An Important Marker of Hypertension in Young Adults: Plasma Renin
Author: Manuela Stoicescu, MD, PhD
Original research
Manuela Stoicescu, MD, PhD
Consultant Internal Medicine, Assistant Professor
Faculty of Medicine and Pharmacy, Medical Disciplines Department
University of Oradea, Romania
ABSTRACT
Introduction: Plasma renin level is an important marker of hypertension in the young adults. The purpose of this study was to determine the role of increased levels of plasmatic renin in the pathogenesis of hypertension in the young adults and to highlight the main conditions underlying the pathogenesis of hypertension in the young people in these circumstances.
Material and methods: The group of patients taking part in the study was of 121 young hypertensive adults (selected from a group of 321 young hypertensive adults), with the age between 18-35 years, with elevated blood pressure exceeding 140/90mmHg in at least three repeated measurements at intervals of one week to exclude white coat phenomenon, or had a blood pressure value greater than 170/100mmHg at the first measurement and increased plasma renin levels above the 4,3ng/ml.
Results and discussion: Of the 121 young hypertensive patients with increased plasma renin levels, 49 were cases of renal artery stenosis representing 40.50% (p<0.001), 8 cases were represented by small unilateral kidneys representing 6.61% (p<0.001), renal cell carcinoma (previously known as “hypernephron” – Grawitz tumor) was responsible for the younger group of patients studied of 4 cases representing 3.30% (p <0.001) of the cases of hypertension in the young adults, and 60 cases representing 49.59% were represented by pheochromocytoma.
Conclusions: The results show the role of plasma renin dosing as being particularly important in the pathogenesis of secondary hypertension in the young adults
Keywords: Renin, arterial hypertension (HBP), young adults
INTRODUCTION
Renin is an enzyme secreted by the juxtaglomerular apparatus to maintain electrolyte balance and blood pressure in the appropriate limits. Plasma renin level is an important marker of hypertension in the young adults. The purpose of this study was to determine the role of increased levels of plasma renin in the pathogenesis of hypertension in the young adults and highlight the main conditions underlying the pathogenesis of hypertension in the young adults with increased plasmatic renin. The principal diseases which had increased plasma renin levels were: renal artery stenosis, pheochromocytoma, congenital unilateral small kidney, primary reninoma (renal cell carcinoma or Grawitz tumor), situations in which renin is secreted in excess, the highest values being in cases of renal cell carcinoma, of 320ng/ml.
MATERIAL AND METHODS
The group of patients taking parts in the study was of 121 hypertensive young adults, with the ages between 18-35 years with elevated blood pressure over 140/90mmHg in at least three repeated measurements at intervals of one week to exclude white coat phenomenon frequently encountered in the young, or had a severely increased blood pressure of >170/100mmHg on the first measurement and plasma renin levels greater than 4,3 ng/ml.
Parameters for assessment of the diseases which had increased plasma rennin levels were clinical, radiological, biological and histopathological. The study was done after the diagnosis of hypertension and staging according OMS. All patients were investigated clinically and fully analyzed paraclinically. They agreed to participate in the trial after they were explained the criteria of professional ethics, scientific and terms of confidentiality. All patients participating in the study had plasmatic renin levels above 4.3ng /ml. The statistical analysis was done with the help of EPIINFO application, version 6.0, program of the Center for Disease Control and Prevention-CDC in Atlanta, suitable for processing of medical statistics. Averages were calculated for the parameters, frequency ranges, standard deviations, tests of statistical significance by Student method (t test) and χ ².
RESULTS
The group of young hypertensive patients with ages 18-35 years, with elevated blood pressure >140/90mmHg, with increased plasma renin levels over 4.3ng/ml we found 49 cases of renal artery stenosis representing 40.50%, 8 cases of congenital small kidney representing 6.61%, 4 cases of Grawitz tumors (renal cell carcinoma) representing 3.30% and 60 cases of pheochromocytoma representing 49.59%. Table No.1
Table 1. The main conditions that were present in the group of young hypertensive patients with increased plasma renin level.
|
Diseases |
No. of cases |
Percentage of cases [%] |
|
Vascular pathology |
49 |
40.50% |
|
Renal parenchymatous pathology Congenital small kidney |
8 |
6.61% |
|
Renal carcinoma (Gravitz tumor) |
4 |
3.30% |
|
60 |
49.59% |
The positive criteria’s of diagnostic for the diseases were included in the study was:
I. Renal artery stenosis
- The increased value of diastolic blood pressure over 110mmHg.
- Paraombilical systolic murmur.
- Imaging of arteriography.
- Increased plasmatic renin level > 4.3 ng / ml.
II. Congenital small kidney
- Values of blood pressure over 140/90mmHg.
- Arteriography – put in evidence the small kidney
- The abdominal MRI
- Increased plasmatic renin level > 4.3 ng / ml.
III. Renal carcinoma (Gravitz tumor)
- Unilateral lumbar pain
- Loss of appetite
- Weight loss
- Macroscopic hematuria (blood in the urine)
- The value of plasma rennin level increased > 4.3ng /ml, mentioning that in this situation the plasma renin values reached the highest values up to 320ng/ml
- The abdominal MRI
- The renal biopsy
IV. Pheochromocytoma
- The paroxysmal outbursts of severe blood pressure over 220/120mmHg
- Headache
- Tremor of extremities
- Nervousness
- Increased serum catecholamine levels above 260pg/ml
- Increased urinary catecholamine values above the 7.4 mg/24 hrs
- Increased plasmatic renin level > 4.3 ng/ml.
- The abdominal MRI used in the detection of adrenal tumors .
Table 2. Diagnostic criteria’s met by patients
|
Diseases |
No. of cases |
Diagnostic criteria’s met by patients |
|
Vascular pathology Renal artery stenosis |
49 |
24 cases 4 of 4 25 cases 3 of 4 |
|
Renal parenchymatous pathology Congenital small kidney |
8 |
8 cases 4 of 4
|
|
Renal carcinoma (Gravitz tumor) |
4 |
2 cases 7 of 7 2 cases 6 of 7 |
|
Pheochromocytoma |
60 |
38 cases 8 of 8 12 cases 7 of 8 10 cases 6 of 8 |
Of the group of young hypertensive patients studied with increased plasma renin activity, 49 of the cases were renal artery stenosis representing 40.50% (p <0.001). The parameters of the clinical assessment were the increased value of diastolic blood pressure over 110mmHg, paraombilical systolic murmur and an imaging of arteriography. Figure 1.
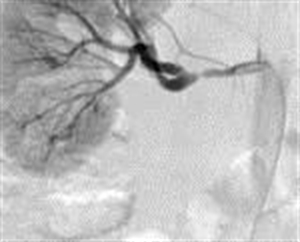
Figure 1. Arteriography of the right renal artery stenosis (M.I. aged 21 years with HBP = 170/120mmHg)
Of the group of young patients participating in the study, we found 8 cases of unilateral small kidney representing 6.61% (p<0.001). The pathogenic mechanism of hypertension was ischemic, in that all cases arterial high blood pressure evolved along with hyperreninemia in congenital unilateral small kidney. The early diagnosis of renal disease is very important, in the best cases before the hypertension causes severe nephroangiosclerosis on the contralateral kidney, leading to nephrectomy which then can then lead to the disappearance of hypertension. The parameters of assessment in this case were the clinical blood pressure values above 140/90mmHg, imaging methods to put in evidence the small kidney: arteriography Figure2 abdominal MRI Figura3 and biological-increased plasmatic renin activity> 4.3 ng / ml.
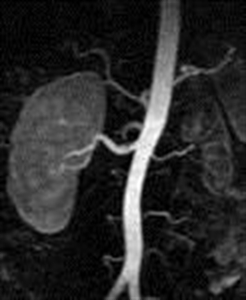
Figure2. Arteriography evidence the congenital small left kidney (D.R. of 19 years old with HBP = 165/110mmHg)
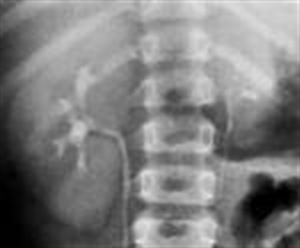
Figure 3. MRI – scan with contrast substance putting in evidence the left renal hypoplasia (D.R. of 19 years old with HBP = 165/110mmHg)
Renal cell carcinoma (renal carcinoma, previously “hypernephroma” – Grawitz tumor) was responsible for the younger group of patients studied, 4 cases representing 3,30% (p<0.001) of the HBP young cases. All had severely elevated blood pressure values over 200/100mmHg. The diagnostic was based on clinical parameters: unilateral lumbar pain, loss of appetite, weight loss, but only two cases had macroscopic hematuria (blood in the urine), biological – the value of increased plasma rennin level > 4.3ng /ml, mentioning that in this situation the plasma renin values reached the highest values up to 320ng/ml. Imaging parameters are represented in the abdominal MRI by Figure 4.
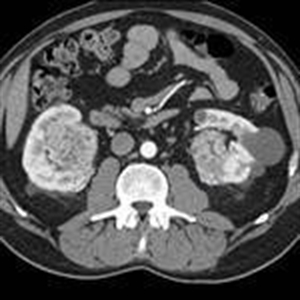
Figure 4. MRI-scan with bilateral renal tumor (F.R.of 34 years with malignant HBP=220/130mmHg — worked with pesticides)
Histopathological parameters were put into evidence in all four cases in which renal biopsy was performed and the histopathological results of which are outlined below:
Two cases were clear cell renal carcinoma based on the histopathology results after renal biopsy – histological preparation with H&E staining with the objective of 10X is shown in (Figure 5a) and (Figure 5b)
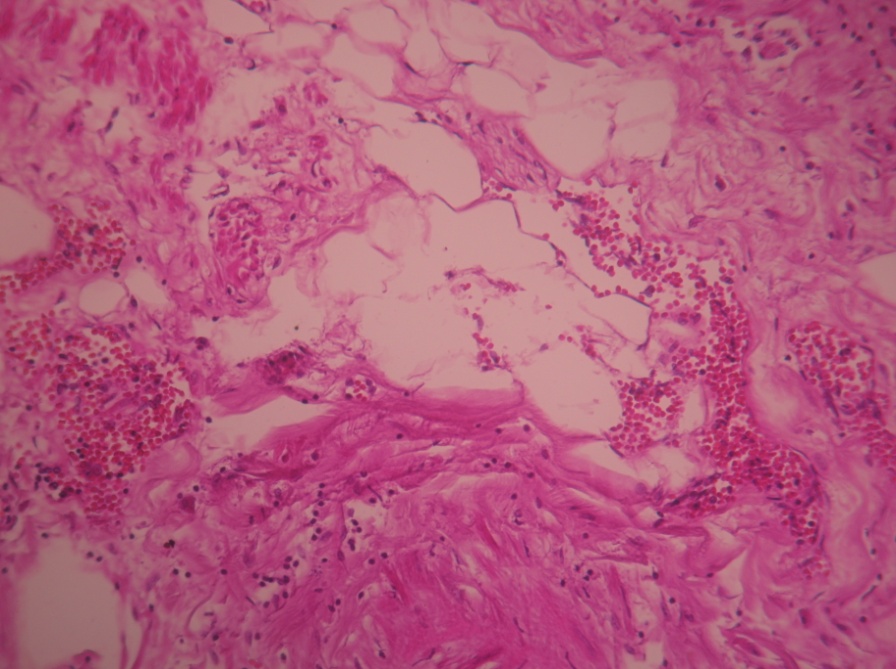
Figure 5a. Clear cell renal carcinoma (objective 10x) – H&E stain (M.I. 21 years with paroxysmal HBP=200/110mmHg)
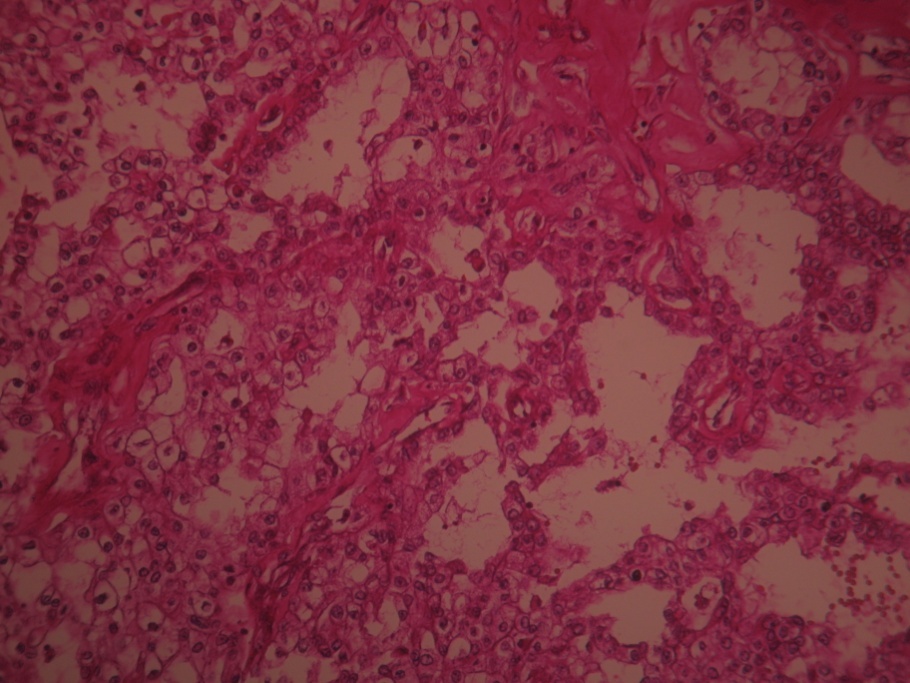
Figure 5b. Clear cell renal carcinoma (objective 10x) – H&E stain (P.R. 28 years with severe form HBP=210/110mmhHg)
The other two cases of renal carcinoma are represented in the following H&E stained images, after the renal biopsy (Figure 6) and (Figure 7).
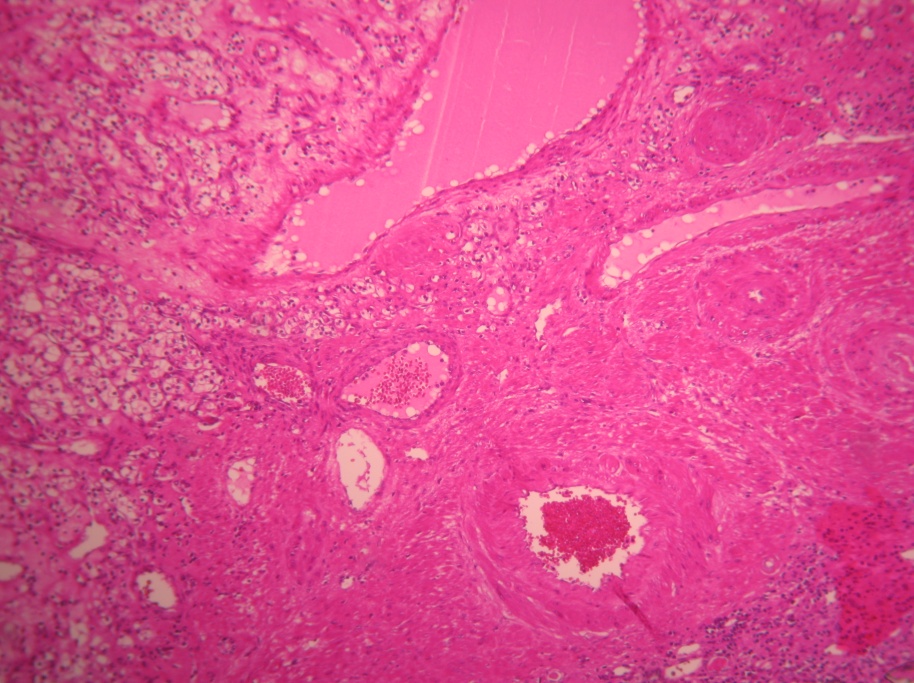
Figure 6. Renal carcinoma (objective 10x). H & E stain (I.G. 31 years with paroxysmal HBP=210/115 mmHg)
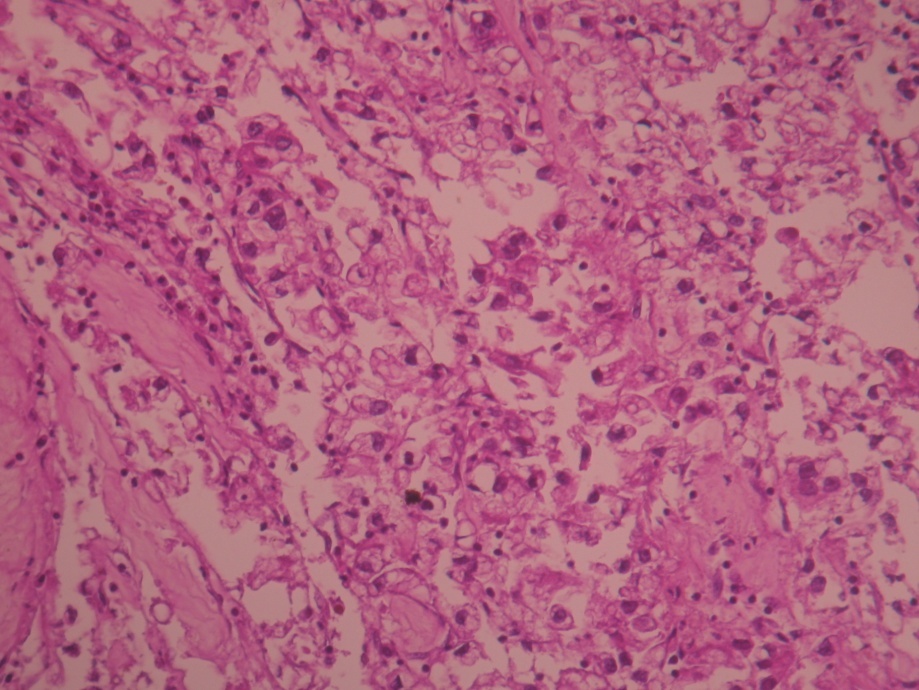
Figure 7. Renal carcinoma (objective 10x). H & E stain (F.R. 34 years with malignant HBP=220/130mmHg — worked with pesticides)
Hypertension in the young patients with renal cell carcinoma took the form of severe paroxysmal HBP=200/110mmHg or above in all four cases, due to excessive secretion of renin produced in large quantities by the tumor and it was the one form which attracted most the clinical attention when it was not manifested by macroscopic hematuria.
Of the group of hypertensive young adults studied, 60 of the cases were of pheochromocytoma representing 49.59%.
The diagnostic criteria used in this clinical situation were: paroxysmal outbursts of severe blood pressure values over 220/120mmHg accompanied by headache, tremor of extremities, nervousness, biological parameters represented by increased serum catecholamine levels above 260pg/ml, increased urinary catecholamine values above the 7.4 mg/24 hrs, imaging parameters which were used in the detection of adrenal tumors by performing an abdominal MRI. Figure 8.
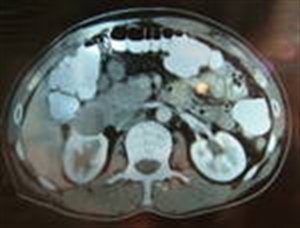
Figure 8. Abdominal MRI-scan with pheochromocytoma of the right adrenal gland (G.R. 24 years with paroxysmal HBP=220/130mmHg)
DISCUSSIONS
The importance of this study was to measure the level of plasma renin of hypertensive young patients with ages between 18-35 years to determine its role in the pathogenesis of secondary hypertension in the young adults. Also the conditions in which plasma renin level is increased in the context of secondary hypertension in the young patients.
Renovascular hypertension was one of the important causes of secondary hypertension in the young, its frequency in the group of patients studied was of 49 cases with renal artery stenosis representing 40.50% (p <0.001), in all these cases the renin plasma level was increased above 4,3ng/ml.
Safian R.D. and Textor S.C. [1] found the frequency of renal artery stenosis in a group of young hypertensive patients with increased plasma renin activity, as being 42.36%, which is slightly higher than in our study, this difference could be explained by a better paraclinical investigation of the young patients with hypertension.
Of the group of young hypertensive patients participating in the study we found 8 cases of unilateral congenital small kidney, representing 6.61% (p<0.001).
Goddard C, et al. [2] found that the incidence of hypertension in young people with kidney hypoplasia was 25%. They suggested that the renin-angiotensin-aldosterone system plays an important role in the pathogenesis of hypertension in the situation of renal hypoplasia. This difference could be explained by the fact that young patients in other countries had an increased teratogenic risk compared with the young in our country.
Renal cell carcinoma (Grawitz tumor) was responsible for the younger group of patients studied, 4 cases representing 3.30% (p<0.001) of all the young hypertensive patients. Two cases were clear cell renal cell carcinoma histopathology analyzed after a renal biopsy. The data obtained are slightly lower than those in the literature (5%) Sukarochana [3] and (4%) Gangurly [4]. This difference could be explained by the fact that our country carcinogenic risk factors are lower.
Rose HJ, Pruitt AW [5] reported the case of a young patient with severe hypertension of 190/110mmHg, which after further investigations had found increased plasma renin levels and after paraclinical investigations a solitary simple kidney cyst was found.
DW Robertson et al. [6] reported the case of a young man who had elevated blood pressure (HBP=180/120mmHg) and after investigations increased plasma renin level was found and a left renal tumor (primary reninoma) was found, whose blood pressure values were normalized after tumor resection.
Pheochromocytoma was found in 60 of the cases representing 49.59% of cases of the young hypertensive adults.
Abrams HL [7] found that the incidence of pheochromocytoma in the young hypertensive cases was 21.03%, Bravo EL [8] found 42.38% cases of pheochromocytoma, and Bravo EL, Gifford RWJr [9] 46.03 % of young hypertensive patients with pheochromocytoma. These results are lower than those obtained in our study. This could be explained by the risk factors in this geographical area and dominant genetic factor has an important role in the etiology of pheochromocytoma.
CONCLUSIONS
- Plasmatic renin level is an important marker of hypertension in the young adults.
- The highest plasmatic renin levels up to 320ng/ml were found in the cases of renal cell carcinoma, because the kidney tumor cells secrete increased amounts of renin.
- This situation suggests that hypertension in the young adults is hyperreninemia hypertension in the most cases dominated by a vasoconstriction and increased peripheral vascular resistance due hyperactivity of the sympathetic nervous system, being a rapidly evolving form of hypertension with vascular complications.
- The results of plasma renin dosing shows its important role in the pathogenesis in secondary hypertension of the young adults, these conditions are not quite as rare as one might think but not enough investigated.
- This marker should be routinely performed in young patients with hypertension, especially those with medium and severe forms of blood pressure > 170/100mmHg, having a role in establishing the etiology of hypertension in the young, however presently it is still not made often enough, but situation must to be change in the future.
REFERENCES:
[1] Safian R.D, Textor S.C, Renal-artery stenosis, N Engl J Med 2001, 344(6):431-42.
[2] Goddard C , Vallothon MB, Broyer M, Plasma rennin activity in segmental hypoplasia of kidneys with hypertension, Nephron 2003, 11:308-17.
[3] Sukarochana K, Nephroblastoma and hypertension J Surg 2005, 7- 573.
[5] Rose HJ, Pruitt AW, Hypertension, hyperreninemia and a solitary renal cyst in an adolescent, Am J Med 2004, 61; 579-82.
[6] Robertson DW, Klidjiana A, Harding KK, Walters G, Lee MR, Robb-Smith AHT, Hypertension due to a renin-secreting renal tumor, Am Med 2005, 43 (9) 63-76.
[8] Bravo EL, Pheochromocytoma: New concepts and future trends, Kidney Int 1991, 40:544-556.
[9] Bravo EL, Gifford RWJr, Pheochromocytoma: Diagnosis, localization and management, N Engl J. Med 1984, 311-1298.
Corresponding author
Manuela Stoicescu, Internal Medicine Department, University of Oradea, Faculty of Medicine and Pharmacy, Oradea, Romania: County Hospital of Oradea, Phone 0723019951, e-mail: manuela_stoicescu@yahoo.com
PUT IT IN CONTEXT OF CANCER CELL MOVEMENT
The contraction of skeletal muscle is triggered by nerve impulses, which stimulate the release of Ca2+ from the sarcoplasmic reticuluma specialized network of internal membranes, similar to the endoplasmic reticulum, that stores high concentrations of Ca2+ ions. The release of Ca2+ from the sarcoplasmic reticulum increases the concentration of Ca2+ in the cytosol from approximately 10-7 to 10-5 M. The increased Ca2+ concentration signals muscle contraction via the action of two accessory proteins bound to the actin filaments: tropomyosin and troponin (Figure 11.25). Tropomyosin is a fibrous protein that binds lengthwise along the groove of actin filaments. In striated muscle, each tropomyosin molecule is bound to troponin, which is a complex of three polypeptides: troponin C (Ca2+-binding), troponin I (inhibitory), and troponin T (tropomyosin-binding). When the concentration of Ca2+ is low, the complex of the troponins with tropomyosin blocks the interaction of actin and myosin, so the muscle does not contract. At high concentrations, Ca2+ binding to troponin C shifts the position of the complex, relieving this inhibition and allowing contraction to proceed.
Figure 11.25
Association of tropomyosin and troponins with actin filaments. (A) Tropomyosin binds lengthwise along actin filaments and, in striated muscle, is associated with a complex of three troponins: troponin I (TnI), troponin C (TnC), and troponin T (TnT). In (more ) Contractile Assemblies of Actin and Myosin in Nonmuscle Cells
Contractile assemblies of actin and myosin, resembling small-scale versions of muscle fibers, are present also in nonmuscle cells. As in muscle, the actin filaments in these contractile assemblies are interdigitated with bipolar filaments of myosin II, consisting of 15 to 20 myosin II molecules, which produce contraction by sliding the actin filaments relative to one another (Figure 11.26). The actin filaments in contractile bundles in nonmuscle cells are also associated with tropomyosin, which facilitates their interaction with myosin II, probably by competing with filamin for binding sites on actin.
Figure 11.26
Contractile assemblies in nonmuscle cells. Bipolar filaments of myosin II produce contraction by sliding actin filaments in opposite directions. Two examples of contractile assemblies in nonmuscle cells, stress fibers and adhesion belts, were discussed earlier with respect to attachment of the actin cytoskeleton to regions of cell-substrate and cell-cell contacts (see Figures 11.13 and 11.14). The contraction of stress fibers produces tension across the cell, allowing the cell to pull on a substrate (e.g., the extracellular matrix) to which it is anchored. The contraction of adhesion belts alters the shape of epithelial cell sheets: a process that is particularly important during embryonic development, when sheets of epithelial cells fold into structures such as tubes.
The most dramatic example of actin-myosin contraction in nonmuscle cells, however, is provided by cytokinesisthe division of a cell into two following mitosis (Figure 11.27). Toward the end of mitosis in animal cells, a contractile ring consisting of actin filaments and myosin II assembles just underneath the plasma membrane. Its contraction pulls the plasma membrane progressively inward, constricting the center of the cell and pinching it in two. Interestingly, the thickness of the contractile ring remains constant as it contracts, implying that actin filaments disassemble as contraction proceeds. The ring then disperses completely following cell division.
Figure 11.27
Cytokinesis. Following completion of mitosis (nuclear division), a contractile ring consisting of actin filaments and myosin II divides the cell in two.
http://www.ncbi.nlm.nih.gov/books/NBK9961/
This is good. I don’t recall seeing it in the original comment. I am very aware of the actin myosin troponin connection in heart and in skeletal muscle, and I did know about the nonmuscle work. I won’t deal with it now, and I have been working with Aviral now online for 2 hours.
I have had a considerable background from way back in atomic orbital theory, physical chemistry, organic chemistry, and the equilibrium necessary for cations and anions. Despite the calcium role in contraction, I would not discount hypomagnesemia in having a disease role because of the intracellular-extracellular connection. The description you pasted reminds me also of a lecture given a few years ago by the Nobel Laureate that year on the mechanism of cell division.
PUT IT IN CONTEXT OF CANCER CELL MOVEMENT
The contraction of skeletal muscle is triggered by nerve impulses, which stimulate the release of Ca2+ from the sarcoplasmic reticuluma specialized network of internal membranes, similar to the endoplasmic reticulum, that stores high concentrations of Ca2+ ions. The release of Ca2+ from the sarcoplasmic reticulum increases the concentration of Ca2+ in the cytosol from approximately 10-7 to 10-5 M. The increased Ca2+ concentration signals muscle contraction via the action of two accessory proteins bound to the actin filaments: tropomyosin and troponin (Figure 11.25). Tropomyosin is a fibrous protein that binds lengthwise along the groove of actin filaments. In striated muscle, each tropomyosin molecule is bound to troponin, which is a complex of three polypeptides: troponin C (Ca2+-binding), troponin I (inhibitory), and troponin T (tropomyosin-binding). When the concentration of Ca2+ is low, the complex of the troponins with tropomyosin blocks the interaction of actin and myosin, so the muscle does not contract. At high concentrations, Ca2+ binding to troponin C shifts the position of the complex, relieving this inhibition and allowing contraction to proceed.
Figure 11.25
Association of tropomyosin and troponins with actin filaments. (A) Tropomyosin binds lengthwise along actin filaments and, in striated muscle, is associated with a complex of three troponins: troponin I (TnI), troponin C (TnC), and troponin T (TnT). In (more ) Contractile Assemblies of Actin and Myosin in Nonmuscle Cells
Contractile assemblies of actin and myosin, resembling small-scale versions of muscle fibers, are present also in nonmuscle cells. As in muscle, the actin filaments in these contractile assemblies are interdigitated with bipolar filaments of myosin II, consisting of 15 to 20 myosin II molecules, which produce contraction by sliding the actin filaments relative to one another (Figure 11.26). The actin filaments in contractile bundles in nonmuscle cells are also associated with tropomyosin, which facilitates their interaction with myosin II, probably by competing with filamin for binding sites on actin.
Figure 11.26
Contractile assemblies in nonmuscle cells. Bipolar filaments of myosin II produce contraction by sliding actin filaments in opposite directions. Two examples of contractile assemblies in nonmuscle cells, stress fibers and adhesion belts, were discussed earlier with respect to attachment of the actin cytoskeleton to regions of cell-substrate and cell-cell contacts (see Figures 11.13 and 11.14). The contraction of stress fibers produces tension across the cell, allowing the cell to pull on a substrate (e.g., the extracellular matrix) to which it is anchored. The contraction of adhesion belts alters the shape of epithelial cell sheets: a process that is particularly important during embryonic development, when sheets of epithelial cells fold into structures such as tubes.
The most dramatic example of actin-myosin contraction in nonmuscle cells, however, is provided by cytokinesisthe division of a cell into two following mitosis (Figure 11.27). Toward the end of mitosis in animal cells, a contractile ring consisting of actin filaments and myosin II assembles just underneath the plasma membrane. Its contraction pulls the plasma membrane progressively inward, constricting the center of the cell and pinching it in two. Interestingly, the thickness of the contractile ring remains constant as it contracts, implying that actin filaments disassemble as contraction proceeds. The ring then disperses completely following cell division.
Figure 11.27
Cytokinesis. Following completion of mitosis (nuclear division), a contractile ring consisting of actin filaments and myosin II divides the cell in two.
http://www.ncbi.nlm.nih.gov/books/NBK9961/
This is good. I don’t recall seeing it in the original comment. I am very aware of the actin myosin troponin connection in heart and in skeletal muscle, and I did know about the nonmuscle work. I won’t deal with it now, and I have been working with Aviral now online for 2 hours.
I have had a considerable background from way back in atomic orbital theory, physical chemistry, organic chemistry, and the equilibrium necessary for cations and anions. Despite the calcium role in contraction, I would not discount hypomagnesemia in having a disease role because of the intracellular-extracellular connection. The description you pasted reminds me also of a lecture given a few years ago by the Nobel Laureate that year on the mechanism of cell division.
PUT IT IN CONTEXT OF CANCER CELL MOVEMENT
The contraction of skeletal muscle is triggered by nerve impulses, which stimulate the release of Ca2+ from the sarcoplasmic reticuluma specialized network of internal membranes, similar to the endoplasmic reticulum, that stores high concentrations of Ca2+ ions. The release of Ca2+ from the sarcoplasmic reticulum increases the concentration of Ca2+ in the cytosol from approximately 10-7 to 10-5 M. The increased Ca2+ concentration signals muscle contraction via the action of two accessory proteins bound to the actin filaments: tropomyosin and troponin (Figure 11.25). Tropomyosin is a fibrous protein that binds lengthwise along the groove of actin filaments. In striated muscle, each tropomyosin molecule is bound to troponin, which is a complex of three polypeptides: troponin C (Ca2+-binding), troponin I (inhibitory), and troponin T (tropomyosin-binding). When the concentration of Ca2+ is low, the complex of the troponins with tropomyosin blocks the interaction of actin and myosin, so the muscle does not contract. At high concentrations, Ca2+ binding to troponin C shifts the position of the complex, relieving this inhibition and allowing contraction to proceed.
Figure 11.25
Association of tropomyosin and troponins with actin filaments. (A) Tropomyosin binds lengthwise along actin filaments and, in striated muscle, is associated with a complex of three troponins: troponin I (TnI), troponin C (TnC), and troponin T (TnT). In (more ) Contractile Assemblies of Actin and Myosin in Nonmuscle Cells
Contractile assemblies of actin and myosin, resembling small-scale versions of muscle fibers, are present also in nonmuscle cells. As in muscle, the actin filaments in these contractile assemblies are interdigitated with bipolar filaments of myosin II, consisting of 15 to 20 myosin II molecules, which produce contraction by sliding the actin filaments relative to one another (Figure 11.26). The actin filaments in contractile bundles in nonmuscle cells are also associated with tropomyosin, which facilitates their interaction with myosin II, probably by competing with filamin for binding sites on actin.
Figure 11.26
Contractile assemblies in nonmuscle cells. Bipolar filaments of myosin II produce contraction by sliding actin filaments in opposite directions. Two examples of contractile assemblies in nonmuscle cells, stress fibers and adhesion belts, were discussed earlier with respect to attachment of the actin cytoskeleton to regions of cell-substrate and cell-cell contacts (see Figures 11.13 and 11.14). The contraction of stress fibers produces tension across the cell, allowing the cell to pull on a substrate (e.g., the extracellular matrix) to which it is anchored. The contraction of adhesion belts alters the shape of epithelial cell sheets: a process that is particularly important during embryonic development, when sheets of epithelial cells fold into structures such as tubes.
The most dramatic example of actin-myosin contraction in nonmuscle cells, however, is provided by cytokinesisthe division of a cell into two following mitosis (Figure 11.27). Toward the end of mitosis in animal cells, a contractile ring consisting of actin filaments and myosin II assembles just underneath the plasma membrane. Its contraction pulls the plasma membrane progressively inward, constricting the center of the cell and pinching it in two. Interestingly, the thickness of the contractile ring remains constant as it contracts, implying that actin filaments disassemble as contraction proceeds. The ring then disperses completely following cell division.
Figure 11.27
Cytokinesis. Following completion of mitosis (nuclear division), a contractile ring consisting of actin filaments and myosin II divides the cell in two.
http://www.ncbi.nlm.nih.gov/books/NBK9961/
This is good. I don’t recall seeing it in the original comment. I am very aware of the actin myosin troponin connection in heart and in skeletal muscle, and I did know about the nonmuscle work. I won’t deal with it now, and I have been working with Aviral now online for 2 hours.
I have had a considerable background from way back in atomic orbital theory, physical chemistry, organic chemistry, and the equilibrium necessary for cations and anions. Despite the calcium role in contraction, I would not discount hypomagnesemia in having a disease role because of the intracellular-extracellular connection. The description you pasted reminds me also of a lecture given a few years ago by the Nobel Laureate that year on the mechanism of cell division.
PUT IT IN CONTEXT OF CANCER CELL MOVEMENT
The contraction of skeletal muscle is triggered by nerve impulses, which stimulate the release of Ca2+ from the sarcoplasmic reticuluma specialized network of internal membranes, similar to the endoplasmic reticulum, that stores high concentrations of Ca2+ ions. The release of Ca2+ from the sarcoplasmic reticulum increases the concentration of Ca2+ in the cytosol from approximately 10-7 to 10-5 M. The increased Ca2+ concentration signals muscle contraction via the action of two accessory proteins bound to the actin filaments: tropomyosin and troponin (Figure 11.25). Tropomyosin is a fibrous protein that binds lengthwise along the groove of actin filaments. In striated muscle, each tropomyosin molecule is bound to troponin, which is a complex of three polypeptides: troponin C (Ca2+-binding), troponin I (inhibitory), and troponin T (tropomyosin-binding). When the concentration of Ca2+ is low, the complex of the troponins with tropomyosin blocks the interaction of actin and myosin, so the muscle does not contract. At high concentrations, Ca2+ binding to troponin C shifts the position of the complex, relieving this inhibition and allowing contraction to proceed.
Figure 11.25
Association of tropomyosin and troponins with actin filaments. (A) Tropomyosin binds lengthwise along actin filaments and, in striated muscle, is associated with a complex of three troponins: troponin I (TnI), troponin C (TnC), and troponin T (TnT). In (more ) Contractile Assemblies of Actin and Myosin in Nonmuscle Cells
Contractile assemblies of actin and myosin, resembling small-scale versions of muscle fibers, are present also in nonmuscle cells. As in muscle, the actin filaments in these contractile assemblies are interdigitated with bipolar filaments of myosin II, consisting of 15 to 20 myosin II molecules, which produce contraction by sliding the actin filaments relative to one another (Figure 11.26). The actin filaments in contractile bundles in nonmuscle cells are also associated with tropomyosin, which facilitates their interaction with myosin II, probably by competing with filamin for binding sites on actin.
Figure 11.26
Contractile assemblies in nonmuscle cells. Bipolar filaments of myosin II produce contraction by sliding actin filaments in opposite directions. Two examples of contractile assemblies in nonmuscle cells, stress fibers and adhesion belts, were discussed earlier with respect to attachment of the actin cytoskeleton to regions of cell-substrate and cell-cell contacts (see Figures 11.13 and 11.14). The contraction of stress fibers produces tension across the cell, allowing the cell to pull on a substrate (e.g., the extracellular matrix) to which it is anchored. The contraction of adhesion belts alters the shape of epithelial cell sheets: a process that is particularly important during embryonic development, when sheets of epithelial cells fold into structures such as tubes.
The most dramatic example of actin-myosin contraction in nonmuscle cells, however, is provided by cytokinesisthe division of a cell into two following mitosis (Figure 11.27). Toward the end of mitosis in animal cells, a contractile ring consisting of actin filaments and myosin II assembles just underneath the plasma membrane. Its contraction pulls the plasma membrane progressively inward, constricting the center of the cell and pinching it in two. Interestingly, the thickness of the contractile ring remains constant as it contracts, implying that actin filaments disassemble as contraction proceeds. The ring then disperses completely following cell division.
Figure 11.27
Cytokinesis. Following completion of mitosis (nuclear division), a contractile ring consisting of actin filaments and myosin II divides the cell in two.
http://www.ncbi.nlm.nih.gov/books/NBK9961/
This is good. I don’t recall seeing it in the original comment. I am very aware of the actin myosin troponin connection in heart and in skeletal muscle, and I did know about the nonmuscle work. I won’t deal with it now, and I have been working with Aviral now online for 2 hours.
I have had a considerable background from way back in atomic orbital theory, physical chemistry, organic chemistry, and the equilibrium necessary for cations and anions. Despite the calcium role in contraction, I would not discount hypomagnesemia in having a disease role because of the intracellular-extracellular connection. The description you pasted reminds me also of a lecture given a few years ago by the Nobel Laureate that year on the mechanism of cell division.
I actually consider this amazing blog , âSAME SCIENTIFIC IMPACT: Scientific Publishing –
Open Journals vs. Subscription-based « Pharmaceutical Intelligenceâ, very compelling plus the blog post ended up being a good read.
Many thanks,Annette