Cell-cell communication in brain
Larry H. Bernstein, MD, FCAP, Curator
LPBI
Adjustable Brain Cells
Neighboring neurons can manipulate astrocytes.
| February 18, 2016 http://www.the-scientist.com/?articles.view/articleNo/45376/title/Adjustable-Brain-Cells

http://www.the-scientist.com/images/News/February2016/astrocyte.jpg
{kind=link}
Neurons in the adult mouse brain can shape the features and physiologies of nearby astroglial cells, according to a study published today (February 18) in Science. Researchers at McGill University in Montreal and their colleagues have identified a molecular signal called sonic hedgehog (Shh), secreted by neurons, that acts as the agent of change.
“What’s very exciting about the paper is this notion that a cell’s fate might be determined—after it has already established its morphology and location in the brain—based on interactions with its neighbors,” said neurologist Ed Ruthazer of the Montreal Neurological Institute at McGill who was not involved in the research. “And the conversion is not superficial,” he added, “it really does seem to fundamentally reorganize the transcriptome of the cell.”
Astroglia are non-neuronal cells in the central nervous system that generally support and modulate neuronal function. The mammalian brain has an assortment of astrocytes, which perform a variety of specialized functions. This diversity was largely thought to be established during embryogenesis and early postnatal development, said Keith Murai of McGill who led the new research. “But after that,” he said, “the properties of these cells were thought to be solidified . . . for the rest of their lives.”
Murai and his colleagues had a different view, however. “Some of these [astrocytes] are so specialized around certain neural circuits that it was hard to imagine that all of the properties of these cells could be determined by that point [in development],” he said. After all, the neural circuitry itself isn’t fully formed until much later.
To investigate whether astrocyte identity might continue to be shaped beyond the perinatal period, Murai’s team searched for gene products in adult neurons and astrocytes that might govern continuing development. To simplify matters, the researchers focused on the cerebellar cortex, where just two types of astrocyte exist—Bergmann glial cells (BGs), which encapsulate the impulse-receiving regions of Purkinji cell neurons (PCs), and velate astrocytes (VAs), which surround granule cell neurons (GCs). Their searches revealed many candidate factors, said Murai, but one pathway kept coming up: Shh signaling.
Shh is a developmental morphogen known to have many important roles in the developing embryo, including the specification of cells in the brain, explained Murai. “People thought that the pathway was shut down and eliminated from the brain after it developed,” he said, “but as it turns out, this pathway is very potent even in the adult brain.”
The team found that the Shh protein itself was produced by PC neurons in the cerebellum, and that Shh receptors were abundantly expressed in BG, but not VA cells.
Furthermore, BGs required Shh signals from PCs to maintain their identities. When transgenic techniques were used to switch off either Shh production in PCs or Shh signaling in BGs in adult mouse brains, the BG cells adopted a transcription profile similar to that of VAs. If Shh signaling in VAs was given a boost on the other hand, these cells became more like BGs.
We could “almost interconvert one type of astrocyte into another based upon the level of Shh signaling,” said Murai. And it wasn’t just a handful of factors, he added: “We’re talking about hundreds of genes that are either being turned on or turned off in response to this pathway.”
The team also found evidence that astrocytes in other brain regions were influenced by Shh manipulations, and that these cells’ electrophysiologies were altered as a result.
“The key message is that astrocytes’ molecular fate is not hardwired,” said cell biologist Cagla Eroglu of Duke University in Durham, North Carolina, who did not participate in the study. The shapes of these cells appear to be less malleable, however. While Shh signaling influenced astrocyte expression profiles and electrical behaviors, the cells’ morphologies remained largely unchanged.
The finding that astrocyte identity is considerably more plastic than previously thought is “exciting and interesting,” added Cagla, “but it remains to be seen what the exact function of this will be in terms of an animal’s behavior or its ability to learn.”
W.T. Farmer et al., “Neurons diversify astrocytes in the adult brain through sonic hedgehog signaling,” Science, 351:849-54, 2016.
Glial cell properties dictated by neurons
Neurons in the brain coexist with astrocytes, a type of glial cell, which help support many functions of their neighboring nerve cells. Farmer et al. now show that the support goes both ways (see the Perspective by Stevens and Muthukumar). They explored the influence of neurons on two specialized types of astrocytes in the mouse cerebellar cortex. The neurons produced the morphogen known as Sonic Hedgehog. Hedgehog signaling adjusted distinctive gene expression within the two astrocyte cell types. Thus, mature neurons appear to promote and maintain specific properties of associated astrocytes.
Science, this issue p. 849; see also p. 813
Astrocytes are specialized and heterogeneous cells that contribute to central nervous system function and homeostasis. However, the mechanisms that create and maintain differences among astrocytes and allow them to fulfill particular physiological roles remain poorly defined. We reveal that neurons actively determine the features of astrocytes in the healthy adult brain and define a role for neuron-derived sonic hedgehog (Shh) in regulating the molecular and functional profile of astrocytes. Thus, the molecular and physiological program of astrocytes is not hardwired during development but, rather, depends on cues from neurons that drive and sustain their specialized properties.
Sonic hedgehog regulates discrete populations of astrocytes in the adult mouse forebrain.
J Neurosci. 2010 Oct 13;30(41):13597-608. http://dx.doi.org:/10.1523/JNEUROSCI.0830-10.2010
Astrocytes in Gli1-expressing regions of the forebrain show signs of mild reactive gliosis following postnatal interruption of Shh signaling
During development, Shh plays a critical role in specification of neuronal and oligodendrocyte precursors (Jessell, 2000). Shh has also been implicated in regulating proliferation of astrocytes in the developing optic nerve (Wallace and Raff, 1999; Dakubo et al., 2008).
Upregulation of GFAP and hypertrophy are hallmarks of reactive astrogliosis, which occurs in response to injury or disease (Eng and Ghirnikar, 1994). Reactive astrocytes have also been shown to undergo increased proliferation and synthesis of nestin and vimentin (Sofroniew, 2009). However BrdU labeling showed no change in proliferation between mutants and controls (data not shown), and staining for nestin and vimentin showed no change in expression of these intermediate filaments (data not shown). Taken together, these data show that the cellular response to interrupting Shh signaling includes key characteristics of reactive astrogliosis and indicates that Shh signaling plays an important role in mediating intracellular properties of specific astrocyte populations.
In this study, we show that high level Shh signaling in the adult CNS occurs in regionally distinct populations of mature, differentiated astrocytes. Our data demonstrate that neurons are a source of Shh, suggesting a novel signaling pathway involved in direct neuronastrocyte communication. Furthermore, we provide evidence that Shh signaling is required to maintain normal cellular functions in specific astrocyte populations. Taken together, our data are the first to demonstrate a critical role for Shh signaling in neuron-astrocyte communication in vivo, in the adult CNS.

http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2966838/bin/nihms-244753-f0004.jpg
{kind=link}
Astrocytes that express Gli1 are differentially distributed between specific forebrain regions
(A–C) Double labeling immunohistochemistry for βGal (red) and S100β (green) in the cortex (A), globus pallidus (B), and hypothalamus (C) of adult Gli1CreER/+;R26lacZ animals given tamoxifen 3 months earlier. Counterstained with DAPI (blue). Scale bar, 25μm. (D) Quantitative analysis of the proportion of S100β-positive astrocytes within the cortex, globus pallidus, and hypothalamus that express Gli1. (mean ± SEM, n≥300 cells/region/animal, from 3 animals, *p < 0.05, **p < 0.01, one-way, repeated measures ANOVA and Tukey’s post-test.)
- Farmer WT, Abrahamsson T, Chierzi S, Lui C, Zaelzer C, Jones EV, Ponroy Bally B, Chen GG, Théroux JF, Peng J, Bourque CW, Charron F, Ernst C, Sjöström PJ, and Murai KK: Neurons Diversify Astrocytes in the Adult Brain Through Sonic Hedgehog Signaling. Science (2016) 351(6275):849-854 DOI: 10.1126/science.aab3103 [abstract] [perspective by Stevens and Muthukumar]
- Lalanne T, Oyrer J, Mancino A, Gregor E, Chung A, Huynh L, Burwell S, Maheux J, Farrant M, and Sjöström PJ: Synapse-specific expression of calcium-permeable AMPA receptors in neocortical layer 5. The Journal of Physiology (2016) 594(4):837-861. (Editor’s Choice) [Abstract]
- Larsen RS & Sjöström: Synapse-type-specific plasticity in local circuits. Current Opinion in Neurobiology (2015) 35:127-135. [Abstract]
- Costa RP, Froemke RC, Sjöström PJ, & van Rossum MCW: Unified pre- and postsynaptic long-term plasticity enables reliable and flexible learning. eLife(2015) DOI:10.7554/eLife.09457. [Abstract]
- Ferreira TA, Blackman AV, Oyrer J, Jayabal S, Chung AJ, Watt AJ, Sjöström PJ, & van Meyel DJ: Neuronal morphometry directly from bitmap images.Nature Methods (2014) 11(10):982-984, DOI:10.1038/nmeth.3125. [Abstract]
- Blackman AV, Grabuschnig S, Legenstein R, & Sjöström PJ: A comparison of manual neuronal reconstruction from biocytin histology or 2-photon imaging: morphometry and computer modeling. Frontiers in Neuroanatomy (2014) 8:65, DOI: 10.3389/fnana.2014.00065. [Abstract]
- Blackman AV, Abrahamsson T, Costa RP, Lalanne T, & Sjöström PJ: Target-cell-specific short-term plasticity in local circuits. Frontiers in Synaptic Neuroscience (2013) 5:11, DOI: 10.3389/fnsyn.2013.00011. [Abstract]
- Costa RP, Sjöström PJ, & van Rossum MCW: Probabilistic inference of short-term synaptic plasticity in neocortical microcircuits. Frontiers in Computational Neuroscience (2013) 7:75, DOI:10.3389/fncom.2013.00075. [Abstract] [ModelDB code]
- Costa RP, Watt AJ, & Sjöström PJ: How to train a neuron. eLife (2013) 2:e00491, DOI: 10.7554/eLife.00491. Commentary on Pawlak et al, eLife 2013.
Neurogenesis in the embryonic and adult brain: same regulators, different roles
Neurogenesis persists in adult mammals in specific brain areas, known as neurogenic niches. Adult neurogenesis is highly dynamic and is modulated by multiple physiological stimuli and pathological states. There is a strong interest in understanding how this process is regulated, particularly since active neuronal production has been demonstrated in both the hippocampus and the subventricular zone (SVZ) of adult humans. The molecular mechanisms that control neurogenesis have been extensively studied during embryonic development. Therefore, we have a broad knowledge of the intrinsic factors and extracellular signaling pathways driving proliferation and differentiation of embryonic neural precursors. Many of these factors also play important roles during adult neurogenesis, but essential differences exist in the biological responses of neural precursors in the embryonic and adult contexts. Because adult neural stem cells (NSCs) are normally found in a quiescent state, regulatory pathways can affect adult neurogenesis in ways that have no clear counterpart during embryogenesis. BMP signaling, for instance, regulates NSC behavior both during embryonic and adult neurogenesis. However, this pathway maintains stem cell proliferation in the embryo, while it promotes quiescence to prevent stem cell exhaustion in the adult brain. In this review, we will compare and contrast the functions of transcription factors (TFs) and other regulatory molecules in the embryonic brain and in adult neurogenic regions of the adult brain in the mouse, with a special focus on the hippocampal niche and on the regulation of the balance between quiescence and activation of adult NSCs in this region.
Neural stem cells (NSCs) in the embryonic and early postnatal murine brain generate neurons and glia, including astrocytes and oligodendrocytes. The transition of proliferative and multipotent NSCs to fully differentiated neurons and glia is called neurogenesis and gliogenesis, respectively. Neurons are generated from early embryonic development until early postnatal stages, with only a few neurogenic zones remaining active in the adult (Götz and Huttner, 2005; Ming and Song, 2011;Paridaen and Huttner, 2014). In contrast, gliogenesis starts during late embryogenesis and continues in postnatal stages, with low but widespread production of both astrocytes and oligodendrocytes also occurring throughout the adult brain (Rowitch and Kriegstein, 2010; Gallo and Deneen, 2014; Guérout et al., 2014). The main neurogenic regions in the adult murine brain are the subependymal zone of the lateral ventricles, also called ventricular-subventricular Zone (V-SVZ) and the subgranular zone (SGZ) of the dentate gyrus (DG) in the hippocampus (Altman and Das, 1965; Doetsch et al., 1999; Ming and Song, 2011;Fuentealba et al., 2012). Both of these neurogenic regions have been shown to also be active in the adult human brain, with the V-SVZ thought to contribute new neurons to the striatum (whereas it produces neurons migrating to the olfactory bulb in mice) and the SGZ contributing neurons to the DG (Eriksson et al., 1998; Spalding et al., 2013; Ernst et al., 2014). The addition of new neurons to the complex circuitry of the adult brain is the focus of intensive research, which is uncovering crucial functions for the newly generated neurons in memory and behavior (Deng et al., 2010). In particular, the integration of adult-born granule cells to the hippocampus circuitry confers an extra degree of plasticity that is crucial for the acquisition of certain types of contextual memory (Jessberger et al., 2009; Sahay et al., 2011). Although adult neurogenesis is an ancient trait, with widespread neurogenesis occurring, for instance, in 16 different adult brain areas of zebrafish, the appearance of the DG as a structural and functional unit seems exclusive to mammals (Treves et al., 2008; Grandel and Brand, 2013). This fact, amongst others, has prompted the idea that hippocampal neurogenesis might be a newly evolved trait in some species, including humans, aimed to enhance adaptation to a continuously changing environment (Kempermann, 2012).
Significant advances have been made in our understanding of the regulation of mouse adult hippocampal neurogenesis in the last few years. Thus, our focus for the rest of the review will be on the mouse model of neurogenesis. The coordinated action of multiple signals acting on embryonic NSCs gives rise to the vast diversity of neuronal and glial populations that populate the mature brain. Embryonic neurogenesis is, thus, tightly linked to cell fate specification. In adult neurogenic regions, however, stem cells are tightly restricted to the generation of one (granule neurons of the DG) or a few types of neurons (granule neurons and periglomerular neurons in the V-SVZ) (Zhao et al., 2008; Ming and Song, 2011). Therefore signals and factors that specify subtype identities during development can control more subtle aspects of adult stem cell behavior.
In recent years, it has become evident that, at the single cell level, stem cells in the embryonic and the adult brain are not as versatile as previously thought. Instead of their classically attributed multipotency, they appear to be already committed to the generation of specific types of neural cells (Taverna et al., 2014). The causes and functions of the emerging heterogeneity of adult NSCs are among the most exciting questions remaining to be addressed in the field (DeCarolis et al., 2013; Encinas et al., 2013; Giachino et al., 2014b). In the case of the murine V-SVZ, different populations of adult NSCs, also called type-B cells, co-exist and give rise to distinct types of periglomerular cells and granule cells in the olfactory bulb. Different adult NSCs are characterized by the differential expression of specific transcription factors (TFs), including Nkx2.1, Pax6, Gsx2 and Nkx6.2, which also pattern the different domains of the embryonic telencephalon (Merkle et al., 2007; Brill et al., 2008;López-Juárez et al., 2013; Merkle et al., 2014). The distinct adult NSC populations are located in different regions along the V-SVZ and their distinct properties are acquired during development (Obernier et al., 2014). Despite the spatial separation of these stem cell populations, all their progeny follow the same long migratory path, the rostral migratory stream (RMS), towards their final destination in the olfactory bulb. In the hippocampus, adult NSCs, also called type-I cells or radial glial-like cells, generate exclusively granule neurons in the DG. The migration of granule neurons is very limited, as they settle, differentiate and integrate into the hippocampal circuitry in the granule cell layer (GCL) located just above the NSC from which they originated in the SGZ. While they appear uniform, adult NSCs in the DG respond to diverse and complex signals, raising the possibility that they are functionally heterogeneous.
Despite their many differences, adult NSCs in the two adult neurogenic niches share several key characteristics. Neural stem cells in both V-SVZ and SGZ, like radial glial stem cells in the embryo, express the molecules GFAP, Nestin and Sox2 and they directly contact blood vessels. Both NSC populations share a restricted potential, as just discussed, with each generating a unique neuronal subtype and one type of glia: in the V-SVZ they generate neurons and oligodendrocytes, while in the SGZ they generate neurons and astrocytes. Perhaps the two characteristics that distinguish adult NSCs most clearly from their embryonic counterparts are the acquisition of quiescence and their situation in a complex and stable cellular niche. While one of the main features of embryonic NSCs is their high proliferative rate, the opposite is true for adult NSCs, which remain for long periods out of the cell cycle, in G0. This is a characteristic that adult NSCs share with many stem cells in other mature tissues and one that is crucial to maintain tissue homeostasis and avoid stem cell exhaustion (Orford and Scadden, 2008;Simons and Clevers, 2011). The existence of adult neurogenic niches (complex cellular microenvironments surrounding adult NSCs) is also a characteristic shared with other tissues (Fuchs et al., 2004; Kuang et al., 2008; Mirzadeh et al., 2008; Ming and Song, 2011; Fuentealba et al., 2012; Goldstein and Horsley, 2012). The niche is comprised of diverse cell types and structures, such as astrocytes, neurons, axon projections and blood vessels, and one of its main functions is to create an appropriate environment that keeps the majority of stem cells quiescent and undifferentiated (Morrison and Spradling, 2008). The niche also provides a great variety of signals that modulate the behavior of adult stem cells and adjust the production of new cells to the needs of the tissue (Fuchs et al., 2004; Blank et al., 2008; Faigle and Song, 2013).
Embryonic and Adult Origin of Granule Cells
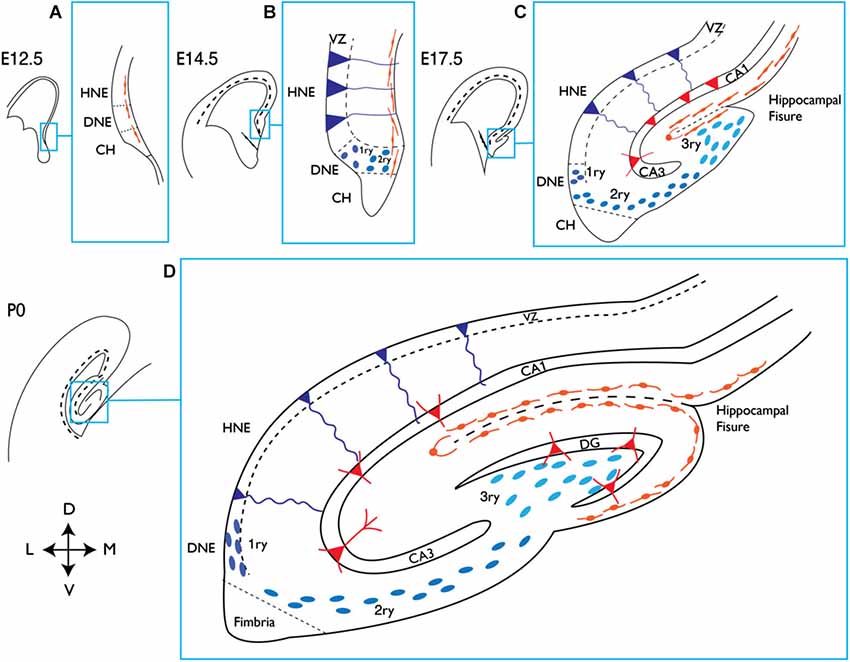
From a developmental point of view, the generation of the DG is unique. While the V-SVZ is seen as a continuation of the embryonic ventricular zone (VZ) of the telencephalon, the formation of the DG involves the generation of a dedicated progenitor cell source away from the VZ and in close proximity to the pial surface. This additional proliferative zone remains active during postnatal stages and eventually becomes the SGZ, which is the site of adult hippocampal neurogenesis (Figure 1;Bayer, 1980a,b; Altman and Bayer, 1990; Pleasure et al., 2000; Khalaf-Nazzal and Francis, 2013; Sugiyama et al., 2013).

http://www.frontiersin.org/files/Articles/118806/fncel-08-00396-r4/image_m/fncel-08-00396-g001.jpg
{kind=link}
Development of the mouse hippocampus. Schematic representation of the dorsal telencephalon at different embryonic (E) stages and at birth (P0). The indicated area in each picture corresponds to the hippocampal region and is magnified on its right handside (blue squares). (A) At E12.5 the presumptive DNE is located between the HNE and the CH, which produces Cajal-Retzius cells (orange), shown lining the pial side of the cortex. (B) At E14.5 dentate precursors of the primary matrix (dark blue circles) are located in the VZ, and precursor cells start to migrate towards the pial side of the cortex forming the secondary matrix. In the VZ of the HNE, radial glial precursors (depicted in dark blue and triangular body shape) will give rise to hippocampal neurons. (C) At E17.5 the hippocampal fissure is formed and dentate precursor cells migrate to and accumulate there, forming the tertiary matrix (light blue). Cajal-Retzius cells are also present and follow the hippocampal fissure. At this stage the glial scaffold (not shown) extends from the CH to the hippocampal fissure and pial surface, directing the migration of dentate precursor cells. From the HNE, hippocampal neurons (red triangles) are born and migrate along radial glial cells towards their location in the hippocampal fields (CA1 and CA3 are shown). (D) At birth the blades of the DG start to form. Granule neurons in the DG (red triangles) appear first in the upper blade, below the hippocampal fissure. The continuous migration of Cajal-Retzius cells reaches the pial side and promotes the formation of the lower blade of the DG. Precursor cells in the primary and secondary matrix will soon disappear, but cells in the tertiary matrix continue actively dividing and producing granule neurons through postnatal DG development. HNE, hippocampal neuroepithelium; DNE, dentate neuroepithelium; CH, cortical hem; VZ, ventricular zone; 1ry, primary matrix; 2ry, secondary matrix; 3ry, tertiary matrix; DG, dentate gyrus; D, dorsal; M, medial; V, ventral; L, lateral.
Regulation of Adult Neurogenesis
The late maturation of the hippocampus, which spans late embryonic and early postnatal stages, means that the process of DG formation and the appearance of NSCs with adult characteristics are overlapping processes. It can therefore be difficult to distinguish between developmental and adult cues regulating hippocampal neurogenesis. However, several physiological and pathological situations, such as physical exercise, task learning, an enriched environment and seizures, have been shown to stimulate neurogenesis specifically in the adult DG (Rolando and Taylor, 2014). Although no direct link has been clearly established between those external stimuli and signaling pathways, numerous extracellular signaling molecules, including Bone Morphogenetic Proteins (BMPs), Notch, GABA, WNT, insulin growth factors (IGFs) and SHH, have been shown to regulate the rate of neurogenesis in the adult DG (Ming and Song, 2011; Faigle and Song, 2013). However, due to limitations of in vivo studies, little is known about the mechanisms by which these signals exert their effects. In the adult DG, NSCs generate granule cells via a well characterized cell lineage that includes a succession of transit amplifying or intermediate progenitor cells (IPCs), characterized by rapid divisions and the expression of a series of neurogenic TFs (Figure 2; Hsieh, 2012). Extrinsic stimuli can affect the proliferation and survival of NSCs but also of IPCs (typeIIa and typeIIb) or differentiating neuroblasts (typeIII) further along the lineage (Figure 2). The selective death of IPCs, for instance, is a major mechanism of regulation of neurogenesis in the DG, with as many as two thirds of these cells being actively eliminated by microglia (Sierra et al., 2010, 2014). Therefore, in order to understand the effects of signaling pathways and intrinsic factors on neurogenesis, it is crucial to determine the stages in the adult neurogenic lineage at which they act, and the cellular processes they regulate. In fact, one of the main difficulties faced by the adult neurogenesis field concerns the scarcity of markers for adult NSCs, which are often shared by other cell types (for instance, GFAP marks subpopulations of astrocytes and Nestin is expressed by early intermediate progenitors). This problem is only more evident in the case of distinguishing quiescent from activated adult NSCs, in which case there is an absolute lack of specific markers apart from the use of cell cycle genes. This issue has been partly addressed in a recent report in which an unbiased approach was used to identify genes differentially expressed by activated and quiescent adult NSCs isolated from the V-SVZ (Codega et al., 2014). This work demonstrates that the quiescent state is a much more complex state than simply the lack of proliferation markers, as the list of differentially expressed genes is enriched in genes related to very diverse cellular processes, such as lipids metabolism, signaling or adhesion. This quiescence signature is shared by adult quiescent stem cells from other organs, such as the blood, muscle or intestine (Cheung and Rando, 2013; Codega et al., 2014). It is thus likely that many of the general characteristics of quiescent stem cells will be shared between DG and SVZ, although no studies on the expression profile of adult DG NSCs have been performed to date.
Ageing of the brain is marked by a major decrease in the number of new neurons generated in the DG. This decrease has been attributed both to a reduction of the NSC pool and to an increased state of quiescence of the remaining stem cells (Lugert et al., 2010; Encinas et al., 2011; Jaskelioff et al., 2011; Seib et al., 2013). The possibility to increase neurogenesis in ageing mice by activating the quiescent stem cell pool is currently the focus of intensive research. In this regard, it was recently shown that systemic factors from young animals can re-activate neurogenesis in aged mice (Katsimpardi et al., 2014). However, disruption of quiescence signals can lead to a short-lived increase in neurogenesis, followed by a sharp decrease caused by a loss of quiescent NSCs (Ehm et al., 2010; Mira et al., 2010; Song et al., 2012). Assessing precisely how factors and signals affect stem cell behavior will be vital to understand their long-term effects on adult neurogenesis. Lineage tracing and particularly clonal analysis of NSCs in the DG have begun to provide evidence of the great diversity of responses of adult NSCs to stimuli, which can affect both their proliferation and differentiation potentials (Bonaguidi et al., 2011; Dranovsky et al., 2011; Song et al., 2012).
Notch Signaling
The functions of Notch signaling during embryonic brain development have been extensively reviewed elsewhere (Kageyama et al., 2008; Imayoshi and Kageyama, 2011). During development of the hippocampus, Notch does not seem to be involved in neural precursor specification or differentiation, but rather in broader decisions, including the regulation of neural lineage commitment, the tempo of neuronal and glial generation and the maintenance of stem cells. Notch receptors and ligands are broadly expressed during all stages of development of the hippocampus (Pleasure et al., 2000). Loss of the essential Notch signaling component RBPJk in the developing brain results in proliferation defects and premature differentiation of embryonic NSCs (Imayoshi et al., 2010). Similarly, loss of RBPJk or of the Notch ligand Jagged1 during hippocampal development leads to defects in proliferation and stem cell maintenance, although the formation of the DG is not prevented (Breunig et al., 2007; Lavado and Oliver, 2014). Therefore, the main function of the Notch pathway in embryonic NSCs is to maintain their proliferative and undifferentiated state.
Other Signaling Pathways: SHH, IGF and Neurotransmitters
We will briefly discuss here the roles of other key signaling pathways for which specific roles in hippocampal development or in adult neurogenesis have not been reported.
Sonic Hedgehog signaling has crucial roles in early patterning and cell fate specification in the embryonic brain. Recently, NSCs in the adult DG have been shown to originate from SHH-responsive progenitors in the ventral hippocampus (Li et al., 2013). Sonic Hedgehog signaling has been implicated in the proliferation and maintenance of both DG and V-SVZ adult NSCs (Machold et al., 2003; Álvarez-Buylla and Ihrie, 2014). Although the sources of SHH that regulate V-SVZ and SGZ neurogenesis have not been clearly identified yet, tracing the activity of SHH by the expression of the SHH-inducible geneGli1 in Gli1nLacZ mice has shown that NSCs in both adult neurogenic regions as well as a fraction of mature astrocytes express the beta galactosidase reporter protein and therefore receive SHH signals (Ahn and Joyner, 2005; Garcia et al., 2010;Ihrie et al., 2011; Petrova et al., 2013). Removal of SHH signaling from V-SVZ stem cells by deletion of the receptor Smoothened has revealed that SHH is necessary for the proliferation and long term maintenance of the stem cells, as well as the subtype specification of the neurons they generate (Palma et al., 2005; Balordi and Fishell, 2007; Kim et al., 2007; Ihrie et al., 2011; Petrova et al., 2013; Merkle et al., 2014). In adult DG stem cells, conditional disruption of primary cilia, which are required for SHH signaling, decreases the production of IPCs, supporting a role for SHH in NSC divisions in the DG as well (Breunig et al., 2008; Amador-Arjona et al., 2011). However, a more direct investigation of the role of SHH in adult DG neurogenesis has not yet been performed.
The Transition from Postnatal to Adult Neurogenesis: NFIX, Tlx, CcnD2 and Ascl1
Granule neurons in the adult DG are exclusively generated by NSCs located in the SGZ. During embryonic and postnatal development, in contrast, neurons are generated by a heterogeneous population of precursor cells in the dentate matrices (Figures 1, 2). The exact time at which the switch from embryonic to adult modes of neurogenesis occurs in the DG is still not well defined. Several independent pieces of evidence suggest that this happens around the second week of life in mice. At postnatal day 14 (P14), the blades of the DG are already formed and the source of new neurons in the DG becomes restricted to the tertiary matrix, which gradually becomes the SGZ (Pleasure et al., 2000; Sugiyama et al., 2013). At the same time, the first presumptive GFAP- and Nestin-positive NSCs adopt their characteristic location, with the nucleus residing in the SGZ and the basal process extending through the GCL (Li and Pleasure, 2005; Martynoga et al., 2013).
Conclusions
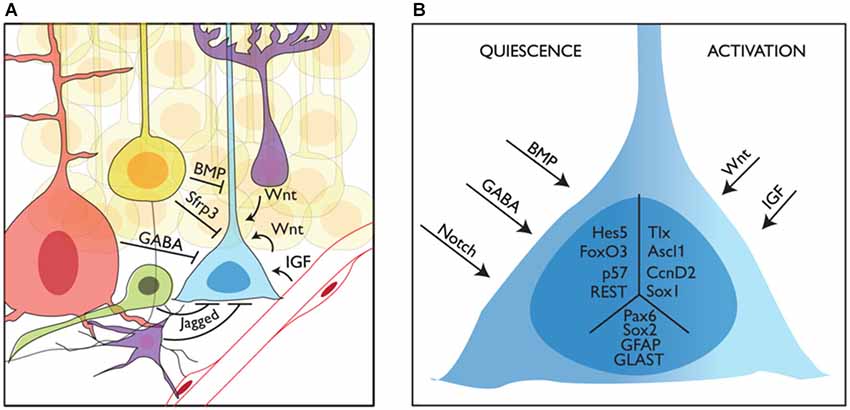
Adult neurogenic niches can be conceptualized as remnants of embryonic signaling centers (i.e., the septum/antihem giving rise to the V-SVZ and the CH generating the SGZ): they are the source of instructive signals that determine the fate of neighboring stem cells. However, in contrast with stem cells in the developing brain that must cope with a continuously changing environment, adult stem cells are surrounded by a relatively stable niche. The V-SVZ and the SGZ niches share many common features. However, while the cellular and molecular composition of the V-SVZ niche has been relatively well investigated, we lack a similar level of understanding of the SGZ niche. Further studies of the signals and cellular interactions that control NSC behavior in the DG will be required before we can appreciate the similarities and divergences in the regulation and function of stem cells in the two adult neurogenic niches (Figure 3).

http://www.frontiersin.org/files/Articles/118806/fncel-08-00396-r4/image_m/fncel-08-00396-g003.jpg
{kind=link}
Niche regulation of mouse adult stem cells in the dentate gyrus. (A) Representation of a neural stem cell (blue) in the adult subgranular zone of the dentate gyrus and some of its interactions with the niche. Granule neurons (yellow), interneurons (red), intermediate precursors (green) and astrocytes (purple) are shown providing quiescence cues, while blood vessels and astrocytes are shown providing activation cues. (B) How quiescence and activation signals are interpreted by adult stem cells is still not known. Here we show several intracellular factors that have been linked to the quiescent (left, Hes5, p. 57, FoxO3 and REST) or active (right, Tlx, Ascl1 and CcnD2) state of stem cells in the adult DG. We also show other factors expressed in NSCs with no clear function in the switch from quiescence to activation (Sox2, Pax6, GFAP and GLAST) in the central part of the schematized cell.