Summary, Metabolic Pathways
Author: Larry H. Bernstein, MD, FCAP
This portion of a series of chapters on metabolism, proteomics and metabolomics dealt mainly with carbohydrate metabolism. Amino acids and lipids are presented more fully in the chapters that follow. There are features on the
- functioning of enzymes and proteins,
- on sequential changes in a chain reaction, and
- on conformational changes that we shall also cover.
These are critical to developing a more complete understanding of life processes.
I needed to lay out the scope of metabolic reactions and pathways, and their complementary changes. These may not appear to be adaptive, if the circumstances and the duration is not clear. The metabolic pathways map in total
is in interaction with environmental conditions – light, heat, external nutrients and minerals, and toxins – all of which give direction and strength to these reactions. A developing goal is to discover how views introduced by molecular biology and genomics don’t clarify functional cellular dynamics that are not related to the classical view. The work is vast.
Carbohydrate metabolism denotes the various biochemical processes responsible for the formation, breakdown and interconversion of carbohydrates in living organisms. The most important carbohydrate is glucose, a simple sugar (monosaccharide) that is metabolized by nearly all known organisms. Glucose and other carbohydrates are part of a wide variety of metabolic pathways across species: plants synthesize carbohydrates from carbon dioxide and water by photosynthesis storing the absorbed energy internally, often in the form of starch or lipids. Plant components are consumed by animals and fungi, and used as fuel for cellular respiration. Oxidation of one gram of carbohydrate yields approximately 4 kcal of energy and from lipids about 9 kcal. Energy obtained from metabolism (e.g. oxidation of glucose) is usually stored temporarily within cells in the form of ATP. Organisms capable of aerobic respiration metabolize glucose and oxygen to release energy with carbon dioxide and water as byproducts.
Carbohydrates are used for short-term fuel, and even though they are simpler to metabolize than fats, they don’t produce as equivalent energy yield measured by ATP. In animals, the concentration of glucose in the blood is linked to the pancreatic endocrine hormone, insulin. . In most organisms, excess carbohydrates are regularly catabolized to form acetyl-CoA, which is a feed stock for the fatty acid synthesis pathway; fatty acids, triglycerides, and other lipids are commonly used for long-term energy storage. The hydrophobic character of lipids makes them a much more compact form of energy storage than hydrophilic carbohydrates.
Glucose is metabolized obtaining ATP and pyruvate by way of first splitting a six-carbon into two three carbon chains, which are converted to lactic acid from pyruvate in the lactic dehydrogenase reaction. The reverse conversion is by a separate unidirectional reaction back to pyruvate after moving through pyruvate dehydrogenase complex.
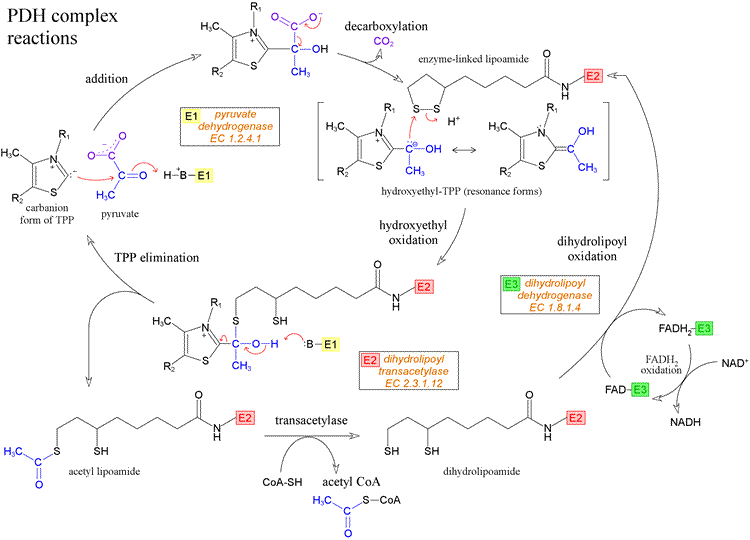
Pyruvate dehydrogenase complex (PDC) is a complex of three enzymes that convert pyruvate into acetyl-CoA by a process called pyruvate decarboxylation. Acetyl-CoA may then be used in the citric acid cycle to carry out cellular respiration, and this complex links the glycolysis metabolic pathway to the citric acid cycle. This multi-enzyme complex is related structurally and functionally to the oxoglutarate dehydrogenase and branched-chain oxo-acid dehydrogenase multi-enzyme complexes. In eukaryotic cells the reaction occurs inside the mitochondria, after transport of the substrate, pyruvate, from the cytosol. The transport of pyruvate into the mitochondria is via a transport protein and is active, consuming energy. On entry to the mitochondria pyruvate decarboxylation occurs, producing acetyl CoA. This irreversible reaction traps the acetyl CoA within the mitochondria. Pyruvate dehydrogenase deficiency from mutations in any of the enzymes or cofactors results in lactic acidosis.

PDH-rxns The acetyl group is transferred to coenzyme A
http://guweb2.gonzaga.edu/faculty/cronk/biochem/images/PDH-rxns.gif
{kind=link}
Typically, a breakdown of one molecule of glucose by aerobic respiration (i.e. involving both glycolysis and Kreb’s cycle) is about 33-35 ATP. This is categorized as:
Glycogenolysis – the breakdown of glycogen into glucose, which provides a glucose supply for glucose-dependent tissues.
Glycogenolysis in liver provides circulating glucose short term.
Glycogenolysis in muscle is obligatory for muscle contraction.
Pyruvate from glycolysis enters the Krebs cycle, also known as the citric acid cycle, in aerobic organisms.
Anaerobic breakdown by glycolysis – yielding 8-10 ATP
Aerobic respiration by Kreb’s cycle – yielding 25 ATP
The pentose phosphate pathway (shunt) converts hexoses into pentoses and regenerates NADPH. NADPH is an essential antioxidant in cells which prevents oxidative damage and acts as precursor for production of many biomolecules.
Glycogenesis – the conversion of excess glucose into glycogen as a cellular storage mechanism; achieving low osmotic pressure.
Gluconeogenesis – de novo synthesis of glucose molecules from simple organic compounds. An example in humans is the conversion of a few amino acids in cellular protein to glucose.
Metabolic use of glucose is highly important as an energy source for muscle cells and in the brain, and red blood cells.
The hormone insulin is the primary glucose regulatory signal in animals. It mainly promotes glucose uptake by the cells, and it causes the liver to store excess glucose as glycogen. Its absence
- turns off glucose uptake,
- reverses electrolyte adjustments,
- begins glycogen breakdown and glucose release into the circulation by some cells,
- begins lipid release from lipid storage cells, etc.
The level of circulatory glucose (known informally as “blood sugar”) is the most important signal to the insulin-producing cells.
- insulin is made by beta cells in the pancreas,
- fat is stored n adipose tissue cells, and
- glycogen is both stored and released as needed by liver cells.
- no glucose is released to the blood from internal glycogen stores from muscle cells.
The hormone glucagon, on the other hand, opposes that of insulin, forcing the conversion of glycogen in liver cells to glucose, and then release into the blood. Growth hormone, cortisol, and certain catecholamines (such as epinepherine) have glucoregulatory actions similar to glucagon. These hormones are referred to as stress hormones because they are released under the influence of catabolic proinflammatory (stress) cytokines – interleukin-1 (IL1) and tumor necrosis factor α (TNFα).
| Net Yield of GlycolysisThe preparatory phase consumes 2 ATP
The pay-off phase produces 4 ATP. The gross yield of glycolysis is therefore 4 ATP – 2 ATP = 2 ATP The pay-off phase also produces 2 molecules of NADH + H+ which can be further converted to a total of 5 molecules of ATP* by the electron transport chain (ETC) during oxidative phosphorylation. Thus the net yield during glycolysis is 7 molecules of ATP* |
Cellular respiration involves 3 stages for the breakdown of glucose – glycolysis, Kreb’s cycle and the electron transport system. Kreb’s cycle produces about 60-70% of ATP for release of energy in the body. It directly or indirectly connects with all the other individual pathways in the body.
The Kreb’s Cycle occurs in two stages:
|
Each pyruvate in the presence of pyruvate dehydrogenase (PDH) complex in the mitochondria gets converted to acetyl CoA which in turn enters the Kreb’s cycle. This reaction is called as oxidative decarboxylation as the carboxyl group is removed from the pyruvate molecule in the form of CO2 thus yielding 2-carbon acetyl group which along with the coenzyme A forms acetyl CoA.
The PDH requires the sequential action of five co-factors or co-enzymes for the combined action of dehydrogenation and decarboxylation to take place. These five are TPP (thiamine phosphate), FAD (flavin adenine dinucleotide), NAD (nicotinamide adenine dinucleotide), coenzyme A (denoted as CoA-SH at times to depict role of -SH group) and lipoamide.
Acetyl CoA condenses with oxaloacetate (4C) to form a citrate (6C) by transferring its acetyl group in the presence of enzyme citrate synthase. The CoA liberated in this reaction is ready to participate in the oxidative decarboxylation of another molecule of pyruvate by PDH complex.
Isocitrate undergoes oxidative decarboxylation by the enzyme isocitrate dehydrogenase to form oxalosuccinate (intermediate- not shown) which in turn forms α-ketoglutarate (also known as oxoglutarate) which is a five carbon compound. CO2 and NADH are released in this step. α-ketoglutarate (5C) undergoes oxidative decarboxylation once again to form succinyl CoA (4C) catalysed by the enzyme α-ketoglutarate dehydrogenase complex.
Succinyl CoA is then converted to succinate by succinate thiokinase or succinyl coA synthetase in a reversible manner. This reaction involves an intermediate step in which the enzyme gets phosphorylated and then the phosphoryl group which has a high group transfer potential is transferred to GDP to form GTP.
Succinate then gets oxidised reversibly to fumarate by succinate dehydrogenase. The enzyme contains iron-sulfur clusters and covalently bound FAD which when undergoes electron exchange in the mitochondria causes the production of FADH2.
Fumarate is then by the enzyme fumarase converted to malate by hydration(addition of H2O) in a reversible manner.
Malate is then reversibly converted to oxaloacetate by malate dehydrogenase which is NAD linked and thus produces NADH.
The oxaloacetate produced is now ready to be utilized in the next cycle by the citrate synthase reaction and thus the equilibrium of the cycle shifts to the right.
The NADH formed in the cytosol can yield variable amounts of ATP depending on the shuttle system utilized to transport them into the mitochondrial matrix. This NADH, formed in the cytosol, is impermeable to the mitochondrial inner-membrane where oxidative phosphorylation takes place. Thus to carry this NADH to the mitochondrial matrix there are special shuttle systems in the body. The most active shuttle is the malate-aspartate shuttle via which 2.5 molecules of ATP are generated for 1 NADH molecule. This shuttle is mainly used by the heart, liver and kidneys. The brain and skeletal muscles use the other shuttle known as glycerol 3-phosphate shuttle which synthesizes 1.5 molecules of ATP for 1 NADH.
Glucose-6-phosphate Dehydrogenase is the committed step of the Pentose Phosphate Pathway. This enzyme is regulated by availability of the substrate NADP+. As NADPH is utilized in reductive synthetic pathways, the increasing concentration of NADP+ stimulates the Pentose Phosphate Pathway, to replenish NADPH. The importance of this pathway can easily be underestimated. The main source for energy in respiration was considered to be tied to the high energy phosphate bond in phosphorylation and utilizes NADPH, converting it to NADP+. The pentose phosphate shunt is essential for the generation of nucleic acids, in regeneration of red cells and lens – requiring NADPH.
NAD+ serves as electron acceptor in catabolic pathways in which metabolites are oxidized. The resultant NADH is reoxidized by the respiratory chain, producing ATP.
The pyridine nucleotide transhydrogenase reaction concerns the energy-dependent reduction of TPN by DPNH. In 1959, Klingenberg and Slenczka made the important observation that incubation of isolated liver mitochondria with DPN-specific substrates or succinate in the absence of phosphate acceptor resulted in a rapid and almost complete reduction of the intramitochondrial TPN. These and related findings led Klingenberg and co-workers (1-3) to postulate the occurrence of a ATP-controlled transhydrogenase reaction catalyzing the reduction of TPN by DPNH. (The role of transhydrogenase in the energy-linked reduction of TPN. Fritz Hommes, Ronald W. Estabrook, The Wenner-Gren Institute, University of Stockholm, Stockholm, Sweden. Biochemical and Biophysical Research Communications 11, (1), 2 Apr 1963, Pp 1–6. http://dx.doi.org:/10.1016/0006-291X(63)90017-2/).
Further studies observed the coupling of TPN-specific dehydrogenases with the transhydrogenase and observing the reduction of large amounts of diphosphopyridine nucleotide (DPN) in the presence of catalytic amounts of triphosphopyridine nucleotide (TPN). The studies showed the direct interaction between TPNHz and DPN, in the presence of transhydrogenase to yield products having the properties of TPN and DPNHZ. The reaction involves a transfer of electrons (or hydrogen) rather than a phosphate. (Pyridine Nucleotide Transhydrogenase II. Direct Evidence for and Mechanism of the Transhydrogenase Reaction* by Nathan 0. Kaplan, Sidney P. Colowick, And Elizabeth F. Neufeld. (From The Mccollum-Pratt Institute, The Johns Hopkins University, Baltimore, Maryland) J. Biol. Chem. 1952, 195:107-119.) http://www.JBC.org/Content/195/1/107.Citation
Notation: TPN, NADP; DPN, NAD+; reduced pyridine nucleotides: TPNH (NADPH2), DPNH (NADH).
Note: In this discussion there is a detailed presentation of the activity of lactic acid conversion in the mitochondria by way of PDH. In a later section there is mention of the bidirectional reaction of lactate dehydrogenase. However, the forward reaction is dominant (pyruvate to lactate) and is described. This is not related to the kinetics of the LD reaction with respect to the defining characteristic – Km.
Biochemical Education Jan 1977; 5(1):15. Kinetics of Lactate Dehydrogenase: A Textbook Problem.
K.L. MANCHESTER. Department of Biochemistry, University of Witwatersrand, Johannesburg South Africa.
One presupposes that determined Km values are meaningful under intracellular conditions. In relation to teaching it is a simple experiment for students to determine for themselves the Km towards pyruvate of LDH in a post-mitochondrial supernatant of rat heart and thigh muscle. The difference in Km may be a factor of 3 or 4-fold.It is pertinent then to ask what is the range of suhstrate concentrations over which a difference in Km may be expected to lead to significant differences in activity and how these concentrations compare with pyruvate concentrations in the cell. The evidence of Vesell and co-workers that inhibition by pyruvate is more readily seen at low than at high enzyme concentration is important in emphasizing that under intracellular conditions enzyme concentrations may be relatively large in relation to the substrate available. This will be particularly so in relation to [NADH] which in the cytoplasm is likely to be in the ~M range.
A final point concerns the kinetic parameters for LDH quoted by Bergmeyer for lactate estimations a pH of 9 is recommended and the Km towards lactate at that pH is likely to be appreciably different from the quoted values at pH 7 — Though still at pH 9 showing a substantially lower value for lactate with the heart preparation. http://onlinelibrary.wiley.com/doi/10.1016/0307-4412%2877%2990013-9/pdf
Several investigators have established that epidermis converts most of the glucose it uses to lactic acid even in the presence of oxygen. This is in contrast to most tissues where lactic acid production is used for energy production only when oxygen is not available. This large amount of lactic acid being continually produced within the epidermal cell must be excreted by the cell and then carried away by the blood stream to other tissues where the lactate can be utilized. The LDH reaction with pyruvate and NADH is reversible although at physiological pH the equilibrium position for the reaction lies very far to the right, i.e., in favor of lactate production. The speed of this reaction depends not only on the amount of enzyme present but also on the concentrations of the substances involved on both sides of the equation. The net direction in which the reaction will proceed depends solely on the relative concentrations of the substances on each side of the equation.
In vivo there is net conversion of pyruvate (formed from glucose) to lactate. Measurements of the speed of lactate production by sheets of epidermis floating on a medium containing glucose indicate a rate of lactate production of approximately 0.7 rn/sm/
mm/mg of fresh epidermis.Slice incubation experiments are presumably much closer to the actual in vivo conditions than
the homogenate experiments. The discrepancy between the
two indicates that in vivo conditions are far from optimal for the conversion of pyruvate to lactate. Only 1/100th of the maximal activity of the enzyme present is being achieved. The concentrations of the various substances involved are not
optimal in vivo since pyruvate and NADH concentrations are
lower than lactate and NAD concentrations and this might explain the in vivo inhibition of LDH activity. (Lactate Production And Lactate Dehydrogenase In The Human Epidermis*. KM. Halprin, A Ohkawara. J Invest Dermat 1966; 47(3): 222-6.)
http://www.nature.com/jid/journal/v47/n3/pdf/jid1966133a.pdf