The SCID Pig II: Researchers Develop Another SCID Pig, And Another Great Model For Cancer Research
Updated 6/25/2019
Writer. Reporter: Stephen J. Williams, Ph.D.

The choice of suitable animal model of disease may define future success or failure for drug development, basic and translational research, or biomarker discovery projects. Indeed, as highlighted in one of my earlier posts “Heroes in Medical Research: Developing Models for Cancer Research”, the choice of animal to model a human disease can have drastic implications in the basic researchers ability to understand metabolic and genetic factors causally associated with disease development. As described in that post the King rat model led to our understanding of the genetics of early development and sex determination while early mouse models helped us to understand the impact of microenvironment on cell fate and the discovery of stem cells. In addition, transgenic and immunodeficient mice resulted in transformational studies on our understanding of cancer. Small rodent models are ideal for following reasons:
- Ease of genetic manipulation
- Availability of well-defined models
- Ease of low cost of use
Regardless of these benefits many investigators in industry and academia are looking to models of human disease in animals more closely resembling human anatomy, physiology, and genetics.
There is a growing need for alternative animal models in cancer research.
As I had discussed in another of my earlier posts “The SCID Pig: How Pigs are becoming a Great Alternate Model for Cancer Research”, the pig is gaining notoriety and acceptance as a very suitable animal to model human disease as minipigs and humans have:
- Similar physiology
- Similar genetics: >90% homology
- Similar anatomic dimensions: i.e. Adult Gottingen minipigs are 70kg (adult human male weight)
- Similar organ size and structure to humans organ size and structure
- Pig genome sequencing project nearly complete
- Ability to manipulate pig genetics
The post had discussed the development of a severe combined immunodeficient (SCID) pig by investigators at Iowa State and Kansas State University. This line of pigs, selected on a specific diet, could act as recipients for human cancer cell lines, a proof of their SCID phenotype.
A report featured on Fierce Biotech Research “MU Scientists Successfully Transplant, Grow Stem Cells in Pigs” discussed the development of a new genetically-modified immunodeficient porcine model by researchers at the University of Missouri, recently published in Proceedings of the National Academy of Sciences[1].
These pigs are available from the National Swine Resource and Research Center (http://nsrrc.missouri.edu).
For the report on Fierce Biotech Research please follow the link below:
The report in FierceBiotech highlights the type of studies an immunocompromised pig model would be useful for including:
- Regenerative medicine
- Xenotransplantation
- Tumor growth and efficacy studies
Comments in the post from the investigators explained the benefits of developing such a porcine model system including:
“The rejection of transplants and grafts by host bodies is a huge hurdle for medical researchers,” said R. Michael Roberts, Curators Professor of Animal Science and Biochemistry and a researcher in the Bond Life Sciences Center. “By establishing that these pigs will support transplants without the fear of rejection, we can move stem cell therapy research forward at a quicker pace.”
The studies main investigators, Drs. Randall Prather and R. Michael Roberts, both of University of Missouri, along with first authors Kiho Lee, Deug-Nam Kwon and Toshihiko Ezashi, used biallellic mutation of the RAG2 gene in Gottingen minipig fibroblasts and then subsequent somatic cell nuclear transfer (SCNT) to produce the RAG2-/- animals. (Rag2 is a protein involved in V(D)J recombination of antibodies during early B and T cell development. See GeneCard link above)
As proof of their SCID phenotype the authors showed that
- these RAG2-/- animals could act as host for human induced pluripotent stem cells
- act as recipient for allogeneic porcine stem cells
- reduced levels of (CD21+) B cells and (CD3+) T cells
- growth retardation if housed under standard, non-sterile conditions
Details of the study are given below:
Methodology Used
For Production of Gottingen minipigs carrying the RAG2 mutation
To produce targeted mutations in RAG2:
- TALENS () were constructed to produced mutation in exon 2 of RAG2
- Constructed TALENS and reporter electroporated in fetal-derived pig fibroblasts
- SCNT used to transfer RAG2 mutant nuclei to donor oocytes
- 9 embryo transfers resulted in 22 live piglets
- Piglets genotyped as either monoallelic or biallelic RAG2 mutant
- RAG2wild-type and mutants housed in either pathogen-free or normal housing conditions
To verify SCID phenotype of litter by either
- Graft acceptance of human iPSCs and teratoma formation
– Fibroblasts from human umbilical cord reprogrammed to pluripotency; verified by pluripotent markers POUSF1, NANOG, SSEA-3)
– Two human and porcine iPSC lines with trophoblastic properties[2] were injected subcutaneously in ear or flank
– Tumor formation analyzed by immunohistochemistry using markers:
CTNNBI (B-catenin)
VWF (von Willebrand
DES and ACTG2
GFAP and ENO2
Human specific MFN1 (both antibody and gene primers)
- Flow Cytometry
– Analysis of piglet spleen cells for B cell population (CD21)
– Analysis of piglet spleen cell for T cell population (CD3)
C. Histology
– histo evaluation of thymus, spleen
– marker evaluation of spleen using anti-CD79A (B cells), CD3 (T cells),
CD335 (NK cells)
Results
TALEN produced a variety of indels (insertion/deletions) and three RAG2 mutatnt colonies (containing monoallelic, mix of mono and biallelic) used for SCNT.
Three litters produced 16 piglets (eight survived [four mono and four biallelic]
Biallelic RAG2 mutants showed slower weight gain than wild type or monoallelic mutants with signs of inflammation and apoptosis in spleen and designated “failure to thrive” in standard housing…needed a clean environment to thrive.
Biallelic mutant pigs lacked mature CD21 B cells and CD3 T cells but contained macrophages and NK cells.
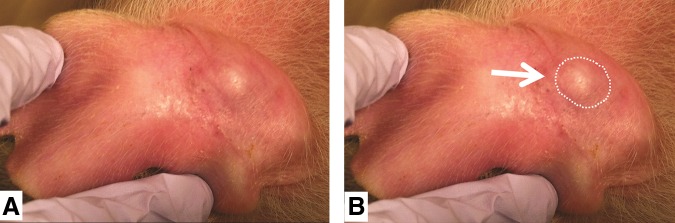
Implantation of human and allogenic porcine pluripotent stem cells (trophoblastic) showed rapid development of teratomas.
References
- Lee K, Kwon DN, Ezashi T, Choi YJ, Park C, Ericsson AC, Brown AN, Samuel MS, Park KW, Walters EM et al: Engraftment of human iPS cells and allogeneic porcine cells into pigs with inactivated RAG2 and accompanying severe combined immunodeficiency. Proceedings of the National Academy of Sciences of the United States of America 2014, 111(20):7260-7265.
- Ezashi T, Matsuyama H, Telugu BP, Roberts RM: Generation of colonies of induced trophoblast cells during standard reprogramming of porcine fibroblasts to induced pluripotent stem cells. Biology of reproduction 2011, 85(4):779-787.
Updated 6/25/2019
The following articles represent an update on the ability to create genetically predisposed porcine models of cancer. The ability to utilize transposable elements to introduce genetic changes in porcine cells in combination with Somatic Cell Nuclear Transfer technology with the ultimate goal to create a transgenic minipig is discussed. The next two articles describe the ability of the scid pig to act as a recipient for human ovarian cancer cells and description of a transgenic inducible porcine intestinal tumor model.
Transgenic Res. 2011 Jun;20(3):533-45. doi: 10.1007/s11248-010-9438-x. Epub 2010 Aug 29.
Pig transgenesis by Sleeping Beauty DNA transposition.
Jakobsen JE1, Li J, Kragh PM, Moldt B, Lin L, Liu Y, Schmidt M, Winther KD, Schyth BD, Holm IE, Vajta G, Bolund L, Callesen H, Jørgensen AL, Nielsen AL, Mikkelsen JG.
Abstract
Modelling of human disease in genetically engineered pigs provides unique possibilities in biomedical research and in studies of disease intervention. Establishment of methodologies that allow efficient gene insertion by non-viral gene carriers is an important step towards development of new disease models. In this report, we present transgenic pigs created by Sleeping Beauty DNA transposition in primary porcine fibroblasts in combination with somatic cell nuclear transfer by handmade cloning. Göttingen minipigs expressing green fluorescent protein are produced by transgenesis with DNA transposon vectors carrying the transgene driven by the human ubiquitin C promoter. These animals carry multiple copies (from 8 to 13) of the transgene and show systemic transgene expression. Transgene-expressing pigs carry both transposase-catalyzed insertions and at least one copy of randomly inserted plasmid DNA. Our findings illustrate critical issues related to DNA transposon-directed transgenesis, including coincidental plasmid insertion and relatively low Sleeping Beauty transposition activity in porcine fibroblasts, but also provide a platform for future development of porcine disease models using the Sleeping Beauty gene insertion technology.
This paper makes use of two technologies: transposon mediated gene transfer to introduce foreign DNA, for example a disease predisposition gene, into oocytes or early embryos, without the use of viral vectors; and use of SCNT to clone a minipig from viable embryos.
The transposon mediated system is based on the Sleeping Beauty (SB) vector system, which is a cut and paste DNA transposon belonging to the Tc1/mariner superfamily of transposable elements(1). Transposable DNA elements are mobile genetic elements which integrate into genomic DNA, in the case of the SB system into discrete sequence elements of actively transcribed genes. The system consists of two entities: 1) a transposase responsible for cutting and pasting the mobile element and 2) the transposon; the vectorial DNA sequence which is inserted into genomic DNA. SB transposition has been used to integrate exogenous genetic elements into the genome of various mammalian species(2) and to make tumor models in mice (3-7) and to transform, ex-vivo, porcine ovarian epithelial cells (8) and to stably integrate GFP containing vectors into porcine fibroblast genome(9). Because of the efficiency and nonviral integration of exogenous vectors into mammalian systems, Sleeping Beauty system has been considered as a potential therapeutic gene transfer modality (10-12).
- Li, Z.H., Liu, D.P., Wang, J., Guo, Z.C., Yin, W.X., and Liang, C.C. Inversion and transposition of Tc1 transposon of C. elegans in mammalian cells. Somat Cell Mol Genet. 1998; 24:363-369.
- Balciuniene, J., Nagelberg, D., Walsh, K.T., Camerota, D., Georlette, D., Biemar, F., et al. Efficient disruption of Zebrafish genes using a Gal4-containing gene trap. BMC Genomics. 2013; 14:619.
- Romano, G., Marino, I.R., Pentimalli, F., Adamo, V., and Giordano, A. Insertional mutagenesis and development of malignancies induced by integrating gene delivery systems: implications for the design of safer gene-based interventions in patients. Drug News Perspect. 2009; 22:185-196.
- Dupuy, A.J. Transposon-based screens for cancer gene discovery in mouse models. Semin Cancer Biol. 2010; 20:261-268.
- Dupuy, A.J., Akagi, K., Largaespada, D.A., Copeland, N.G., and Jenkins, N.A. Mammalian mutagenesis using a highly mobile somatic Sleeping Beauty transposon system. Nature. 2005; 436:221-226.
- Dupuy, A.J., Clark, K., Carlson, C.M., Fritz, S., Davidson, A.E., Markley, K.M., et al. Mammalian germ-line transgenesis by transposition. Proc Natl Acad Sci U S A. 2002; 99:4495-4499.
- Dupuy, A.J., Fritz, S., and Largaespada, D.A. Transposition and gene disruption in the male germline of the mouse. Genesis. 2001; 30:82-88.
- Hamilton, T.C., Williams, S.J., and Cvetkovic, D. 2010. Cancer Compositions, Animal Models, and Methods of Use Thereof. U.S.P. Office, editor. USA: Fox Chase Cancer Center.
- Clark, K.J., Carlson, D.F., Foster, L.K., Kong, B.W., Foster, D.N., and Fahrenkrug, S.C. Enzymatic engineering of the porcine genome with transposons and recombinases. BMC Biotechnol. 2007; 7:42.
- Ivics, Z., and Izsvak, Z. Transposable elements for transgenesis and insertional mutagenesis in vertebrates: a contemporary review of experimental strategies. Methods Mol Biol. 2004; 260:255-276.
- Liu, H., Liu, L., Fletcher, B.S., and Visner, G.A. Sleeping Beauty-based gene therapy with indoleamine 2,3-dioxygenase inhibits lung allograft fibrosis. FASEB J. 2006; 20:2384-2386.
- Ohlfest, J.R., Lobitz, P.D., Perkinson, S.G., and Largaespada, D.A. Integration and long-term expression in xenografted human glioblastoma cells using a plasmid-based transposon system. Mol Ther. 2004; 10:260-268.
A second paper, by Larry Shook and Geoffrey Clark’s groups describe the production of ex vivo transformed porcine breast cancer line, driven by inactivation of BRCA1. In this paper normal porcine breast epithelial cells were immortalized by transfection with SV large T antigen (SV-LT) and upon inactivation of porcine BRCA1 in these immortalized cell lines, developed phenotype characteristic of transformed cells and exhibited cancer stem cell characteristics. The end point assay for transformation was growth in soft agar however the authors did not confirm malignancy in either SCID mice or SCID pigs.
Front Genet. 2015 Aug 25;6:269. doi: 10.3389/fgene.2015.00269. eCollection 2015.
A porcine model system of BRCA1 driven breast cancer.
Donninger H1, Hobbing K2, Schmidt ML3, Walters E4, Rund L5, Schook L5, Clark GJ2.
Author information
Abstract
BRCA1 is a breast and ovarian tumor suppressor. Hereditary mutations in BRCA1 result in a predisposition to breast cancer, and BRCA1expression is down-regulated in ~30% of sporadic cases. The function of BRCA1 remains poorly understood, but it appears to play an important role in DNA repair and the maintenance of genetic stability. Mouse models of BRCA1 deficiency have been developed in an attempt to understand the role of the gene in vivo. However, the subtle nature of BRCA1 function and the well-known discrepancies between human and murine breast cancer biology and genetics may limit the utility of mouse systems in defining the function of BRCA1 in cancer and validating the development of novel therapeutics for breast cancer. In contrast to mice, pig biological systems, and cancer genetics appear to more closely resemble their human counterparts. To determine if BRCA1 inactivation in pig cells promotes their transformation and may serve as a model for the human disease, we developed an immortalized porcine breast cell line and stably inactivated BRCA1 using miRNA. The cell line developed characteristics of breast cancer stem cells and exhibited a transformed phenotype. These results validate the concept of using pigs as a model to study BRCA1 defects in breast cancer and establish the first porcine breast tumor cell line.
Figure 1. Immortalization of pig mammary epithelial cells. Primary pig breast epithelial cells were stably transfected with an SV40 LT expression construct and selected in puromycin. Surviving cells were serially passaged to confirm immortalization.

Figure 3. Loss of BRCA1 enhances pig mammary epithelial cell growth. (A) Serially passaging the pig mammary epithelial cells stably knocked down for BRCA1 resulted in an altered morphology compared to those cells stably expressing the LacZ miRNA. (B) 2 × 104 cells/well were plated in 6-well plates and cell growth was determined by counting the number of cells at the indicated times. Error bars show standard error, p < 0.05.

Figure 5. Loss of BRCA1 enhances the transformed phenotype of pig mammary epithelial cells. (A) The pig breast epithelial cells stably expressing BRCA1 miRNA were plated in soft agar and scored for growth 14 days later. Representative photomicrographs are shown in the top panel and data from three independent experiments quantitated in the bar graph in the lower panel. (B) 1 × 106 cells/well were plated in polyHEMA-coated 12-well plates and cell viability assessed 48 h later by trypan blue staining. Error bars show standard error, p < 0.05.

A third paper describes the development, in Gottingen minipigs, of a transgenic inducible model of intestinal cancer.
Mol Oncol. 2017 Nov;11(11):1616-1629. doi: 10.1002/1878-0261.12136. Epub 2017 Oct 10.
A genetically inducible porcine model of intestinal cancer.
Callesen MM1, Árnadóttir SS1, Lyskjaer I1, Ørntoft MW1, Høyer S2, Dagnaes-Hansen F3, Liu Y4, Li R4, Callesen H4, Rasmussen MH1, Berthelsen MF3, Thomsen MK3, Schweiger PJ5, Jensen KB5, Laurberg S6, Ørntoft TF1, Elverløv-Jakobsen JE3, Andersen CL1.
Author information
Abstract
Transgenic porcine cancer models bring novel possibilities for research. Their physical similarities with humans enable the use of surgical procedures and treatment approaches used for patients, which facilitates clinical translation. Here, we aimed to develop an inducible oncopig model of intestinal cancer. Transgenic (TG) minipigs were generated using somatic cell nuclear transfer by handmade cloning. The pigs encode two TG cassettes: (a) an Flp recombinase-inducible oncogene cassette containing KRAS-G12D, cMYC, SV40LT – which inhibits p53 – and pRB and (b) a 4-hydroxytamoxifen (4-OHT)-inducible Flp recombinase activator cassette controlled by the intestinal epithelium-specific villin promoter. Thirteen viable transgenic minipigs were born. The ability of 4-OHT to activate the oncogene cassette was confirmed in vitro in TG colonic organoids and ex vivo in tissue biopsies obtained by colonoscopy. In order to provide proof of principle that the oncogene cassette could also successfully be activated in vivo, three pigs were perorally treated with 400 mg tamoxifen for 2 × 5 days. After two months, one pig developed a duodenal neuroendocrine carcinoma with a lymph node metastasis. Molecular analysis of the carcinoma and metastasis confirmed activation of the oncogene cassette. No tumor formation was observed in untreated TG pigs or in the remaining two treated pigs. The latter indicates that tamoxifen delivery can probably be improved. In summary, we have generated a novel inducible oncopig model of intestinal cancer, which has the ability to form metastatic disease already two months after induction. The model may be helpful in bridging the gap between basic research and clinical usage. It opens new venues for longitudinal studies of tumor development and evolution, for preclinical assessment of new anticancer regimens, for pharmacology and toxicology assessments, as well as for studies into biological mechanisms of tumor formation and metastasis.
Other posts on this site related to Cancer Research Tools include
The SCID Pig: How Pigs are becoming a Great Alternate Model for Cancer Research
Heroes in Medical Research: Developing Models for Cancer Research
Reprogramming Induced Pleuripotent Stem Cells
The Cancer Research Concentration @ Leaders in Pharmaceutical Business Intelligence
A Synthesis of the Beauty and Complexity of How We View Cancer
Guidelines for the welfare and use of animals in cancer research
Gene Therapy and the Genetic Study of Disease: @Berkeley and @UCSF – New DNA-editing technology spawns bold UC initiative as Crispr Goes Global