Cilia and Tubulin
Larry H. Bernstein, MD, FCAP, Curator
LPBI
RESEARCHERS IDENTIFY LIPID RESPONSIBLE FOR KEEPING BRAIN FLUID MOVING

Cilia labelled in ependymal cells with tubulin antibodies by indirect immunofluorescence. 1) Acetylated tubulin (cilia labelling)/ Alexa 488, 2) Détyrosinated tubulin (cilia labelling)/ Cy3, 3) Tyrosinated tubulin (cilia labelling in pseudo color magenta)/ Cy5, 4) Cell nuclei stained with Hoescht Courtesy of Dr Saoudi, GIN, Grenoble.
Previous studies show that in the case of algae, motile cilia work like arms on a swimmer, to help algae reach sunlight, which they use to make sugar for energy and survival. Motile cilia can make 40-50 whip-like motions per second and are continuously being resorbed and replaced. These motile cilia also inhabit a type of support brain cell called ependymal cells; the cells that line the cavities of the brain containing cerebrospinal fluid which nourishes and cleanses the brain.
Earlier studies from the lab were the first to show that ceramide is present in these fast-moving algae cilia where it has likely been at work for a billion years. The group reasoned that algae, an old and simple plant, seemed a good first model to figure out what regulates cilia. Cilia in algae are comparatively big, about 10 times those in mammalian cells, so with just a regular microscope, the researchers could easily watch cilia movement. The current study showed that when they applied even a miniscule amount of a fungus toxin known to inhibit ceramide production, the usually agile algae stopped, the cilia shrank and some fell off.
The current study shows that one of the protein kinases that is highly conserved from Chlamydomonas to mammals and critical for ciliogenesis is glycogen synthase kinase-3 (GSK3). The lab explain that in Chlamydomonas an GSK3 isoform regulates flagellar length. Results show that ceramide present in the cilia of human ependymal cells activates GSK3 to regulate cilia. The researchers conclude that in both algae and ependymal cells, the two can be found together and, without their teamwork, hydrocephalus, excess fluid on the brain, can result.
The team surmise that their findings suggest that if ceramide is one of the key regulators of cerebrospinal fluid movement, it may also be a drug target to help normalize the function of cilia. For the future, the researchers plan to visualize cilia movement in ependymal cells.
Source: University of Georgia
Regulation of Chlamydomonas flagella and ependymal cell motile cilia by ceramidemediated translocation of GSK3
Ji Na Kong1 , Kara Hardin1 , Michael Dinkins1 , Guanghu Wang1 , Qian He1 , Tarik Mujadzic1 , Gu Zhu1 , et al.
Mol. Biol. Cell October 7, 2015mbc.E15-06-0371 http://www.molbiolcell.org/content/early/2015/10/05/mbc.E15-06-0371.full.pdf
Cilia are important organelles formed by cell membrane protrusions, however, little is known about their regulation by membrane lipids. We characterize a novel activation mechanism for glycogen synthase kinase-3 (GSK3) by the sphingolipids phytoceramide and ceramide, which is critical for ciliogenesis in Chlamydomonas and murine ependymal cells, respectively. We show for the first time that Chlamydomonas expresses serine palmitoyl transferase (SPT), the first enzyme in (phyto)ceramide biosynthesis. Inhibition of SPT in Chlamydomonas by myriocin led to loss of flagella and reduced tubulin acetylation, which was prevented by supplementation with the precursor dihydrosphingosine. Immunocytochemistry showed that (phyto)ceramide was colocalized with phospho-tyr216-GSK3 (pYGSK3) at the base and tip of Chlamydomonas flagella and motile cilia in ependymal cells. The (phyto)ceramide distribution was consistent with that of a bifunctional ceramide analog UV-crosslinked and visualized by click-chemistry mediated fluorescent labeling. Ceramide depletion, by myriocin or neutral sphingomyelinase deficiency (fro/fro mouse), led to GSK3 dephosphorylation and defective flagella and cilia. Motile cilia were rescued and pYGSK3 localization restored by incubation of fro/fro ependymal cells with exogenous C24:1 ceramide, which directly bound to pYGSK3. Our findings suggest that (phyto)ceramide-mediated translocation of pYGSK into flagella and cilia is an evolutionarily conserved mechanism fundamental to the regulation of ciliogenesis.
Flagella and cilia are slender organellar protrusions of the eukaryotic cell membrane with sensory and motor function. Flagella and cilia are similar in that they share a microtubule-based cytoskeleton called the axoneme (Snell et al., 2004; Sung and Leroux, 2013). Primary, nonmotile cilia have a ring of nine outer microtubule doublets and are studded with growth factor receptors to function as sensory organelles regulating cell migration and differentiation. Motile cilia and flagella contain two central microtubules and the motor protein dynein attached to the nine peripheral microtubule doublets (see (Silflow and Lefebvre, 2001; Snell et al., 2004; Satir et al., 2010) for comprehensive reviews on the structure of primary and motile cilia/flagella). Motile cilia and flagella move extracellular fluids and particles, or propel cells such as sperm or unicellular organisms such as the green algae Chlamydomonas reinhardtii (Vincensini et al., 2011). Flagella dysfunction leads to reduced or absent motility, while aberrant motile cilia lead to ependymal cell malfunction and hydrocephalus (Young et al., 2013). It is vital for the signaling and motility functions of cilia that length, number, and intraciliary or intraflagellar transport (IFT) of cargo are dynamically regulated. The protein machinery regulating assembly, length, movement, and transport within these organelles is conserved from Chlamydomonas to mammals (Snell et al., 2004; Vincensini et al., 2011; Sung and Leroux, 2013). Although cilia and flagella are localized extensions of the plasma mebrane, little is known about the role of lipids in this regulation of ciliogenesis and cilium function.
Sphingolipids are essential components of eukaryotic membranes and regulate vital cell signaling pathways (Bartke and Hannun, 2009; Milhas et al., 2010; Bieberich, 2012; Markham et al., 2013). The common long chain base precursor for de novo biosynthesis of animal, plant, fungus, and algae sphingolipids is dihydrosphingosine (DHS, Figure 1). DHS is converted to ceramide (animals) and phytoceramide (plants, algae, and yeast). In animals, ceramide and its metabolite, sphingomyelin are unique in that their levels can be rapidly regulated by enzymatic conversion, the so-called sphingomyelin cycle (Figure 1)(Hannun, 1996). Ceramide is converted to sphingomyelin by sphingomyelin synthases, while sphingomyelin is hydrolyzed to ceramide by acid or neutral sphingomyelinases (aSMase, nSMase) (Nikolova-Karakashian et al., 2008; Milhas et al., 2010; Shamseddine et al., 2015). We previously reported that aSMase and nSMase-mediated generation of ceramide is critical for ciliogenesis in MDCK cells and neural progenitors, respectively (Wang et al., 2009a; He et al., 2012; He et al., 2014). In contrast to mammalian cells, Chlamydomonas and plants do not synthesize sphingomyelin, which therefore is excluded as a source for ceramide. However, in all eukaryotic cells, ceramides can be generated by de novo biosynthesis (Figure 1) (Sperling and Heinz, 2003; Gault et al., 2010; Pata et al., 2010; Hannun and Obeid, 2011; Markham et al., 2013). The first enzyme in this pathway, serine palmitoyltransferase (SPT) can be selectively inhibited by the fungal toxin myriocin, allowing for loss-of-function experiments to understand the significance of ceramide for ciliogenesis in animal cells and algae (Miyake et al., 1995; Hanada et al., 2000; Wadsworth et al., 2013). The long chain base in algae and plant sphingolipids is more complex than that in animals. Hydroxylation of DHS at C4 results in biosynthesis of phytosphingosine, which can be acylated to give rise to phytoceramides (Figure 1) (Spassieva et al., 2002; Sperling and Heinz, 2003; Pata et al., 2010; Markham et al., 2013). While plants and algae sphingolipids are relatively well known, the sphingolipid pathway in Chlamydomonas has not been investigated. Physical interaction of (phyto)ceramide with proteins that are critical for ciliogenesis is likely to underlie the regulation of flagella and motile cilia, respectively. Previously, we found that in MDCK cells and human ES cell-derived neuroprogenitors, atypical PKCδ (aPKCδ) directly interacts with ceramide and promotes primary cilium extension ultimately through inhibition of histone deacetylase 6 (HDAC6) (Bieberich et al., 2000; Wang et al., 2005; Wang et al., 2009b; He et al., 2012; He et al., 2014). HDAC6 inhibition prevents deacetylation of lysine 40 of α-tubulin, a reaction that destabilizes microtubules and promotes cilium disassembly (Pugacheva et al., 2007; Loktev et al., 2008). A Chlamydomonas tubulin deacetylase has been described, but it is not clear whether it has the same function for flagella regulation as HDAC6 for cilia in animals (Maruta et al., 1986). It is likely that ceramide interacts with additional proteins to regulate HDAC6 and/or cilium length. One candidate protein, conserved in Chlamydomonas and mammalian cilia is GSK-3β (Wilson and Lefebvre, 2004; Thoma et al., 2007). GSK-3β (GSK3) is critical for ciliogenesis, regulates HDAC6, and phosphorylates kinesin light chains that control cargo binding to microtubules (Szebenyi et al., 2002; Thoma et al., 2007; Song et al., 2014).
To identify ceramide-protein complexes in cilia, we used chemoproteomics technology for in vitro and in vivo UV cross-linking of a bifunctional ceramide analog (pacFACer) to interacting proteins and functionalizing this covalent complex for visualization using “Click chemistry” (azide-alkyne cycloaddition, Figure S1 for structure). In addition, we have developed a novel anti-ceramide antibody that recognizes ceramide and phytoceramide. Using pacFACer cross-linking and anti-ceramide antibody, we demonstrate that Chlamydomonas flagella and ependymal cell cilia show a similar distribution of ceramides that bind to GSK3, which is critical for the regulation of ciliogenesis by an evolutionarily conserved mechanism of ceramidemediated translocation of GSK3 into flagella and cilia.
Figures not shown
Chlamydomonas expresses serine palmitoyltransferase and generates plant sphingolipids There is no prior information on the presence of sphingolipids or enzymes required for sphingolipid biosynthesis in Chlamydomonas. Using reverse transcriptase (RT)-PCR and oligonucleotide primers based on sequence similarities between mouse serine palmitoyltransferase (SPT), the first enzyme in the sphingolipid biosynthetic pathway, and Chlamydomonas expressed sequence tagged (EST) clones, we found that Chlamydomonas expresses mRNAs for the subunits SPT1 and SPT2 (Figure 2A). mRNAs for these SPT subunits are similar to those expressed in mouse ependymal cells enriched from primary cultured glial cells (Weibel et al., 1986). In addition to SPT, we also detected mRNAs for lag1 and lag2, two ceramide synthases expressed in plants, suggesting that Chlamydomonas expresses several enzymes in the phytoceramide biosynthesis pathway (Figure S1B). Expression of SPT was confirmed by immunoblot. Using an antibody raised against mammalian SPTs, we were able to detect SPT2 at the predicted molecular weight (72 kD) but not SPT1 in Chlamydomonas (Figure 2B). The expression of SPTs and ceramide synthases in Chlamydomonas suggested that sphingolipids are synthesized. Using LC-MS/MS, we found that Chlamydomonas generates very long chain C26:0 (11±3 pmol/mg cells) and C28:0 (8±2 pmol/mg cells) phytoceramides, which are common plant sphingolipids (Figure 2C). In comparison, glial cells contain relatively small proportions of phytoceramide (<10% of total ceramides) (Dasgupta et al., 2013) but larger amounts of ceramide (800±150 pmol/mg cells), with C18:0 ceramide (35%) and C24:1 ceramide (25%) being the most abundant (Wang et al., 2012). Based on the observation that SPT2 is expressed in Chlamydomonas as well as mammalian cells, we tested the effect of myriocin, a fungal toxin and highly selective inhibitor of SPT by covalent binding to SPT2 (Wadsworth et al., 2013), on (phyto)ceramide biosynthesis. Figure 2D shows that myriocin is an extremely potent inhibitor of phytoceramide biosynthesis (>90% at 5 nM) in Chlamydomonas, suggesting that myriocin can be effectively used to test the function of phytoceramide in Chlamydomonas.
Phytoceramide and ceramide are critical for motility and ciliary length regulation Based on our previous studies showing that ceramide is critical for primary ciliogenesis (Wang et al., 2009a; He et al., 2012; He et al., 2014), we tested if a similar function exists for the regulation of flagella in Chlamydomonas and motile cilia in ependymal cells. To determine the effect of inhibition of de novo sphingolipid biosynthesis by myriocin on flagella, we first performed a phototaxis motility assay (Lechtreck et al., 2009). Following incubation of Chlamydomonas in a 6-well dish with 0-50 nM myriocin, half of each well was shielded from light. At 7 hours, most of the algae were still motile and swimming towards light. By 15 hours, only the algae not exposed to myriocin were fully motile (Figure 3A). This was confirmed both by cell counting with a hemocytometer and by measuring optical absorbance of chlorophyll at 450 nm. At a concentration of myriocin as low as 5 nM, approximately half of the cells were immotile (Figure 3B). At 10 nM myriocin, phase contrast microscopy showed that >70% of cells had extremely shortened (<2 µm length) flagella (Figure 3C) and often formed multicellular clusters (Figure S2A). Only a small population of cells (<20%) did not show significant flagella shortening. The inhibitory effect of myriocin on cilium length and motility was reversible since myriocin-treated cells regained motility within 48 h when resuspended in myriocin-free medium (Figure S2B). This result was consistent with a trypan blue exclusion assay showing that myriocin-treated cells did not undergo cell death (Figure S2C), further suggesting that the effect of myriocin on motility was specific for ciliogenesis and not due to general toxicity. To test if the effect of myriocin was caused by reduced synthesis of sphingolipids, we performed motility rescue experiments by supplementing the medium with various sphingolipid precursors 2 h prior to myriocin exposure. Among the sphingolipids tested, dihydrosphingosine (DHS), a metabolic precursor for (phyto)ceramide biosynthesis (Figure 1), and phytoceramide itself were the most effective (60±10% and 42±12%, respectively) in rescuing flagella length and motility (Figure 3D and E). Sphingosine and various ceramides were not effective for restoration of cilia (not shown). Fumonisin B1, a ceramide synthase inhibitor, failed to reduce motility in Chlamydomonas. This inhibitor has a higher dissociation constant than myriocin (100 nM vs. 0.28 nM) and in the case of lower uptake, the intracellular concentration of fumonisin B1 may not allow for effective inhibition of phytoceramide biosynthesis (Wang et al., 1991; Miyake et al., 1995). In ependymal cells, both myriocin and fumonisin B1 (FB1; Figure 1) reduced the length of motile cilia (Figure 3G). Addition of C24:1 ceramide to the culture medium rescued cilia in cells treated with FB1 and induced elongation of cilia in cells not treated with ceramide biosynthesis inhibitors (Figure 3F and G). In addition to FB1-treated wild type ependymal cells, we used cells from the fragilitis ossium (fro/fro) mouse, a mouse model for genetic deficiency of nSMase2 (Guenet et al., 1981; Poirier et al., 2012). In these cells, lack of nSMase2-catalyzed ceramide generation led to shortened cilia, which was rescued by exogenous C24:1ceramide (Figure S3). Other ceramides such as C18:0 ceramide were not effective in rescuing cilia in ependymal cells (not shown). These results suggest that phytoceramides in Chlamydomonas and C24:1 ceramide in ependymal cells are critical for cilium length regulation, which is likely to rely on a conserved downstream mechanism.
Phytoceramide and ceramide are enriched in compartments that regulate ciliogenesis Any effect of (phyto)ceramide on cilium regulation is likely to require the physical interaction of (phyto)ceramide with proteins in compartments that are known to regulate cilia, in particular vesicles and membranous compartments at the cilium base and the ciliary membrane. We previously generated a ceramide-specific antibody that detected an apical ceramide-enriched compartment (ACEC) at the base of primary cilia (Wang et al., 2009a; Bieberich, 2011; He et al., 2012; He et al., 2014).To test (phyto)ceramide distribution in Chlamydomonas and ependymal cells, we have generated a new antibody that reacts with both ceramide and phytoceramide as shown by lipid ELISAs (Figure 4A; data shown are background subtracted; (He et al., 2014; Dinkins et al., 2015). This antibody does not react with fatty acid (nervonic acid was tested), but it recognizes sphingosine and phytosphingosine, which are only minor cellular sphingolipids (≤10% of (phyto)ceramide) under physiological conditions. Immunocytochemistry using this antibody showed punctate labeling for (phyto)ceramide at tips of flagella and motile cilia (Figure 4B). In addition, (phyto)ceramide was enriched in vesicles or a compartment at the bases of flagella and motile cilia (Figure 4B).
We next tested if this compartment supplies (phyto)ceramide for the ciliary membrane. We used Chlamydomonas because it can be subjected to several cycles of de- and reflagellation where functionality is noted by return of motility (Wilson and Lefebvre, 2004). Chlamydomonas was 4-times deflagellated at pH 4.5 followed by rapid neutralization and reflagellation in the absence or presence of myriocin. With each de/reflagellation cycle, fewer cells retained motility. After 4 cycles, myriocin-treated cells completely lost motility while control cells were not affected (Figure 4C). In contrast to overnight incubation (Figure 3A), continuous myriocin incubation for 5 h of cells not undergoing de/reflagellation did not show loss of motility. Immunocytochemistry using the anti-(phyto)ceramide antibody showed that loss of motility in myriocin-treated cells undergoing repeated de/reflagellation was concurrent with the inability to regrow flagella and disappearance of the phytoceramide-enriched compartment at the flagella base (Figure 4D). This result suggests that the phytoceramide-enriched compartment in the cell body is dependent on de novo phytoceramide biosynthesis and supplies phytoceramide for the flagellar membrane. In the presence of myriocin, phytoceramide transported into the flagellar membrane and lost by repeated deflagellation can no longer be replenished from the cell body and flagella are no longer formed.
To test if phytoceramide is cotransported with proteins from the base into flagella, we used a bifunctional ceramide analog pacFA Ceramide (pacFACer; Figure S1A) to covalently label protein interaction partners of phytoceramide in the compartment at the flagella base and the flagellar membrane. After UV cross-linking, the pacFACer-protein complex was visualized by covalent linking to Alexa Fluor 594 azide using click chemistry. Alexa Fluor 594-labeled pacFACer was enriched in the base and tip of flagella (Figure 4E), consistent with the results obtained with anti-(phyto)ceramide antibody (Figure 4B). Next, we incubated Chlamydomonas with pacFACer followed by UV cross-linking and one cycle of de/reflagellation in the presence of myriocin before labeling of the pacFACer-protein complex with Alexa Fluor 594 azide. A proportion of the cells (15±5%) formed single very long (20±3 µm) flagella approximately double the length of normal flagella, which was not observed in the absence of UV cross-linking (Figure 4F). In these cells, the ceramide-enriched compartment at the flagella base was absent, consistent with the results obtained from repeated de/reflagellation (Figure 4D). The formation of a single, elongated flagellum labeled with pacFACer suggests that the pacFACer-protein complex is cross-linked at the flagella base and is then transported into the newly formed flagellum. Failure to dissociate after initial binding and transport due to covalent cross-linking leads to irreversible membrane association and excess cilium extension, similar to the phenotype of Chlamydomonas mutants with defective protein kinases involved in cilium length regulation (Berman et al., 2003; Wilson and Lefebvre, 2004; Hilton et al., 2013). These results suggest that (phyto)ceramide at the cilium base transiently interacts with proteins, in particular protein kinases that are critical for cilium length regulation.
Motile ciliogenesis in Chlamydomonas and ependymal cells relies on active GSK-3β One of the protein kinases that is highly conserved from Chlamydomonas to mammals and critical for ciliogenesis is GSK-3β. In Chlamydomonas, whose genome encodes for a single GSK3 isoform, activated pYGSK3 has been shown to be enriched in flagella and to regulate flagellar length (Wilson and Lefebvre, 2004). We performed immunoblotting of proteins from myriocin-treated Chlamydomonas and found a reduction of pYGSK3 levels, which was prevented by exogenous DHS, the phytoceramide precursor (Figure 5A). Consistently, acetylated tubulin was also reduced by myriocin and rescued by DHS, supporting the hypothesis that pYGSK3 levels are correlated with tubulin acetylation in flagella. In addition to pYGSK3, total GSK3 levels were reduced by myriocin and rescued by DHS, suggesting that ceramide depletion leads to decreased expression or increased degradation of Chlamydomonas GSK3. To show the direct effect of GSK3 activity on flagella and cilia we incubated Chlamydomonas and ependymal cells with BIO ((2’Z,3’E)-6-bromoindirubin-3′-oxime), a highly selective GSK3 inhibitor (Sato et al., 2004), for 60 min. GSK3 inhibition led to immotility and flagellar length reduction (Figure 5B) similar to that observed with myriocin (Figure 3C), which is consistent with the hypothesis that phytoceramide and ceramide are critical for activation of GSK3 to regulate cilium length. Lithium chloride (LiCl), a non-competitive GSK3 inhibitor also led to immotility, although flagella were first elongated (after 2h incubation, Figure 5B) and then lost (after 24 h, not shown) consistent with previously published studies (Wilson and Lefebvre, 2004).
The enrichment of pYGSK3 in flagella indicates that it may be colocalized with phytoceramide and ceramide in Chlamydomonas and ependymal cells, respectively. We found that pYGSK3 was colocalized with phytoceramide in the flagellar membrane, particularly in the flagella tips (Figure 5C). In ependymal cells, cilia were colabeled for pYGSK3 and ceramide (Figure 5D). Interestingly, we found that while pYGSK3 colocalized with acetylated tubulin in cilia (Figure 5E, left panel) inactive GSK3 phosphorylated at ser9 (pSGSK) was not found in cilia (Figure 5E, right panel). This result suggested that pYGSK3 may bind directly to ceramide in the ciliary membrane. A lipid ELISA-based binding assay using recombinant human GSK-3β expressed in Sf9 cells confirmed that GSK3 binds to different ceramide species (Figure 5F). Affinity of human GSK3 was highest to C24:1 ceramide, which was able to rescue cilia in ceramide-depleted ependymal cells (Figure 3F and G). Immunoblots showed that human GSK3 was phosphorylated at tyr216, consistent with the hypothesis that pYGSK3 binds directly to ceramide (Figure 5G).
Immunoblots of ependymal cells from wild type and fro/fro mice showed reduced pYGSK3 and increased pSGSK3 in fro/fro cells, suggesting that reduced cellular ceramide leads to less activation of GSK3 (Figure 6A), concurrent with reduced cilium length and diminished labeling of pYGSK3 in motile cilia (Figure 6B and Figure S3). Immunohistochemistry on cryosections of fro/fro brains showed that the length of motile cilia in the ependyma was also reduced by more than 50% (Figure 6C and D). Motile cilia in the ependyma showed colocalization of ceramide with pYGSK3, predominantly in punctate structures along the membrane and cilium tip (Figure 6E and F), which was consistent with the results obtained with primary cultured ependymal cells (Figure 6B and C). Taken together, these results suggest that phytoceramide and ceramide may induce activation and translocation of pYGSK3 into flagella and cilia, which is instrumental for the regulation of their length. Ceramide interaction with aPKCζ regulation of their length.
Ceramide interaction with aPKCζ regulates ciliogenesis in ependymal cells Our previous studies showed that atypical PKCδ (aPKCδ), another protein kinase that directly binds to ceramide regulates primary ciliogenesis in mammalian cells (Lozano et al., 1994; Muller et al., 1995; Wang et al., 1999; Bieberich et al., 2000; Bourbon et al., 2000; Fan et al., 2004; Ossipova et al., 2007; Pruliere et al., 2011; He et al., 2012; He et al., 2014). GSK-3β can be inactivated by aPKCδ-mediated phosphorylation of Ser9 (pSGSK3), suggesting that mammalian ciliogenesis can be regulated by binding of ceramide to both, GSK-3β and aPKCδ (Etienne-Manneville and Hall, 2003; Kim et al., 2007; Krishnamurthy et al., 2007b). Genome analysis shows that plants and Chlamydomonas do not express a homolog of aPKCδ and GSK3 does not contain an equivalent to the Ser9 phosphorylation site found in mammalian GSK-3β (Kruse et al., 1996; Wilson and Lefebvre, 2004). Therefore, we limited the analysis motile cilia regulation by ceamide and aPKCδ to ependymal cells. An orthogonal view of a confocal z-scan shows that UV cross-linked pacFACer colocalizes with aPKCδ in the ceramide-enriched compartment at the cilium base (Figure 7A). The physical interaction of pacFACer with aPKCδ was demonstrated by a lipid ELISA-based assay using a surface coat of pacFACer for binding to recombinant aPKCδ. Binding was enhanced by UV cross-linking, suggesting that aPKCδ could be covalently linked to pacFACer and is indicative of high affinity for ceramide. The reaction did not occur in the absence of pacFACer (pCer), aPKCδ, or aPKCδ-specific antibody (Figure 7B). The ceramide binding affinity of aPKCδ was tested using lipid-ELISA with surface coats of ceramides differing in the chain length and degree of saturation of the fatty acid moiety. Affinity of aPKCδ was higher for ceramides with very long chain and unsaturated fatty acids, with highest affinity for C24:1 ceramide (Figure 7C), similar to the affinity found with GSK-3β (Fig. 5F). This result is consistent with rescue of cilia length in myriocin-treated ependymal cells and elongation of cilia in untreated cells by C24:1 ceramide.
We tested whether aPKCδ activation or inhibition is critical for ceramide-regulated motile ciliogenesis. Incubation of ependymal cells with fumonisin B1 (FB1), a ceramide synthase inhibitor, led to reduction of motile cilium length (Figure 7D). The cell-permeable myristoylated aPKCδ pseudosubstrate inhibitor (PZI) prevented reduction of motile cilium length by FB1, suggesting that ceramide-mediated inhibition of aPKCδ is critical for motile ciliogenesis (Figure 7D). Consistent with this hypothesis, addition of PZI to cells not treated with FB1 resulted in increase of cilium length by 50-60% (Figure 7D). Taken together, these results suggest that in ependymal cells, ceramide binds and inhibits aPKCδ, which leads to increase of cilium length. However, since co-localization of aPKCδ with ceramide was only detected at the cilium base, but not in the ciliary membrane (Figure 7A), we concluded that binding of aPKCδ to ceramide at the cilium base affects a secondary target that regulates cilium length by its translocation into cilia. Consistent with the observation that GSK3 is critical for flagella formation and a target for mammalian aPKCδ, we found that inhibition of GSK3 with BIO obliterated motile cilia in ependymal cells (Figure 7D). Therefore, we hypothesize that GSK3 is this secondary target by being phosphorylated at Ser9 and inactivated by aPKCδ, unless aPKCδ is bound to ceramide at the cilium base and sequestered from GSK3 that is translocated to the ciliary membrane.
Discussion
Length regulation of primary and motile cilia and flagella is essential for their biological function. To date, there are more than 20 proteins known to regulate cilium length, among which many are protein kinases (e.g., Aurora A kinase (AurA), MAPK/MAK/MRK overlapping kinase or MOK/RAGE1), IFT proteins, and enzymes involved in tubulin modification (e.g., HDAC6) (Pan et al., 2004; Pugacheva et al., 2007; Cao et al., 2013; Broekhuis et al., 2014). Little is known about the dynamic role of lipids in ciliogenesis. Since the cell membrane cannot be stretched any cilium length extension is inevitably accompanied by net synthesis and transport of membrane lipids, by either vesicle traffic or lateral movement of lipids within the cell membrane toward the cilium base. While new membrane is added at the cilium base, tubulin and other proteins for cilium extension are transported along the cilium and then added at the tip. It is not clear how membrane lipid and protein transport are coregulated to ensure that elongation of the axoneme is adjusted to the adequate expansion of the ciliary membrane. Our study presents new evidence that the sphingolipid (phyto)ceramide is instrumental for length regulation of flagella and cilia based on data from the green algae Chlamydomonas flagella and murine ependymal cells.
Chlamydomonas is a well-established model to study regulation of motile cilium length by testing mechanisms that affect flagella growth and regeneration. It has been shown that the flagellar membrane is enriched with phosphatidylethanolamine, which can be hydrolyzed by phospholipase D (PLD) to phosphatidic acid (PA), a lipid known to induce deflagellation (Quarmby et al., 1992; Goedhart and Gadella, 2004; Lechtreck et al., 2013). However, it is not clear if PLD-mediated generation of PA is part of a process regulating flagella length. It is also not known whether Chlamydomonas expresses enzymes that generate sphingolipids and whether these lipids, if present, are essential for regulation of flagella. Using RT-PCR and immunoblotting we showed that SPT, the first enzyme in de novo sphingolipid biosynthesis, in particular its subunit SPT2, is conserved and expressed in Chlamydomonas. LC-MS/MS confirmed that C26:0 and C28:0 phytoceramide are generated, suggesting that Chlamydomonas expresses a complete set of enzymes for de novo sphingolipid biosynthesis. While the analysis of the complete set of enzymes in (phyto)ceramide biosynthesis is part of our future studies, we could show that Chlamydomonas expresses the mRNAs of two putative ceramide synthases, lag1 and lag2. Among the enzyme inhibitors tested, the SPT inhibitor myriocin blocked phytoceramide biosynthesis and led to flagella length reduction and immotility. The specificity of this effect was demonstrated by the recovery of flagella growth and motility after removal of myriocin and by rescuing flagella with exogenous DHS, a downstream product of SPT and precursor for phytoceramide. Flagella were also partially rescued by exogenous phytoceramide, suggesting that flagella length is regulated by DHS that is metabolically converted to phytoceramide. Exogenous ceramide did not rescue flagella in Chlamydomonas, but it prevented loss of motile cilia in ependymal cells treated with inhibitors of ceramide biosynthesis. These results demonstrate that phytoceramide and ceramide are ciliogenic sphingolipids that regulate cilium length in Chlamydomonas and ependymal cells, respectively.
In animals, ceramide is synthesized in the endoplasmic reticulum (ER), transported to the Golgi, where it can be converted into glycosphingolipids or sphingomyelin, which are further transported to the cell membrane. Sphingomyelin can be internalized by endocytosis and hydrolyzed to ceramide by sphingomyelinases (SMases). Our previous studies have shown that SMases in mammalian cells generate ciliogenic ceramide that is accumulated in an apical ceramide-enriched compartment (ACEC) at the base of primary cilia (Wang et al., 2009a; He et al., 2012; He et al., 2014). In plants and yeast, phytoceramide is also synthesized in the ER and derivatized to complex phytosphingolipids in the Golgi. However, plants and algae do not contain sphingomyelin, but instead they generate inositolphosphorylsphingolipids (Lester and Dickson, 1993; Sperling and Heinz, 2003; Markham et al., 2013). Therefore, phytoceramide in Chlamydomonas is likely to be generated at the flagella base, either by de novo biosynthesis or degradation of complex phytosphingolipids, and then further transported into the ciliary membrane. This transport was tested using two techniques: labeling of phytoceramide and ceramide with an antibody and direct fluorescent labeling of the bifunctional ceramide analog pacFACer after UV cross-linking to interacting protein(s). Anti-ceramide IgG was originally generated in our laboratory and has been extensively tested in independent laboratories using immunocytochemistry (Krishnamurthy et al., 2007a; Wang et al., 2009a; Muscoli et al., 2010; He et al., 2012; He et al., 2014). Anti-ceramide IgG was found to detect ceramide and phytoceramide in lipid ELISAs (He et al., 2014; Dinkins et al., 2015). Using this antibody, we showed that phytoceramide and ceramide are enriched at the base and the tip of flagella and motile cilia in Chlamydomonas and ependymal cells, respectively. These results suggest that (phyto)ceramide is transported from the ciliary base to the tip.
Figure 8A shows a model consistent with a by co-transport of (phyto)ceramide and flagella/cilia length-regulating proteins. In this “flux equilibrium model”, lipid vesicles from the ceramide compartment are incorporated into the ciliary membrane at the cilium base. Lipidcargo protein co-transport ensures the stoichiometry required for simultaneous cilium elongation and membrane expansion. (Phyto)ceramide may serve as a membrane anchor for co-transported cargo proteins, or alternatively, it may activate a loading/unloading mechanism at the base or tip of the cilium. The cilium length is regulated by the size or ceramide content of the compartment at the base and its lipid flux to the cilium: more ceramide favors cilium assembly, less favors disassembly until flux rates in both directions are equal and the cilium length is maintained. Consistent with this model, repeated de- and reflagellation in the presence of myriocin led to the inability to regenerate flagella, indicating that phytoceramide is continuously incorporated into the ciliary membrane and replenished by de novo biosynthesis. Appearance of fluorescently labeled pacFACer in newly formed flagella was concurrent with disappearance of labeling at the base, further supporting the hypothesis that (phyto)ceramide is transported from the base into the ciliary membrane.
Any specific regulation of cilia by (phyto)ceramide requires its interaction with proteins involved in ciliogenesis. It has been shown that several evolutionary conserved kinases with mammalian homologs such as Chlamydomonas AurA-like kinase (CALK), LF4p (MOK in mammals), and GSK3 are important for flagella length regulation (Silflow and Lefebvre, 2001; Berman et al., 2003; Pan et al., 2004; Wilson and Lefebvre, 2004; Thoma et al., 2007; Cao et al., 2009; Ou et al., 2009; Cao et al., 2013; Hilton et al., 2013; Broekhuis et al., 2014). GSK3 is of particular importance since its role in ciliogenesis has been confirmed in numerous studies, but it is unclear if cilium length extension is induced by activation or inhibition of GSK3, or both. Lithium chloride (LiCl), a non-competitive inhibitor of GSK3, has been shown to induce cilium extension in Chlamydomonas and mammalian cells (Berman et al., 2003; Nakakura et al., 2014). However, LiCl is not specific and requires mM concentrations for effectiveness, and other studies have shown that its effect on ciliogenesis is independent of GSK3 (Ou et al., 2009). Specific GSK3 inhibitors have not been tested in Chlamydomonas, and they have given inconsistent results when used to test their effect on mammalian ciliogenesis (Ou et al., 2009; Wang et al., 2009a). We administered two specific GSK3 inhibitors, BIO and indirubin-3- monoxime, to Chlamydomonas and found that they lead to complete shortening of flagella within 60 min, consistent with their effect on ependymal cells. Therefore, we conclude that active GSK3 is critical for maintenance or extension of cilia. This conclusion is consistent with previous studies showing that active pYGSK3 is translocated into flagella and that GSK3 knockdown leads to flagella shortening (Wilson and Lefebvre, 2004). It is also consistent with studies showing that specific inhibition or knockdown of GSK3 impairs ciliogenesis in mammalian cells (Ou et al., 2009). The critical role of pYGSK3 for ciliogenesis was further substantiated by our observation that pYGSK3 is colocalized with phytoceramide and ceramide in Chlamydomonas flagella and ependymal cell motile cilia, respectively. In ependymal cells, nSMase2 deficiency led to decreased levels of pYGSK and increased levels of pSGSK, clearly showing that nSMase2-mediated generation of ceramide is instrumental for increasing pYGSK3 and reducing pSGSK levels.
Activation of GSK3 by autophosphorylation of tyr216 is a cotranslational process, while inactivation by desphosphorylation at this residue and phosphorylation of ser9 is posttranslational and regulated by phosphatases and kinases, respectively (Beurel et al., 2015). Since nSMase2-deficient ependymal cells show reduction of tyr216 and increase of ser9 phosphorylation, ceramide is likely to affect phosphatases and kinases that regulate GSK3 phosphorylation. We and other laboratories have found that ceramide binds and sequesters aPKCδ (Lozano et al., 1994; Muller et al., 1995; Wang et al., 1999; Bieberich et al., 2000; Bourbon et al., 2000; Wang et al., 2005; Fox et al., 2007; Wang et al., 2009b), which was confirmed by lipid-ELISAs and UV cross-linking to pacFACer. Ceramide-induced sequestration may prevent aPKCδ-mediated phosphorylation of GSK3 at ser9 and lead to enrichment of pYGSK3 in the cilium tip (Figure 8B). The hypothesis that ceramide-mediated sequestration of aPKCδ is critical for cilium length extension is supported by the observation that inhibition of aPKCδ with PZI promotes ciliogenesis in ependymal cells. Ceramide depletion prevents sequestration of aPKCδ and enrichment of pYGSK3 in cilia, while it promotes phosphorylation of GSK3 at ser9 and elevation of pSGSK3 levels (Figure 6A). Neither aPKCδ nor pSGSK3 are transported into cilia, suggesting that ciliary pYGSK3 is critical for cilium length regulation.
In Chlamydomonas, pYGSK3 levels and transport into flagella are elevated by phytoceramide, but the interaction partner is currently unknown as plants and algae do not possess aPKC homologs (Kruse et al., 1996). Moreover, Chlamydomonas GSK3 does not have a phosphorylation site homologous to ser9 in mammalian GSK-3β (Wilson and Lefebvre, 2004). Therefore, phytoceramide may sustain pYGSK3 levels and its enrichment into flagella by a different mechanism. Our data showing that phytoceramide is colocalized with pYGSK3in flagella strongly suggests that phytoceramide may regulate IFT of pYGSK3 (Figure 8B). Likewise, colocalization of ceramide with pYGSK3 in motile cilia suggests that ceramide may interact directly with pYGSK3 within the ciliary membrane and promote its transport to the tip. Support for this hypothesis comes from lipid-based ELISAs showing that recombinant human GSK-3β binds to C24:1 ceramide, the ceramide species rescuing motile cilia in ceramide depleted ependymal cells. Potential binding to pYGSK3 may be the evolutionarily older mechanism by which (phyto)ceramide regulates flagella/cilia, while ceramide-mediated sequestration of aPKC may have developed more recently as complementary mechanism of GSK3 activation in animals. It is currently unknown how the interaction of phytoceramide or ceramide with pYGSK3 in the cilium tip may regulate cilium length extension. It is possible that pYGSK downregulates HDAC6, thereby preserving tubulin acetylation (Figure 8B). Support for this hypothesis comes from a previous study showing that dysfunctional GSK3 leads to upregulation of HDAC6 and ciliary defects (Song et al., 2014). It is also possible that pYGSK does not directly regulate tubulin acetylation but instead cargo release at the cilium tip. It has been shown that GSK3- mediated phosphorylation of kinesin light chain leads to release of transport vesicles in neuronal axons (Szebenyi et al., 2002). It is likely that a similar mechanism may induce phosphorylation of kinesin 2 in cilia, which then allows for release of proteins that are critical for cilium assembly at the tip. The interaction of phytoceramide and ceramide with pYGSK at the cilium tip may trigger this cargo release and promote ciliogenesis.
In summary, our study shows that (phyto)ceramide is critical for the regulation of flagella in Chlamydomonas and motile cilia in ependymal cells in vitro and in vivo. We present new evidence that nSMase2-deficiency results in shortening of motile cilia in the ependyma, suggesting that reduction of ceramide generation leads to ciliary dysregulation in vivo. While it has not been described that mutations of enzymes in ceramide metabolism are linked to ciliopathies, our study shows that (phyto)ceramide levels and composition are important for adjusting cilium length. Therefore, (phyto)ceramide is the first signaling sphingolipid demonstrated to metabolically regulate ciliogenesis by an evolutionarily conserved mechanism in Chlamydomonas and mammalian cells.
RESEARCHERS VIDEO AND MEASURE TUBULIN TRANSPORT IN CILIA FOR THE FIRST TIME.
http://health-innovations.org/2015/01/27/researchers-image-and-measure-tubulin-transport-in-cilia/
Defective cilia can lead to a host of diseases and conditions in the human body, from rare, inherited bone malformations to blindness, male infertility, kidney disease and obesity. It is known that these tiny cell organelles become deformed and cause these diseases because of a problem related to their assembly, which requires the translocation of vast quantities of the vital cell protein tubulin. What they didn’t know was how tubulin and another cell organelle known as flagella fit into the process.
Now, a new study from University of Georgia shows the mechanism behind tubulin transport and its assembly into cilia, including the first video imagery of the process. The study was published in the Journal of Cell Biology.
Cilia are found throughout the body, so defects in cilia formation affect cells that line airways, brain ventricles or the reproductive track. One of the main causes of male infertility is the cilia won’t function properly.
The team used total internal reflection fluorescence microscopy to analyze moving protein particles inside the cilia of Chlamydomonas reinhardtii, a green alga widely used as a model for cilia analysis.
The team exploited the natural behaviour of the organism, which is to attach by its cilia to a smooth surface, such as a microscope glass cover. This positions the cilia within the 200-nanometer reach of the total internal reflection fluorescence microscope allowing for the imaging of individual proteins as they move inside the cilia. A video explaining the process was published along with the study.
Tubulin is transported by this process called intraflagellar transport, or IFT. Though it has long been suspected in the field and there was indirect evidence to support the theory, this is the first time it has been shown directly, through live imaging, that IFT does function as a tubulin pump. The team observed that about 400,000 tubulin dimers need to be transported within 60 minutes to assemble a single cilium. Being able to see tubulin moving into cilia allowed for first insights into how this transport is regulated to make sure cilia will have the correct size.
The new findings are expected to have wide implications for a variety of diseases and conditions related to cilia defects in the body. The team state that they are on the very basic side of this research. But because more and more diseases are being connected to cilia-related conditions, including obesity and even diabetes, the number of people working on cilia has greatly expanded over the last few years.
The team summise that unicellular models are great tools to address the many fundamental questions which remain unanswered such as controlling the size of cilia and how cells determine the proteins to be placed into these sensory organelles. In Chlamydomonas, the researchers were able to initiate cilia formation experimentally, which allowed them to see cilia during construction and analyze protein traffic while they are made.
Source: UGA Public Affairs Division

Repeated FRAP of growing and nongrowing cilia is followed by similar rates of recovery. (A and B) Segments of a steady-state (A) and a re-generating (B) cilium were repeatedly bleached (indicated by arrowheads). Kymograms (top) and FRAP quantification (bottom) indicate similar rates (in percentage of pre-bleach GFP–a-tubulin fluorescence) of recovery after each bleaching step. (C and D) Individual frames (C) and kymograms (D) of a long-short cell. Bleached areas are marked by dashed circles. The kymogram (D) is a composite of several recordings, and arrowheads labeled a–e indicate the positions of the frames in C. The time (in seconds) for each recording and the position the bleaching laser (Brackets) is indicated; overexposed frames caused by photobleaching were deleted. Arrows in D: GFP–a-tubulin trajectories. Note fast and strong recovery in subsequent bleachings of the short cilium while the initially bleached area in the long cilium remains visible. The extended observation time will also bleach some of the (axonemal) GFP–a-tubulin outside of the spot bleaching area. This loss of fluorescence in the nonbleached areas results in a higher apparent recovery in the bleached areas. Tubulin transport by IFT is upregulated during ciliary growth by a cilium-autonomous mechanism. Lechtreck et al 2015.
Tubulin transport by IFT is upregulated during ciliary growth by a cilium-autonomous mechanism
Julie M. Craft,1, J. Aaron Harris,1, Sebastian Hyman,1, Peter Kner,2 and Karl F. Lechtreck1
JCB Jan 2015; 208(2): 223-237 Craft et al. 208 (2): 223 http://dx.doi.org:/10.1083/jcb.201409036
The assembly of the axoneme, the structural scaffold of cilia and flagella, requires translocation of a vast quantity of tubulin into the growing cilium, but the mechanisms that regulate the targeting, quantity, and timing of tubulin transport are largely unknown. InChlamydomonas, GFP-tagged α-tubulin enters cilia as an intraflagellar transport (IFT) cargo and by diffusion. IFT-based transport of GFP-tubulin is elevated in growing cilia and IFT trains carry more tubulin. Cells possessing both nongrowing and growing cilia selectively target GFP-tubulin into the latter. The preferential delivery of tubulin boosts the concentration of soluble tubulin in the matrix of growing versus steady-state cilia. Cilia length mutants show abnormal kinetics of tubulin transport. We propose that cells regulate the extent of occupancy of IFT trains by tubulin cargoes. During ciliary growth, IFT concentrates soluble tubulin in cilia and thereby promotes elongation of the axonemal microtubules.
Microtubules, polymers of α- and β-tubulin dimers, are the major structural element of motile and nonmotile cilia and flagella (Borisy and Taylor, 1967). Microtubules form the scaffold onto which other axonemal structures such as dynein arms and radial spokes are attached; they also serve as tracks for intraflagellar transport (IFT), a bidirectional motility which is required for ciliary assembly (Kozminski et al., 1993, 1995). During ciliary growth, large amounts of tubulin dimers need to be transported into the organelle. Like cytoplasmic microtubules, the axonemal microtubules grow through the addition of tubulin subunits to the distal plus-ends (Witman, 1975; Euteneuer and McIntosh, 1981; Johnson and Rosenbaum, 1992). Thus, the delivery of tubulin into cilia and its translocation to the ciliary tip are prerequisites for ciliary assembly.
Several lines of evidence suggest that inside cilia, tubulin is transported by IFT. In the conditionalChlamydomonas fla10-1 mutant, the incorporation of epitope-tagged tubulin at the distal end of cilia is decreased after a shutdown of IFT (Marshall and Rosenbaum, 2001). In vitro, tubulin dimers directly interact with a tubulin-binding module formed by the IFT particle proteins IFT74 and IFT81 (Bhogaraju et al., 2013). Expression of fluorescent tubulin in Caenorhabditis elegans revealed weak tracks, possibly representing tubulin transport in sensory cilia, and FRAP analysis supported the notion that tubulin transport is IFT-dependent (Hao et al., 2011). It remains largely unknown how tubulin transport is coordinated with ciliary growth. Insufficient delivery of tubulin could result in slower growth and shorter cilia, whereas an excess of tubulin might promote the assembly of excessively long cilia. Defects in ciliary length impair the motile and sensory functions of cilia and have been linked to ciliary diseases (Mahjoub et al., 2005; Tammachote et al., 2009; Ko et al., 2010; Özgül et al., 2011; Mahjoub and Stearns, 2012; Tam et al., 2013).
In the unicellular alga Chlamydomonas reinhardtii, IFT and protein transport inside cilia can be imaged with single particle sensitivity using total internal reflection fluorescence (TIRF) microscopy (Lechtreck, 2013;Wren et al., 2013). Recently, stable expression of a fluorescent protein-tagged α-tubulin was accomplished inC. reinhardtii (Rasala et al., 2013). Ciliogenesis can be experimentally induced by removing the existing cilia (Rosenbaum et al., 1969), allowing for an in-depth study of the relationship between tubulin transport and ciliary growth using direct imaging. A genetic analysis of the regulation of tubulin transport is aided by numerous C. reinhardtii mutants with defects in ciliary assembly and length control.
Here, using two-color imaging, we show that GFP–α-tubulin is primarily transported by IFT but some GFP-tubulin also enters cilia by diffusion. The frequency of tubulin transport events by anterograde IFT was greatly increased during ciliary growth. We find that C. reinhardtii cells possess the ability to preferentially direct IFT-bound tubulin into growing cilia, indicating that tubulin transport is regulated in a cilium-autonomous manner. Dysregulation of tubulin transport was observed in the cilia length mutants short flagella2 and long flagella2-1, indicating a possible link between the ciliary length regulation and tubulin transport by IFT. During ciliary growth, the concentration of soluble tubulin in the ciliary matrix was elevated substantially above that of steady-state cilia. We propose a model in which cells use IFT to regulate the concentration of soluble tubulin inside cilia; a high concentration of tubulin in the matrix will promote microtubule polymerization and ciliary growth.
To express GFP–α-tubulin in C. reinhardtii for imaging in vivo, we used the recently described fusion construct consisting of the zeocin resistance gene BLE and the TUA2 gene separated by the viral 2A sequence encoding a self-cleaving peptide and replaced the original mCerulean with the brighter superfolder GFP (hereafter referred to as GFP; Rasala et al., 2013). In vivo microscopy revealed the presence of GFP–α-tubulin in cilia and cell body microtubules (Fig. 1 A; Rasala et al., 2013). Western blotting of whole cells with anti-GFP identified a major ∼78 kD and a minor ∼104 kD band that were both also recognized by anti–α-tubulin (Fig. 1 B). The former represents GFP–α-tubulin; the latter, largely excluded from cilia (Fig. 1 B), is the uncleaved ble-GFP–α-tubulin fusion protein. The expression levels of GFP–α-tubulin varied between individual transformants. In the strain GFP-Tub1, which was used in most experiments, GFP–α-tubulin amounted to ∼15% of the endogenous α-tubulin (Fig. 1 B). To determine the distribution of GFP-tagged and endogenous α-tubulin, cells were fractionated and analyzed by Western blotting (Fig. 1, B–D). Staining with anti–α-tubulin revealed that ∼80% of endogenous tubulin resides in the cell body and ∼20% enters the cilia; a similar partition was observed for GFP–α-tubulin with ∼90% in the cell body and ∼10% in cilia (Fig. 1, C and F). Inside steady-state cilia, both tagged and endogenous tubulin showed a similar distribution with ∼90% incorporated into the axoneme and <10% remaining soluble in the membrane+matrix fraction (MM fraction; Fig. 1, D and G). Immunoprecipitation using anti-GFP beads revealed that GFP–α-tubulin forms a complex with endogenous β-tubulin in the ciliary matrix (Fig. S1, A and B). Axonemal tubulin is known to undergo numerous posttranslational modifications (PTMs; Gaertig and Wloga, 2008). Western blots of fractionated cilia probed with PTM-specific antibodies showed that GFP–α-tubulin undergoes K40 acetylation and polyglutamylation, especially in the axoneme (Fig. 1 E and Fig. S1 C). In summary, GFP–α-tubulin behaves largely similar to the endogenous protein, validating its use as a reporter of tubulin transport.

To image GFP–α-tubulin during transport, cilia were first photobleached to eliminate the signal resulting from the presence of GFP–α-tubulin in the axonemes and then analyzed by TIRF microscopy. Kymograms (time-space plots) showed individual GFP–α-tubulin particles diffusing inside cilia or undergoing active transport with constant velocities indicative for IFT (Fig. 2 A; Videos 1 and 2). Anterograde transport progressed at 1.68 µm/s (±0.21 µm/s; n = 1,404 particles) and occurred with a frequency of 0.3 particles/min in steady-state cilia (± 0.7 particles/min, n = 78 cilia); retrograde transport events were rarely observed and had a velocity of 3.05 µm/s (±1.1 µm/s, n = 35; Fig. 2 A, a and b). IFT moved the majority of GFP–α-tubulin particles directly from the ciliary base to the tip (98% of 1,281 particles; Fig. 2 B, a); transitions of GFP–α-tubulin from IFT to diffusion indicative of unloading along the ciliary shaft were observed as well (Fig. S2, C and D). To test directly if tubulin moves in association with IFT trains, GFP–α-tubulin was expressed in anift20-1 IFT20-mCherry strain (Fig. S3 B). Two-color imaging showed comigration of the tagged tubulin and IFT20-mCherry indicating that tubulin is a bona fide cargo of IFT (Fig. 2 B and Video 3).

Tubulin enters cilia by IFT and diffusion. (A) Gallery of kymograms depicting GFP–α-tubulin (or, in c, mNeonGreen-tubulin) moving inside cilia by anterograde IFT (open arrow in a), retrograde IFT (open arrow in b), and diffusion (arrowheads in c and d). Anterograde transport results in trajectories running from the bottom left (ciliary base) to the top right (ciliary tip; T); retrograde transport events result in top-left to bottom-right trajectories. (c) Tubulin diffusing inside the ciliary shaft; (d) reduced mobility of GFP–α-tubulin in the vicinity of the tip (filled arrows). Bars, 1 µm and 1 s. (B) Kymograms from simultaneous imaging of mNeonGreen–α-tubulin (a and b) and IFT20-mCherry (c and d) in growing cilia; IFT-like trajectories are marked in b and d. A merged kymogram is shown in e. A Western blot of this strain is shown in Fig. S3 B. Bars, 5 µm and 5 s. (C) Mean square displacement versus time for 64 sfGFP–α-tubulin particles diffusing inside the shaft and 58 particles diffusing near the tip of steady-state cilia. The standard error of the mean at each value is indicated. A linear fit to the data at the short time points, which is likely to represent pure diffusion, results in diffusion coefficients of ∼1.8 µm2/s and ∼0.2 µm2/s for 1D diffusion of GFP-tubulin along the ciliary shaft and at the tip, respectively. (D) Still images and kymograms of fla10-1 cells expressing either GFP–α-tubulin (a and b) or IFT20-mCherry (c and d) at the permissive temperature (22°C; a and c) and after >180 min at 32°C (b and d). IFT-like trajectories for tubulin and IFT20 were only observed at 22°C (open arrows in a and c). Diffusion of GFP–α-tubulin (arrowhead in b) into photobleached cilia continued at 32°C in the absence of detectable IFT (d). Bars, 1 µm and 1 s. (E) Frequency of anterograde GFP–α-tubulin transport by IFT in fla10-1 and control cells (FLA10 ift20-1 IFT20 mCherry). Steady-state cilia and cilia regenerated in the presence of CHX were compared; at 32°C, IFT-like tubulin transport was robust in control cells but not observed in fla10-1 cells. fla10-1 regenerates cilia only slowly at room temperature (∼22°C); most measurements are based on cells regenerating cilia at 16°C. Error bars indicate SEM. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4298693/bin/JCB_201409036_Fig2.jpg
Recent data revealed that the acidic C-terminal domain (E-hook) of β-tubulin but not that of α-tubulin is required for the in vitro interaction between tubulin dimers and IFT74–IFT81 complexes (Bhogaraju et al., 2013). We designed modified GFP–α-tubulins in which the E-hook was replaced with the corresponding region of β-tubulin, altered to substitute seven glutamate residues with alanine or glutamine, or removed entirely (Table S1). The altered α-tubulin molecules were expressed (as ∼5–15% of the endogenous α-tubulin; Fig. S3 A), transported by IFT, and incorporated into the axoneme (Table S1). Thus, our in vivo observations agree with the earlier in vitro study showing that the C-terminal E-hook of α-tubulin is dispensable for tubulin binding to IFT trains. The modified tubulins were expressed in the presence of endogenous wild-type tubulin indicating that C. reinhardtii tolerates some E-hook–deficient GFP–α-tubulins; a deletion of the E-hook from all of either α- or β-tubulin is lethal in Tetrahymena (Duan and Gorovsky, 2002).
Tubulin enters cilia by diffusion
To improve imaging of diffusing tubulin, photobleached cilia were observed at elevated laser intensities which quickly bleached most GFP preventing crowding of the cilia with fluorescent α-tubulin. For some experiments, superfolder GFP was replaced with the brighter mNeonGreen (Shaner et al., 2013). 64 individual GFP–α-tubulin particles were tracked while diffusing inside the ciliary shaft and the mean of the square displacement was determined in 30 subsequent frames (Fig. 2 C). This resulted in a 1D diffusion coefficient of 1.76 ± 0.18 µm2/s for GFP–α-tubulin, a value similar to that determined for similarly sized soluble proteins diffusing in cilia (Lin et al., 2013). In agreement with this high mobility of GFP–α-tubulin, we observed “jumps” over almost the entire length of a cilium (Fig. 2 A, d; and Fig. S2, A, E, and I). In the vicinity of the ciliary tip, GFP-tagged tubulin mostly displayed a reduced mobility as reflected by a diffusion coefficient of ∼0.18 ± 0.02 µm2/s (n = 58 trajectories; Fig. 2 A, d; Fig. S2, A, C, D, and F, white arrowheads; and Video 4). The reduced mobility of GFP-tubulin in the distal ∼0.5-µm ciliary segment could result, for example, from interactions with other proteins. Infrequently, GFP–α-tubulin diffusing near the tip of steady-state cilia was observed becoming stationary, which is indicative of incorporation into the axoneme (Fig. S2, F–H).
Although IFT-based transport of tubulin was rare in steady-state cilia, diffusing GFP-tubulin continuously entered fully photobleached cilia from the base (Fig. 2 A, c; Fig. S2 I; and Video 2). To test whether active IFT is required for the diffusional entry of tubulin into cilia, we expressed GFP–α-tubulin in the C. reinhardtii fla10-1 mutant; IFT can be switched-off in this strain by incubating the cells at 32°C for several hours (Kozminski et al., 1995). IFT of GFP–α-tubulin was observed in fla10-1 and wild-type steady-state cilia at the permissive temperature albeit at low frequencies (Fig. 2, D, a, and E). Cycloheximide (CHX) limits ciliary regeneration to half-length and we previously observed elevated transport of the axonemal protein DRC4 in such cilia (Wren et al., 2013). Similarly, an increased frequency of tubulin transport was observed in half-length cilia assembled by control or fla10-1 in the presence of CHX at the permissive temperature (Fig. 2 E). After shifting such cells to 32°C for 180 min, IFT-like movements of GFP–α-tubulin were frequent in cilia of control cells but were not observed in the fla10-1 cells; entry of GFP–α-tubulin by diffusion, however, continued (Fig. 2, D, b, and E). A strain expressing IFT20-mCherry in the fla10-1 ift20-1background was used to verify that IFT was indeed abolished at the conditions used (Fig. 2 D, c and d). The data indicate that GFP–α-tubulin enters cilia by diffusion.
The frequency of tubulin transport by IFT is regulated by the assembly status of cilia
To determine whether IFT transports more tubulin when cilia grow, we compared the transport frequencies of GFP–α-tubulin between steady-state and growing cilia. After amputation of cilia by a pH shock, cells regenerate full-length cilia within ∼90 min. Partially regenerated cilia were bleached and tubulin influx and incorporation into the cilia were analyzed (Fig. 3 A and Video 5). Fluorescent tubulin was rapidly added to the tip and the fluorescent distal parts of cilia lengthened, revealing that cilia continue to grow under TIRF illumination while immobilized in the observation chamber (Fig. 3 B). In growing cilia (∼3–10 µm in length), GFP–α-tubulin moved by anterograde IFT with a greatly elevated mean frequency of 18.3 ± 6.9 particles/min (n = 110 cilia) versus ∼0.3 particles/min observed in steady-state cilia of the same strain (Fig. 3 C). The transport frequency of GFP–α-tubulin remained elevated until cilia reached ∼10 µm in length and was then reduced to 1 ± 2.2 particles/min in cilia of 10–12 µm in length (n = 49 cilia) before returning to the pre-deciliation level (∼0.3 ± 0.75 particles/min, n = 80 cilia at >2 h after pH shock; Fig. 3, C and D). The pattern indicates a strong length dependency of IFT-based tubulin transport and matches the deceleratory kinetics of cilia regeneration in C. reinhardtii with rapid elongation until cilia reach ∼10 µm, followed by slower growth until cilia reach their final length of ∼12 µm (Rosenbaum et al., 1969; Engel et al., 2009). In summary, tubulin transport by anterograde IFT is up-regulated during ciliary growth.

The observed increase in the transport of tubulin in growing cilia could be caused by an increase in the amount of tubulin carried by a given number of IFT trains or by an increase in the number of IFT trains without changing the tubulin load per train. In C. reinhardtii, similar amounts of IFT material are thought to be present in cilia regardless of their length (Marshall et al., 2005; Engel et al., 2009). Two-color imaging was used to determine the share of IFT particles participating in GFP-tubulin transport in regenerating and steady-state cilia (Fig. 3, E–G). Photobleaching of the entire cilia using increased 488-nm laser intensity also greatly diminished the signal from IFT20-mCherry making quantitative analyses unreliable. We therefore used a brief, focused laser beam in epifluorescence (∼3 µm diameter; ∼100–300 ms in duration) to bleach a ciliary segment and imaged the cilia in TIRF as particles reentered the bleached area (Fig. 3, E and F). IFT occupancy, i.e., the share of IFT20-mCherry–tagged anterograde IFT trains carrying GFP–α-tubulin, decreased from ∼80% during the initial rapid growth to ∼10% as cilia approached full length and even lower rates in fully regenerated and steady-state cilia (means were 45 ± 21% vs. 4.5 ± 3.5%, respectively; P = 2.2 × 10−8; Fig. 3 G). As previously shown, the IFT frequency was largely independent of ciliary length (Fig. S4 A; Dentler, 2005; Engel et al., 2009). In conclusion, IFT trains in regenerating cilia carry a larger load of tubulin than those in nongrowing cilia.
IFT transport of tubulin is regulated in a cilium-autonomous manner
How do cells regulate the loading of cargo onto IFT particles? To begin addressing this question, we tested whether tubulin transport is regulated at the cilium or cell level. After shearing of just one of the two cilia, cells will partially resorb the remaining cilium while regrowing the missing one giving so-called long-short cells (Fig. 4 A and see Fig. 6 E; Rosenbaum et al., 1969). An elevated tubulin transport in both cilia of a long-short cell would indicate a cell-wide regulation, whereas an increase limited to the growing cilium would indicate a regulation that operates at the single cilium level. The mean GFP–α-tubulin IFT frequencies were 20.5 particles/min for the short and 3.3 particles/min for the long cilia of long-short cells (n = 17; ±11.6 and 3.2 particles/min, respectively; Fig. 4, B and C; Video 6). In all long-short cells analyzed, the transport frequency in the short cilium exceeded that of the long one; this difference decreased as both cilia approached a similar length (Fig. 4 D).


As an alternative approach that does not require mechanical shearing, we generated long-short cells by mating GFP–α-tubulin–expressing gametes with full-length cilia to wild-type gametes that were deciliated by a pH shock and regenerating cilia (Fig. 5, A and B, a and b). The resulting zygotes, initially possessing two long GFP-positive and two short GFP-negative cilia, will equalize the length of all four cilia by partially resorbing the long ones and adding new tubulin to the short ones (Fig. 5 B, c–g; Ludington et al., 2012). Tubulin transport in all four cilia of such zygotes was analyzed after photobleaching (Fig. 5 C; Video 7). The frequency of tubulin transport by anterograde IFT in the short growing cilia surpassed that of the nongrowing long cilia (Fig. 5 D). As the four cilia approached equal length, the transport frequency in the initially long cilia increased, whereas that in the initially short ones decreased (Fig. S4 D). Based on the study of long-short cells, we conclude that the frequency of IFT-based tubulin transport events is controlled cilium-autonomously.

In steady-state cilia, ∼10% of the total tubulin was in the detergent-soluble MM fraction (Fig. 1 D). The elongation of microtubules depends, among other factors, on the availability of αβ-tubulin dimers (Desai and Mitchison, 1997; Howard, 2001). Western blotting and FRAP analysis were used to test whether steady-state and growing cilia differ in the concentration of soluble tubulin. Steady-state, growing, and fully regenerated cilia were separated into axonemal and MM fractions; loading was adjusted to represent equal volumes of MM (Fig. 6 A). Western blots revealed that the amount of soluble tubulin was doubled from ∼10 to 20% of the total ciliary tubulin in growing versus nongrowing flagella (Fig. 6 A; for quantification see Fig. S4, B and C).
For FRAP analysis, ciliary segments were spot bleached (Video 8). An exchange of axonemal tubulin with GFP–α-tubulin laterally along the ciliary shaft was not observed in this study. Thus, axonemal GFP–α-tubulin will remain bleached. Soluble, unbleached GFP–α-tubulin from other ciliary segments or de novo imported from the cell body will move into the bleached region, and fluorescence recovery reflects the concentration of mobile GFP-tubulin in the cilium (Fig. 6 B). FRAP analysis showed recovery rates of 2 ± 1% of the pre-bleach fluorescence for nongrowing cilia (n = 16), whereas ∼15.6 ± 10% of the signal intensity recovered in growing cilia (n = 18; Fig. 6, C and D). In growing cilia, GFP-tubulin moving on IFT trains through the bleached region contributes to the recovery, but over time the diffused background staining significantly increased, indicating an increase in the concentration of GFP-tubulin diffusing inside cilia (Figs. 3 F, 6 F, and S5 B). Repeated bleaching of the same ciliary segment was followed by a recovery of similar strength (∼5% for steady-state and ∼15% for growing cilia; Fig. S5), indicative of an ongoing de novo import of GFP–α-tubulin into cilia.
An increase in the concentration of soluble tubulin in cilia could result from an increase in soluble tubulin in the cell body of regenerating cells. FRAP of long-short cells, however, showed that recovery was significantly stronger in the short cilium than in the long one (39 ± 10% vs. 4.3 ± 3%; Fig. 6, E–H; Fig. S5, C and D; Video 9). Thus, distinct concentrations of soluble tubulin can be established in the two cilia of the same cell. We conclude that the concentration of soluble tubulin is elevated in growing cilia. A high concentration of tubulin in the ciliary matrix is likely to promote elongation of the axonemal microtubules.
Defective regulation of tubulin transport in ciliary length mutants
Because microtubules are the major structural elements of cilia, defective regulation of tubulin transport could affect ciliary length. We expressed GFP–α-tubulin in lf2-1 (long flagella2) and shf2 (short flagella2) mutants and selected transformants expressing similar amounts of GFP-tubulin as the control strain GFP-Tub1 (Fig. S3 C). lf2-1 has cilia of variable length with some 2–3× longer than wild type, whereas cilia reach only 6–8 µm in shf2. LF2 encodes a CDK-like kinase, whereas the molecular defect in shf2 is unknown (Kuchka and Jarvik, 1987; Tam et al., 2007). In steady-state lf2-1 cilia, the mean frequency of GFP–α-tubulin transport was higher than in steady-state wild-type cilia; a length-dependent decrease in frequency was not observed (Fig. 7 A). Cilia regeneration in lf2-1 was slow and irregular in onset with many cells failing to reassemble cilia (Barsel et al., 1988). In the lf2-1 cells that regenerated cilia, the frequency of tubulin transport was somewhat higher but showed a similar lack of length dependency. The frequency of IFT was slightly reduced in lf2-1 (unpublished data). lf2-1 cells apparently fail to properly regulate the IFT-based transport of tubulin in response to changes in ciliary length.

In steady-state cilia of shf2, the frequency of tubulin transport decreased with increasing ciliary length and tubulin transport was rarely observed in shf2 cilia longer than ∼6 µm, a length at which wild-type cilia showed robust trafficking of tubulin (Figs. 7 A and 3 D). IFT (based on DIC microscopy) appeared normal in the longer shf2 cilia (unpublished data). Many shf2 cells failed to regenerate cilia but the regrowth kinetics was largely normal in those that did (Kuchka and Jarvik, 1987). Regenerating shf2 cilia showed a similar steep down-regulation of tubulin transport frequency with increasing ciliary length as described for shf2 in steady-state. The data suggest that shf2, although able to increase tubulin transport frequency in short regenerating cilia, prematurely down-regulates tubulin transport by IFT. The data show that an abnormal regulation of tubulin transport occurs in mutants with defects in ciliary length regulation.
Ciliary assembly requires a massive translocation of tubulin from the cell body into the growing organelle. InC. reinhardtii, almost 800,000 tubulin dimers, corresponding to ∼20% of the cell’s total tubulin, are required to assemble its two 12-µm-long 9+2 cilia (Bhogaraju et al., 2014b; see Materials and methods). IFT is essential for the assembly of cilia and flagella in most eukaryotes and thought to be the major pathway of protein transport in cilia (Rosenbaum and Witman, 2002). Recent data, however, have shown that numerous proteins move into cilia in an IFT-independent manner. Ectopic cytosolic proteins up to a size of ∼50 kD enter cilia by passive diffusion; even proteins up to ∼650 kD appear to diffuse slowly into cilia (Kee et al., 2012; Breslow et al., 2013; Lin et al., 2013). In C. reinhardtii, the transmembrane protein SAG1 and the membrane-associated phospholipase D will enter cilia in the absence of IFT (Belzile et al., 2013; Lechtreck et al., 2013). Furthermore, certain transmembrane proteins (SSTR3, Smo) diffuse inside the ciliary membrane while associating only transiently—if ever—with IFT (Ye et al., 2013). These findings raised the question which proteins are the actual cargoes of IFT. Several TRP channels were reported to move by IFT and the BBSome cycles through cilia by associating to IFT trains (Blacque et al., 2004; Qin et al., 2004; Huang et al., 2007; Lechtreck et al., 2009). Also, IFT powers gliding and ciliary surface motility in C. reinhardtiipresumably due to its interaction with the ciliary transmembrane protein FMG-1B (Collingridge et al., 2013;Shih et al., 2013). We recently showed that the axonemal proteins PF16, DRC2, and DRC4 are cargoes of IFT and that entry of DRC4-GFP into cilia is largely diminished in the absence of IFT (Wren et al., 2013). Defects in the IFT complex B protein IFT46 and or its interacting partner ODA16p largely prevent the transport of outer arm dynein to its ciliary docking sites (Hou et al., 2007; Ahmed et al., 2008). Thus, IFT plays a role in the transport of axonemal proteins, several of which are transported in large complexes feasibly rendering transport by diffusion inefficient. Several studies provided evidence that tubulin is a cargo of IFT but direct imaging of tubulin transport by IFT and the quantification of such events has been challenging (Marshall and Rosenbaum, 2001; Hao et al., 2011; Bhogaraju et al., 2013; Brust-Mascher et al., 2013). Here, we show that GFP-tagged tubulin moves in association with IFT particles inside cilia indicating that IFT functions as tubulin transporter.
Cilium-autonomous regulation of tubulin transport by IFT
In growing cilia, the frequency of GFP-tubulin transport by IFT was greatly elevated. Taking into account that only a fraction of the total tubulin in cilia was tagged, extrapolation of the data indicates that IFT trains in growing cilia carry a larger tubulin load than those in steady-state cilia. Previously, we showed that the IFT-based transport of the axonemal protein DRC4 is strongly increased during ciliary growth (Wren et al., 2013). It is likely that the transport of many other ciliary proteins is also enhanced when cilia elongate. Although some differences in the velocity, size, and frequency of IFT in growing vs. steady-state cilia of C. reinhardtii have been reported, the number of IFT particles inside cilia is largely length-independent (Dentler, 2005; Marshall et al., 2005; Engel et al., 2009). In conclusion, IFT trains are highly loaded with cargo during ciliary growth; then, the number and velocity of IFT trains might limit the rate of ciliary elongation. Indeed, the 60% reduction in IFT frequency observed in the IFT kinesin mutant fla3-1 results in sluggish regrowth of nearly full-length cilia (Mueller et al., 2005). In steady-state cilia, IFT particles carry less cargo probably running well below capacity. This would allow the cells to quickly adjust ciliary protein content and length in response to developmental or environmental cues by altering cargo loading of IFT (Wren et al., 2013).
Currently, it is unknown how the amount of cargo transported by IFT into cilia is regulated. In a simple model, an increase in the concentration of soluble tubulin in the cell body would make more tubulin available as cargo for IFT and promote tubulin transport into cilia (Chan and Marshall, 2012; Goehring and Hyman, 2012). Indeed, primary cilia elongate when the level of soluble cytoplasmic tubulin is increased by drug-induced microtubule depolymerization (Sharma et al., 2011). During ciliary growth, kinesin-13 may depolymerize cell body microtubules in C. reinhardtii to provide tubulin for axonemal assembly (Wang et al., 2013). Remarkably, in cells possessing growing and nongrowing cilia, the frequency of IFT-based GFP-tubulin transport was strongly elevated in the growing over the nongrowing cilia. We conclude that a general increase in tubulin supply in the cell body is insufficient to explain the increase in IFT-based tubulin transport just into growing cilia of long-short cells. Our data suggest that tubulin import into cilia is regulated locally within the basal body–cilium entity. An increase in admission of tubulin to the cilium could involve a change in the transition zone (TZ), located between the basal body and the cilium proper. TZ proteins are hotspots for ciliopathy-related mutations which alter the protein composition of cilia, and the TZ is thought to function as a ciliary gate regulating protein entry into the organelle (Craige et al., 2010; Omran, 2010; Williams et al., 2011; Kee and Verhey, 2013). The TZ could respond to changes in the ciliary length by permitting IFT particles carrying a large load into growing cilia while preventing such particles from entering steady-state cilia. Such a model of size-based exclusion of IFT trains at the TZ would require the constant presence of both loaded and unloaded IFT particles. The share of IFT trains occupied with GFP-tubulin in nongrowing and growing cilia ranges from <10 to ∼80%, rendering this model unlikely.
Alternatively, cargo loading onto IFT particles could be regulated. Dikaryon experiments showed that IFT proteins from distinct basal body–cilia units mix rapidly (Wren et al., 2013). Therefore, an increase in cargo loading onto IFT must happen locally at the base of a growing cilium, for example, by increasing the affinity of IFT carriers for cargo proteins. In C. reinhardtii, IFT particle and motor proteins are concentrated at the transitional fibers, which are located proximal to the TZ (Deane et al., 2001). The transitional fibers could increase the local concentration of IFT and cargoes and thereby promote the formation of IFT–cargo complexes. The anterograde IFT motor subunit KAP is more abundant at the base of growing cilia (Ludington et al., 2013), which might suggest that transitional fibers at the base of growing cilia have a higher capacity to bind IFT material. We propose a model in which cells measure the length of their cilia; cilia of insufficient length generate a signal that alters the property of the ciliary base, resulting in an increased loading of IFT particles and/or facilitated admission of highly loaded IFT trains into the cilium, which will augment the amount of tubulin transported into the cilium in need (Fig. 7 B).
Ciliary length defects are associated with dysregulation of tubulin transport
Sensing of ciliary length is likely to involve various protein kinases whose mutations lead to length defects (Berman et al., 2003; Tam et al., 2007; Lefebvre, 2009; Cao et al., 2013; Tam et al., 2013). LF2p, for example, is a CDK kinase which, together with LF1p and LF3p, forms the length regulating complex (LRC;Tam et al., 2007). LF2p homologues are also involved in ciliary length control in mammals and C. elegans(Ko et al., 2010; Phirke et al., 2011). In lf2-1, the frequency of tubulin transport was largely independent of ciliary length and remained elevated in lf2-1 cilia that were longer than wild-type cilia. A failure to adjust tubulin transport with respect to ciliary length could explain the phenotype of lf2-1 characterized by slow regeneration kinetics, and variable ciliary length with some abnormally long cilia (Barsel et al., 1988). The LRC is located in the cell body of C. reinhardtii, but its molecular role is unknown (Tam et al., 2007). Based on its localization, the LRC could function in adjusting IFT-based tubulin transport in response to signals from the ciliary length sensor.
Shorter-than-normal cilia could result from a failure to incorporate precursors into the cilium, an increased disassembly rate, or an insufficient delivery of precursors. The latter was observed in the short flagella mutantshf2. Tubulin transport was down-regulated well before cilia reached wild-type length. The phosphorylation pattern of the aurora-like kinase CALK can be used as an indicator of the assembly state of cilia. At the onset of ciliary growth, phosphorylation and dephosphorylation occur at distinct sites, followed by a gradual return to a pattern characteristic for full-length steady-state cilia (Luo et al., 2011; Cao et al., 2013). shf2 displays a CALK phosphorylation state typical for short, growing cilia (Luo et al., 2011; Cao et al., 2013). This suggests that shf2 cells still sense the insufficient length of their cilia but fail to respond appropriately by maintaining a high tubulin influx via IFT.
In our study, we did not address the question of why the longer cilia of long-short cells retract. In those long cilia, tubulin transport via IFT—although well below that of short cilia—was actually somewhat above the steady-state levels (Fig. 4 C). Also, the breakdown of the axoneme is likely to increase the levels of soluble tubulin inside the shortening long cilia. Thus, ciliary shortening appears not to be driven by a shortage of tubulin, suggesting that the intraciliary tubulin concentration is not the sole factor regulating ciliary growth. We posit that the resorption involves an up-regulation of the cilia disassembly pathway in the long cilia. Putative players in the disassembly pathway are microtubule-destabilizing proteins such as kinesin-13 and CNK2. Mutants in the latter assemble the missing cilium of long-zero cells at normal rates but fail to resorb the remaining long cilium, supporting the notion that ciliary assembly and disassembly in long-short cells are distinctly regulated (Hilton et al., 2013).
IFT promotes ciliary growth by concentrating tubulin
GFP-tubulin enters cilia by both diffusion and IFT, raising the question of the respective roles in supplying tubulin for ciliary assembly. Tubulin dimers (110 kD) are above the predicted size exclusion limit of ∼50 kD for free diffusion of soluble protein into the cilium (Kee et al., 2012; Breslow et al., 2013). Distinct concentrations of soluble tubulin were observed in cilia of a given cell, indicating the absence of diffusive equilibration. We estimate (see Materials and methods) that the concentration of soluble tubulin in the ciliary matrix of growing cilia is higher than that in the cytoplasm; the absence of a cell body–to–cilia gradient in tubulin concentration rules out a net influx of tubulin into cilia by diffusion. In fla10-1, GFP-tubulin continued to enter cilia by diffusion in the absence of IFT but the cilia do not elongate. Similarly, ciliary elongation ceased in shf2 after IFT of GFP-tubulin was down-regulated despite the continued entry of GFP-tubulin by diffusion. We conclude that the entry of tubulin into cilia by diffusion is insufficient to provide enough tubulin to promote ciliary growth.
Tubulin concentration has a strong impact on microtubule growth in vitro and in vivo, with high concentrations of tubulin suppressing catastrophe frequency and increasing the polymerization rate (Pedigo and Williams, 2002). In the cytoplasm, proteins such as XMAP215 act as microtubule polymerases at MT plus ends putatively by increasing the local tubulin dimer concentration (Al-Bassam and Chang, 2011). Messages of specific tubulin isoforms are recruited to microtubule plus-ends in the axonal growth cone to locally increase tubulin expression (Preitner et al., 2014). In growing cilia, large amounts of tubulin released from IFT into the cilium will increase the concentration of soluble tubulin in the matrix. We noticed a reduced mobility of tubulin in the vicinity of the ciliary tip, which could result from tubulin interactions with other proteins; IFT proteins, for example, are typically accumulated at the ciliary tip and possess tubulin binding motifs (Bhogaraju et al., 2014b). Further, GFP-tubulin was predominately released from IFT near the ciliary tip; reduced mobility and preferred unloading could further increase the tubulin concentration locally at the tip.
We predict that the concentration of soluble tubulin in the matrix of growing cilia is in the range of 300 µM (see Materials and methods). Although some of the tubulin is sequestered onto IFT, the concentration is well above the concentrations (∼20 µM) required for microtubule assembly in vitro or in the cytoplasm. C. reinhardtii cilia elongate at ∼350 nm/min compared with 20–60 µm/min observed for microtubules in the cytoplasm (Srayko et al., 2005). However, during axonemal assembly, nine doublet microtubules and two singlet microtubules are assembled simultaneously within the comparably small volume of the cilium. About 2,500 dimers are required to elongate a singlet microtubule by 20 µm, whereas the assembly of 350 nm of 9+2 microtubules consumes ∼10,000 dimers. We estimate ∼18,000 dimers to be present in the matrix of a 4-µm-long cilium (see Materials and methods). Thus, soluble tubulin in the ciliary matrix will be rapidly consumed during ciliary growth and the matrix pool must be equally rapidly replenished to ensure continued elongation of the axoneme. A unifying feature of most cilia is doublet microtubules, the assembly of which has not yet been reconstituted in vitro and could require very high concentrations of tubulin dimers. In summary, due to its architecture and its position within the small volume of the cilium, axonemal assembly could require exceptionally high concentrations of soluble tubulin. Several IFT complex B proteins contain tubulin-binding motifs—confirmed and predicted—and estimates suggest that IFT has the capacity to transport tubulin in amounts required during ciliary assembly (Bhogaraju et al., 2014). Although additional factors are likely involved in determining the rate of ciliary elongation (most notably the rate of axoneme depolymerization), tubulin transport by IFT and the resulting high concentration of tubulin in the ciliary matrix are likely prerequisites for the rapid elongation of axonemal microtubules. We propose that IFT functions as a tubulin pump to increase the concentration of tubulin inside growing cilia. Higher tubulin concentrations will promote the growth of the axonemal microtubules and thereby elongation of the cilium.
 Their first paper demonstrated that
Their first paper demonstrated that 


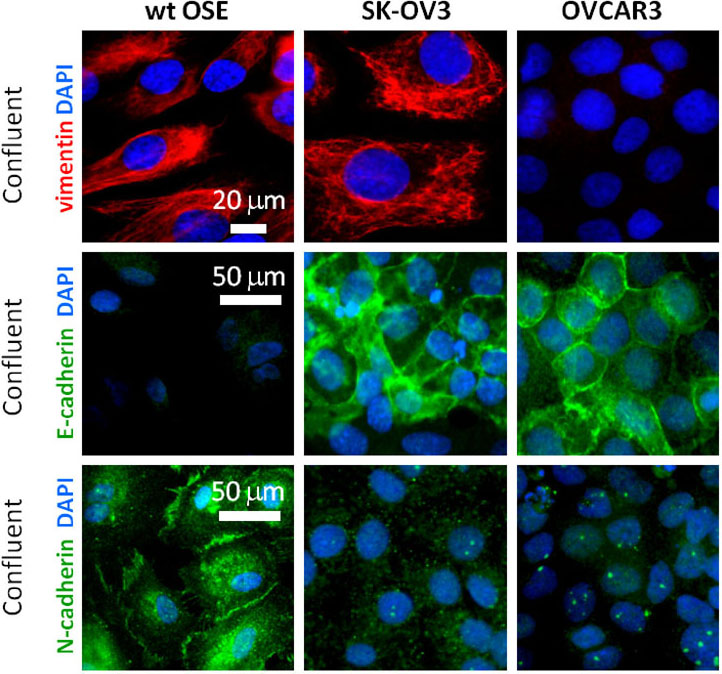
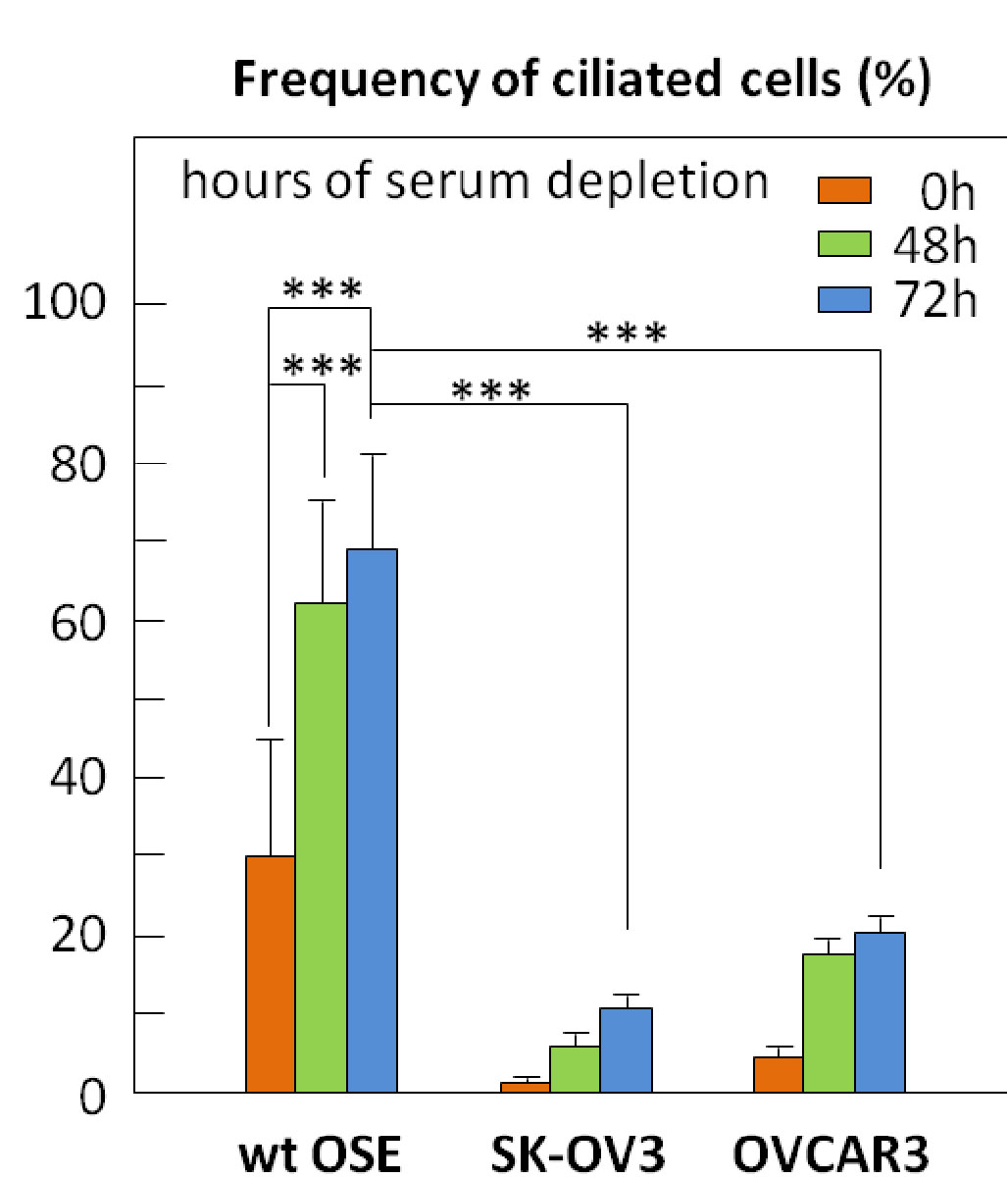

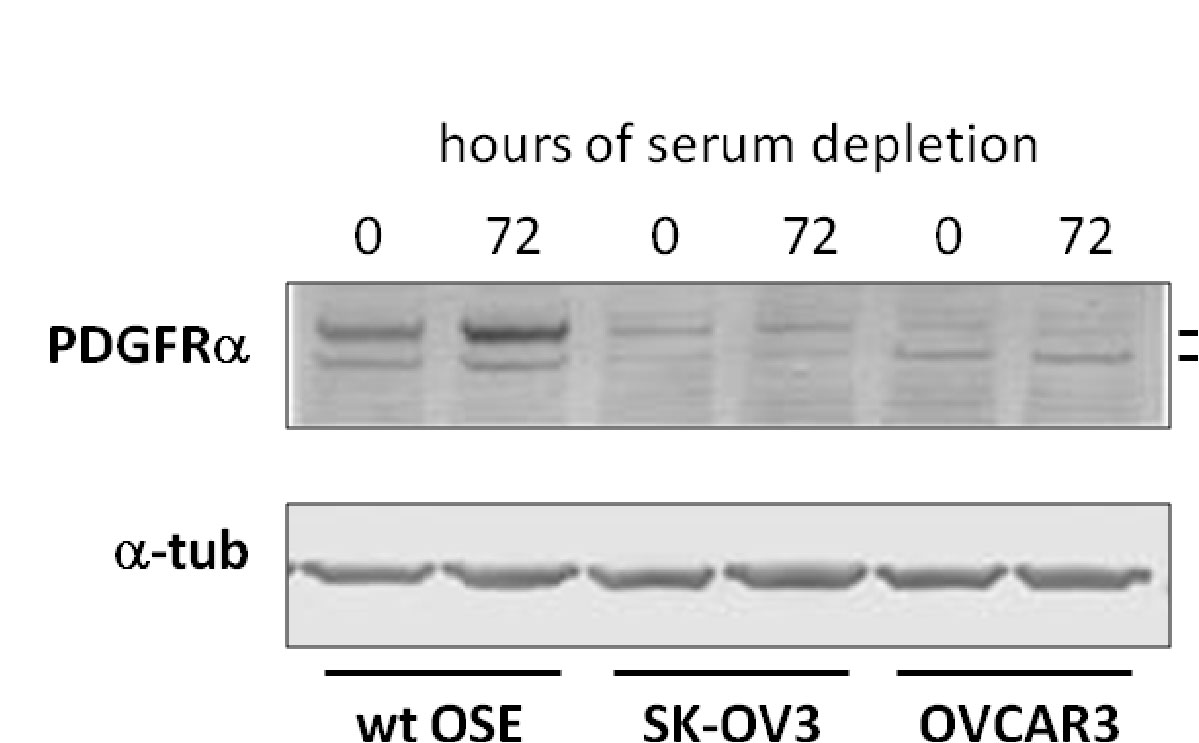
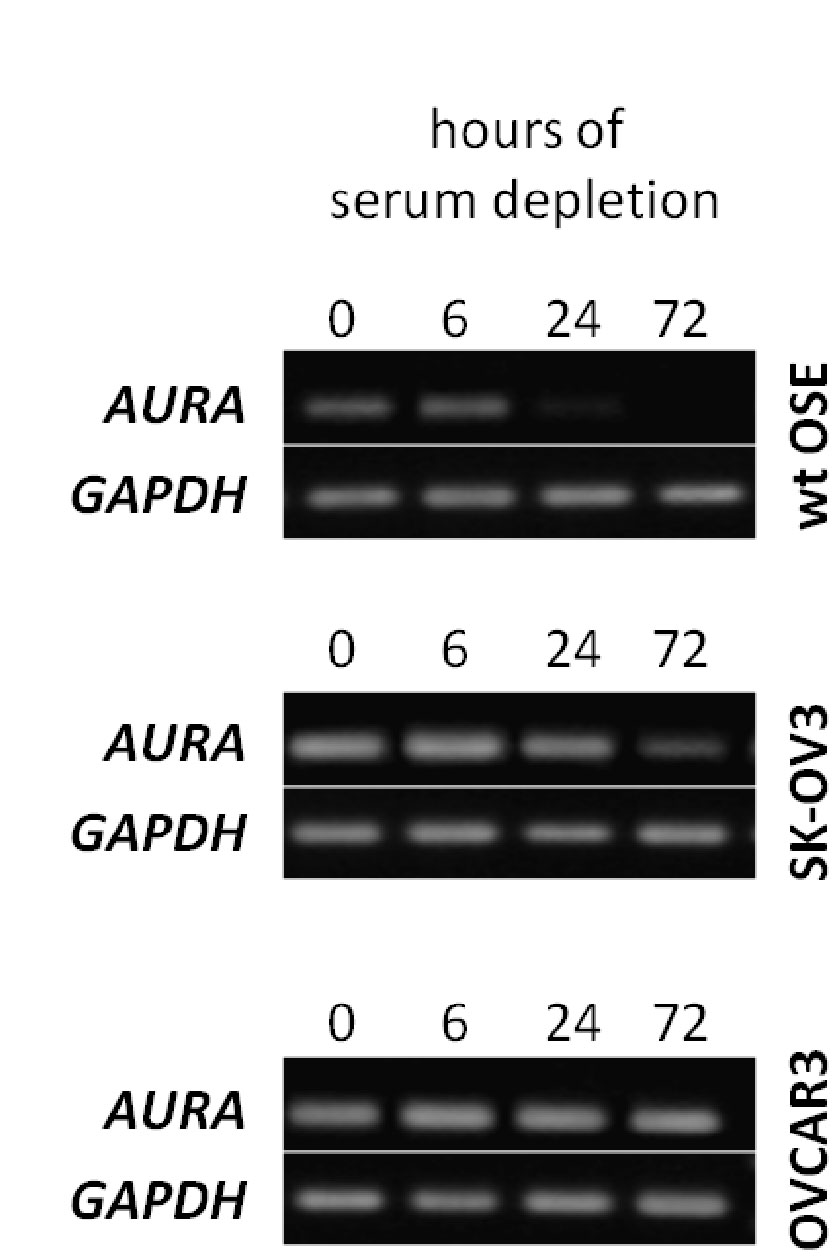

 Physiologically, the proteins play a role in calcium and Pi homeostasis as demonstrated by studies on mouse transgenic models. In addition to cancer, the protein has been linked to atherosclerosis, hypoxia response and in wound repair (Lal et al., 2001; Iyer et al., 1999). Pharmacologically, an STC1 receptor has been deduced from studies and thought to be localized to the mitochondria where it has been shown to have a relationship with the mitochondrial electron transfer (McCudden et al., 2002). Recent studies show that STC1 activates the mitochondrial antioxidant pathway through its regulation of intracellular calcium (Sheikh-Hamad, 2010). Overall, STC1 and STC2 are thought to be secreted as phosphoproteins as demonstrated by coimmunoprecipitations of cellular lysates. And, it’s thought the proteins play a role in mineralizing tissues (e.g. bone) to control the levels of calcium and Pi via their influence on calcium channels and sodium/Pi co-transporters. A schematic diagram showing how stannniocalcin might be have pro-apoptic functions is shown in Figure 1 (Yeung et al., 2012).
Physiologically, the proteins play a role in calcium and Pi homeostasis as demonstrated by studies on mouse transgenic models. In addition to cancer, the protein has been linked to atherosclerosis, hypoxia response and in wound repair (Lal et al., 2001; Iyer et al., 1999). Pharmacologically, an STC1 receptor has been deduced from studies and thought to be localized to the mitochondria where it has been shown to have a relationship with the mitochondrial electron transfer (McCudden et al., 2002). Recent studies show that STC1 activates the mitochondrial antioxidant pathway through its regulation of intracellular calcium (Sheikh-Hamad, 2010). Overall, STC1 and STC2 are thought to be secreted as phosphoproteins as demonstrated by coimmunoprecipitations of cellular lysates. And, it’s thought the proteins play a role in mineralizing tissues (e.g. bone) to control the levels of calcium and Pi via their influence on calcium channels and sodium/Pi co-transporters. A schematic diagram showing how stannniocalcin might be have pro-apoptic functions is shown in Figure 1 (Yeung et al., 2012). However, stanniocalcin’s more prominent role is arguably as a cancer biomarker. Its expression has been shown to be affected in a number of different cancer pathologies. Table 1 shows a representative list of cancers where stanniocalcin levels are differentially expressed depending on the cancer. Thus, it appears that stanniocalcin is a good candidate cancer biomarker. It is hypothesized that this is due in part to its role in tumor vasculature (Chang et al., 2003). It should be noted that the list is but a brief compilation while stanniocalcin has been linked to other cancers as well.
However, stanniocalcin’s more prominent role is arguably as a cancer biomarker. Its expression has been shown to be affected in a number of different cancer pathologies. Table 1 shows a representative list of cancers where stanniocalcin levels are differentially expressed depending on the cancer. Thus, it appears that stanniocalcin is a good candidate cancer biomarker. It is hypothesized that this is due in part to its role in tumor vasculature (Chang et al., 2003). It should be noted that the list is but a brief compilation while stanniocalcin has been linked to other cancers as well.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}