority over continual androgen ablation. In castration-resistant disease, response and improvement in subjective quality of life with therapeutic testosterone has been observed, but so too has symptomatic and life-threatening disease acceleration. The multifunctional and paradoxical role of steroid hormones in regulating proliferation and differentiation, as well as cell death, requires deeper understanding. The lack of molecular biomarkers that predict the outcome of hormone supplementation in a particular clinical context remains an obstacle to individualized therapy. Biphasic patterns of response to hormones are identifiable in vitro, and endocrine-regulated neoplasms that proliferate after prolonged periods of hormone deprivation appear preferentially sex steroid–suppressible. This review examines the relevance of a translational framework for studying therapeutic androgens in prostate cancer.
Protection and Damage from Acute and Chronic Stress: Allostasis and Allostatic Overload and Relevance to the Pathophysiology of Psychiatric Disorders
Bruce S. Mcewen*
Annals of the New York Academy of Sciences 12 JAN 2006;
1032 (Biobehavioral Stress Response: Protective and Damaging Effects): Pp1–328
http://dx.doi.org:/10.1196/annals.1314.001
Keywords:
stress;psychiatric disorders;depression;allostasis;allostatic overload;homeostasis
Abstract: Stress promotes adaptation, but prolonged stress leads over time to wear-and-tear on the body (allostatic load). Neural changes mirror the pattern seen in other body systems, that is, short-term adaptation vs. long-term damage. Allostatic load leads to impaired immunity, atherosclerosis, obesity, bone demineralization, and atrophy of nerve cells in the brain. Many of these processes are seen in major depressive illness and may be expressed also in other chronic anxiety disorders. The brain controls the physiological and behavioral coping responses to daily events and stressors. The hippocampal formation expresses high levels of adrenal steroid receptors and is a malleable brain structure that is important for certain types of learning and memory. It is also vulnerable to the effects of stress and trauma. The amygdala mediates physiological and behavioral responses associated with fear. The prefrontal cortex plays an important role in working memory and executive function and is also involved in extinction of learning. All three regions are targets of stress hormones. In animal models, neurons in the hippocampus and prefrontal cortex respond to repeated stress by showing atrophy, whereas neurons in amygdala show a growth response. Yet, these are not necessarily “damaged” and may be treatable with the right medications.