Highlights of a Green Evolution
Reporter and Curator: Larry H Bernstein, MD, FCAP
Chlorophyll

chlorophyll coloration to leaves
Paul May
School of Chemistry, University of Bristol
VRML, Jmol, and Chime versions
Chlorophyll is the molecule that absorbs sunlight and uses its energy to
synthesize carbohydrates from CO2 and water. This process is known as
photosynthesis. Animals and humans obtain their food supply by eating plants.
In 1780, the famous English chemist Joseph Priestley found that plants could “restore air which has been injured by the burning of candles.” He placed a mint
plant into a vessel of water for several days, then found that “the air would neither extinguish a candle, nor was it all inconvenient to a mouse which I put into it”.
He discovered that plants produce oxygen. Then, in 1794, Antoine Lavoisier
discovered oxidation. It fell to a Dutchman, Jan Ingenhousz, to make the next
major contribution to the mechanism of photosynthesis.
Having heard of Priestley’s experiments, he spent a summer near London doing
over 500 experiments, to discover that light plays a major role in photosynthesis.
He noted that plants not only have the faculty to correct bad air in six to ten days,
but they perform this in a few hours; owing to the influence of light of the sun
upon the plant.
Very soon after, more pieces of the puzzle were found by two chemists working
in Geneva. Jean Senebier, found that “fixed air” (CO2) was taken up during photosynthesis, and Theodore de Saussure discovered that the other reactant
necessary was water. The final contribution came from a German surgeon,
Julius Robert Mayer ,

Julius Robert Mayer
who recognised that plants convert solar energy into chemical energy. He said:
“Nature has put itself the problem of how to catch in flight light streaming to
the Earth and to store the most elusive of all powers in rigid form. The plants
take in one form of power, light; and produce another power, chemical
difference.” The actual chemical equation which takes place is the reaction
between carbon dioxide and water, catalyzed by sunlight, to produce glucose
and a waste product, oxygen. The glucose sugar is either directly used as an
energy source by the plant for metabolism or growth, or is polymerized to form
starch, so it can be stored until needed. The waste oxygen is excreted into the
atmosphere, where it is made use of by plants and animals for respiration.
http://www.chm.bris.ac.uk/motm/chlorophyll/photosth.gif
{kind=link}
Chlorophyll as a Photoreceptor
Chlorophyll is the molecule that traps this ‘most elusive of all powers’ – and is
called a photoreceptor. It is found in the chloroplasts of green plants,
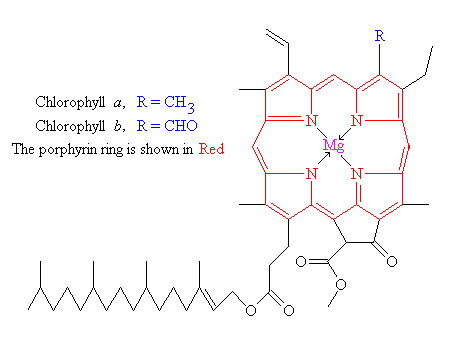
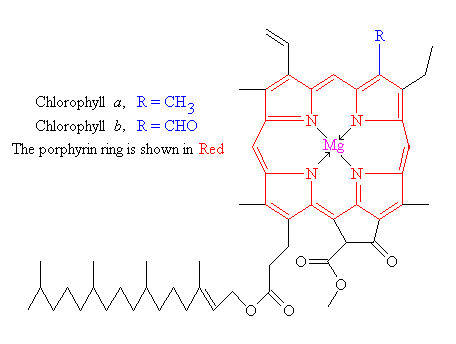
and is what makes green plants, green. The basic structure of a chlorophyll
molecule is a porphyrin ring, co-ordinated to a central atom. This is very
similar in structure to the heme group found in hemoglobin, except that in
heme the central atom is iron, whereas in chlorophyll it is magnesium.
http://www.chm.bris.ac.uk/motm/chlorophyll/chphyll.gif
{kind=link}
Click for 3D structure file
There are actually 2 main types of chlorophyll, named a and b. They differ only
slightly, in the composition of a sidechain (in a it is – H3, in b it is CHO). Both of these
two chlorophylls are very effective photoreceptors because they contain a network of
alternating single and double bonds, and the orbitals can delocalize stabilizing the
structure. Such delocalised polyenes have very strong absorption bands in the visible
regions of the spectrum, allowing the plant to absorb the energy from sunlight.
http://www.chm.bris.ac.uk/motm/chlorophyll/chloroabs.gif
{kind=link}
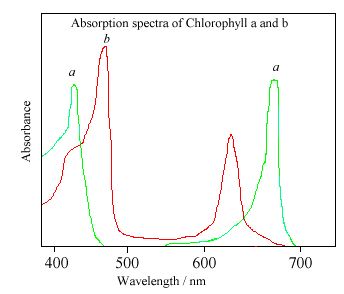
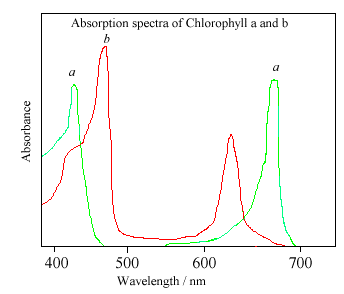
The different side groups in the 2 chlorophylls ‘tune’ the absorption spectrum to
slightly different wavelengths, so that light that is not significantly absorbed by
chlorophyll a, at, say, 460nm, will instead be captured by chlorophyll b, which
absorbs strongly at that wavelength. Thus these two kinds of chlorophyll
complement each other in absorbing sunlight. Plants can obtain all their energy
requirements from the blue and red parts of the spectrum, however, there is still
a large spectral region, between 500-600nm, where very little light is absorbed.
This light is in the green region of the spectrum, and since it is reflected, this
is the reason plants appear green. Chlorophyll absorbs so strongly that it can
mask other less intense colours. Some of these more delicate colours (from
molecules such as carotene and quercetin) are revealed when the chlorophyll
molecule decays in the Autumn, and the woodlands turn red, orange,and
golden brown. Chlorophyll can also be damaged when vegetation is cooked,
since the central Mg atom is replaced by hydrogen ions. This affects the energy
levels within the molecule, causing its absorbance spectrum to alter. Thus cooked
leaves change colour – often becoming a paler, insipid yellowy green.
As the chlorophyll in leaves decays in the autumn, the green colour fades and is
replaced by the oranges and reds of carotenoids.
Chlorophyll in Plants
The chlorophyll molecule is the active part that absorbs the sunlight, but just as with
hemoglobin, in order to do its job (synthesising carbohydrates) it needs to be attached
to the backbone of a very complicated protein. This protein may look haphazard in
design, but it has exactly the correct structure to orient the chlorophyll molecules in
the optimal position to enable them to react with nearby CO2 and H2O molecules in
a very efficient manner. Several chlorophyll molecules are lurking inside this bacterial
photoreceptor protein (right).
References:
Introduction to Organic Chemistry, Streitweiser and Heathcock (MacMillan, New York,
1981).
Biochemistry, L. Stryer (W.H. Freeman and Co, San Francisco, 1975).
Wikipedia – Chlorophyll
Chlorophyll (also chlorophyl) is a green pigment found in cyanobacteria and the
chloroplasts of algae and plants. Its name is derived from the Greek words χλωρός,
chloros (“green”) and φύλλον, phyllon (“leaf”). Chlorophyll is an extremely important
biomolecule, critical in photosynthesis, which allows plants to absorb energy from light. Chlorophyll absorbs light most strongly in the blue portion of the
electromagnetic spectrum, followed by the red portion. Conversely, it is a poor
absorber of green and near-green portions of the spectrum, hence the green
color of chlorophyll-containing tissues. chlorophyll was first isolated by
Joseph Bienaimé Caventou and Pierre Joseph Pelletier in 1817.
Absorption maxima of chlorophylls against the spectrum of white light
![]()
Chlorophyll is found in high concentrations in chloroplasts of plant cells.

http://upload.wikimedia.org/wikipedia/commons/thumb/0/05/Clorofila_3.jpg/
120px-Clorofila_3.jpg
{kind=link}
These chlorophyll maps show milligrams of chlorophyll per cubic meter of seawater
each month. Places where chlorophyll amounts were very low, indicating very low
numbers of phytoplankton, are blue. Places where chlorophyll concentrations were
high, meaning many phytoplankton were growing, are yellow.

chlophyll world map
{kind=link}
Chlorophyll and photosynthesis
Chlorophyll is vital for photosynthesis, which allows plants to absorb energy from light.
Chlorophyll molecules are specifically arranged in and around photosystems that are
embedded in the thylakoid membranes of chloroplasts. In these complexes,
chlorophyll serves two primary functions. The function of the vast majority of
chlorophyll (up to several hundred molecules per photosystem) is to absorb light and
transfer that light energy by resonance energy transfer to a specific chlorophyll pair
in the reaction center of the photosystems.
The two currently accepted photosystem units are Photosystem II and Photosystem I,
which have their own distinct reaction center chlorophylls, named P680 and P700,
respectively. These pigments are named after the wavelength (in nanometers) of their
red-peak absorption maximum. The identity, function and spectral properties of the
types of chlorophyll in each photosystem are distinct and determined by each other
and the protein structure surrounding them. Once extracted from the protein into a
solvent (such as acetone or methanol), these chlorophyll pigments can be separated
in a simple paper chromatography experiment and, based on the number of polar
groups between chlorophyll a and chlorophyll b, will chemically separate out on the
paper.
The function of the reaction center chlorophyll is to use the energy absorbed by and
transferred to it from the other chlorophyll pigments in the photosystems to undergo
a charge separation, a specific redox reaction in which the chlorophyll donates an
electron into a series of molecular intermediates called an electron transport chain.
The charged reaction center chlorophyll (P680+) is then reduced back to its ground
state by accepting an electron. In Photosystem II, the electron that reduces P680+
ultimately comes from the oxidation of water into O2 and H+ through several
intermediates.
This reaction is how photosynthetic organisms such as plants produce O2 gas, and
is the source for practically all the O2 in Earth’s atmosphere. Photosystem I typically
works in series with Photosystem II; thus the P700+ of Photosystem I is usually
reduced, via many intermediates in the thylakoid membrane, by electrons ultimately
from Photosystem II. Electron transfer reactions in the thylakoid membranes are
complex, however, and the source of electrons used to reduce P700+ can vary.
The electron flow produced by the reaction center chlorophyll pigments is used to
shuttle H+ ions across the thylakoid membrane, setting up a chemiosmotic potential
used mainly to produce ATP chemical energy; and those electrons ultimately reduce
NADP+ to NADPH, a universal reductant used to reduce CO2 into sugars as well as
for other biosynthetic reductions.
Reaction center chlorophyll–protein complexes are capable of directly absorbing light
and performing charge separation events without other chlorophyll pigments, but the
absorption cross section (the likelihood of absorbing a photon under a given light
intensity) is small. Thus, the remaining chlorophylls in the photosystem and antenna
pigment protein complexes associated with the photosystems all cooperatively absorb
and funnel light energy to the reaction center. Besides chlorophyll a, there are other
pigments, called accessory pigments, which occur in these pigment–protein
antenna complexes.
Chemical structure
Chlorophyll is a chlorin pigment, which is structurally similar to and produced through the same metabolic pathway as other porphyrin pigments such as heme. At the center
of the chlorin ring is a magnesium ion. This was discovered in 1906, and was the first
time that magnesium had been detected in living tissue. or the structures depicted in
this article, some of the ligands attached to the Mg2+ center are omitted for clarity.
The chlorin ring can have several different side chains, usually including a long
phytol chain. There are a few different forms that occur naturally, but the most
widely distributed form in terrestrial plants is chlorophyll a.

Chlorophyll-a-3D
{kind=link}
Space-filling model of the chlorophyll a molecule
After initial work done by German chemist Richard Willstätter spanning from 1905 to
1915, the general structure of chlorophyll a was elucidated by Hans Fischer in 1940.
By 1960, when most of the stereochemistry of chlorophyll a was known, Robert Burns
Woodward published a total synthesis of the molecule. In 1967, the last remaining
stereochemical elucidation was completed by Ian Fleming, and in 1990 Woodward
and co-authors published an updated synthesis. Chlorophyll was announced to be
present in cyanobacteria and other oxygenic microorganisms that form stromatolites
in 2010; a molecular formula of C55H70O6N4Mg and a structure of (2-formyl)-chlorophyll a were deduced based on NMR, optical and mass spectra.
When leaves degreen in the process of plant senescence, chlorophyll is converted
to a group of colourless tetrapyrroles known as nonfluorescent chlorophyll catabolites
(NCC’s) with the general structure:
These compounds have also been identified in several ripening fruits

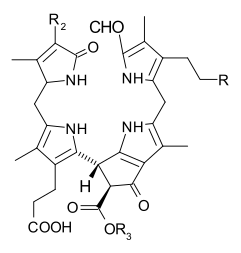{kind=link}
Absorbance spectra of free chlorophyll a (blue) and b (red) in a solvent. The spectra
of chlorophyll molecules are slightly modified in vivo depending on specific pigment-
protein interactions.

Chlorophyll_ab_spectra
{kind=link}
Complementary light absorbance of anthocyanins with chlorophylls
Anthocyanins are other plant pigments. The absorbance pattern responsible for the
red color of anthocyanins may be complementary to that of green chlorophyll in
photosynthetically active tissues such as young Quercus coccifera leaves. It may
protect the leaves from attacks by plant eaters that may be attracted by green color.
Superposition of spectra of chlorophyll a and b with oenin (malvidin 3O glucoside),
a typical anthocyanidin, showing that, while chlorophylls absorb in the blue and
yellow/red parts of the visible spectrum, oenin absorbs mainly in the green part
of the spectrum, where chlorophylls don’t absorb at all.

Superposition of spectra of chlorophyll a and b with oenin
.PNG/220px-Spectra_Chlorophyll_ab_oenin_(1).PNG){kind=link}
Many important natural substances are chelates. In chelates a central metal ion is
bonded to a large organic molecule, a molecule composed of carbon, hydrogen, and
other elements such as oxygen and nitrogen. One such chelate is chlorophyll, the
green pigment of plants. In chlorophyll the central ion is magnesium, and the large
organic molecule is a porphyrin. The porphyrin contains four nitrogen atoms that form
bonds to magnesium in a square planar arrangement. There are several forms of
chlorophyll. The structure of one form, chlorophyll a, is shown.
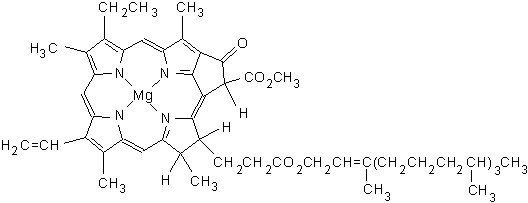
http://scifun.chem.wisc.edu/chemweek/chlrphyl/chlrphyl.gif
{kind=link}
(As you can see from the molecular structure, the “chloro” in chlorophyll does not
mean that it contains the element chlorine. The chloro portion of the word is from
the Greek chloros, which means yellowish green. The name of the element chlorine
comes from the same source. Chlorine is a yellowish green gas.)
Chlorophyll is one of the most important chelates in nature. It is capable of
channeling the energy of sunlight into chemical energy through the process of
photosynthesis. In photosynthesis, the energy absorbed by chlorophyll transforms
carbon dioxide and
water into carbohydrates and oxygen.
CO2 + H2O ——- (CH2O) + O2
(In this equation, (CH2O) is the empirical formula of carbohydrates.) The chemical
energy stored by photosynthesis in carbohydrates drives biochemical reactions in
nearly all living organisms.
In the photosynthetic reaction, carbon dioxide is reduced by water; in other words,
electrons are transferred from water to carbon dioxide. Chlorophyll assists this
transfer. When chlorophyll absorbs light energy, an electron in chlorophyll is excited
from a lower energy state to a higher energy state. In this higher energy state, this
electron is more readily transferred to another molecule. This starts a chain of
electron-transfer steps, which ends with an electron transferred to carbon dioxide.
Meanwhile, the chlorophyll which gave up an electron can accept an electron from
another molecule. This is the end of a process which starts with the removal of an
electron from water. Thus, chlorophyll is at the center of the photosynthetic
oxidation-reduction reaction between carbon dioxide and water.
Other molecules with structures similar to that of chlorophyll play important roles in
other biochemical electron-transfer (oxidation-reduction) reactions. Heme consists
of a porphyrin similar to that in chlorophyll and an iron(II) ion in the center of the
porphyrin. Heme is bright red. In the red blood cells of vertebrates, heme is bound
to proteins forming hemoglobin. Hemoglobin combines with oxygen in the lungs, gills,
or other respiratory surfaces and releases it in the tissues. In muscle cells, myoglobin,
the name given to hemoglobin in muscles, stores oxygen as an electron source for
energy-releasing oxidation-reduction reactions.
Another relative of chlorophyll is vitamin B12. Vitamin B12 contains a cobalt ion at
the center of the porphyrin. Like heme, vitamin B12 is bright red. It is essential to
digestion and nutritional absorption in animals. The exact way it functions is not
known. Because vitamin B12 is not produced by higher plants, a strictly vegetarian
diet can lead to vitamin B12 deficiency. However, it is produced by molds and
bacteria which grow on most foods.
The intense color of chlorophyll suggests that it may be useful as a commercial
pigment. In fact, chlorophyll a is a green dye (Natural Green 3) used in soaps and
cosmetics. The absorption spectrum of chlorophyll (below) shows that it absorbs
strongly in the red and blue-violet regions of the visible spectrum. Because it absorbs
red and blue-violet light, the light it reflects and transmits appears green. Commercial
pigments with structures similar to chlorophyll have been produced in a range of colors.
Some of these have slightly modified porphyrins, such as having hydrogen atoms
replaced with chlorine atoms. Others have different metal ions. For example, one
bright blue pigment has a copper(I) ion at the center of the porphyrin and is used
primarily in coloring fabrics.
http://scifun.chem.wisc.edu/chemweek/chlrphyl/clrphlsp.gif
{kind=link}