Acetylation and Deacetylation of non-Histone Proteins
Author and Curator: Larry H Bernstein, MD, FCAP
Acetylation and Deacetylation of non-histone proteins
MA Glozak, N Sengupta, X Zhang, E Seto
Gene 2005; 363(19): 15-23 http://dx.doi.org/10.1016/j.gene.2005.09.010
Since the first report of p53 as a non-histone target of a histone acetyltransferase (HAT), there has been a rapid proliferation in the description of new non-histone targets of HATs. Of these,
- transcription factors comprise the largest class of new targets.
The substrates for HATs extend to
- cytoskeletal proteins,
- molecular chaperones and
- nuclear import factors.
Deacetylation of these non-histone proteins by histone deacetylases (HDACs) opens yet another exciting new field of discovery in
- the role of the dynamic acetylation and deacetylation on cellular function.
This review will focus on these non-histone targets of HATs and HDACs and the consequences of their modification.
Abbreviations:
HAT, histone acetyltransferase; HDAC, histone deacetylase; TSA, trichostatin A; CtBP, C-terminal binding protein; YY1, yin yang 1; HMG, high mobility group; NR, nuclear receptor; AR, androgen receptor; ER α, estrogen receptor α; SHP, short heterodimer partner; EKLF, erythroid Kruppel like factor; Rb, retinoblastoma; GR, glucocorticoid receptor; HDV, hepatitis delta virus; L-HDAg, large delta antigen; S-HDAg, small delta antigen
Keywords HATs; HDACs; Post-translational modification
Histone deacetylases (EC 3.5.1.98, HDAC) are a class of enzymes that
- remove acetyl groups (O=C-CH3) from an ε-N-acetyl lysine amino acid on a histone,
- allowing the histones to wrap the DNA more tightly.
This is important because DNA is wrapped around histones, and
- DNA expression is regulated by acetylation and de-acetylation.
Its action is opposite to that of histone acetyltransferase. HDAC proteins are now also called
- lysine deacetylases (KDAC),
- to describe their function rather than their target, which also
- includes non-histone proteins
Histone modification
Histone tails are normally positively charged due to
These positive charges help the histone tails to
- interact with and bind to the negatively charged phosphate groups on the DNA backbone.
Acetylation, which occurs normally in a cell,
- neutralizes the positive charges on the histone by changing amines into amides and
- decreases the ability of the histones to bind to DNA.
This decreased binding
- allows chromatin expansion,
- permitting genetic transcription to take place.
Histone deacetylases
- remove those acetyl groups,
- increasing the positive charge of histone tails and
- encouraging high-affinity binding between the histones and DNA backbone.
The increased DNA binding
- condenses DNA structure,
- preventing transcription.
Histone deacetylase is involved in a series of pathways within the living system. According to the Kyoto Encyclopedia of Genes and Genomes (KEGG), these are:
- Environmental information processing; signal transduction; notch signaling pathway PATH:ko04330
- Cellular processes; cell growth and death; cell cycle PATH:ko04110
- Human diseases; cancers; chronic myeloid leukemia PATH:ko05220
Histone acetylation plays an important role in the regulation of gene expression.
Hyperacetylated chromatin is
- transcriptionally active, and
hypoacetylated chromatin
- is silent.
A study on mice found that a
- specific subset of mouse genes (7%) was
- deregulated in the absence of HDAC1.[10]
Their study also found a
- regulatory crosstalk between HDAC1 and HDAC2 and suggest
- a novel function for HDAC1 as a transcriptional coactivator.
HDAC1 expression was found to be
- increased in the prefrontal cortex of schizophrenia subjects,[11]
- negatively correlating with the expression of GAD67 mRNA.
Non-histone effects
It is a mistake to regard HDACs solely in the context of regulating gene transcription by modifying histones and chromatin structure, although
- that appears to be the predominant function.
The function, activity, and stability of proteins can be controlled by
Protein phosphorylation is perhaps the most widely studied and understood modification in which
- certain amino acid residues are phosphorylated by the action of protein kinases or
- dephosphorylated by the action of phosphatases.
The acetylation of lysine residues is emerging as an analogous mechanism, in which
- non-histone proteins are acted on by acetylases and deacetylases.[12]
It is in this context that HDACs are being found to interact with a variety of non-histone proteins—
some of these are transcription factors and co-regulators, some are not. Note the following four examples:
- HDAC6 is associated with aggresomes.Misfolded protein aggregates are
- tagged by ubiquitination and removed from the cytoplasm by dynein motors via the microtubule network to an organelle termed the aggresome.
- HDAC 6 binds polyubiquitinated misfolded proteins and links to dynein motors, thereby
- allowing the misfolded protein cargo to be physically transported to chaperones and proteasomes for subsequent destruction.[13]
- PTEN is an important phosphatase involved in cell signaling via phosphoinositols and the AKT/PI3 kinase pathway.
- PTEN is subject to complex regulatory control via phosphorylation, ubiquitination, oxidation and acetylation.
- Acetylation of PTEN by the histone acetyltransferase p300/CBP-associated factor (PCAF) can repress its activity; on the converse,
- deacetylation of PTEN by SIRT1 deacetylase and, by HDAC1, can stimulate its activity.[14][15]
- APE1/Ref-1 (APEX1) is a multifunctional protein possessing both
- DNA repair activity (on abasic and single-strand break sites) and
- transcriptional regulatory activity associated with oxidative stress.
- APE1/Ref-1isacetylatedbyPCAF; on the converse,
- it is stably associated with and deacetylated by Class I HDACs.
- The acetylation state of APE1/Ref-1 does not appear to affect its DNA repair activity, but it does
- regulate its transcriptional activity such as
- its ability to bind to the PTH promoter and initiate transcription of the parathyroid hormone gene.[16][17]
- NF-κB is a key transcription factor and
HDAC inhibitors
Main article: Histone deacetylase inhibitor
Histone deacetylase inhibitors (HDIs) have a long history of use in psychiatry and neurology as mood stabilizers and anti-epileptics,
- for example, valproic acid.
In more recent times, HDIs are being studied as
- a mitigator or treatment for neurodegenerative diseases.[19][20]
- there has been an effort to develop HDIs for cancer therapy.[21][22]
- Vorinostat (SAHA) was approved in 2006 for the treatment of cutaneous manifestations in patients with cutaneous T cell lymphoma (CTCL) that have failed previous treatments.
- A second HDI, Istodax (romidepsin), was approved in 2009 for patients with CTCL.
The exact mechanisms by which the compounds may work are unclear, but
- epigenetic pathways are proposed.[23] In addition, a clinical trial is studying valproic acid effects on the latent pools of HIV in infected persons.[24]
HDIs are currently being investigated as chemosensitizers for
- cytotoxic chemotherapy or radiation therapy, or in association with DNA methylation inhibitors based on in vitro synergy.[25]
Recent research has focused on developing isoform selective HDIs which can aid in elucidating role of
- individual HDAC isoforms and device strategy for effective treatment of
- diseases related to relevant HDAC isoform.[26][27][28]
HDAC inhibitors have effects on non-histone proteins that are related to acetylation. HDIs can
- alter the degree of acetylation of these molecules and, therefore,
- increase or repress their activity.
For the four examples given above (see Function) on HDACs acting on non-histone proteins, in each of those instances
- the HDAC inhibitor Trichostatin A (TSA) blocks the effect.
HDIs have been shown to alter the activity of many transcription factors, including
ACTR, cMyb, E2F1, EKLF, FEN 1, GATA, HNF-4, HSP90, Ku70, NFκB, PCNA, p53, RB, Runx, SF1 Sp3, STAT, TFIIE, TCF, YY1.[29][30]
To carry out gene expression, a cell must control the coiling and uncoiling of DNA around histones. This is accomplished with the assistance of histone acetyl transferases (HAT), which
- acetylate the lysine residues in core histones leading to
- a less compact and more transcriptionally active chromatin, and, on the converse,
- the actions of histone deacetylases (HDAC), which
- remove the acetyl groups from the lysine residues
- leading to the formation of a condensed and transcriptionally silenced chromatin.
Reversible modification of the terminal tails of core histones constitutes
- the major epigenetic mechanism for remodeling higher-order chromatin structure and controlling gene expression.
HDAC inhibitors (HDI) block this action and
The histone deacetylase inhibitors are a new class of cytostatic agents that inhibit the proliferation of tumor cells in culture and in vivo
- by inducing cell cycle arrest,
- differentiation
- and/or apoptosis.
Histone deacetylase inhibitors exert their anti-tumour effects via
- the induction of expression changes of oncogenes or tumour suppressor, through
- modulating that the acetylation/deactylation of histones and/or non-histone proteins such as transcription factors[8].
Histone acetylation and deacetylation play important roles in the modulation of chromatin topology and the regulation of gene transcription.
Histone deacetylase inhibition induces
- the accumulation of hyperacetylated nucleosome core histones in most regions of chromatin
but affects the expression of only a small subset of genes, leading to transcriptional activation of some genes, but repression of an equal or larger number of other genes.
Non-histone proteins such as transcription factors are also targets for acetylation with varying functional effects. Acetylation
- enhances the activity of some transcription factors such as the tumor suppressor p53 and
- the erythroid differentiation factor GATA-1
- but may repress transcriptional activity of others including T cell factor and the co-activator ACTR.
Recent studies […] have shown that the estrogen receptor alpha (ERalpha) can be hyperacetylated
- in response to histone deacetylase inhibition,
- suppressing ligand sensitivity and regulating transcriptional activation by histone deacetylase inhibitors.[9]
Conservation of the acetylated ER-alpha motif in other nuclear receptors suggests that
- acetylation may play an important regulatory role in diverse nuclear receptor signaling functions.
A number of structurally diverse histone deacetylase inhibitors have shown potent antitumor efficacy with little toxicity in vivo in animal models. Several compounds are currently in early phase clinical development as potential treatments for solid and hematological cancers both as monotherapy and in combination with cytotoxics and differentiation agents.”[10]
HDIs MI · Granger, A.; Abdullah, I.; Huebner, F.; Stout, A.; Wang, T.; Huebner, T.; Epstein, J. A.; Gruber, P. J. (2008). “Histone deacetylase inhibition reduces myocardial ischemia-reperfusion injury in mice”. The FASEB Journal 22 (10): 3549–60. http://dx.doi.org/10.1096/fj.08-108548. PMC 2537432. PMID 18606865.
Protein Acetylation: Much More than Histone Acetylation
By Tom Brock, Ph.D.
Just last decade, everyone was excited about the Human Genome Project, and the gene was king. Today, epigenetics is reminding us that
- non-genetic factors are important in shaping gene expression and development.
Similarly, where phosphorylation once seemed the primary way to modulate proteins,
- epigenetics has re-introduced us to acetylation as an important force in defining protein function.
In particular, the acetylation of histones has moved to center stage, even though it was described over 45 years ago. Research on histone acetylation has
- led to a resurgence in the interest in enzymatically-mediated acetylation of other proteins.
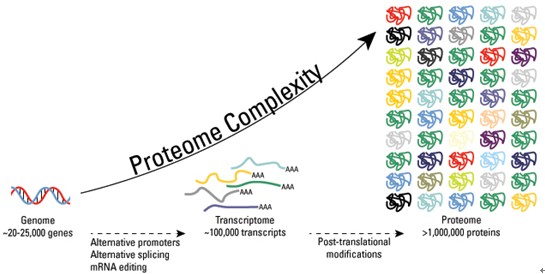
This article examines acetylation as a post-translational modification of proteins that impacts gene expression and plays a role in epigenetics.
The Basics
Acetylation refers to the addition of an acetyl group (CH3CO) to organic compounds. Proteins can be acetylated by both enzymatic and non-enzymatic processes.
One group of acetyltransferases commonly catalyze the transfer of an acetyl group from acetyl-CoA to the terminal amine on the side chain of lysine residues (Figure 1).
These enzymes are commonly called HATs, because their best-known substrates have been histones.
However, the nomenclature is being revised to lysine acetyltransferases (KATs), reflecting their ability to acetylate lysine (denoted ‘K’) on many proteins.
1 The KATs are numerous, with many assigned, based on structural similarities, to either
- the GNAT (Gcn5-related N-acetyltransferases) superfamily or
- the MYST (MOZ, YBF2/Sas3, Sas2, Tip60) family.
Other important KATs include
- p300 (E1A-associated protein 300 kDa),
- CBP (cAMP response element binding (CREB)-binding protein), and
- TAFII 250 (TATA-binding protein associated factor II 250).
The conversion of the positively charged lysine to acetyl-lysine, like the addition of negative phosphates to uncharged amino acids during phosphorylation,
alters protein structure and interactions with other biomolecules. For example, acetylation of histones typically
- promotes the recruitment of effector proteins,
- relaxation of chromatin conformation, and
- an increase in transcription.
Like phosphorylation,
- acetylation is reversible.
Histone deacetylases (HDACs, a.k.a. KDACs) are a smaller group of evolutionarily conserved enzymes.
The human class I HDACs are
- homologous to the yeast enzyme Rpd3 and include HDAC1, 2, 3, and 8.
Class II HDACs are
- homologous to yeast HDA1 and are divided into class IIa (HDAC4, 5, 7, 9) and class IIb (HDAC6 and 10) based on structure.
- The human class III HDACs include the sirtuin family of NAD+-dependent protein deacetylases.
- The novel HDAC11 has a distinct structure and is a class IV HDAC.
The HDACs often participate in the formation of transcriptional repressor complexes, inducing
- chromatin compaction through histone deacetylation, and silencing gene expression.
A Diversity of Partners
A great resource for the research scientist is the National Center for Biotechnology Information (NCBI), your tax dollars at work compiling information about everything molecular. This site should be your first stopping point when trying to learn authoritative information about a new protein or gene that you’re studying. Information at this site helps to underscore two points about KATs and deacetylases: they are social enzymes, always interacting with other proteins, and they are promiscuous, binding to an astounding array of partners. Take, for example, the KAT known commonly as p300. At the NCBI gene link, entering ‘human p300’ finds the gene EP300 (KAT3B), with a summary stating that it associates with the adenovirus protein E1A, acetylates histones, binds CREB, and is a co-activator of HIF-1α (hypoxia-inducible factor 1α). Further down, we find that it binds three different proteins produced by the lentivirus human immunodeficiency virus (HIV)-1. Then, impressively, is a list of over two hundred proteins that have been documented to directly interact with p300 (with links to references and other interactome datasets included). Similarly, the deacetylase HDAC1 is summarized as a histone deacetylase that also interacts with retinoblastoma tumor-suppressor to control cell growth and, together with metastasis-associated protein-2, deacetylates the tumor suppressor p53. Like p300, HDAC1 has an amazing list of partners: it interacts with some 300 proteins, with over 125 of these documented as direct binding partners.
The abundance of protein partners, for both KATs and HDACs, suggests that these enzymes tend to form multimeric complexes. In fact, such complexes serve the critical purpose of positioning the (de)acetylases at specific sites to perform their functions. Certainly, KATs can directly acetylate substrates in vitro. However, KAT activity in vivo is regulated, at least in part, by where it is positioned. For example, the classical model for activation of PPARs (peroxisome proliferator-activated receptors) posits that this receptor heterodimerizes at specific response elements with RXR (retinoid X receptor). In the absence of ligand, the unactivated heterodimer binds co-repressor proteins, such as nuclear receptor co-repressors (NCoR), G-protein pathways suppressor 2 (GPS2), and HDACs (Figure 2). The HDACs help prevent expression of PPAR-specific genes by keeping the neighboring histones deacetylated. The appearance of a ligand for PPAR causes dissociation of the co-repressor proteins followed by the recruitment of co-activators, including PPAR co-activator (PGC-1), CREB binding protein (CBP), and p300. Formation of the PPAR activation complex leads to histone acetylation by CBP and p300, giving rise to altered expression of genes involved in fatty acid metabolism, lipid homeostasis, and adipocyte differentiation. In this example, ligand binding to its receptor causes a large scale switch from a cluster of proteins serving various roles in preventing transcription to a different group designed to facilitate gene transcription.
Acetylation Patterns
In its simplest form acetylation is merely another form of post-translational modification of proteins. A good example is the acetylation of tubulin, which can be deacetylated by HDAC6 or SIRT2. Acetylation of this key microtubule component appears to alter its affinity for kinesin-1 and redirect motor-based trafficking of vesicles.2,3 In short, acetylation changes protein function by adjusting protein-protein interactions. The net ‘global’ acetylation, in this case, may be determined by the balance of overall KAT and HDAC activities.
More commonly, acetylation is targeted to specific proteins and, possibly, specific lysine residues on those protein targets. One way that this can be achieved is by the formation of protein complexes containing either KATs or HDACs, as in the PPAR case described above. The assembly of the complex serves to place the KATs/HDACs near histones, transcription factors, or other targets. Histones, assembled as an octamer core surrounded by DNA, have amino termini that are freely exposed (Figure 3). Positively-charged lysine residues on these tails interact electrostatically with negatively-charged phosphate groups along the DNA backbone. Acetylation reduces these interactions and loosens the DNA, facilitating transcription. Bear in mind that, while it is generally true that histone acetylation increases transcriptional activation, there are exceptions. For example, acetylation of estrogen receptor-α suppresses ligand sensitivity and reduces ligand-induced transcriptional activity.4,5
References
1. Glozak, M.A., Sengubpta, N., Zhang, X., et al. Gene 363, 15-23 (2005).
2. Hammond, J.W., Cai, D., and Verhey, K.J. Curr. Opin. Cell Biol. 20, 71-76 (2008).
3. Gao, Y., Hubber, C.C., and Yao, T.P. J. Biol. Chem. epub ahead of print (2010).
4. Wang, C., Fu, M., Angeletti, R.H., et al. J. Biol. Chem. 276, 18375-18383 (2001).
5. Popov, V.M., Wang, C., Shirley, L.A., et al. Steroids 72, 221-230 (2007).
6. Mellert, H.S. and McMahon, S.B. Trends Biochem. Sci. 34, 571-578 (2009).
7. Yang, X.J. and Seto, E. Mol. Cell 31, 49-461 (2008).
8. Wilson, A.J., Byun, D.S., Popova, N., et al. J. Biol. Chem. 281, 13548-13558 (2006).
9. Vincent, A. and Van Seuningen, I. Differentiation 78, 99-107 (2009).
10. Li, Z., Chen, L., Kabra, N., et al. J. Biol. Chem. 284, 10361-10366 (2009).
From Protein Acetylation: Much More than Histone Acetylation by Brock, T.G.
2 Figures saved
PTM modifications


3-d-genome-map

graphs_superdex-both-high-resolution-size-exclusion-gel-filtration-chromatography