EpCAM
Writer and Curator: Larry H. Bernstein, MD, FCAP
7.4 EpCAM
7.4.1 Cell-Cell Adhesion
7.4.2 The cell–cell adhesion molecule EpCAM interacts directly with the tight junction protein claudin-7
7.4.3 EpCAM. Structure and function in health and disease
7.4.4 EpCAM contributes to formation of functional tight junction in the intestinal epithelium by recruiting claudin proteins
7.4.5 EpCAM is up-regulated by EGF via ERK1.2 signaling and suppresses human epithelial ovarian cancer cell migration
7.4.6 The tumor-associated antigen EpCAM upregulates the fatty acid binding protein E-FABP
7.4.7 Argonaute and Triman Generate Dicer-Independent priRNAs and Mature siRNAs to Initiate Heterochromatin Formation
7.4.8 Argonaute2 Cleaves the Anti-Guide Strand of siRNA during RISC Activation
7.4.1 Cell-Cell Adhesion
Alberts B, Johnson A, Lewis J, et al.
Molecular Biology of the Cell. 4th edition.
New York: Garland Science; 2002
Cell-Cell Adhesion
http://www.ncbi.nlm.nih.gov/books/NBK26937/
To form an anchoring junction, cells must first adhere. A bulky cytoskeletal apparatus must then be assembled around the molecules that directly mediate the adhesion. The result is a well-defined structure—a desmosome, a hemidesmosome, a focal adhesion, or an adherens junction—that is easily identified in the electron microscope. Indeed, electron microscopy provided the basis for the original classification of cell junctions. In the early stages of cell junction development, however, before the cytoskeletal apparatus has assembled, cells often adhere to one another without clearly displaying these characteristic structures; in the electron microscope, one may simply see two plasma membranes separated by a small gap of a definite width. Functional tests show, nevertheless, that the two cells are stuck to each other, and biochemical analysis can reveal the molecules responsible for the adhesion.
The study of cell-cell junctions and the study of cell-cell adhesion were once quite distinct endeavors, originating from two different experimental approaches—junctions through electron microscopic description, and adhesion through functional tests and biochemistry. Only in recent years have these two approaches begun to converge in a unified view of the molecular basis of cell junctions and cell adhesion. In the previous section, we concentrated on the structures of mature cell junctions. In this section, we turn to functional and biochemical studies of the cell-cell adhesion mechanisms that operate when cells migrate over other cells and when they assemble into tissues—mechanisms that precede the construction of mature cell-cell anchoring junctions. We begin with a critical question for embryonic development: what mechanisms ensure that a cell attaches to appropriate neighbors at the right time?
Animal Cells Can Assemble into Tissues Either in Place or After They Migrate
Many simple tissues, including most epithelial tissues, derive from precursor cells whose progeny are prevented from wandering away by being attached to the extracellular matrix, to other cells, or to both (Figure 19-22). But the accumulating cells do not simply remain passively stuck together; instead, the tissue architecture is generated and actively maintained by selective adhesions that the cells make and progressively adjust.

mechanism by which cells assemble ch19f22
Figure 19-22 The simplest mechanism by which cells assemble to form a tissue. The progeny of the founder cell are retained in the epithelium by the basal lamina and by cell-cell adhesion mechanisms, including the formation of intercellular junctions.
http://www.ncbi.nlm.nih.gov/books/NBK26937/bin/ch19f22.jpg
{kind=link}
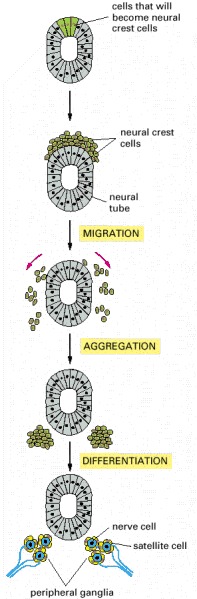
Selective adhesion is even more essential for the development of tissues that have more complex origins involving cell migration. In these tissues, one population of cells invades another and assembles with it, and perhaps with other migrant cells, to form an orderly structure. In vertebrate embryos, for example, cells from the neural crest break away from the epithelial neural tube, of which they are initially a part, and migrate along specific paths to many other regions (discussed in Chapter 21). There they assemble with other cells and with one another to differentiate into a variety of tissues, including those of the peripheral nervous system (Figure 19-23).

more complex mechanism by which cells assemble ch19f23
Figure 19-23 An example of a more complex mechanism by which cells assemble to form a tissue. Some cells that are initially part of the epithelial neural tube alter their adhesive properties and disengage from the epithelium to form the neural crest on the upper surface (more…)
http://www.ncbi.nlm.nih.gov/books/NBK26937/bin/ch19f23.jpg
{kind=link}
Dissociated Vertebrate Cells Can Reassemble into Organized Tissues Through Selective Cell-Cell Adhesion
Embryonic vertebrate tissues are easily dissociated by treating the tissue with low concentrations of a proteolytic enzyme such as trypsin, sometimes combined with the removal of extracellular Ca2+ and Mg2+ with a divalent-cation chelator (such as EDTA). The dissociated cells often reassemble in vitro into structures that resemble the original tissue. Tissue structure is not just a product of history; it is actively maintained and stabilized by the system of affinities that cells have for one another and for the extracellular matrix.
Cells adhere to each other and to the extracellular matrix through cell-surface proteins called cell adhesion molecules (CAMs)— transmembrane adhesion proteins. CAMs can be cell-cell adhesion molecules or cell-matrix adhesion molecules. Some CAMs are Ca2+-dependent, whereas others are Ca2+-independent. The Ca2+-dependent CAMs seem to be primarily responsible for the tissue-specific cell-cell adhesion seen in early vertebrate embryos, explaining why these cells can be disaggregated with Ca2+-chelating agents.
Cadherins Mediate Ca2+-dependent Cell-Cell Adhesion
The cadherins are the major CAMs responsible for Ca2+-dependent cell-cell adhesion in vertebrate tissues. The first three cadherins that were discovered were named according to the main tissues in which they were found: E-cadherin is present on many types of epithelial cells; N-cadherin on nerve, muscle, and lens cells; and P-cadherin on cells in the placenta and epidermis.
These and other classical cadherins are related in sequence throughout their extracellular and intracellular domains. There are also a large number of nonclassical cadherins, with more than 50 expressed in the brain alone.
7.4.2 The cell–cell adhesion molecule EpCAM interacts directly with the tight junction protein claudin-7
Ladwein M, Pape UF, Schmidt DS, Schnölzer M, Fiedler S, Langbein L, et al.
Exp Cell Res. 2005 Oct 1;309(2):345-57.
http://dx.doi.org/10.1016/j.yexcr.2005.06.013
We recently described that in the metastasizing rat pancreatic carcinoma line BSp73ASML the cell–cell adhesion molecule EpCAM, CD44 variant isoforms and the tetraspanins D6.1A and CD9 form a complex that is located in glycolipid-enriched membrane microdomains. This complex contains, in addition, an undefined 20 kDa protein. As such complex formation influenced cell–cell adhesion and apoptosis resistance, it became of interest to identify the 20 kDa polypeptide. This 20 kDa protein, which co-precipitated with EpCAM in BSp73ASML lysates, was identified as the tight junction protein claudin-7. Correspondingly, an association between EpCAM and claudin-7 was noted in rat and human tumors and in non-transformed tissues of the gastrointestinal tract. Co-localization of the two molecules was most pronounced at basolateral membranes, but was also observed in tight junctions. Evidence for direct protein–protein interactions between EpCAM and claudin-7 was obtained by co-immunoprecipitation after treatment of tumor cells with a membrane-permeable chemical cross-linker. The complex, which is located in glycolipid-enriched membrane microdomains, is not disrupted by partial cholesterol depletion, but claudin-7 phosphorylation is restricted to the localization in glycolipid-enriched membrane microdomains. This is the first report on an association between EpCAM and claudins in both non-transformed tissues and metastasizing tumor cell lines.
Fig. 1. EpCAM associates directly with a 20 kDa protein: (A) BSp73ASML cells were lysed in 1% Triton X-100 or 1% Lubrol. Lysates were precipitated with the indicated antibodies. The lysates and the precipitates were incubated with 10ACi [32P]g-ATP for 15 min at 37-C. Precipitates were separated by SDS-PAGE, proteins were transferred to nitrocellulose membranes and exposed for autoradiography. Lubrol lysates and immunoprecipitates of Lubrol lysates with antiCD44- and, most strongly, anti-EpCAM-precipitates contain a phosphorylated protein of 20 kDa. (B) BSp73ASML cells were biotinylated and Triton X-100 lysates were precipitated with anti-EpCAM, anti-D6.1A and anti-CD44v6. After SDS-PAGE and transfer, membranes were blotted with anti-EpCAM, antiD6.1A, anti-CD44v6 and secondary HRP conjugated Ab or Extravidin-HRP. A 20 kDa protein was detected only in the anti-EpCAM immunoprecipitate. (C) BSp73ASML cells were biotinylated or treated with the cross-linker DSP (1 mM, 30 min, RT). Triton X-100 lysates were precipitated with anti-EpCAM. After SDS-PAGE and transfer, proteins were detected with anti-EpCAM and Extravidin. In the biotinylated, and strongly in the DSP lysate, EpCAM and an EpCAMcontaining 55 kDa protein complex was recovered. (D) BSp73ASML cells were treated with the membrane-permeable cross-linker DSP or the membraneimpermeable cross-linker DTSSP (1 mM, 30 min, RT) and Triton X-100 lysates were precipitated with anti-EpCAM. Besides EpCAM itself, a 55 kDa EpCAM-containing complex was only detected after cross-linking with DSP.
Fig. 2. The EpCAM-associated 20 kDa protein is claudin-7: (A) Lysates (1% Triton X-100) of BSp73ASML cells were precipitated with anti-EpCAM or anticlaudin-7 and proteins were separated by SDS-PAGE. Both anti-EpCAM and anti-claudin-7 precipitated EpCAM as well as claudin-7. (B) BSp73ASML cells were incubated with DSP (as above) or DMSO (control), lysed in 1% Triton X-100 and lysates were immunoprecipitated with the indicated antibodies. After SDS-PAGE and transfer, membranes were incubated with anti-EpCAM or anti-claudin-7. The lysates and the EpCAM precipitates contained EpCAM and claudin-7. Only the immunoprecipitates of DSP-treated lysates contained, in addition, the 55 kDa complex, which was stained by anti-EpCAM and anticlaudin-7. (C) BSp73ASML cells were cleared from EpCAM or claudin-7 by 5 repeated rounds of immunoprecipitation. The ‘‘cleared’’ lysates and the precipitates were separated by SDS-PAGE and blotted with anti-EpCAM and anti-claudin-7. After 3–4 rounds of immunoprecipitation, claudin-7 was no longer detectable in anti-EpCAM-precipitates and vice versa. However, even after 5 rounds of immunoprecipitation, the cleared lysates of anti-EpCAM immunoprecipitates contained claudin-7 and cleared lysates of anti-claudin-7 immunoprecipitates contained EpCAM. This implies that only part of EpCAM and claudin-7 molecules associate. Precipitations with anti-transferrin receptor and medium not containing an antibody served as controls. (D) BSp73ASML cells were double stained with anti-EpCAM/anti-mIgG-Cy2 and anti-claudin-7/anti-guinea pig IgG-Cy3. Staining was analyzed using a confocal microscope and digital overlays. Yellow staining indicates co-localization (arrowheads). EpCAM and claudin-7 co-localize in BSp73ASML, though only partly. Colocalization is exclusively seen at the cell membrane. Scale bar: 10 Am.
http://ars.els-cdn.com/content/image/1-s2.0-S0014482705002892-gr2cd.sml
Fig. 3. Co-localization and co-immunoprecipitation of EpCAM and claudin-7 are not restricted to rat tumor cell lines: (A) Lysates of BSp73ASML, Progressor, 804 G and BSp73AS cells were immunoprecipitated with anti-EpCAM. After SDS-PAGE and transfer, membranes were blotted with antiEpCAM and anti-claudin-7. Anti-EpCAM precipitates of BSp73ASML and Progressor cells contained both EpCAM and claudin-7. 804 G cells, which contain EpCAM at a very low level, but not claudin-7 and BSp73AS cells, which are EpCAM- and claudin-7-negative, served as controls. (B) Human pancreatic (6) and colorectal (4) adenocarcinoma lines were lysed and lysates were immunoprecipitated with anti-EpCAM. Proteins in the lysates and immunoprecipitates were separated by SDS-PAGE and stained with anti-EpCAM and anti-claudin-7 after blotting. With the exception of MiaPaca1 cell lysate, which contains neither EpCAM nor claudin-7, all lines express EpCAM and claudin-7. In 8 of these 9 lines (exception SW707), claudin-7 coimmunoprecipitates with EpCAM. (D) Capan-1 and QGP-1 cells were double stained for EpCAM and claudin-7. Co-staining of Capan-1 cells with anticlaudin-7 and anti-EpCAM shows little colocalization of both proteins (arrowheads) at cell–cell contacts. In contrast, in QGP-1 cells, co-staining with anticlaudin-7/anti-guinea pig IgG-Alexa-488 and anti-EpCAM/anti-mIgG-Cy3 shows extensive co-localization of both proteins by confocal laser scanning microscopy. Scale bars: 10 Am. (D) Sections (5Am) of shock frozen human pancreatic normal and carcinoma tissue were stained with anti-EpCAM and anticlaudin-7. One normal pancreatic gland and four pancreatic carcinoma tissues are shown. The pancreatic adenocarcinoma tissues are strongly positive for EpCAM and claudin-7. Normal pancreatic tissue stains weakly with anti-EpCAM and very weakly with anti-claudin-7. Scale bar: 100 Am. Human jejunal (E, F) mucosa was double stained for EpCAM (anti-EpCAM/anti-mIgG-Alexa 488) and claudin-7 (anti-claudin-7/anti-guinea pig IgG-Cy3). Co-staining with anti-claudin-7 and anti-EpCAM reveals extensive colocalization of both proteins along the lateral cell membranes of neighboring enterocytes by confocal laser scanning microscopy. Note claudin-7-staining along the lateral cell membrane (brackets) with only a small fraction of total claudin-7 localizing to the region of the apical junctional complex (arrows). Anti-EpCAM stains the lateral membranes and the brushborder of the enterocytes (arrowheads). Some goblet cells within the upper small intestinal mucosa are indicated by an asterisk. Lamina propria vessels are weakly stained with anti-claudin-7 (empty arrow). Scale bars: (E): 20 Am, (F): 10 Am. (G, H) Electron microscopy of human duodenal mucosal cells after staining with anti-claudin-7 (G) and antiEpCAM (H) is shown at a 16,000-fold magnification. Note immunoreactivity for claudin-7 (G) at apical localization typical for tight junctions (arrow) and at the lateral cell membrane (arrowheads). Immunoreactivity for EpCAM (H) is also observed at the lateral cell membrane (arrowheads) as well as at a typical tight junction localization (arrow) and at microvilli of the brush border (empty arrow).
http://ars.els-cdn.com/content/image/1-s2.0-S0014482705002892-gr3eh.sml
Fig. 4. EpCAM-associated claudin-7 phosphorylation depends on glycolipid-enriched membrane domain localization of the complex: (A) Control or MhCD(20 mM, 20 min, 37-C) treated BSp73ASML cells were lysed in 1% Lubrol and immunoprecipitated with the indicated antibodies. Precipitates were incubated with 10 ACi [32P]g-ATP for 15 min at 37-C and separated by SDS-PAGE. Proteins were transferred to nitrocellulose membranes and exposed to autoradiography. After MhCD treatment, EpCAM and claudin-7 precipitates did no longer contain a phosphorylated 20 kDa protein. (B) BSp73ASML cells were treated as described in (A) and lysates were precipitated with anti-EpCAM or anti-claudin-7. After SDS-PAGE and transfer, membranes were blotted with anti-EpCAM and anti-claudin-7. After MhCD treatment, EpCAM-precipitates contained claudin-7 and claudin-7-precipitates contained EpCAM. (C) To control, whether the phosphorylated 20 kDa protein in panel (A) is claudin-7, BSp73ASML lysates were cleared with anti-EpCAM and anti-claudin-7. Cleared lysated were immunoprecipitated, incubated with 10 ACi [32P]g-ATP, separated by SDS-PAGE and proteins were transferred to nitrocellulose membranes and exposed to autoradiography as described in panel (A). The 20 kDa phosphorylated protein was absent in anti-claudin-7 cleared lysated and was strongly reduced in anti-EpCAM cleared lysates.
7.4.3 EpCAM. Structure and function in health and disease
Schnell U, Cirulli V, Giepmans BN.
Biochim Biophys Acta. 2013 Aug; 1828(8):1989-2001
http://dx.doi.org/10.1016/j.bbamem.2013.04.018
Highlights
- EpCAM structure/ function under (patho)physiological conditions reviewed
- EpCAM is a target for tumor therapy and diagnosis, as well as regenerative medicine
- EpCAM signals in tetraspanin-enriched microdomains and via regulated proteolysis
- These models emerge as generic signaling mechanisms by cell surface molecules
- Mutant EpCAM absent from the cell surface leads to intestinal abnormalities
Abstract
Injection of tumor cells in mice more than 30 years ago resulted in the discovery of an epithelial antigen, later defined as a cell adhesion molecule (EpCAM). Although EpCAM has since evoked significant interest as a target in cancer therapy, mechanistic insights on the functions of this glycoprotein have been emerging only very recently. This may have been caused by the multitude of functions attributed to the glycoprotein, its localization at different subcellular sites and complex posttranslational modifications. Here, we review how EpCAM modifies cell–cell contact adhesion strength and tissue plasticity, and how it regulates cell proliferation and differentiation. Major knowledge derived from human diseases will be highlighted: Mutant EpCAM that is absent from the cell surface leads to fatal intestinal abnormalities (congenital tufting enteropathy). EpCAM-mediated cell proliferation in cancer may result from signaling (i) via regulated intramembrane proteolysis and/or (ii) the localization and association with binding partners in specialized membrane microdomains. New insight in EpCAM signaling will help to develop optimized cancer therapies and open new avenues in the field of regenerative medicine.

EpCAM signaling in cross-talk with E-Cadherin.
Graphical abstract
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-fx1.sml
In multicellular organisms, epithelium lining all external and internal surfaces provides a major barrier that prevents unrestricted exchange between the organism and the external world. The two main types of epithelium are (i) simple epithelium and (ii) stratified epithelium. Simple epithelium consists of a tightly packed and organized monolayer of polarized cells and can generally be found in organs in which absorption, secretion, or filtration occurs, such as in the intestine, the exocrine pancreas and the kidney. Stratified epithelium consists of multiple cell layers, and e.g. can be found in the skin, the cornea and the oral cavity. Characteristic features for simple epithelia are microvilli or cilia on the apical domain, high abundance of cell-cell junctions on the lateral domains, and cell attachment to basal membranes via focal adhesions and hemidesmosomes (Fig. 1). Dysfunction of epithelia due to cellular or molecular abnormalities can give rise to a number of diseases, including cancer.

cell adhesion molecules
Fig. 1. Cell adhesion molecules (CAMs) and junctional complexes are abundant in epithelial tissues. (1) Tight junctions build a seal between adjacent cells and are connected to actin filaments. (2) Adherens junctions are plaques of classical cadherins linked to the actin cytoskeleton. (3) Desmosomes are formed by desmosomal cadherins, linked to intermediate filaments. (4) Gap junctions connect the cytoplasm of two adjacent cells and are linked to microfilaments. (5) Selectins, Ig-superfamily CAMs, but also other CAMs not belonging to the classical families can promote homophilic adhesion outside of junctions. Integrins bind in a heterophilic manner. (6) Focal adhesions (linked to actin) and hemi-desmosomes (linked to intermediate filaments) are cell–matrix junctions that are formed by integrins.
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-gr1.sml
The epithelial barrier function is primarily maintained by tight junctions (TJ; Zonula occludens; Fig. 1), which form an impermeable, or regulated semi-permeable, seal between the apical and basolateral domains of the cells. Cell adhesion is a critical regulator of cell polarity which in turn affects cellular functions as diverse as cell migration, proliferation and differentiation when epithelial cells adhere to neighboring cells, so-called contact inhibition [1]. Contact inhibition is crucial in organogenesis and wound healing [2] and [3]. In cancer, loss of contact inhibition may result in uncontrolled cell movement and proliferation, allowing cells to invade neighboring tissue and to metastasize [4]. Cell adhesion is mediated by transmembrane cell adhesion molecules (CAMs), some of which are subdivided into four families: cadherins, selectins, integrins, and immunoglobulin (Ig)-like CAMs ( Fig. 1). Additionally, there are several CAMs that do not belong to any of these families [5]. In general, CAMs can promote cell–cell adhesion or cell–matrix adhesion by homophilic or heterophilic interaction ( Fig. 1). One cell adhesion protein that does not fit into any of the “classical” families of adhesion molecules, which was discovered as one of the first cancer markers, is the epithelial cell adhesion molecule (EpCAM; CD326). In the 1970s, functional screens were performed for novel tumor-specific cell surface antigens [6] and [7], in which monoclonal antibodies were generated by immunization of mice with cancer cells and production of antibody-secreting hybridomas. Using the newly generated antibodies, EpCAM was identified as a tumor antigen [7]. EpCAM is a cell surface glycoprotein of approximately 40 kDa (detailed below) that is highly expressed in epithelial cancers and at lower levels in normal simple epithelia. Due to its simultaneous discovery by many research groups, EpCAM received a variety of names ( Table 1). Several of these names are still in use, which complicates the linking of studies to the properties of EpCAM [8]. Since 2007, consensus has been reached to use EpCAM as the primary name [8].
Table 1. EpCAM’s various names based on monoclonal antibodies or cDNA clones. *murine EpCAM.
Due to its frequent overexpression in tumors, EpCAM has been of significant interest for the diagnosis and therapy of various epithelial cancers, and several EpCAM-directed antibody- or vaccine-based clinical trials for a wide variety of carcinomas have been conducted [8] and [9]. Apart from being targeted as a molecular biomarker, EpCAM plays a morphoregulatory role in normal epithelia and stem/progenitor cells, as well as it may actively drive tumor progression in cancer cells. High expression of EpCAM is often associated with decreased patient survival [10], [11], [12], [13], [14] and [15]. Recently, mutations in the EpCAM gene have been discovered to be responsible for a disease that affects the intestinal epithelium, leading to intractable diarrhea in infants [16]. While the name EpCAM originates from studies demonstrating that the protein can promote homophilic adhesion [17], forced expression of EpCAM has also been shown to interfere with E-Cadherin-mediated cell adhesion, suggesting a role as a functional antagonist for classic cadherin-mediated cell–cell interactions [18]. In addition, EpCAM has been implicated to be involved in a diversity of processes including cell proliferation, migration, adhesion, differentiation, and cell signaling [5] and [19]. Proliferation induced by EpCAM-overexpression has been shown to correlate with upregulation of cell cycle-related proteins [20]. Although several models of EpCAM signaling have been suggested, the molecular basis of EpCAM-mediated signaling in healthy tissue, cancer, and in conditions such as congenital tufting enteropathy is not yet fully understood. In this review, we provide a general overview of the current knowledge on EpCAM with a focus on functional and structural aspects of this molecule in health and disease.EpCAM: gene, protein, and structure
Human EpCAM is a polypeptide of 314 amino acids (aa), consisting of a large extracellular domain (N-terminal) of 242 aa, a single-spanning transmembrane domain of 23 aa and a short cytoplasmic domain of 26 aa (C-terminal) [27] (Fig. 2 and Fig. 3).

Structure of the EpCAM encoding gene (A) and protein (B)
Fig. 2. Structure of the EpCAM encoding gene (A) and protein (B). The EpCAM encoding gene (GA733-2, epcam) consists of 9 exons located on chromosome 2. The protein is encoded as indicated. SP = signal peptide; TY = thyroglobulin-like domain; TM = transmembrane domain.
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-gr2.sml
The extracellular domain of human EpCAM
EpCAM’s extracellular domain starts with the signal peptide, which is cleaved off predominantly between alanine 23 and glutamine 24 [27], [41] and
[43] (Fig. 3). N-terminal sequencing revealed a minor alternative cleavage of the signal peptide after residue 21 (1%) [43].
Balzar and colleages have suggested that EpCAM’s extracellular domain contains a tandem of epidermal growth factor(EGF)-like repeats (amino acids 27–59 and 66–135), which closely resemble the fourth and fifth EGF-like motif in the rod domain of nidogen, a basement membrane glycoprotein involved in cell–matrix adhesion [44]. However, determination of the exact disulphide bond assignments of the protein by Chong and Speicher has revealed that the first motif in EpCAM’s ectodomain shows a novel pattern of disulphide linkage that does not resemble an EGF-like domain [43]. The second motif does not represent an EGF-like repeat, but it rather resembles a thyroglobulin (TY) type 1A repeat, as proposed in earlier studies [41], [42], [43] and [45] (Fig. 3).
EpCAM cleavage
Proteolytic cleavage of EpCAM at multiple sites has been implicated in its function. Like many other transmembrane proteins, EpCAM contains a signal peptide that is cleaved off by a signal peptidase (see before; Fig. 3, arrow 1). Furthermore, EpCAM can be cleaved between Arg80 and Arg81, located in the second motif (TY-repeat) of the ectodomain [27], [42] and [49] (Fig. 3, arrow 2).
N-terminal cleavage of EpCAM. (A) Schematic representation of EpCAM in its non-cleaved (1) and cleaved state (2). After reduction of disulfide bonds (blue lines), the cleaved part is no longer connected (3). Red arrow: Arg80/81 cleavage site. (B) Lysates of colon cancer cell line SW480 were immunoprecipitated with antibody MH99, recognizing EpCAM motif 1. [35S]Methionine-labeled immunoprecipitates were analyzed under non-reducing (1) and reducing conditions (2) on 15% acrylamide gels (reproduced and adapted from Thampoe et al. [26]; Schnell et al. [58]).

N-terminal cleavage of EpCAM.
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-gr4.sml
EpCAM function
EpCAM and adhesion?
While EpCAM was discovered in a tumor-marker assay, its function remained elusive. Sequence homology to nidogen, a basement membrane protein mediating cell-matrix adhesion, suggested that EpCAM might be involved in cell–matrix or cell–cell adhesion [42]. The name EpCAM -or epithelial cell adhesion molecule—was introduced by Litvinov and colleagues [17], who showed that EpCAM can mediate Ca2 +-independent homophilic cell–cell adhesion in cells that normally lack cell–cell interactions[17]. Expression of EpCAM in EpCAM-negative cells led to aggregation of cells and the formation of cell–cell contacts. In a mix of EpCAM-positive and EpCAM-negative L-cells, cell aggregates mainly consisted of EpCAM-positive cells (90%), pointing to homophilic adhesion. Using a range of EpCAM deletion mutants, Balzar et al. have demonstrated that all the motifs in EpCAM’s extracellular domain are required for the formation of homophilic intermolecular binding and EpCAM accumulation at cell-cell adhesion sites [57]. However, also the short intracellular domain, which might connect EpCAM with the actin cytoskeleton via α-actinin (see Section 2.2), is essential for EpCAM localization at cell-cell contacts and therefore for EpCAM’s ability to mediate adhesion [56].
While being able to mediate homophilic adhesion when introduced in cells that lack their own means of cell-cell interaction, EpCAM is neither structurally related to any of the four major families of CAMs [18] nor associated with any classical junctional structure [61]. In human colon epithelium as well as in epithelial cell lines, EpCAM could not be detected at tight junctions, desmosomes or cell–matrix adhesions, whereas it partly co-localized with E-Cadherin at the lateral membrane [61] and [62].
When compared to E-Cadherin, EpCAM is a relatively weak cell–cell adhesion molecule [17] and [18]. Unlike E-Cadherin expressing cells, which are tightly connected due to adherens junctions, EpCAM expressing cells are only loosely interconnected [18]. Remarkably, when co-expressed in E-Cadherin expressing cells, EpCAM weakens E-cadherin-mediated intercellular adhesion [18]. This weakening is not due to lower E-Cadherin levels or lack of its cell-surface localization, but results from reduced stability of E-Cadherin-mediated adhesion due to disturbance of E-Cadherin association with the cytoskeleton via α-actinin [18].
Although EpCAM is able to promote homophilic cell–cell interactions, its functional antagonism on E-Cadherin-mediated adhesions suggests that it operates as a modulator of the strength of cell adhesion rather than as a promoter of epithelial cell aggregation and junctional complex formation [63]. In this regard, compared to Cadherin-mediated adhesions, EpCAM’s anti-adhesive effect might well be tightly regulated and coordinated during morphogenesis and tissue regeneration [18] but disrupted during neoplasm developmen
EpCAM and proliferation
Besides EpCAM’s increased/de novo expression in metaplasia and neoplasia, its abundant expression on fast proliferating tumors points to a causal relationship and the involvement of EpCAM in growth control. Several in vitro and in vivo studies have shown that overexpression of EpCAM, or parts of it (see below), induces cell proliferation, whereas downregulation of EpCAM decreases cell proliferation [20], [74], [75],[76] and [77]. Induction of EpCAM in some in vitro models results in rapid upregulation of the oncogenic transcription factor c-Myc, and consequently in upregulation of cell cycle-related proteins Cyclin A and E, as well as epidermal fatty acid binding protein [20] and [78]. Using EpCAM mutants, Münz and colleagues have shown that EpCAM´s intracellular domain is not only necessary but also sufficient to induce c-Myc upregulation [20].
Fig. 5. EpCAM as a substrate for regulated intramembrane proteolysis (RIP). Full-length EpCAM (1) is cleaved by ADAM17, releasing EpCAM’s ectodomain (EpEX) (2). EpEX might act as a homophilic ligand for non-cleaved EpCAM, inducing RIP (grey dotted arrow). Following the first cleavage step, EpCAM´s cytoplasmic tail (EpICD) is released due to cleavage by PS-2 (3). EpICD associates with FHL-2 and β-catenin and translocates to the nucleus (4), inducing transcription of EpCAM target genes via LEF-1 consensus sites. Adapted from Maetzel et al. [74]. See main text for details and abbreviations.

EpCAM as a substrate for regulated intramembrane proteolysis (RIP).
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-gr5.sml
Models of EpCAM-mediated signaling
Regulated intramembrane proteolysis
The mechanism of EpCAM-induced proliferation in cancer cells has been shown to involve regulated intracellular membrane proteolysis (RIP) [74]. RIP describes an evolutionarily conserved mechanism combining regulated e.g. ligand-induced ectodomain shedding with the consecutive release of an intracellular domain (ICD) from transmembrane proteins [79] (Fig. 5). Both the shed ectodomain and/or the ICD may activate signaling events. Matrix metalloproteases (MMPs) and the disintegrin-type metalloproteases (ADAMs, ´A disintegrin and metalloprotease´) are often involved in protein ectodomain shedding, whereas, in a second step, γ-secretases including presenilin-1 and 2 (PS-1, PS-2) as active subunits are generating the soluble ICD. Besides proteins such as Notch and amyloid precursor protein (APP), a number of CAMs have been shown to undergo proteolytic cleavage, including E-Cadherin, CD44 and L1-CAM [1], [80], [81], [82] and [83]. While non-cleaved E-Cadherin, e.g., acts as a tumor suppressor, different proteolytic fragments of E-Cadherin have been shown to promote tumor progression [84].
Fig. 6. EpCAM signaling in cross-talk with E-Cadherin. EpCAM may weaken E-Cadherin-mediated adhesion by interrupting the link between E-Cadherin and the actin cytoskeleton. This might in turn result in increased availability of non-bound β-catenin that may be stabilized by association with EpICD. Adapted from Maetzel et al. [74].

EpCAM signaling in cross-talk with E-Cadherin.
http://ars.els-cdn.com/content/image/1-s2.0-S0005273613001338-gr6.sml
Fig. 7. (not shown) EpCAM in tetraspanin-enriched microdomains (TEMs). EpCAM associates with Claudin-7 (1) and is recruited to TEMs (2), where it builds a complex with tetraspanins CD9 and CO-029, and CD44v6. ADAM10, also present in TEMs, might be involved in activating EpCAM signaling as presented in Fig. 6 (3).
Metalloproteases have also been shown to contribute to the tetraspanin web; ADAM10 for example associates with CD9 and other [102]. ADAM10 and ADAM17, which has been shown to be involved in EpCAM cleavage [74], are key players in ectodomain shedding or RIP [103],[104] and [105]. Notably, tetraspanins are involved in the regulation of ADAM activity, and also affect MMPs and associate with γ-secretases (reviewed in Yáñez-Mó et al. [110]), implicating that TEMs might have an important role in the regulation of RIP. Since EpCAM associates with tetraspanins CD9 and CO-029 in TEMs, ADAM10 seems to be a likely candidate for the ectodomain cleavage of EpCAM (Fig. 7).
EpCAM in health & disease
EpCAM in normal tissue and during development
EpCAM expression
In healthy adult tissue, EpCAM is expressed at the basolateral cell membrane of simple, pseudo-stratified, and transitional epithelia. No expression can be detected in the differentiated cells of normal squamous stratified epithelia. In adults, EpCAM is expressed in most organs and glands, with the highest expression in colon. Generally, the level of expression differs between tissues. Typically, in tissues where EpCAM is present it is high in proliferating cells, and low in differentiated cells. EpCAM is neither found in cells of lymphoid origin and bone-marrow-derived cells, nor in mesenchymal, muscular, and neuro-endocrine tissues [25] and [111] (see Balzar et al. [19] and Went et al. [112]).
EpCAM in cancer
Prognostic marker
EpCAM levels are increased in most epithelium-derived tumors. High expression levels of EpCAM usually correlate with poor prognosis, e.g. in breast cancer and ovarian cancer as well as in pancreatic, urothelial, and gallbladder carcinoma [10], [11], [12], [13], [14] and [15]. Exceptional cases are renal and thyroid carcinoma, where high levels of EpCAM have been shown to correlate with increased survival[127] and [128]. In several cancer types EpCAM seems to play a dual role, either promoting or reducing cancer progression [129]. Overviews of EpCAM regarding tumor prognosis are given in van der Gun et al.[129] and Patriarca et al. [130]. EpCAM’s prognostic value has also been assessed in combination with E-Cadherin and tight junction protein Claudin7, which are both functionally related to EpCAM (Section 3.1). However, co-expression analysis did not improve EpCAM’s prognostic value for the presence of nodal metastases in oral and oropharyngeal squamous cell carcinoma [131].
Cancer therapy
In addition to being of prognostic value, EpCAM is an attractive target for tumor diagnosis and therapy because of its tumor-specific overexpression. EpCAM has been used as a target for many immunotherapeutic approaches, including treatment with monoclonal antibodies [9] and the development cancer targeting antibodies [135]. Furthermore, vaccination strategies [136], an EpCAM-specific antibody fragment fused to TRAIL (Tumor necrosis factor–related apoptosis–inducing ligand) [137] and [138], and toxin-conjugated antibody fragments [130], [139] and [140] have been developed. Since 2009, the anti-EpCAM trispecific antibody catumaxomab (Removab) has been authorized for treatment of malignant ascites in cancer patients; other EpCAM-directed antibodies and antibody-based constructs are currently in clinical development [8] and [9].
Lynch syndrome
In addition to its direct involvement in tumor biology (Section 3), EpCAM plays an indirect role in Lynch syndrome (also known as hereditary nonpolyposis colorectal cancer; HNPCC), an autosomal dominant disorder that predisposes to colorectal adenocarcinoma (60–90%), endometrial carcinoma (20–60%), and various other cancers [141], [142] and [143]. Tumors observed in Lynch syndrome families are diagnosed at an unusual early age and can be multiple. Lynch syndrome is caused by mutations in mismatch repair genes, including MLH1 and MHL2 (mutL homolog 1 and mutS homolog 2). These mutations prevent the repair of DNA mistakes occurring during cell divisions that may eventually lead to the development of cancer. Mutations in MLH1 and MSH2 account for about 80% of Lynch syndrome cases [144].
Conclusions, remarks & future perspectives
Although EpCAM has been discovered as a tumor marker more than three decades ago and is since exploited as a tool for tumor diagnosis/prognosis and as a target for cancer therapy, it is still not fully understood how EpCAM is functionally involved in tumor biology. Only in recent years, models of EpCAM-mediated signaling on the molecular level emerged. The discovery of the causal correlation between EpCAM mutations and CTE highlights the importance of EpCAM in normal development. From these studies we learned that EpCAM needs to be present at the cell surface for proper function.
However, the signaling underlying balancing cell proliferation and differentiation remains pleiotropic. Post-translational modification of EpCAM such as glycosylation and proteolytic cleavage at various sites, as well as association with binding partners and localization in TEMs might be crucial for activation and regulation of EpCAM signaling and dictate the functional outcome.
The differences in EpCAM function in malignant versus healthy tissues might be explained by differential expression of the various proteins interacting with EpCAM. Although EpCAM-induced proliferation is normally triggered by increased, or de novo, expression in cancer as well as in developing or regenerating tissues, it remains to be determined if these events are regulated at the level of gene transcription, possibly involving the Wnt pathway [85]. While proteolytic cleavage of EpCAM may induce genes via Wnt, EpCAM itself may trigger a positive feedback loop on its expression [74]. Furthermore, ablation or downregulation of tumor suppressor protein p53 is associated with an increase in EpCAM expression [157], which might explain EpCAM´s frequent overexpression in tumors.
7.4.4 EpCAM contributes to formation of functional tight junction in the intestinal epithelium by recruiting claudin proteins
Lei Z, Maeda T, Tamura A, Nakamura T, Yamazaki Y, Shiratori H, et al.
Dev Biol. 2012 Nov 15; 371(2):136-45
http://dx.doi.org/10.1016/j.ydbio.2012.07.005
Highlights
► Mice lacking EpCAM manifest intestinal barrier defects (55). ► EpCAM colocalizes with claudin-7 at cell–cell junctions (57). ► Claudins such as claudin-7 are down-regulated in EpCAM mutant mouse (68). ► Network of tight junction strands is scattered in the EpCAM mutant (67). ► Barrier function of the intestinal epithelium was impaired in the mutant (74).
Tight junctions (TJs) connect epithelial cells and form a semipermeable barrier that only allows selective passage of ions and solutes across epithelia. Here we show that mice lacking EpCAM, a putative cell adhesion protein frequently overexpressed in human cancers, manifest intestinal barrier defects and die shortly after birth as a result of intestinal erosion. EpCAM was found to be highly expressed in the developing intestinal epithelium of wild-type mice and to localize to cell–cell junctions including TJs. Claudin-7 colocalized with EpCAM at cell–cell junctions, and the two proteins were found to associate with each other. Claudins 2, 3, 7, and 15 were down-regulated in the intestine of EpCAM mutant mice, with claudin-7 being reduced to undetectable levels. TJs in the mutant intestinal epithelium were morphologically abnormal with the network of TJ strands scattered and dispersed. Finally, the barrier function of the intestinal epithelium was impaired in the mutant animals. These results suggest that EpCAM contributes to formation of intestinal barrier by recruiting claudins to cell–cell junctions.
During our search for pluripotency-associated genes in embryonal carcinoma cell lines (Saijoh et al., 1996), we identified EpCAM as a gene that is specifically expressed in undifferentiated cells. Previous studies have revealed that EpCAM contributes to various biological processes including cell signaling, migration, and proliferation ( de Boer et al., 1999, Litvinov et al., 1997 and Maghzal et al., 2010). EpCAM was originally identified as a tumor-associated antigen on the basis of its high level of expression in rapidly growing tumors of epithelial origin ( Trzpis et al., 2007). EpCAM-positive cells have been suggested to serve as cancer stem cells for various human cancers including colorectal and hepatocellular carcinoma (Dalerba et al., 2007 and Yamashita et al., 2009). A proteolytic fragment of EpCAM containing the intracellular domain was recently shown to form a complex with β-catenin and Lef-1 that translocates to the nucleus and activates the transcription of genes related to cell proliferation, such as those for c-Myc and cyclins A and E, and thereby to promote oncogenesis ( Maetzel et al., 2009). EpCAM is also implicated in maintenance of pluripotency in embryonic stem cells ( Gonzalez et al., 2009 and Lu et al., 2010) as well as in somatic stem cells such as hepatic stem cells ( Okabe et al., 2009, Schmelzer et al., 2006, Schmelzer et al., 2007 and Tanaka et al., 2009).
EpCAM (CD326, also known as Tacstd1), a type I transmembrane glycoprotein that contains two EGF-like domains, a single transmembrane domain, and a small intra cellular domain (Winter et al., 2003), has been implicated in homotypic cell–cell adhesion in epithelia (Litvinov et al., 1994), but its precise role in epithelial cell adhesion remains unclear. A recent study of zebrafish lacking EpCAM suggested that this protein functions as a partner of E-cadherin in epithelial morphogenesis during epiboly and skin development (Slanchev et al., 2009). Adhesive structures between adjacent cells, including tight junctions (TJs) and adherens junctions (AJs), are not only required for tissue integrity but also act as signaling structures. While adhesion at AJs is mediated by cadherins, a conserved family of adhesion molecules, TJs contain many adhesive molecules including claudins, occludin, ZO-1 and junctional adhesion molecules (JAMs) (Tsukita et al., 2001).
EpCAM mutant mice exhibit intestinal defects
We previously searched for pluripotency-associated genes in the mouse P19 embryonal carcinoma cell line and its derivatives (Saijoh et al., 1996), and we found that EpCAM is expressed at a high level in undifferentiated P19 cells but is down-regulated in differentiated P19 cells ( Fig. S1A) ( Shimazaki et al., 1993). In the developing mouse embryo, EpCAM is expressed specifically in the inner cell mass of the blastocyst, in the epiblast at embryonic day (E) 5.5 and E6.5, and in the developing gonads at E12.5 ( Fig. S1B). This expression pattern is similar to that of Oct3/4 ( Saijoh et al., 1996 and Scholer et al., 1990), a functional marker of pluripotency.
To determine the physiological function of EpCAM, we generated mice deficient in this protein. Two types of mutant allele were generated: EpCAM– and EpCAMβgeo ( Fig. S2). The phenotypes of EpCAM–/– andEpCAMβgeo/βgeo mice were indistinguishable, suggesting that both alleles are null. In fact, EpCAM protein was not detected in the intestine of EpCAM–/– or EpCAMβgeo/βgeo mice ( Fig. S3A and B). Examination of newborn offspring of heterozygote intercrosses revealed that the frequency of homozygous mutant mice was slightly smaller than expected (22.43% for EpCAM–/– and 13.76% for EpCAMβgeo/βgeo) ( Table S1), suggesting that the mutant alleles are embryonic lethal in a small proportion of mutant homozygotes. A previous study found that EpCAM mutant mice die during embryogenesis as a result of impaired development of the placenta ( Nagao et al., 2009). The homozygous mutant mice in the present study had a body size similar to that of wild-type (WT) mice at birth and were able to take milk from the mother. However, the mutant mice had diarrhea and did not gain body weight, with most dying within 10 days after birth ( Fig. S4). Examination of abdominal organs revealed that the intestine was most prominently affected in the mutant animals. At the newborn stage [postnatal day (P) 0 to P1], the intestine of the mutant mice appeared relatively normal both macroscopically ( Fig. 1A) and histologically ( Fig. S5). At P4, however, bleeding was apparent within the mutant intestine, rendering it brown in color (data not shown). At P6, severe intestinal bleeding and obstruction were evident in the mutant mice ( Fig. 1A). Perforation was also observed at many sites of the mutant intestine. Histological analysis revealed disruption of mucosal architecture and sloughing epithelial cells in the mutant intestine at P5, which were more obvious in the upper intestine, the duodenum and jejunum ( Fig. 1B).
In the normal intestine, Ki67-positive (proliferating) cells are localized to the intervillus domains between villi at both embryonic and adult stages (Fig. S6A) (van der Flier et al., 2009). The number of Ki67-positive cells in intervillus domains of EpCAM mutant mice at E18.5 was similar to that in WT animals throughout the intestine (Fig. S6A). Intestinal stem cells, which express Lgr5, are also located in intervillus domains ( Kim et al., 2012). Again, the number of Lgr5-expressing cells in the intestine of mutant mice was similar that in WT controls ( Fig. S6B). These results thus suggested that intestinal stem and progenitor cells as well as regulation of their cell cycle are maintained in the intestine of EpCAM mutant mice.
EpCAM is highly expressed in the developing intestine
In WT mice, EpCAM mRNA was detected in the developing gut from E9.5 to E15.5 as well as throughout the intestine, from the duodenum to the colon, at E18.5 ( Fig. S8). EpCAM protein was found in villi and intervillus domains and was localized to cell–cell junctions of the intestinal epithelium at E18.5 and at P0 (Figs. 1C and 2A). As expected, EpCAM protein was not detected in the intestine of EpCAM–/– orEpCAMβgeo/βgeo mice ( Fig. 1C, Fig. S2B). Staining of the intestinal epithelium of WT mice with both antibodies to EpCAM and either those to E-cadherin or those to ZO-1, markers of adherens junctions (AJs) and tight junctions (TJs), respectively, revealed that EpCAM is localized to TJs, AJs and the lateral membranes of the epithelial cells lining the mouse intestines ( Fig. 2A).
Fig. 2. Colocalization of EpCAM with claudin-7 in the intestine. EpCAM localized not only to apical junctions containing E-cadherin and ZO-1 but also to the lateral membrane of neighboring epithelial cells. It also colocalized extensively with claudin-7 along the lateral cell membrane of epithelial cells, indicating that claudin-7 is not a specific TJ marker.
Claudin proteins are down-regulated in the intestine of EpCAM mutant mice
We next examined the expression of proteins that contribute to cell–cell junctions, including that of claudin-7, which interacts with EpCAM in cultured tumor cells (Kuhn et al., 2007 and Ladwein et al., 2005) and is one of the major claudin proteins expressed in the intestine (Fujita et al., 2006) (Fig. 3). In EpCAM mutant embryos at E18.5, however, claudin-7 was down-regulated to undetectable levels in all regions of the intestine examined, including the duodenum, jejunum, ileum, and colon (Fig. 3A, Fig. S3B). Claudin-7 was thus undetectable not only at cell–cell junctions but also in the interior of intestinal cells. Furthermore, whereas claudin-7 is expressed in the epithelium of the normal developing gut at E12.5, E13.5, and E14.5, it was undetectable in the developing gut of the mutant mice at any stage examined (Fig. 3B).
Mice lacking claudin-7 manifest renal salt wasting and chronic dehydration and intestinal anomalies similar to those of EpCAM mutant mice (Ding et al., 2012 and Tatum et al., 2010). To examine whether deficiency of claudin-7 alone is responsible for intestinal defects of the EpCAM mutant mouse, we examined the expression of additional junctional proteins, including other claudins as well as AJ and desmosome proteins. Claudin-3 is predominantly expressed in intervillus domains of the developing intestine in WT mice; its abundance was reduced in the intestine of EpCAM mutant mice at P0, however, with this effect being most prominent in the duodenum and jejunum (Fig. 4A). Claudin-15, which is normally expressed at TJs of the intestinal epithelium (Fujita et al., 2006), was also down-regulated in the mutant intestine, especially in the duodenum and jejunum (Fig. 4B). Whereas claudin-2 was detected in intestinal villi and intervillus domains of WT mice at P0, it was down-regulated in the mutant intestine (Fig. 4C).
EpCAM interacts with claudins in the intestinal epithelium
Claudin-7, like EpCAM, is abundantly localized to lateral membrane while other claudins localize mainly to apical TJs in the intestinal epithelium. Whereas the abundance of claudin-7 protein was greatly reduced in the intestine of EpCAM mutant mice, that of claudin-7 mRNA was not ( Fig. 3C), suggesting that the down-regulation of claudin-7 expression occurs at a post-transcriptional level.
EpCAM and claudin-7 were found to be coexpressed and colocalized in epithelia of lung, pancreas, stomach, and kidney in WT embryos (Fig. 5A). Claudin-7 was not detected in epithelia of the lung, stomach, and pancreas of EpCAM mutant mice, but it was detected in the kidney of EpCAM mutant mice albeit at a reduced level (Fig. 5B).
Barrier function is impaired in the EpCAM mutant intestinal epithelium
Given that TJs are required for paracellular barrier function (Tsukita and Furuse, 2002 and Tsukita et al., 2008), we next examined the barrier function of the intestinal epithelium. Injection of sulfo-NHS-biotin, a probe that physically labels cell membrane proteins, into the intestinal lumen of E18.5 WT embryos revealed that the resulting biotin signal at the basal membrane of epithelial cells in the jejunum was less intense than that at the apical membrane (Fig. 6A). In contrast, similar analysis of the intestine of EpCAM mutant mice showed that the biotin signal at the basal membrane was similar to that at the apical membrane (Fig. 6A). In addition, the intensity of the biotin signal at the lateral membrane of the intestinal epithelium was higher for the mutant mice than for WT mice (Fig. 6A). These results thus suggested that the barrier function of TJs is impaired in the EpCAM mutant animals.
Claudin-2 and claudin-15 are responsible for paracellular permeability of Na+ (Tamura et al., 2011). In EpCAM mutant mice in which claudin-2 and claudin-15 are down-regulated (Fig. 4), NaCl-dilution potential was lowered (Fig. 6C). Na+-selective paracellular permeability was reduced while Cl−-selective permeability remained normal (Fig. 6C), like in the claudin-15 mutant mouse (Tamura et al., 2011).
Structural anomalies of tight junctions in the intestinal epithelium of EpCAM mutant mice
Freeze-fracture electron microscopy revealed the branching network of sealing strands of TJs apparent in the intestinal epithelium of WT mice to be scattered and dispersed in that of the mutant mice (Fig. 7A). Analysis of ultrathin sections (Fig. 7B) revealed the presence of cell–cell junctions including TJs, AJs, and desmosomes in the intestinal epithelium of WT mice. In the intestinal epithelium of EpCAM mutant mice, however, TJs expanded and appeared markedly dispersed and less well organized (Fig. 7B). On the other hand, the structure of AJs appeared to be normal in the mutant animals (Fig. 7A). These structural anomalies of TJs likely account for the impaired barrier function of the mutant intestinal epithelium (Fig. 6).
We have uncovered a role for EpCAM in formation of functional TJs of the intestinal epithelium. TJs form a barrier that separates the apical from the basolateral membrane of the epithelium, allowing the selective passage of ions and solutes (Van Campenhout et al., 2011). Members of the claudin family, which includes at least 27 proteins in human and mouse (Mineta et al., 2011, Tsukita and Furuse, 2002, Tsukita et al., 2008, Van Itallie and Anderson, 2006 and Yu and Turner, 2008), are essential components of TJs (Angelow et al., 2008). They have been shown to determine not only the barrier function but also the paracellular permeability of epithelial cell sheets in culture (Colegio et al., 2002, Furuse and Tsukita, 2006 and Van Itallie and Anderson, 2006). The combined down-regulation of claudins-2, 3, 7, and 15 observed in the intestinal epithelium of EpCAM mutant mice in the present study might thus account for the impaired barrier function in the intestine of these animals. In fact, a recent study (Ding et al., 2012) reported that mutant mouse lacking claudin-7 manifests intestinal defects similar to those of the EpCAM mutant mice. With the barrier function impaired, large molecules such as proteases may have penetrated the mutant intestinal epithelium, which may be the reason for the intestinal erosion found in the EpCAM mutant mouse. In this regard, it is interesting to note that the intestinal defects of EpCAM mutant mice were more severe in the upper parts of the intestine ( Fig. 1B), where proteases are more abundant.
EpCAM is expressed in embryonic and somatic stem cells and is implicated in pluripotency. Although our EpCAM mutant mice appear to develop normally until birth, a maternal contribution of EpCAM might have prevented early embryonic death of the mutant animals. A minor proportion (3/50, 6%) of the mutant mice survives until adulthood, with both males and females being fertile. EpCAM therefore appears to be dispensable for germ cell formation, even though it is highly expressed in primordial germ cells (Fig. S1B). Such EpCAM mutant mice that survive until adulthood should prove useful for investigation of the possible role of EpCAM in maintenance of somatic stem cells such as those in the liver (Okabe et al., 2009 and Schmelzer et al., 2006) as well as that of cancer stem cells.
Claudin-7 was the most extensively down-regulated claudin in the intestinal epithelium of EpCAM mutant mice. However, the combined down-regulation of multiple claudins may thus underlie the intestinal defects of EpCAM mutant mice. Claudin-7 localizes to TJs and the lateral membrane of epithelial cells in many mouse and human tissues (Ladwein et al., 2005 and Tatum et al., 2010). What then is the mechanism by which claudin-7 is down-regulated to undetectable levels in the intestine of EpCAM mutant mice? Given that the abundance of claudin-7 mRNA was maintained in the intestine of these animals, the reduction in the amount of claudin-7 protein appears to occur at a posttranscriptional step. Previous studies (Kuhn et al., 2007 and Ladwein et al., 2005) and our present results suggest that EpCAM associates with claudin-7 in various cultured cells and tissues. Interaction with EpCAM may result in the recruitment of claudin-7 to AJs and TJs, such that, in the absence of EpCAM, claudin-7 becomes mislocalized and susceptible to degradation by the ubiquitin-proteasome system.
7.4.5 EpCAM is up-regulated by EGF via ERK1.2 signaling and suppresses human epithelial ovarian cancer cell migration
Fan Q, Cheng JC, Qiu X, Chang HM, Leung PC
Biochem Biophys Res Commun. 2015 Feb 13; 457(3):256-61
http://dx.doi.org/10.1016/j.bbrc.2014.12.097
Highlights
- EGF up-regulates EpCAM in human EOC cells. • ERK1/2 is involved in EGF-up-regulated EpCAM expression. • Overexpression of EpCAM inhibits EOC cell migration. • Knockdown of EpCAM stimulates EOC cell migration.
Although epithelial cell adhesion molecule (EpCAM) is overexpressed in human epithelial ovarian cancer (EOC), some contradictory results have been reported regarding the correlation between EpCAM overexpression and patient survival. In addition to this controversy, the function and regulation of EpCAM in EOC remain largely unknown. Here, we show that epidermal growth factor (EGF) up-regulates EpCAM expression by activating ERK1/2 signaling in a human EOC cell line, SKOV3. Additionally, EpCAM overexpression suppresses not only basal but also EGF-stimulated SKOV3 cell migration, whereas EpCAM knockdown increases both basal and EGF-stimulated cell migration in another human EOC cell line, OVCAR4. This study demonstrates the regulation of EpCAM and its role in mediating the effects of EGF on human EOC cell migration.
7.4.6 The tumor-associated antigen EpCAM upregulates the fatty acid binding protein E-FABP
Münz M, Zeidler R, Gires O
Cancer Lett. 2005 Jul 8; 225(1):151-7
http://dx.doi.org/10.1016/j.canlet.2004.11.048
The epithelial cell adhesion molecule, EpCAM, is a transmembrane glycoprotein associated with both benign and malignant proliferation. In cancer cells, expression levels of this tumour-associated antigen correlate positively with the grade of dysplasia and are also a negative prognostic factor for breast cancer patients. De novo expression of EpCAM resulted in the rapid upregulation of the proto-oncogene c-Myc along with enhanced cell proliferation and metabolism. Here, we analyzed the effects of EpCAM onto the proteome of epithelial cells. The epidermal fatty acid binding protein, E-FABP, was identified as a new EpCAM-regulated protein. E-FABP is a major target of c-Myc and was rapidly upregulated upon induction of EpCAM. Additionally, E-FABP levels correlated with the amount of EpCAM in permanent squamous cell carcinoma lines and in vivo in primary head and neck carcinomas. Taken together, these results provide further evidence for the direct involvement of EpCAM in signalling processes, gene regulation, and cellular metabolism supporting its important role in tumour biology.
7.4.7 Argonaute and Triman Generate Dicer-Independent priRNAs and Mature siRNAs to Initiate Heterochromatin Formation
Marasovic M, Zocco M, Halic M
Mol Cell. 2013 Oct 24; 52(2):173-83
http://dx.doi.org/10.1016/j.molcel.2013.08.046
Highlights • Argonaute recruits Triman to generate Dicer-independent priRNAs and mature siRNAs • Longer sRNAs are inefficient in guiding Argonaute to slice complementary targets • Small RNA trimming is required for establishment of centromeric heterochromatin • Triman is required for formation of facultative and ectopic heterochromatin
RNAi is a conserved mechanism in which small RNAs induce silencing of complementary targets. We have previously identified priRNAs, a class of Dicer-independent small RNAs in fission yeast. The mechanism by which Dicer-independent small RNAs are generated is not well understood in any species. Here we reconstitute the final steps of priRNA and siRNA biogenesis in vitro. We identify the 3′-5′ exonuclease Triman and demonstrate that Argonaute, loaded with longer RNA precursors, recruits Triman to generate mature priRNAs and siRNAs. We show that priRNA and siRNA trimming is required for de novo assembly of heterochromatin at centromeric repeats and the mat locus and for maintenance of heterochromatin at developmental genes. Furthermore, in rrp6Δ cells RNAi targets diverse genes in a Triman-dependent way, indicating that the exosome protects the genome from spurious RNAi. Our results suggest that Argonaute association with RNA degradation products generates priRNAs and triggers RNAi in a process of transcriptome surveillance.

Argonaute recruits Triman to generate Dicer-independent priRNAs and mature siRNAs
http://www.cell.com/cms/attachment/2024627216/2044258742/fx1.jpg
{kind=link}
Regulation of genome expression is essential for many cellular processes, including cell proliferation, differentiation, development, and viability. Small RNA silencing pathways are involved in the cellular control of gene expression and protection of the genome against mobile repetitive DNA sequences, retroelements, and transposons (Aravin et al., 2007, Ghildiyal and Zamore, 2009 and Mello and Conte, 2004). The Argonaute family of proteins binds small RNAs that interact with target RNAs via base-pairing interactions. Small RNAs promote DNA and chromatin modifications, translational inhibition, and degradation of complementary RNAs to induce gene silencing (Ghildiyal and Zamore, 2009, Hutvagner and Simard, 2008, Law and Jacobsen, 2010 and Moazed, 2009).
piRNAs, the most studied class of Dicer-independent small RNAs, are associated with the PIWI clade of Argonaute proteins and are expressed in germline cells to suppress transposon activity (Aravin et al., 2006, Girard et al., 2006, Grivna et al., 2006 and Watanabe et al., 2006). Primary piRNAs arise from long single-stranded precursors, and their biogenesis is dependent on PIWI proteins (Siomi et al., 2011). It is unknown how primary piRNAs are generated and how PIWI proteins mediate their biogenesis. The slicer activity of two PIWI proteins generates the 5′ end of secondary piRNAs by the ping-pong cycle, a feedforward amplification mechanism. However, nucleases involved in generation of the 3′ end have not yet been identified (Brennecke et al., 2007 and Gunawardane et al., 2007). A class of Dicer-independent small RNAs associates with Ago1/2 in human cells and has similar genomic distribution to S. pombepriRNAs ( Valen et al., 2011), suggesting that priRNA function might be conserved in higher eukaryotes. Dicer-independent small RNAs have been described in many species; nevertheless, little is known about their biogenesis and function.
Here we identify a nuclease involved in small RNA processing and provide the first mechanistic explanation of how Dicer-independent small RNAs are generated. We show that the previously uncharacterized gene Triman is required for generation and 3′ end processing of Dicer-independent priRNAs. priRNAs are generated by a trimming/protection mechanism that requires joint activity of Argonaute and Triman. This resembles biogenesis and 3′ end processing of piRNAs in the animal kingdom, where similar nuclease has been suggested (Kawaoka et al., 2011). To our surprise, we found that Triman is required also for 3′ end processing of siRNAs. We show that longer small RNAs accumulating in tri1Δ cells were less competent in guiding Argonaute to slice complementary targets and to maintain heterochromatin at developmental genes. Furthermore, we show that centromeric heterochromatin cannot be reestablished in tri1Δ cells, indicating that priRNA and siRNA trimming is required for establishment of heterochromatin at the centromeric region. In the absence of siRNAs, priRNAs can induce very low levels of H3K9me at centromeric repeats and thus might provide specificity for heterochromatin formation. The specificity lies in a prevalence of antisense transcription and antisense priRNAs that are generated at centromeric repeats. We elaborate further that an increase in antisense transcription can recruit RNAi and demonstrate that, in exosome mutant cells, RNAi-dependent heterochromatin was established at diverse genes, including developmental genes, a proteosomal subunit, and ribosomal proteins. This ectopic nucleation of siRNA generation and heterochromatin formation was Triman dependent, indicating that Dicer-independent priRNAs are required for de novo siRNA generation. Our data show that exosome-mediated RNA quality control protects the genome from spurious RNAi. We propose that Argonaute association with RNA degradation products generates priRNAs and triggers RNAi and heterochromatin formation in a process of transcriptome surveillance.
Figure 1. Triman Is Required for Biogenesis of Dicer-Independent priRNAs
(A) Detection of Argonaute-associated centromeric priRNAs in indicated mutant cells by splinted ligation. Five different centromeric small RNAs mapping to dg transcripts were assayed. One-third and one-ninth of Argonaute-associated RNAs fromdcr1Δ cells were assayed.
(B) Autoradiograph of denaturing polyacrylamide gel showing Argonaute-associated RNAs purified from wild-type,SPBC29A10.09cΔ, dcr1Δ, and dcr1ΔSPBC29A10.09cΔ cells. Quantification is based on five independent biological replicates. Lower panels show western blot detection of FLAG-Argonaute protein in immunoprecipitation assays and total cell lysates. Argonaute protein is reduced in dcr1Δ and dcr1ΔSPBC29A10.09cΔ cells in both input and IP, suggesting that empty Argonaute is targeted for degradation, which is consistent with recent findings in plants and mammalian cells ( Frohn et al., 2012 and Gibbings et al., 2012). (C) Length distribution of Argonaute-associated priRNAs and siRNAs in indicated cells. Twenty- to twenty-seven-nucleotide-long Argonaute-associated small RNAs were analyzed by high-throughput sequencing. See also Figure S1.
The observation that mature priRNAs are almost eliminated in SPBC29A10.09cΔ cells prompted us to examine the length and distribution of remaining priRNAs by small RNA sequencing. In wild-type cells, Argonaute-associated siRNAs and priRNAs were predominately 22–23 nt long and had strong preference for 5′ uracil ( Halic and Moazed, 2010). Sequencing of small RNAs in SPBC29A10.09cΔ cells showed that both Argonaute-associated priRNAs and siRNAs were longer: in SPBC29A10.09cΔ cells, remaining priRNAs were 23–27 nt long, while siRNAs had a broad length distribution of 20–27 nt ( Figure 1C). The length of Argonaute-associated centromeric siRNAs in SPBC29A10.09cΔ cells is similar to the length of centromeric siRNAs purified from total RNA fraction in wild-type cells ( Figure S1D). This is in agreement with previous findings that S. pombe Dicer does not generate siRNAs of a very defined length (Colmenares et al., 2007) and suggests that Argonaute binds longer siRNAs that are then trimmed to the mature length. Low residual amounts of priRNAs in dcr1ΔSPBC29A10.09cΔ cells had a strong preference for 5′ uracil, confirming their association with Argonaute ( Frank et al., 2010 and Halic and Moazed, 2010) (Figure S1E).
Our results suggest that SPBC29A10.09c generates priRNA and trims siRNAs to the mature length. Therefore, we named the previously uncharacterized gene triman (tri1). The catalytic domain of Triman belongs to the CAF1 family of 3′-5′ exonucleases, which are conserved in most eukaryotes ( Table S1). Canonical Caf1 nuclease had no effect on small RNA biogenesis in fission yeast ( Figure S1F)
Triman Is a 3′-5′ Exonuclease
Although Triman is homologous to the CAF1 family of deadenylases, it was also capable of degrading non-poly(A) RNA sequences ( Figures 2A and S2B–S2F). β-eliminated and 2′-O-methyl-containing RNAs were not degraded, confirming that Triman is a 3′-5′ exonuclease and needs the 2′OH group for its activity (Figures S2D–S2F). Triman nuclease activity was observed on both short RNA templates (22–30 nt) as well as longer RNA templates (100–130 nt) ( Figures 2A and S2B–E). However, Triman could not degrade either long dsRNA templates or double-stranded siRNAs ( Figure S2G and S2H).
Argonaute and Triman Cooperate to Generate priRNAs and Mature siRNAs
The most probable factor that cooperates with Triman to generate Dicer-independent priRNAs and mature siRNAs is Argonaute. To reconstitute the final step of Triman-dependent small RNA biogenesis in vitro, we loaded 5′ 32P-labeled small RNAs of various lengths onto resin-bound FLAG-Argonaute. Unbound small RNAs were washed away from the resin-bound Argonaute, and Argonaute-associated small RNAs were incubated with Triman. While incubation of 30-nt-long RNA with Triman resulted in a complete degradation of the RNA, incubation of Argonaute-associated 30 nt RNA generated specific product migrating just above the 22 nt marker (Figure 2B). Similar results were observed with 26- and 65-nt-long RNAs associated with Argonaute (Figure S3A and B). In contrast, incubation of 22 nt Argonaute-associated small RNA with Triman did not change the length of the RNA (Figure 2B). Unbound 22 nt RNA was completely degraded under the same conditions. These results indicate that Argonaute associates with longer priRNA and siRNA precursors, which are then processed to the mature length by Triman.
To determine whether tri1Δ cells have a defect in heterochromatin establishment, we perturbed existing heterochromatin structure with the histone deacetylase inhibitor trichostatin (TSA) ( Ekwall et al., 1997). Wild-type cells were fully functional in establishment of heterochromatin at the centromeric ura4+ reporter gene; however, tri1Δ cells show >10-fold reduction in the efficiency of heterochromatin re-establishment (Figure 4A). This suggests that Triman might have function in the nucleation of RNAi.
Our data show that trimming of priRNAs and siRNAs is necessary for de novo assembly of centromeric heterochromatin. Although maintenance of heterochromatin at the mat locus is RNAi independent, it has been proposed that heterochromatin establishment is dependent on RNAi ( Hall et al., 2002). Consistent with this finding, heterochromatin was not established at the mat locus in clr4Δdcr1Δtri1Δ/clr4+dcr1+ cells, indicating that Triman is also required for de novo assembly of heterochromatin at this locus ( Figure 4D). Reintroduction of clr4 gene intoclr4Δdcr1Δtri1Δ cells that lack siRNAs and priRNAs did not induce H3K9me at centromeric dg repeats, indicating that small RNAs are essential for de novo establishment of centromeric heterochromatin (Figure 4E).
While priRNAs are amplified by a RDRC and Dicer-based mechanism, in the case of piRNAs in fly and mammalian germline cells, the amplification is carried out by the ping-pong mechanism. Ping-pong amplification also requires 3′ end processing of secondary piRNAs by an unknown nuclease ( Brennecke et al., 2007, Gunawardane et al., 2007 and Ishizu et al., 2012). Here, we identified a nuclease required for 3’ end formation of Dicer-independent small RNAs in S. pombe. Our results show that priRNAs are generated and siRNAs achieve their mature size by a trimming/protection mechanism that involves Argonaute and Triman. Argonaute binds longer priRNA and siRNA precursors and recruits Triman to process them to the final length. Triman belongs to the CAF1 family of exonucleases that are conserved in most eukaryotes, indicating that Triman homologs might be involved in biogenesis of Dicer-independent small RNAs in other species as well. We propose that priRNA biogenesis in fission yeast resembles biogenesis of primary piRNAs and 3’ end formation of secondary piRNAs in the animal kingdom.
We show that 3’ end processing of siRNAs and priRNAs is required for their function in guiding Argonaute to slice target RNAs. Our results suggest that longer small RNAs interact with Argonaute in a different mode that does not allow proper positioning of the target RNA in an active site. Likely interaction of the 3’ end of slightly longer small RNA (23–27 nt) with the PAZ domain of Argonaute positions RNA away from the catalytic site and prevents the cleavage (Wang et al., 2008). RNAs that are longer than 28 nt bind Argonaute with much lower affinity and eventually dissociate. In this case, the 3′ end of small RNA is likely too far from the PAZ domain of Argonaute and cannot bind it anymore. The MID domain of Argonaute binds the 5’ end (Wang et al., 2008) of small RNA, and trimming to the 22 nt length is required for the correct interaction of the 3’ end with the PAZ domain.
Our results suggest that priRNAs and Argonaute surveillance is one possible mechanism by which repetitive and foreign DNA elements can be recognized by a host cell. Changes in antisense transcription or antisense transcript levels could recruit RNAi to targeted locus to control gene expression. This resembles recognition of transposable and other foreign DNA elements that frequently generate high levels of antisense transcripts after incorporation into genomic DNA. The uncontrolled antisense transcription is scanned by an Argonaute and priRNAs in a process of genome surveillance and triggers silencing. Our results suggest that Argonaute associates with random degradation products to generate priRNAs, which in turn could nucleate RNAi (Figure 7). In this regard, Argonaute transcriptome surveillance may play an essential role in genome defense and initial recognition of transposable and other invading genomic elements.

Model of priRNA Biogenesis and RNAi Nucleation
http://ars.els-cdn.com/content/image/1-s2.0-S1097276513006461-gr7.jpg
{kind=link}
7.4.8 Argonaute2 Cleaves the Anti-Guide Strand of siRNA during RISC Activation
Rand TA, Petersen S, Du F, Wang X.
Cell. 2005 Nov 18; 123(4):621-9
http://dx.doi.org/10.1016/j.cell.2005.10.020
The mRNA-cleavage step of RNA interference is mediated by an endonuclease, Argonaute2 (Ago2), within the RNA-induced silencing complex (RISC). Ago2 uses one strand of the small interfering (si) RNA duplex as a guide to find messenger RNAs containing complementary sequences and cleaves the phosphodiester backbone at a specific site measured from the guide strand’s 5′ end. Here, we show that both strands of siRNA get loaded onto Ago2 protein in Drosophila S2 cell extracts. The anti-guide strand behaves as a RISC substrate and is cleaved by Ago2. This cleavage event is important for the removal of the anti-guide strand from Ago2 protein and activation of RISC.
RNA interference (RNAi) is a gene-silencing process during which endogenous messenger RNA is destroyed upon introduction of the corresponding double-stranded RNA (Fire et al., 1998). RNAi has found widespread application as a technique within research laboratories allowing simple yet effective knockdown of genes of interest (reviewed by Dykxhoorn et al., 2003 and Meister and Tuschl, 2004). Biologically, RNAi-related processes are critical for development (Brennecke et al., 2003, Giraldez et al., 2005, Lee et al., 1993, Reinhart et al., 2000 and Rhoades et al., 2002), heterochromatin formation (Motamedi et al., 2004, Verdel et al., 2004 and Volpe et al., 2002), and offer cellular protection from virus (Baulcombe, 2004 and Vaucheret et al., 2001) and transposon proliferation (Sijen and Plasterk, 2003).
Recently, Ago2’s biochemical role within the RISC complex was clarified when its PIWI domain was identified as RNase H-like by crystallography (Song et al., 2004) and endonuclease V-like by a bioinformatics tool, metaBASIC analysis (Rand et al., 2004). Importantly, Ago2 was identified as the sole protein required for mRNA cleaving RISC activity (Liu et al., 2004, Meister et al., 2004 and Rand et al., 2004). The endonucleolytic RISC activity has also been demonstrated using bacterially expressed human Ago2 loaded with a single-stranded siRNA (Rivas et al., 2005).
Three previous observations are consistent with the existence of a helicase during RNA interference. First, siRNA-triggered RISC activity was reduced upon removal of ATP in a Drosophila embryo extract ( Nykanen et al., 2001). Helicases utilize ATP to separate the RNA strands. Secondly, though both double-stranded (ds) and single-stranded (ss) siRNA (at higher concentrations) can activate RISC in cell extracts, only ss siRNA can be directly loaded onto recombinant Ago2 ( Nykanen et al., 2001, Martinez et al., 2002 and Rivas et al., 2005). Finally, though their biochemical roles have not been demonstrated, several helicases have been identified by genetic or biochemical studies as potentially playing a part in RNAi in various systems ( Meister and Tuschl, 2004).
To illustrate the molecular mechanism of siRNA loading onto RISC and its subsequent activation process, we conducted a detailed biochemical analysis of this process. Our results are surprising and as reported here, suggest an alternative model for siRNA loading and RISC activation.
Loading and Activation of RISC Is ATP-Independent in S2 Cell Extracts
In an effort to further understand the siRNA loading mechanism of RISC, we depleted ATP from the S100 extracts prepared from Drosophila S2 cells by treating the extracts with Hexokinase in the presence of 100 mM glucose. The removal of ATP was followed using Sigma’s luciferase-based ATP assay. After treatment, the ATP level remaining, 0.8 pM, was at the detection limit of the kit (∼2 pM) and orders of magnitude less than the mock-treated extracts (data not shown). The ATP-depleted extracts and mock-treated extracts (addition of glucose and buffer in place of Hexokinase supplemented with 1 mM ATP) were then used to measure mRNA-cleaving RISC activity and double-stranded RNA-cleaving Dicer activity. As shown in Figure 1, when compared to mock-treated extracts (+ATP lanes), the ATP-depleted extracts (−ATP lanes) demonstrated comparable siRNA-induced RISC activity (upper panel). As a control for ATP removal, we checked the Dicer activity in the same samples (lower panel). The main Dicer activity in S2 100,000 × g supernatant (S100) is Dicer-2, which requires ATP to process dsRNA into siRNA ( Liu et al., 2003). The Hexokinase-treated extracts (−ATP lanes, Figure 1, bottom panel) were deficient for Dicer activity relative to the mock-treated sample (+ATP lanes, Figure 1, bottom panel), which demonstrated strong Dicer activity. This result, together with the luciferase-based quantification of ATP, demonstrated that siRNA-activated RISC activity is largely ATP independent in S2 extract. This result does not support the requirement of an ATP-dependent helicase in removing the anti-guide strand during RISC activation.
Anti-Guide Strand of siRNA Is Cleaved by Ago2 at the Ninth Position from the 5’ Phosphate
In light of the intrinsic endonuclease activity of Ago2, we entertained the possibility that the anti-guide strand of siRNA was removed from RISC through cleavage by Ago2 itself via the same ATP-independent cleavage mechanism used for mRNA substrates. The hypothesized cleavage of the anti-guide RNA and the subsequent release of the cleaved fragment from RISC would result in the production of two fragments from the original 21 nucleotide anti-guide siRNA strand. The 5′ fragment, containing the 5′ end phosphate, would be 9 bases long. The 3′ fragment would consist of the remaining 12 bases. In an attempt to detect the 5′ cleavage product, one strand of the let-7 siRNA was radio-phosphorylated with P32 and the other strand was phosphorylated with nonradioactive phosphate (Figure 2A). After loading onto RISC, the RNA was recovered and analyzed by urea/polyacrylamide gel electrophoresis followed by phosphorimaging. … we added 100-fold excess (relative to the siRNA) of a nuclease-resistant, 2′-O-methylated, single-stranded RNA oligonucleotide that could base pair with the radiolabeled product once it was released from its complementary siRNA strand (as shown in cartoon form in Figure 2A). Using this trapping technique, we easily detected a radiolabeled RNA ( Figure 2A, lanes 1–4) that perfectly comigrates with a radiolabeled synthetic RNA 9-mer of the same sequence ( Figure 2A, lanes 5 and 10). A 2′-O-methylated RNA trap consisting of the complementary sequence (UGA…) did not rescue the 9-mer, demonstrating that the trap was working by sequence-specific annealing, rather than, for instance, clogging an RNA degradation pathway ( Figure 2B, lane 12).
The cleaved anti-guide strand 9-mer was observed at comparable levels with or without ATP (Figure 2E). This is in agreement with the observation that ATP has little effect on the total process of siRNA-triggered RISC loading and activity as shown in Figure 1.
2’-O-Methyl Modification Decreases 9-Mer Production and RISC Activity
The effect of 2′-O-methyl inhibition is due to the ability of the methyl group to cause steric hindrance of the Mg2+-based cleavage mechanism ( Schwarz et al., 2004). We incorporated this chemical modification on base 9 of the anti-guide RNA. To ensure that the chemically modified RNA would in fact become the anti-guide, rather than the guide RNA, we used an asymmetrical RNA with a bubble A-G mismatch on one end as previously described ( Tomari et al., 2004b; see Figure 3A for the sequence). No RISC activity against the anti-guide strand’s target was detected ( Figure 3C, lanes 1–9), whereas the RISC activity generated by the guiding strand (green strand) was readily detected ( Figure 3B, lanes 1–9). As shown in Figure 3B, the 2′-O-methyl modification at position 9 of the anti-guide strand reduced the RISC activity of the guide strand by over 50% ( Figure 3B, lanes 1–3 compared to lanes 4–6). When the modification was moved to the neighboring position, base 8 from the 5′ end of the anti-guide strand, activity returned to the level of unmodified siRNA ( Figure 3B, lanes 1–3 and 7–9). Similarly, the modification at the 10 position also had full activity (data not shown). This indicates the sensitivity to chemical modification was site specific and the effect was limited only to the predicted Ago2 cleavage site.
As demonstrated in Figures 3B and 3C, the green-labeled strand is exclusively selected as the guide strand of RISC. We checked to see if 9-mer fragments were produced from the guide as well as the anti-guide strand. As shown in Figure 3E, 9-mer was only generated from the anti-guide (lanes 1–4), while the guide produced no detectable 9-mer (lanes 5–8).
Ago2 Interacts with the Anti-Guide Strand of siRNA Transiently
Because 2′-O-methyl 9 modification of the anti-guide strand of siRNA reduces RISC activation, we attempted to detect an increased interaction between the endogenous nuclease-active Ago2 with modified siRNA. Using the same modification strategy, we synthesized biotinylated unmodified, 9-modified, and 8-modified anti-guide strands. The biotin modification of 3′ hydroxyl group of either the guide or anti-guide strand of siRNA does not affect RISC activity ( Martinez et al., 2002 and data not shown). Each of these anti-guide strands was annealed to a nonbiotinylated guide strand. Likewise, we synthesized a biotinylated guide strand, which was annealed to nonbiotinylated unmodified, 9-modified, or 8-modified anti-guide RNAs. Each of the six siRNAs was then added into two types of S2 cell extracts. The first type of extracts contained HisFlagAgo2 as well as the endogenous Ago2 protein. The second type of extracts contained exogenously expressed HisFlag-tagged Ago2 containing a D965A active site mutation (again, in addition to the endogenous Ago2). The D965 position of Drosophila Ago2 is part of a well-conserved GADV motif previously demonstrated to be required for nuclease activity in human Ago2 ( Liu et al., 2004).
The above experimental results support a model for how siRNA, generated from long double-stranded RNA by Dicer enzyme, becomes the single-stranded guide of Ago2 nuclease. It seems that both strands of siRNA are loaded onto Ago2 and the anti-guide strand becomes the first substrate of Ago2. The cleavage and subsequent disassociation of the anti-guide strand fragments produces active RISC capable of targeting cognate mRNA for endonucleolytic cleavage.
Both Strands of siRNA Interact with Ago2
The ATP independence of RISC activation by siRNA suggested that both strands of siRNA might get passed to Ago2 from D2/R2/siRNA complex without being separated by a helicase. Indeed, using a 3’ biotin-modified siRNA, we detected interaction between both strands of siRNA and Ago2 protein (Figure 5). As predicted, the interaction between the guide strand and Ago2 was readily detected. However, the anti-guide strand and Ago2 interaction was only detected when we used the 2’-O-methyl-modified siRNA at the ninth position or when an active-site mutant Ago2 protein was used. Since in both cases, the cleavage of anti-guide strand is impaired, it seems to us that the cleavage event is important for the anti-guide strand to leave Ago2 and therefore activate RISC to cleave mRNA substrates.
Ago2 Cleaves the Anti-Guide Strand of siRNA
If both strands of siRNA get loaded onto Ago2 protein without separation, it is logical that the anti-guide strand will be cleaved by Ago2 endonuclease since it could closely mimic the mRNA substrate, differing only in length. However, direct detection of cleaved products from the anti-guide strand was not possible because of its instability and was only observed when a chemically modified single-stranded RNA“trap”that annealed to and protected it from degradation was used. The cleavage is ATP independent and occurs at the 9-10 bond counting from the 5’ phosphate of the anti-guide strand (or alternatively, if counting from the guide strand’s 5’ end, across from the guide strand’s 10-11 bond). The cleaved product contains a 5’ phosphate and a 3’ hydroxyl group since it can be ligated by T4 RNA ligase. Furthermore, the cleavage fragment was not produced in Ago2 null fly extracts. These features are consistent with the notion that the cleavage is executed by Ago2 endonuclease.
Is Anti-Guide Strand Cleavage Only for the Nuclease Competent Argonaut Proteins?
Evidence is mounting that many of these proteins use guide RNAs to perform their cellular functions. Do these proteins undergo a similar loading process including cleavage of the anti-guide strand? It is thought that the small, noncoding, imperfectly base-paired hairpin microRNAs are processed by Dicer to form siRNA-like species. Such a species would differ from siRNA in that there would be bubbles where mismatches occur. Mismatches between the guide and target RNA have been shown to decrease RISC cleavage activity against mRNA targets when the bubble occurs close to the cleavage site (Martinez and Tuschl, 2004). This argues that certain microRNAs will not be cleaved by their respective Ago proteins during the loading process. Of course, the mismatch bubbles might serve the same function as cleavage by helping to decrease the overall melting temperature of the duplex. Nevertheless, family members such as human Ago1, 3, and 4 lacking the nuclease activity will be unable to cleave the anti-guide RNA no matter what level of complementarity the two strands have.
Leave a Reply