charged gold nanoparticles influence the structure of DNA and RNA
Larry H. Bernstein, MD, FCAP, Curator
LPBI
How to Control Shape, Structure of DNA and RNA
http://www.technologynetworks.com/news.aspx?ID=185126
Researchers have used computational modelling to shed light on precisely how charged gold nanoparticles influence the structure of DNA and RNA.
The work holds promise for developing applications that can store and transport genetic information, create custom scaffolds for bioelectronics and create new drug delivery technologies.
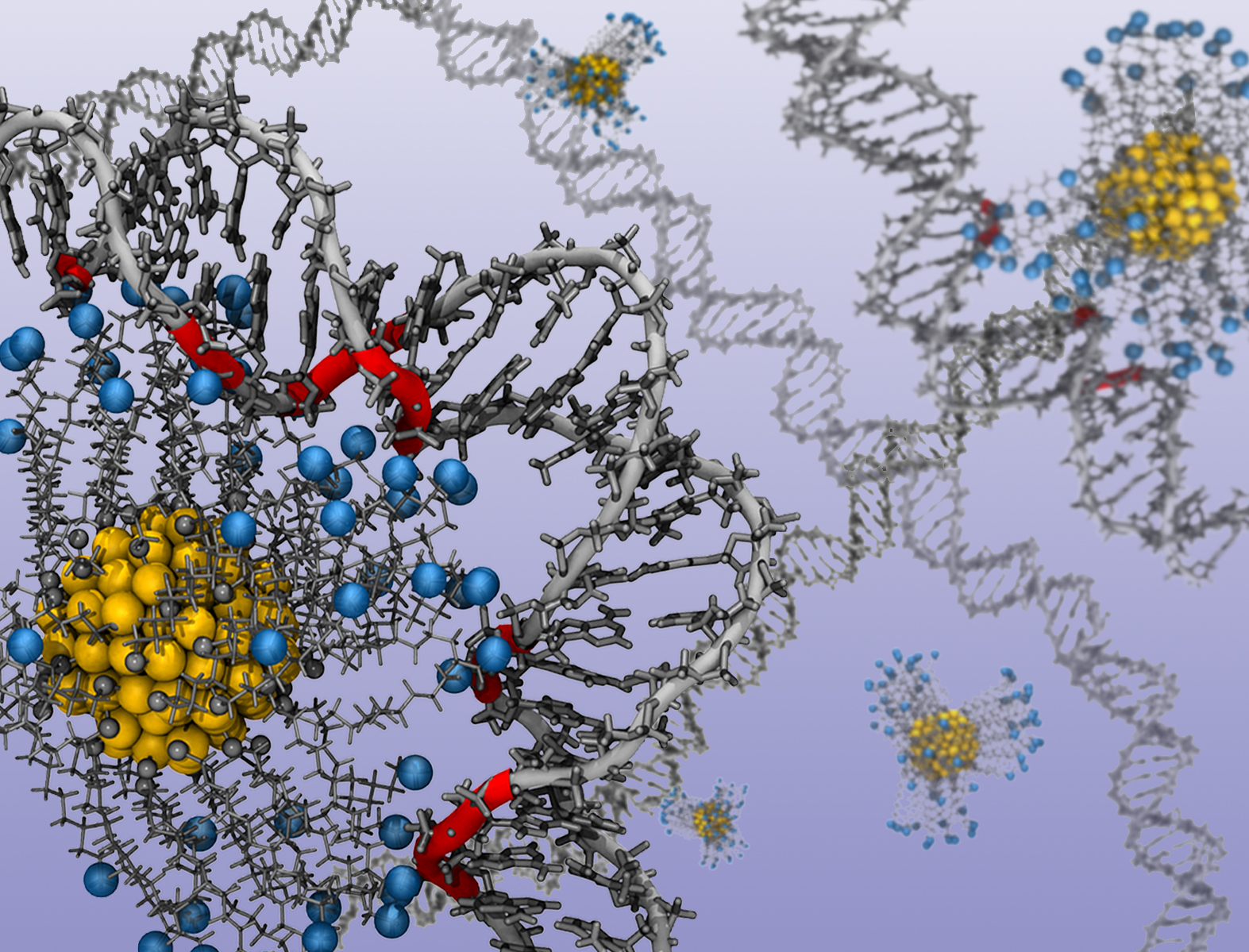
“In nature, meters of DNA are packed tightly into every living cell,” says Jessica Nash, a Ph.D. student at NC State and lead author of a paper on the work. “This is possible because the DNA is wrapped tightly around a positively charged protein called a histone. We’d like to be able to shape DNA using a similar approach that replaces the histone with a charged gold nanoparticle. So we used computational techniques to determine exactly how different charges influence the curvature of nucleic acids – DNA and RNA.”
In their model, the researchers manipulated the charge of the gold nanoparticles by adding or removing positively charged ligands – organic molecules attached to the surface of the nanoparticle. This allowed them to determine how the nucleic acid responded to each level of charge. An animation of a nanoparticle and ligands shaping a strand of DNA is available on the further information link below.
“This will let researchers know what to expect – how much charge they need in order to get the desired curvature in the nucleic acid,” says Yaroslava Yingling, an associate professor of materials science and engineering at NC State and corresponding author of the paper.
“We used ligands in the model, but there are other ways to manipulate the charge of the nanoparticles,” says Abhishek Singh, a postdoctoral researcher at NC State and co-author of the paper. “For example, if the nanoparticles and nucleic acid are in solution, you can change the charge by changing the pH of the solution.”
The work is also significant because it highlights how far computational research has come in materials science.
“Our large-scale models account for every atom involved in the process,” says Nan Li, a Ph.D. student at NC State and co-author of the paper. “This is an example of how we can use advanced computational hardware, such as the GPUs – or graphics processing units – developed for use in videogames, to conduct state-of-the-art scientific simulations.”
The research team is now building on these findings to design new nanoparticles with different shapes and surface chemistries to get even more control over the shape and structure of nucleic acids.
“No one has come close to matching nature’s efficiency when it comes to wrapping and unwrapping nucleic acids,” Yingling says. “We’re trying to advance our understanding of precisely how that works.”
Researchers Detail How to Control Shape, Structure of DNA and RNA
Yaroslava Yingling, Matt Shipman
https://news.ncsu.edu/2015/11/yingling-shaping-dna-2015/
https://news.ncsu.edu/wp-content/uploads/2015/11/Yingling-histone-mimic-FULL.jpg
{kind=link}
Researchers used a computer model of gold nanoparticles and ligands to determine how nucleic acids respond to various charges. In technical language, the image shows the binding of alkyl ligand functionalized gold nanoparticles with protonated amine end groups (the blue spheres) to double stranded DNA. Image credit: Jessica Nash. Click to enlarge.
Researchers at North Carolina State University have used computational modelling to shed light on precisely how charged gold nanoparticles influence the structure of DNA and RNA – which may lead to new techniques for manipulating these genetic materials.
The work holds promise for developing applications that can store and transport genetic information, create custom scaffolds for bioelectronics and create new drug delivery technologies.
“In nature, meters of DNA are packed tightly into every living cell,” says Jessica Nash, a Ph.D. student at NC State and lead author of a paper on the work. “This is possible because the DNA is wrapped tightly around a positively charged protein called a histone. We’d like to be able to shape DNA using a similar approach that replaces the histone with a charged gold nanoparticle. So we used computational techniques to determine exactly how different charges influence the curvature of nucleic acids – DNA and RNA.”
In their model, the researchers manipulated the charge of the gold nanoparticles by adding or removing positively charged ligands – organic molecules attached to the surface of the nanoparticle. This allowed them to determine how the nucleic acid responded to each level of charge. An animation of a nanoparticle and ligands shaping a strand of DNA is available at https://www.youtube.com/watch?v=kNpvPyc2bmc&feature=youtu.be.
Explains what is meant by specific and non-specific binding and how they are measured in a radioligand binding assay.
“This will let researchers know what to expect – how much charge they need in order to get the desired curvature in the nucleic acid,” says Yaroslava Yingling, an associate professor of materials science and engineering at NC State and corresponding author of the paper.
“We used ligands in the model, but there are other ways to manipulate the charge of the nanoparticles,” says Abhishek Singh, a postdoctoral researcher at NC State and co-author of the paper. “For example, if the nanoparticles and nucleic acid are in solution, you can change the charge by changing the pH of the solution.”
The work is also significant because it highlights how far computational research has come in materials science.
“Our large-scale models account for every atom involved in the process,” says Nan Li, a Ph.D. student at NC State and co-author of the paper. “This is an example of how we can use advanced computational hardware, such as the GPUs – or graphics processing units – developed for use in videogames, to conduct state-of-the-art scientific simulations.”
Chromosomes
http://www.nature.com/scitable/topicpage/chromosomes-14121320
Cells package their DNA not only to protect it, but also to regulate which genes are accessed and when. Cellular genes are therefore similar to valuable files stored in a file cabinet — but in this case, the cabinet’s drawers are constantly opening and closing; various files are continually being located, pulled, and copied; and the original files are always returned to the correct location.
Of course, just as file drawers help conserve space in an office, DNA packaging helps conserve space in cells. Packaging is the reason why the approximately two meters of human DNA can fit into a cell that is only a few micrometers wide. But how, exactly, is DNA compacted to fit within eukaryotic and prokaryotic cells? And what mechanisms do cells use to access this highly compacted genetic material?

http://www.nature.com/scitable/content/ne0000/ne0000/ne0000/ne0000/113367985/1_2.jpg
{kind=link}
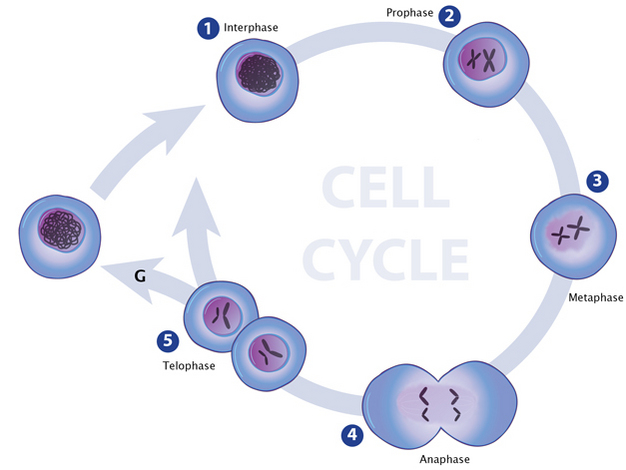
In the first stage of mitosis, an illustration of chromatin looks like a loosely packed clump of thread contained inside a cell’s nucleus (early interphase). In late interphase, the chromatin appears tightly packed into two chromosome-like structures within the nucleus. The structures look like two hockey sticks. In the 2ndstage of mitosis (prophase), true chromosomes have formed inside the nucleus: each chromosome pair looks like two hockey sticks connected to one another where they bend to form an “X” shape. There are two pairs of chromosomes, so the nucleus contains two “X” shaped structures. In stage 3 (metaphase), each chromosome has separated from its partner in each pair, so that four individual chromosomes are visible inside the nucleus. In stage 4 (anaphase), two separate cells are forming, and each contains its own developing nucleus. Inside each nucleus are two chromosomes. In stage 5 (telophase), two individual cells are shown. Inside each cell’s nucleus are two chromosomes. An arrow connects each stage to the one succeeding it. Between telophase and interphase an arrow completes the cycle.
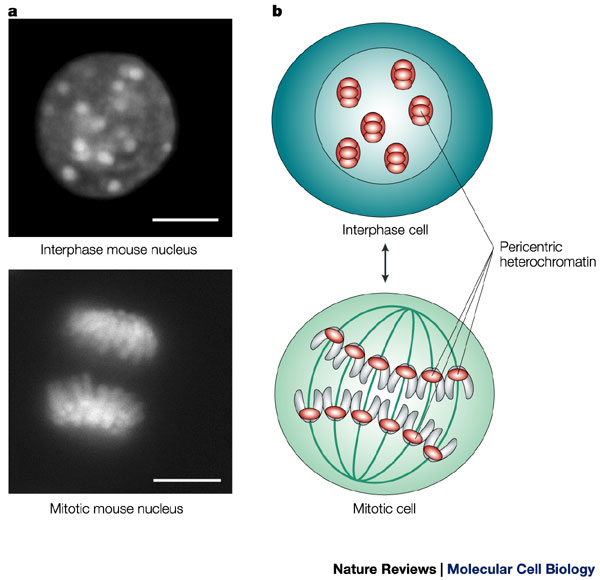
Eukaryotes typically possess multiple pairs of linear chromosomes, all of which are contained in the cellular nucleus, and these chromosomes have characteristic and changeable forms. During cell division, for example, they become more tightly packed, and their condensed form can be visualized with a light microscope. This condensed form is approximately 10,000 times shorter than the linear DNA strand would be if it was devoid of proteins and pulled taut. However, when eukaryotic cells are not dividing — a stage called interphase — the chromatin within their chromosomes is less tightly packed. This looser configuration is important because it permits transcription to take place (Figure 1, Figure 2).

http://www.nature.com/scitable/content/ne0000/ne0000/ne0000/ne0000/14668934/U2.cp2.1_nrm1355-f1.jpg
{kind=link}
Histones carry positive charges and bind negatively charged DNA in a specific conformation. In particular, a segment of the DNA double helix wraps around each histone core particle a little less than twice. The exact length of the DNA segment associated with each histone core varies from species to species, but most such segments are approximately 150 base pairs in length. Furthermore, each histone molecule within the core particle has one end that sticks out from the particle. These ends are called N-terminal tails, and they play an important role in higher-order chromatin structure and gene expression.
…….
Chromatin
Getting a Lot of Information into a Small Package
http://modencode.sciencemag.org/chromatin/introduction
P. J. Horn and C. L. Peterson, Science 297, 1824-1827 (2002)
- Learn how the cell packs six feet of DNA into a nucleus 5 micrometers in length, through the amazing structure known as chromatin.
- Begin to decipher the “histone code” and the mechanisms, beyond transcription, by which the expression of genes is regulated.
- Explore how the modENCODE project has expanded our understanding of how chromatin states affect gene expression, in a Vignette.
Compared to prokaryotes, eukaryotes have enormous genomes. For example, inside each of your cells is a genome containing more than 3 billion base pairs. Lined up, that’s more thansix feet (~2 meters) of DNA.
Fitting that much DNA in a cell nucleus (average width: 5 micrometers) is like fitting a string the length of the Empire State building underneath your fingernail! How do our cells store these extremely long, information-packed molecules?
The answer, in one word: Chromatin.
Chromatin — found only in the cells of eukaryotic organisms — is the complex of DNA and a specialized suite of of proteins that are organized in beads, wrapped with DNA, and the string folded, at multiple scales, to allow the DNA to be packaged into the neat nuclear structures called chromosomes that we see through the microscope. But chromatin is about more than just solving a storage problem. In the past few decades, scientists have come to increasingly appreciate the role of chromatin, and of modifications to the histone proteins central to chromatin structure, in regulating gene expression. The packaging is even as a mechanism for heritable changes in phenotype that don’t involve actual changes to the genome sequence. The study of such mechanisms makes up an exciting, and rapidly growing, branch of biology called epigenetics.
In this section of the site, we’ll take a brief look at chromatin as a regulator of gene expression in some ways complementary to transcription factors. And we’ll see how the techniques used in the modENCODE project have helped to broaden our understanding of the system.
Strings, Beads, and Nucleosomes

To fit our genomes into a tiny cell, the DNA of each chromosome is coiled, compacted, and coiled up some more. At the primary level of compaction, the DNA is wrapped around a group of special proteins called histones. When DNA wraps around a group of histones, it forms a nucleosome. You can think of the system as DNA “thread” wound around a protein “spool”. The first scientists who saw nucleosomes with an electron microscope remarked that they looked like “beads on a string,” though we now know that nucleosomes are more like “string wrapped around beads.”
Each nucleosome is made of four different histones — H2A, H2B, H3, and H4. Two molecules of each histone come together to form an octamer. (Note the prefix “octa” — an octamer is just a complex made of eight proteins.) The DNA string wraps around the histone octamer bead to create the nucleosome.
So the process of fitting all of that DNA into a tiny cell nucleus begins with wrapping the DNA around histones into a nucleosome. But it doesn’t end there — the chain of nucleosomes coils around a central axis to get even more compact. As depicted in the diagram at the right, the packaging actually takes place at a number of scales:
- The DNA wraps around histone octamers to form a “beads on string” fiber approximately 10nanometers (nm) in width.
- The beads-on-string structure in turn coils into a 30-nm-diameter fiber that packs the nucleosomes more closely together.
- During cellular interphase — the period in which the cell is not actively dividing — “scaffold” proteins fold the 30-nm fibers into a somewhat more compact structure to fit within the nucleus.
- During cell division, the chromatin, through the action of additional scaffold proteins, is radically packed and condensed to form the metaphase chromosome that divides and passes the DNA carrying the genetic code to the two daughter cells.
You might be starting to see a problem here. You already know that the genome contains important information — the instructions for making all of the proteins of your cells. If the DNA is all twisted up on itself, how can the cell access that information so it can make proteins?
Getting at the Data: Histone Modification

{kind=link}
At the end of the holiday season, you may store your holiday decorations away in a closet or garage. You don’t need them immediately at hand in the house, since you don’t use them regularly — but you know where they are in storage, and can find them when you need them.
Something very similar is happening in your cells. Cells have ways of opening up the DNA to be read — or of hiding it so that it isn’t read by mistake. And different types of cells have different sets of genes that are accessible for transcription, and other sets of genes that they have closed up and stored away. That’s why your nerve cells are so different from your muscle cells, even though both types of cells contain exactly the same genome with exactly the same DNA code. And, just as you can push or slide boxes out of the way in your garage or closet to get at the holiday decorations when you need them, the cell can remove or slide nucleosomes so that RNA polymerase can get to a gene it needs to transcribe.
One way that cells can open or close a certain gene is by modifying the histones around which the DNA is wrapped. The ends of histone proteins form so-called “tails”, and certain chemical groups can attach to those tails, changing their chemical properties and affecting how the tails interact with the DNA. Sometimes, chemical modification of histones makes a whole region of the genome easier to access –- sort of like putting the boxes you use most often on a lower shelf, or at the front of the closet. Other times, these chemical markers are labels that can be read by other proteins, as you might label a box with a marker to make it easier to find what you need.

Examples of chemical modifications to histones are methylation, acetylation, orphosphorylation (the names just refer to the types of chemical group that is attached – methyl, acetyl, phosphate, etc.). These modifications can be added and removed by special enzymes, so that each type of cell can organize its genome to make it easier to synthesize the proteins that cell type uses most often. These enzymes can also change the modifications as the organism develops, or in response to the environment — for example, in response to a hormone signal or temperature change.
These histone modifications can directly affect how tightly DNA binds to histones. In unmodified histones, the positively charged (basic) histone tails bind very tightly to the negatively charged (acidic) DNA. Some modifications, like the acetylation of a lysine, help neutralize the positive charge of the histone tails. This means the tails bind the DNA less tightly, and the chromatin is more open. Acetylation is thus a histone modification that is associated with genes that are very active, and expressed at high levels. (Check out an animation that illustrates how acetylation weakens the binding between DNA and the histone octamer, making it easier to disassemble the nucleosome.)
As already noted, other histone modifications work differently — instead of directly affecting how tightly the tails bind to DNA, they are instead interpreted by other proteins. These modifications are believed to be part of a histone code, which is “read” by proteins that respond by making the chromatin either more open or more compact, depending on the specific histone modification.
Vignette: modENCODE, the Fly, and Chromatin States

{kind=link}
The modENCODE scientists worked hard to understand how chromatin functions in model organisms like flies and worms. This has uncovered a lot of important and useful information about how chromatin is structured in general, which can be applied to other organisms, including humans. It’s time to drill down into some of this work. Click on the image to the right to explore what detailed work in the fly has taught us about chromatin.
Thought Questions
- Why would modifications like acetylation, that make histones bind to DNA less tightly, affect how active a gene is?
- Imagine a gene that encodes a protein important for developing the axons of neurons. There are histones binding your DNA coding for this gene in every cell. Do you think these histones are more likely to be acetylated in your skin cells, brain cells, or the cells of your immune system?
- What effect do you think histone deacetylases have on gene expression, in general?
- Why would we study chromatin in flies and worms? Can you think of living organisms that might not be helpful for studying chromatin structure?