Reprogramming Cell in Tissue Repair
Reporter and Curator: Larry H Bernstein, MD, FCAP
This is a novel concept in regenerative medicine that needs attention.
Lin28 enhances tissue repair by reprogramming cellular metabolism
Shyh-Chang N, Zhu H, Yvanka de Soysa T, Shinoda G, Seligson M T, Tsanov K M, Nguyen L, Asara J M, Cantley L C and Daley G Q.
Stem Cell Transplantation Program,Boston Children’s Hospital and Dana Farber Cancer Institute, Boston; Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School; Harvard Stem Cell Institute;
Manton Center for Orphan Disease Research; Howard Hughes Medical Institute; Department of Medicine, Division of Signal Transduction, Beth Israel Deaconess Medical Center, Boston, MA 02115.
Cell. 7 Nov 2013; 155(4):778-792. http://dx.doi.org/10.1016/j.cell.2013.09.059.
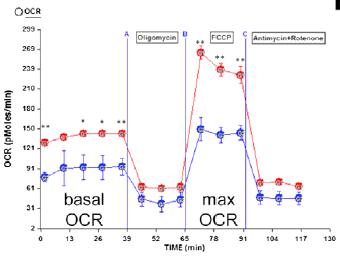
Copyright © 2013 Elsevier Inc. PMID: 23561442 PMCID: PMC3652335
Abstract
In recent years, the highly conserved Lin28 RNA-binding proteins have emerged as factors that define stemness in several tissue lineages. Lin28 proteins repress let-7 microRNAs and influence mRNA translation, thereby regulating the self-renewal of mammalian embryonic stem cells. Subsequent discoveries revealed that Lin28a and Lin28b are also important in organismal growth and metabolism, tissue development, somatic reprogramming, and cancer. In this review, we discuss the Lin28 pathway and its regulation, outline its roles in stem cells, tissue development, and pathogenesis, and examine the ramifications for re-engineering mammalian physiology.
Figure 1. Overview of Molecular Mechanisms Underlying Lin28 Function. From: Lin28: Primal Regulator of Growth and Metabolism in Stem Cells.

Both Lin28a and Lin28b have been observed to shuttle between the nucleus and cytoplasm, binding both mRNAs and pri-/prelet-7. In the nucleus, Lin28a/b could potentially work in tandem with the heterogeneous nuclear ribonucleoproteins (hnRNPs) to regulate splicing, or with Musashi-1 (Msi1) to block pri-let-7 processing. In the cytoplasm, Lin28a recruits Tut4/7 to oligouridylate pre-let-7, and block Dicer processing to mature let-7 miRNA (right, violet). Lin28a also recruits RNA helicase A (RHA) to regulate mRNA processing in messenger ribonucleoprotein (mRNP) complexes, in tandem with the Igf2bp’s, poly(A)-binding protein (PABP), and the eukaryotic translation initiation factors (eIFs). In response to unknown signals and stimuli, the mRNAs are either shuttled into poly-ribosomes for translation, stress granules for temporary sequestering, or P-bodies for degradation, in part via miRNAs and the Ago2 endonuclease (left, orange).
Figure 2. Signals Upstream and Targets Downstream of Lin28 in the Lin28 Pathway. From: Lin28: Primal Regulator of Growth and Metabolism in Stem Cells.

The lin-4 homolog miR-125a/b represses both Lin28a and Lin28b during stem cell differentiation. The core pluripotency transcription factors Oct4, Sox2, Nanog and Tcf3 can activate Lin28a transcription in ESCs and iPSCs, whereas the growth regulator Myc and the inflammation-/stress-responsive NF-κB can transactivate Lin28b. A putative steroid hormone-activated nuclear receptor, conserved from C. elegans daf-12, might also regulate both Lin28a/b and let-7 expression. Downstream of Lin28a/b, the let-7 family represses a network of proto-oncogenes, including the insulin-PI3K-mTOR pathway, Ras, Myc, Hmga2, and the Igf2bp’s. At the same time, Lin28a can also directly bind to and regulate translation of mRNAs, including Igf2bp’s, Igf2, Hmga1, and mRNAs encoding metabolic enzymes, ribosomal peptides, and cell-cycle regulators. Together, this broad network of targets allows Lin28 to program both metabolism and growth to regulate self-renewal.
Figure 3. Potential of Lin28 in Re-Engineering Adult Mammalian Physiology. From: Lin28: Primal Regulator of Growth and Metabolism in Stem Cells.

Lin28a, in conjunction with the pluripotency factors Oct4, Sox2 and Nanog, can reprogram somatic cells into iPSCs. Alone, Lin28a/b can reprogram adult HSPCs into a fetal-like state, and enhance insulin sensitivity in the skeletal muscles to improve glucose homeostasis, resist obesity and prevent diabetes. Emergent clues suggest that optimal doses of Lin28a/b might have the potential to re-engineer adult mammalian tissue repair capacities and extend longevity, although Lin28a/b could also cooperate with oncogenes to initiate tumorigenesis. Future work might elucidate these mysteries.
Cell. 2013 Nov 7;155(4):778-92. doi: 10.1016/j.cell.2013.09.059.


